Гаплогруппа u
| Гаплогруппа u | |
|---|---|
| Возможное время происхождения | 46 500 ± 3300 лет назад [ 1 ] |
| Возможное место происхождения | Западная Азия, [ 2 ] |
| Предок | Ведущий |
| Потомки | U1, U5, U6, U2'3'4'7'8'9 |
| Определение мутаций | 11467, 12308, 12372 [ 3 ] |
Гаплогруппа U представляет собой гаплогруппу митохондриальной ДНК человека (мтДНК). Клада возникла из гаплогруппы R , вероятно, во время ранней верхней палеолита . Различные подклады (помеченные U1 - U9, расходящиеся в течение верхнего палеолита), находятся широко распространенными по всей северной и восточной Европе , Центральной , Западной и Южной Азии , а также в Северной Африке , Африке и на Канарских островах. Полем
Происхождение
[ редактировать ]Гаплогруппа U выходит из гаплогруппы R Mtdna ветви филогенетического дерева. Определяющие мутации (A11467G, A12308G, G12372A), по оценкам, составляют между 43 000 и 50 000 лет назад, в раннем верхнем палеолите (около 46 530 ± 3290 лет до настоящего, с 95% доверительным интервалом на Behar et al., 2012). Полем
Древняя ДНК , классифицированная как принадлежащая к митохондриальной гаплогруппе, была извлечена из скелетных останков человека, обнаруженных в западной Сибири, которые были датированы c. 45 000 лет назад. [ 4 ] Митогеном (33-кратный охват) индивидуума Peştera muierii 1 (PM1) из Румынии (35 Ky Cal BP) был идентифицирован как базальная гаплогруппа U6*, ранее не обнаруженная ни у древних или современных людей. [ 5 ]
Гаплогруппа U была обнаружена среди иберомавровских образцов, датируемых эпипалеолитом в доисторических местах Taforalt и Afalou . [ 6 ] Среди лиц тафоральта около 13%наблюдаемых гаплотипов принадлежали различным подкладам U, включая U4A2B (1/24; 4%), U4C1 (1/24; 4%) и U6D3 (1/24; 4%). Еще 41% анализируемых гаплотипов могут быть назначены либо гаплогруппе, либо гаплогруппе h . Среди индивидуумов Afalou 44% проанализированных гаплотипов могут быть назначены либо гаплогруппе U, либо гаплогруппе H (3/9; 33%). [ 7 ]
Гаплогруппа U также наблюдалась среди древних египетских мумий, раскопаемых в археологическом участке Abusir el-Meleq в Среднем Египте, датированном 1-м тысячелетию до н.э., 13 из 90 мумий, несущих гаплогруппу U (U носители все поздний период) и различные подклады этого, u, u1, u3, u5, u6, u7 и u8. [ 8 ] и в отдельном исследовании ДНК, извлеченная из зуба, мумифицированная голова гораздо более старшей мамы, около 4000 назад лет в современной популяции носителей U5) из статьи 2018 года Odile Loreille et al. [ 9 ]
Кроме того, гаплогруппа U наблюдалась в древних окаменелостях ганчика , раскопаемых в Гран-Канарии и Тенерифе на Канарских островах , которые были переданы радиоуглеродом между 7-м и 11-м веками. Все люди, несущие по кладке, были внушаются на сайте Tenerife, и эти образцы принадлежат к подкладам U6B1A (4/7; 57%) и U6B (1/7; 14%). [ 10 ]
Распределение
[ редактировать ]Гаплогруппа U находится в 15% индийской касты и 8% индийских племенных популяций. [ 11 ] Гаплогруппа U обнаруживается примерно у 11% местных европейцев и считается самой старой материнской гаплогруппой, обнаруженной в этом регионе. [ 11 ] [ 12 ] [ 13 ] В исследовании 2013 года все, кроме одной из древних современных человеческих последовательностей из Европы, принадлежали к материнской гаплогруппе U, что подтверждает предыдущие выводы о том, что гаплогруппа U была доминирующим типом митохондриальной ДНК (мтДНК) в Европе до распространения сельского хозяйства в Европу Ближний Восток. [ 14 ]
Гаплогруппа U имеет различные подклады, пронумерованные U1 до U9. Гаплогруппа K является подкладом U8. [ 15 ] Старость привела к широкому распределению подгрупп потомков по западной Евразии, Северной Африке и Южной Азии. Некоторые подклады гаплогруппы U имеют более специфический географический диапазон.
