ALDH18A1 , ARCL3A, GSAS, P5CS, PYCS, семейство альдегиддегидрогеназы 18, член A1, ADCL3, SPG9A, SPG9B, Альдегиддегидрогеназа 18 Член семейства A1, SPG9
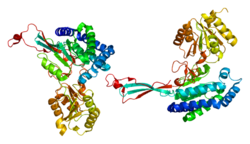
P5CS состоит из двух доменов: гамма-глютамилкиназы и гамма-глутамилфосфатредуктазы, каждая из которых используется для завершения двух этапов для создания орнитина и пролина. В домене гамма-глютамилкиназы используется цепочка 367- остатков , которая складывается в N-концевую аминокислотную киназную домен, ответственный за катализ и ингибирование пролина, а также С-концевой PUA домен связывания . Этот фермент также является тетрамером, образованным двумя димерами , а мономеры и димеры собираются таким образом, что позволяет аминокислотные активные карманы в тетрамере. альтернативно ориентировать [ 8 ] Короткая версия (p5cs.short) и длинная версия (p5cs.long) PC5S представляют собой две изоформы этого фермента, которые отличаются от добавления двух аминокислот в длинной форме и дополнительной вставкой 6 п.н. после BP+711 Полем Эта небольшая разница создает драматическую разницу в том, как на них влияет ингибирование орнитина . [ 9 ] Ген ALDH18A1 охватывает 15 т.п.н. , нанесен на карту на 10Q24.3 и имеет количество экзонов 18. [ 6 ] [ 10 ]
P5Cs катализирует фосфорилирование -и восстановление -конверсию глутамата с дельта-1-пирролин-5-карбоксилатом (P5C). Это происходит посредством процесса, в котором глутамат превращается в гамма-глютамилфосфат в домене гамма-глутамилкиназы, а затем гамма-глютамилфосфат превращается в гамма-глютамический семяльдегид в домене гамма-глутамил фосфат. Гамма-глутамический полуалдегид находится в таутомерном равновесии с p5c и является обязательным промежуточным соединением в взаимосвязи пролина, орнитина и глутамата. [ 10 ] Две изоформы (PC5S.Short и PC5S.Long) также участвуют и в различных действиях. Короткая версия обладает высокой активностью в кишечнике и является основным участником биосинтеза аргинина . Длинная версия PC5S экспрессируется в различных тканях и является важной для его способности синтезировать пролин из глутамата. Кроме того, короткая версия ингибируется орнитином, тогда как длинная версия нечувствительна к аминокислоте. [ 11 ]
Отдельные домены эукариотических P5C находятся в прокариотах и одноклеточных эукариотах, присутствующих в двух отдельных ферментах, ProA (гамма-глютамилфосфатредуктаза) [ 12 ] и проб (гамма-глютамилкиназа). [ 13 ] Последовательность и структурные данные обоих наследственных ферментов позволяют предположить, что ProA и Prob одного Genes происходят из дупликации гена и последующей субфункционализации . [ 14 ] Слиянию ProA и Prob , вероятно, предшествовало делеции домена PUA в вероятности , снижая чувствительность к ингибированию обратной связи пролина и обеспечивая наблюдаемое направление промежуточного звена гамма-глутамилфосфата. [ 15 ]
Орнитин и/или аргинин являются ключевыми промежуточными продуктами для синтеза мочевины , креатина , оксида азота , полиаминов и белка ; в то время как пролин является основным компонентом соединительной ткани
белки, коллаген и эластин . Поскольку все три из этих аминокислот являются частью очень значительных процессов, наличие P5C становится важным регулятором, который гарантирует, что ни один из этих трех не станет дефицитом. [ 16 ] Следовательно, отсутствие p5C, из -за мутаций в гене ALDH18A1 , часто приводит к нейродегенерации, слабости сустава , гиперладности кожи , двусторонней понука -капсульной катаракте и множеству других осложнений, связанных с нарушением синтеза пролина и орнитина. [ 11 ]
У всех видов растений активность растений P5CS также связана с путем биосинтеза пролина, как во время оптимальных, так и во время абиотических стрессовых условий. Подобно млекопитающим, растения обладают двумя изоферментами P5CS, кодируемыми паралогическими генами, P5CS1 и P5CS2 . Различные виды растений развивались для использования различных паралогов P5CS в зависимости от окружающей среды. [ 14 ] Таким образом, P5CS2 действует как домашнее хозяйство в Arabidopsis, эта функция обеспечивается P5CS1 в рисе; В отличие от Arabidopsis P5CS1 обеспечивает лучшую производительность при абиотическом стрессе, в то время как P5CS2 отвечает за улучшение реакции абиотического стресса в рисе. Arabidopsis p5cs2 локализован в цитоплазме и хлоропластах. [ 17 ] [ 18 ] P5CS-зависимый синтез p5C является ограничивающей скоростью стадией биосинтеза пролина и ингибируется связыванием конечного продукта, L-пролин, предположительно предотвращая связывание глутамата с сайтом входа в гамма-глютамилкиназной субъединице. [ 8 ]
^ «Мышь Pubmed ссылка:» . Национальный центр информации о биотехнологии, Национальная медицина США .