Подклады
[ редактировать ]Подклады помечены U1 - U9; Гаплогруппа K является подкладом U8.
Van Oven and Kayser (2009) предложили подклады «U2'3'4'7'8» и «U4'9». [ 3 ] Behar et al. (2012) внесли изменения в это, группировав «U4'9» как подчиненное «U2'3'4'7'8» для нового промежуточного подклада «U2'3'4'7'8'9».
Гаплогруппа u
[ редактировать ]Базальный U был найден в 26 000 -летних останках древнего Северного Евразийца , мальчика из Малты (MA1).
Гаплогруппа U1
[ редактировать ]Подклады U1: U1A (с глубокими субкладами U1A1, U1A1A, U1A1A1, U1A1B) [ 16 ] и U1B. [ 16 ]
По оценкам, гаплогруппа U1 произошла между 26 000 и 37 000 лет назад. Он встречается на очень низкой частоте по всей Европе. Это чаще наблюдается в Восточной Европе, Анатолии и на Ближнем Востоке. Это также встречается на низких частотах в Индии. U1 встречается в регионе Сванетии в Грузии в 4,2%. Подклад U1A встречается из Индии в Европу, но крайне редко среди северных и атлантических полос Европы, включая Британские острова и Скандинавия. В Индии U1A был найден в регионе Керала . U1B имеет аналогичный спред, но более реже, чем U1A. Разнообразие подкласса U1B1 с мутациями G14070A! и A3426G встречается в ашкенази -евреях . [ 17 ] Подклады U1A и U1B появляются в одинаковой частоте в Восточной Европе. [ 18 ]
Редкая клада U1 также обнаружена среди алжирцев в Оране -1,08%) и племени Регубата Сахрави (0,83 % (0,93%). [ 19 ]
Подклад U1A1A наблюдался в древнем отдельном отделении, раскопанном на кладбище Келлис 2 в Оазисе Дакле , расположенном в юго -западной пустыне Египта. 21 из захоронения Келлиса были радиоуглеродными до 80-445 г. н.э., сроки в римско-христианском периоде. [ 20 ] Гаплогруппа U1 также была обнаружена среди образцов на материковом кладбище в Кулубнарти , Судан , которые датируются ранним христианским периодом (550-800 гг. До н.э.). [ 21 ]
Анализ ДНК раскопанных останков, которые теперь расположены в руинах Церкви Святого Августина в Гоа , Индия также выявила уникальный подклад мтДНК U1B. Эта подсолингом отсутствует в Индии, но присутствует в Грузии и окружающих регионах. [ 22 ] Поскольку генетический анализ подтверждает археологические и литературные доказательства, считается, что раскопанные останки принадлежат кевану мученику , королеве Грузии. [ 22 ]
Гаплогруппа U5
[ редактировать ]Возраст U5 оценивается в 25 000 и 35 000 лет, [ 23 ] Примерно соответствует гравите . Приблизительно 11% европейцев (10% европейских американцев) имеют некоторый вариант гаплогруппы U5. Гаплогруппа, скорее всего, возникла в Европе. [ 24 ] [ 25 ]
U5 был преобладающей мтДНК мезолитических западных охотников (WHG).
U5 был обнаружен в человеческих останках, встречающихся из мезолита в Англии, Германии, Литве, Польше, Португалии, России, [ 26 ] Швеция, [ 27 ] Франция [ 28 ] и Испания. [ 29 ] Неолитические скелеты (~ 7000 лет), которые были раскопаны из пещеры Avellaner в Каталонии , Северо -Восточная Испания включал в себя образец с гаплогруппой U5. [ 30 ]
Гаплогруппа U5 и его подклады U5A и U5B сегодня формируют самые высокие концентрации населения на дальнем севере, среди сами , финны и эстонцы . Тем не менее, он широко распространен на более низких уровнях по всей Европе. Это распределение и возраст гаплогруппы указывают на то, что люди, принадлежащие к этой кладе, были частью первоначального расширения, отслеживающего отступление ледяных щитов из Европы около 10 000 лет назад.
Современные баски и кантабрийцы имеют почти исключительно линии U5B (U5B1F, U5B1C1, U5B2). [ 31 ] [ 32 ]
Кроме того, гаплогруппа U5 встречается на небольших частотах и при гораздо более низком разнообразии на Ближнем Востоке и в некоторых частях Северной Африки (районы с значительными концентрациями U6), что указывает на обратную миграцию людей из Европы на юг. [ 33 ]
Митохондриальная гаплогруппа U5A также была связана с ВИЧ -инфицированными людьми, демонстрирующими ускоренное прогрессирование в СПИДе и смерти. [ 34 ]
U5 был основной гаплогруппой мезолитических европейских охотников. U-гаплогруппы присутствовали на 83% у европейских охотников, прежде чем приток ближневосточного фермера и индоевропейского происхождения Steppe Endo-European снизил свою частоту до менее чем 21%. [ 20 ]
- U5
- U5A возник около 17 000 и 27 000 лет назад
- U5A1 возник между 14 000 и 20 000 лет назад. Обнаружено у этрусского человека (700-600 г. до н.э.) из Южной Этрурии , Италия . [ 35 ]
- U5A1A возник между 8000 и 16 000 лет назад
- U5A1A1 появился от 3000 до 11 000 лет назад
- U5A1A1A возник менее 6000 лет назад [ 36 ]
- U5A1A1B возникла около 600 и 6000 лет назад
- U5A1A1C
- U5A1A1D возникла менее 4300 лет назад
- U5A1A2 появился между 7000 и 14 000 лет назад
- U5A1A2A возник менее 5400 лет назад
- U5A1A2A1 появился менее 3400 лет назад
- U5A1A2A возник менее 5400 лет назад
- U5A1A1 появился от 3000 до 11 000 лет назад
- U5A1B появился между 6000 и 11 000 лет назад
- U5A1B1 появился между 5000 и 9 000 лет назад
- U5A1B1A возник между 2500 и 7500 годами
- U5A1B1A1 Менее 4000 лет назад
- U5A1B1B возникла менее 8000 лет назад
- U5A1B1C появился от 3000 до 7000 лет назад
- U5A1B1C1 возник менее 5000 лет назад
- U5A1B1C2 возник менее 5000 лет назад
- U5A1B1D
- U5A1B1E
- U5A1B1A возник между 2500 и 7500 годами
- U5A1B2
- U5A1B3
- U5A1B1 появился между 5000 и 9 000 лет назад
- U5A1C
- U5A1C1
- U5A1C2
- U5A1D возник около 19000 лет назад
- U5A1D1
- U5A1D2
- U5A1D2A
- U5A1E
- U5A1F
- U5A1A возник между 8000 и 16 000 лет назад
- U5A2 возник около 14000 лет назад
- U5A2A возникла около 6000 лет назад. Он был обнаружен в древнем мезолитическом образце (6000-5000 кал BCE) из пещеры Сантимаминьи в Баскской стране , Испания . [ 37 ]
- U5A2B возник около 8000 лет назад
- U5A2C возник около 13000 лет назад
- U5A2D обнаружил на мезолитическом участке Huseby Klev в Западной Швеции.
- U5A2E
- U5A1 возник между 14 000 и 20 000 лет назад. Обнаружено у этрусского человека (700-600 г. до н.э.) из Южной Этрурии , Италия . [ 35 ]
- U5B появился между 19 000 и 26 000 лет назад [ 38 ] и имеет полиморфизмы в 150 7768 14182 ( + полиморфизмы U5). Найдены среди сии -берберов Сива Оазис . [ 39 ]
- U5B1 возник между 11 000 и 20 000 лет назад. [ 40 ]
- U5B1A
- U5B1B: был найден в Саами из Скандинавии, финских и берберов Северной Африки, которые, как было установлено, разделяют чрезвычайно молодой филиал, в возрасте всего лишь ~ 9000 лет. U5B1B также был найден у людей Fulbe и Papel в Гвинеи-Бисау и Якутс людях с северо-восточной Сибири . [ 41 ] [ 42 ] Это возникло около 11000 лет назад.
- U5B1B1
- U5B1B1B Основной элемент в западных евразийских линии матери в Пуэрто -Рико , соответствующий образцам из Сенегамбии и северного Камеруна, указывающий на его присутствие как продукт ранней колонизации и порабощения сенегамбийцев. [ 43 ]
- U5B1B1
- U5B1C возник около 13 000 лет назад.