^ Лю Г., Маунури С., Камоун П., Арал Б (октябрь 1996 г.). «Назначение человеческого гена, кодирующего дельта-пирролин-5-карбоксилат-синтетазу (P5CS) до 10Q24.3 с помощью гибридизации in situ». Геномика . 37 (1): 145–6. doi : 10.1006/geno.1996.0535 . PMID 8921385 .
^ Martinelli D, Häberle J, Rubio V, Giunta C, Hausser I, Carrozzo R, et al. (Сентябрь 2012 г.). «Понимание дефицита пирролина-5-карбоксилат-синтетазы: клинические, молекулярные, функциональные и экспрессирующие исследования, структурный анализ и новая терапия аргинином». Журнал наследственного метаболического заболевания . 35 (5): 761–76. doi : 10.1007/s10545-011-9411-8 . PMID 22170564 . S2CID 6881255 .
^ Хур Дж, Юнг К.Х., Ли Ч., Г. (сентябрь 2004 г.). «Индуцируемый стрессом ген OSP5CS2 необходим для соли и устойчивости к холоду в рисе». Наука растений . 167 (3): 417–426. doi : 10.1016/j.plantsci.2004.04.009 .
Maruyama K, Sugano S (январь 1994 г.). «Олиго-капитализация: простой метод замены структуры крышки эукариотических мРНК олигорибонуклеотидами». Ген . 138 (1–2): 171–4. doi : 10.1016/0378-1119 (94) 90802-8 . PMID 8125298 .
Aral B, Schlenzig JS, Liu G, Kamoun P (март 1996 г.). «Клонирование базы данных Delta 1-пирролин-5-карболин-карболин-карбоксилат-синтетазы (P5CS): бифункциональный фермент, катализирующий первые 2 стадии биосинтеза пролина». Comptes rendus de l'Académie des Sciences, Série III . 319 (3): 171–8. PMID 8761662 .
Suzuki Y, Yoshitomo-Nakagawa K, Maruyama K, Suyama A, Sugano S (октябрь 1997 г.). «Строительство и характеристика обогащенной полной длиной и библиотекой кДНК с 5'-эндом». Ген . 200 (1–2): 149–56. doi : 10.1016/s0378-1119 (97) 00411-3 . PMID 9373149 .
Arc.Ask3.Ru Номер скриншота №: a32b1a611380d2c6f9d89b3c8f4f5873__1695818640 URL1:https://arc.ask3.ru/arc/aa/a3/73/a32b1a611380d2c6f9d89b3c8f4f5873.html Заголовок, (Title) документа по адресу, URL1: Aldehyde dehydrogenase 18 family, member A1 - Wikipedia
Данный printscreen веб страницы (снимок веб страницы, скриншот веб страницы), визуально-программная копия документа расположенного по адресу URL1 и сохраненная в файл, имеет: квалифицированную, усовершенствованную (подтверждены: метки времени, валидность сертификата), открепленную ЭЦП (приложена к данному файлу), что может быть использовано для подтверждения содержания и факта существования документа в этот момент времени. Права на данный скриншот принадлежат администрации Ask3.ru, использование в качестве доказательства только с письменного разрешения правообладателя скриншота. Администрация Ask3.ru не несет ответственности за информацию размещенную на данном скриншоте. Права на прочие зарегистрированные элементы любого права, изображенные на снимках принадлежат их владельцам. Качество перевода предоставляется как есть. Любые претензии, иски не могут быть предъявлены. Если вы не согласны с любым пунктом перечисленным выше, вы не можете использовать данный сайт и информация размещенную на нем (сайте/странице), немедленно покиньте данный сайт. В случае нарушения любого пункта перечисленного выше, штраф 55! (Пятьдесят пять факториал, Денежную единицу (имеющую самостоятельную стоимость) можете выбрать самостоятельно, выплаичвается товарами в течение 7 дней с момента нарушения.)