- U5B1D
- U5B1E возникла около 6600 лет назад. U5B1E в основном виден в центральной Европе среди чехов, словаков, венгров и южных русских. [ 44 ]
- U5B1G
- U5B2 возник между 17 000 и 23 000 лет назад [ 45 ] и имеет полиморфизмы в 1721 году 13637 ( + полиморфизмы U5B). Клада была обнаружена в останках, встречающихся с доисторических времен в Европе, таких как подклад U5B2C1 La Braña Man (найденный на участке La Braña в Испании). U5B2 встречается редко среди французских басккс (2,5%) и чаще в испанских басках.
- U5B2A от 12 000 до 19 000 лет назад, [ 46 ] Распространен в Центральной Европе. [ 44 ]
- U5B2A1 с 9 000 до 18 000 лет назад
- U5B2A2 с 7000 до 14 000 лет назад,
- U5B2A3 от 3000 до 14 000 лет назад
- U5B2A4 от 1000 до 10 000 лет назад
- U5B2A5 Менее 2600 лет назад
- U5B2A6 Менее 12 000 лет назад.
- U5B2B от 12 000 до 17 000 лет назад. [ 47 ] Клада была заметно связана с Неве, которая во время ее открытия была самой старой идентифицированной женской детской захородой в Европе, уравновешенным около 10 000 лет назад. [ 48 ] [ 49 ]
- U5B2C от 7000 до 18 000 лет назад. [ 50 ]
- U5B2C1 Менее 8000 лет назад. [ 51 ] Найден у финикийского человека из гробницы Карфагена в Бирса -Хилл , Тунис. [ 52 ]
- U5B2A от 12 000 до 19 000 лет назад, [ 46 ] Распространен в Центральной Европе. [ 44 ]
- U5B3: подклад, вероятно, происходит на итальянском полуострове; [ 44 ] Он находится на самом высоком распределении на юго -западе Европы, достигая пика среди сардинцев (3,84%), за которыми следуют балеарские из севера люди (1,56%) и португальские (1,09%). [ 53 ] Согласно другому исследованию, U5B3 встречается на частоте 2,53% среди основных мест и 0,96% среди Sephardi Chuetas . [ 54 ]
- U5B3B: Этот подклад, вероятно, также является таким же западным средиземноморским/ибералиатическим по происхождению, но распространяется на части северо-западной и средней Европы посредством римской экспансии, с образцами, найденными на Крите (Греция) , Испании , Центральной Италии , Англии , Германии Палатит и Богемии. Полем [ 55 ]
- U5B1 возник между 11 000 и 20 000 лет назад. [ 40 ]
- U5A возник около 17 000 и 27 000 лет назад
Гаплогруппа U6
[ редактировать ]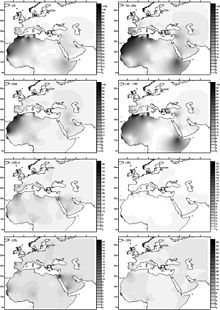
Гаплогруппа U6 была датирована от 31 000 до 43 000 лет назад Behar et al. (2012). Базальный U6* был найден в румынском образе древней ДНК ( Peștera muierilor ), датированного 35 000 лет назад. [ 56 ] Hervella et al. (2016) принять это в качестве доказательства палеолитической миграции Homo Sapiens из Евразии в Африку. Обнаружение базальной U6* в древней ДНК способствовало устранению расчетного возраста до 6 до 46 000 лет назад. [ 57 ]
Обычно генетическая история U6 рассматривается как миграция из Юго -Западной Азии через Северную Африку. Эта гипотеза основана на общем происхождении подкладов гаплогруппы U в Юго-Западной Азии, которая также является центром географического распределения UB-клад: Европа, Индия, Центральная Азия, Восточная Африка и Северная Африка. Два возможных сценария для первого гаплотипа U6 (Matering Mutations 3348 и 16172) могут быть выдвинуты: i) эти мутации, вызванные в области основателя, но не оставляли какого -либо генетического наследия в современных популяциях человека; ii) Они возникли, вероятно, где -то в Северной Африке, после прибытия гаплотипа основателя U6. В Северной Африке U6 находится значительно частым на его западном краю (а также в юго-западной Европе). Что еще более важно, все самые базальные ветви практически ограничены этой регионом (U6B, U6C и U6D), что может указывать на его западное происхождение. Тем не менее, он не может быть исключен из основного подклада U6A, который показывает богатство субклад в северо-западной Африке, хотя некоторые из производных филиалов также включают последовательности из восточноафриканских и ближневосточных популяций (например, U6A2).
Гаплогруппа U6 распространена (с распространенностью около 10%) [ 33 ] в северо -западной Африке (с максимум 29% в алжирских мозабитах [ 58 ] ) и Канарские острова (в среднем 18% с пиковой частотой 50,1% в La Gomera ). Это также найдено на Иберийском полуострове , где он имеет самое высокое разнообразие (10 из 19 сублинсов встречаются только в этом регионе, а не в Африке), [ 59 ] Северо -восточная Африка и иногда в других местах. U6 также встречается на низких частотах в бассейне Чад , включая редкую канарскую ветвь. Это говорит о том, что древние носители клады U6, возможно, заселивали или проходили через бассейн Чада на западе к Канарским островам. [ 60 ]
Считается, что U6 вошел в Северную Африку с Ближнего Востока около 30 000 лет назад. Это было найдено среди иберомавровских образцов, датируемых эпипалеолитом на доисторическом участке Taforalt . [ 61 ] Несмотря на самое высокое разнообразие иберийского U6, Мака-Мейер утверждает о ближнем восточном происхождении этой клады на основе самого высокого разнообразия подклада U6A в этом регионе, [ 59 ] Где это прибыло бы из Западной Азии, с иберийской частотой, в основном представляющей миграцию из Магриба , а не настойчивость европейского корневого населения. [ почему? ]
Согласно Hernández et al. 2015 «Расчетный вход североафриканских линий U6 в Iberia в 10 ky хорошо коррелирует с другими африканскими кладами L, что указывает на то, что U6 и некоторые линии L перешли вместе из Африки в Иберию в раннем голоцене ». [ 62 ]
U6 имеет четыре основных подклада: [ 59 ] [ 61 ]
Подгруппа U6A отражает первое африканское расширение от Магриба, возвращающегося на восток. Деривативный клад U6A1 сигнализирует о заднем движении от Восточной Африки обратно в Магриб и Ближний Восток. Эта миграция совпадает с вероятной афроазиатской лингвистической экспансией. U6B и U6C Clades, ограниченные Западной Африкой, имели более локализованные расширения. U6B, вероятно, достиг иберийского полуострова во время диффузии капсианской диффузии в Северной Африке. Два автохтонных производных этих клад (U6B1 и U6C1) указывают на прибытие североафриканских поселенцев к канарскому архипелагу в доисторические времена, скорее всего, из -за сахарского высыхания. Отсутствие этих канарских линий в настоящее время в Африке предполагает важные демографические движения в западной области этого континента.
- Macker 2003
- U6A: Подклад является наиболее распространенным, простирающимся от Канарских островов и иберийского полуострова до Африканского Рога и Ближнего Востока. Субхаплогруппа имеет самое высокое разнообразие в северо -восточной Африке. Древний анализ ДНК иберамовра скелетных останков в месте Тафоральта в Марокко , которые были датированы более поздним каменным веком между 15 100 до 13 900 YBP, наблюдал подклад U6A среди большинства окаменелостей (6/7; ~ 86%). [ 63 ] Окаменелости в раннем неолитическом месте Ифри Н'Амра или Муссы в Марокко, которые были обнаружены около 5000 г. до н. Эти древние люди имели автохтонную северо -западную африканскую геномную компонент, который достигает пика среди современных берберов , что указывает на то, что они были наследственными населениями в этом районе. [ 64 ] Расчетный возраст U6A составляет 24-27 500 п.н. У него есть один крупный подклад:
- U6B: показывает более исправленное и западное распределение. На иберийском полуострове U6B более часты на севере, тогда как U6A чаще встречается на юге. Он также был обнаружен на низких частотах в Марокко, Алжире, Сенегале и Нигерии. Расчетный возраст: 8500-24 500 п.н. У него есть один подплат:
- U6B1: найдено только на Канарских островах и на иберийском полуострове. Расчетный возраст: c. 6000 п.н.
- U6C: только найдено на Марокко и Канарских островах. Расчетный возраст: 6000-17 500 п.н.
- U6D: наиболее тесно связано с U6B. Локализован в Магрибе, с присутствием в Европе. Это возникло от 10 000 до 13 000 п.н.
U6A, U6B и U6D имеют общую базальную мутацию (16219), которой нет в U6C, тогда как U6C имеет 11 уникальных мутаций. U6B и U6D имеют мутацию (16311), не разделяемая U6A, которая имеет три уникальных мутации.
U2'3'4'7'8'9
[ редактировать ]Подклады U2, U3, U4, U7, U8 и U9 теперь считаются монофилетическими, их общий предок «U2'3'4'7'8'9», определяется мутацией A1811G, возникшим от 42 000 до 48 000 лет назад (Behar et al., 2012). Внутри U2'3'4'7'8'9, U4 и U9 могут быть монофилетическими, так как «U4'9» (мутации T195C!, G499A, T5999C), возникающие от 31 000 до 43 000 лет назад (Behar et al., 2012) Полем
U2'3'4'7'8'9 был найден в остатках двух 32 000 лет древних северных сибирцев (ANS) с участка Yana RHS на реке Яна . [ 66 ]
Гаплогруппа U2
[ редактировать ]Гаплогруппа U2 наиболее распространена в Южной Азии [ 67 ] но также встречается в низкой частоте в Центральной и Западной Азии, а также в Европе как U2E (европейское разнообразие U2 называется U2E). [ 68 ] Общая частота U2 в Южной Азии в значительной степени учитывается группой U2I в Индии, тогда как гаплогруппа U2E, обычная в Европе, редко; Учитывая, что эти линии расходились примерно на 50 000 лет, эти данные были интерпретированы как указывающие на очень низкий уровень материнской линии генов между Южной Азией и Европой на протяжении всего этого периода. [ 67 ] Приблизительно половина U-Mtdnas в Индии принадлежат к индийским ветвям гаплогруппы U2 (U2I: U2A, U2B и U2C). Гаплогруппа U2B2 была обнаружена в остатках 4500 -летней женщины, раскопанной на месте Рахигархи Цивилизации долины Инда , в современном штате Харьяна, Индия. [ 69 ] В то время как U2 обычно встречается в Индии, [ 67 ] Он также присутствует в Nogais , потомках различных монгольских и туркских племен, которые сформировали Орду Ногай . [ 70 ] Как U2, так и U4 встречаются в народах Кет и Нгагасан , коренных жителях бассейна реки Енисей и на полуострове Таймир . [ 71 ]
Подклады U2: U2A, [ 72 ] U2B, [ 73 ] U2C, [ 74 ] U2D, [ 75 ] и U2E. [ 76 ] С специфичными для Индии подклады U2A, U2B и U2C в совокупности, называемых U2I, евразийская гаплогруппа U2D, по-видимому, является родственной кладкой с индийской гаплогруппой U2C, [ 77 ] в то время как U2E считается европейским специфическим подкладом, но также и встречается в Южной Индии. [ 68 ]
Гаплогруппа U2 была обнаружена в остатках 37 000 [ 78 ] и 30 000-летний охотник-собиратель из Костионки, Воронежский вскрытия в Центральной Южной Европе России. [ 79 ] В 4800-4000-летних человеческих останках из культурного участка покойного неолита в Кромдорф Германии, [ 80 ] и у 2000-летнего человеческого останка от Bøgebjerggård на юге Дании. Тем не менее, гаплогруппа U2 встречается редко у современных скандинавцев. [ 81 ] Остатки 2000-летнего западно-евразийского мужчины гаплогруппы U2E1 были обнаружены на кладбище Xiongnu на северо-восточной Монголии. [ 82 ]
Гаплогруппа U3
[ редактировать ]Гаплогруппа U3 делится на два подклада :: u3a [ 83 ] и U3b. [ 83 ]
Возраст коацензии для U3A оценивается в 18 000 до 26 000 лет, в то время как возраст слияния для U3B оценивается как от 18 000 до 24 000 лет. U3A встречается в Европе, на Ближнем Востоке, Кавказе и Северной Африке. Почти по европейским распределенным подкладам U3A1, от 4000 до 7000 лет, предполагает относительно недавнее (позднее голоценовое или более позднее) расширение этих линий в Европе. Существует незначительный подклад U3C (получен из U3A), представленную одной азербаймстским мтДНК из Кавказа. U3B широко распространен на Ближнем Востоке и Кавказе, и он обнаруживается, особенно в Иране, Ираке и Йемене, с незначительным европейским подкладом U3B1B, от 2000 до 3000 лет. [ 84 ] Гаплогруппа U3 определяется HVR1 переходом A16343G. Он обнаруживается на низких уровнях по всей Европе (около 1% населения), на Ближнем Востоке (около 2,5% населения) и Центральной Азии (около 1% населения). U3 присутствует в популяции Свана из региона Сванети (около 4,2% населения) и среди литовских цыган, польских романи и испанских римья (36-56%) [ 85 ] [ 86 ] [ 87 ] [ 88 ]
Клада U3 также найдена среди мозабитских берберов (10,59%), [ 19 ] а также египтяне в Эль-Хейе (2,9%) [ 89 ] и герна оазисы (2,9%), [ 90 ] и алжирцы в Оране (1,08%-1,25%). [ 19 ] Редкий подклад U3A происходит среди туарега, населяющих Нигер (3,23%) [ 91 ] и среди сомалийцев (1,6%). [ 92 ]
Гаплогруппа U3 была обнаружена в некоторых из 6400-летних останков (U3A), обнаруженных в пещерах в Вади-эль-Маккух, недалеко от Иерихона, связанного с халколитическим периодом. [ 93 ] Haplogroup U3 уже присутствовала в западном евразийском геном пуле около 6000 лет, а также, вероятно, также его подклада U3A. [ 93 ]
Гаплогруппа U4
[ редактировать ]Гаплогруппа U4 имеет свое происхождение между 21 000 и 14 000 лет назад. Его распределение связано с узким местом населения из -за последнего ледникового максимума . [ 85 ]
U4 был найден в древней ДНК, [ 94 ] и это относительно редко в современных популяциях, [ 44 ] Хотя он обнаруживается в значительных соотношениях в некоторых коренных населениях Северной Азии и Северной Европы, связанные с остатками древних европейских охотников-собирателей, сохранившихся в коренном населении Сибири. [ 95 ] [ 96 ] [ 97 ] U4 встречается в исчезающем народе Nganasan на полуострове Таймир, [ 71 ] [ 98 ] в манси (16,3%), [ 97 ] и в кете (28,9%) реки Йенисей. [ 97 ] Он встречается в Европе с самыми высокими концентрациями в Скандинавии и в Балтийских штатах. [ 99 ] и встречается в сами -популяции скандинавского полуострова (хотя U5B имеет более высокое представление). [ 100 ] U4 также сохранился у людей Калаш (текущий размер населения 3700) [ 101 ] Уникальное племя среди индоарийских народов Пакистана, где U4 (подклад U4A1 [ 102 ] ) достигает самой высокой частоты 34%. [ 86 ] [ 103 ]
Подклады U4: U4A, [ 104 ] U4b, [ 105 ] U4C, [ 106 ] и U4D. [ 107 ]
Гаплогруппа U4 связана с древними европейскими охотниками-собирателями и была обнаружена у 7200-6000-летних остатков в стиле « Культура по борьбе» в Готланде Швеции и в от 4 400 до 3800-летнего останков с датчика датской культуры стакана Полем [ 108 ] [ 27 ] [ 109 ] Останки, идентифицированные как подклад U4A2, связаны с культурой проводной посуды , которая процветала от 5200 до 4300 лет назад в Восточной и Центральной Европе и охватывала большую часть континентальной северной Европы от реки Волги на востоке до Рейна на западе. [ 110 ] Митохондриальная ДНК, извлеченная у 3500 до 3300-летних, остается на участке Bredtoftegård в Дании, связанном с северным бронзовым веком, включают гаплогруппу U4 с 16179t в своем HVR1, указывающем на подклад U4C1. [ 109 ] [ 111 ] [ 112 ] 2 из 9 1700-летних остались на крайнем юго-западе региона Иваново были U4C1. [ 113 ]
Гаплогруппа U7
[ редактировать ]Гаплогруппа U7 считается западной евразийской гаплогруппой мтДНК, которая, как полагают, возникла в районе Черного моря примерно 30 000 лет назад. [ 67 ] [ 114 ] [ 115 ] В современных популяциях U7 встречается на низкой частоте в Кавказе, [ 115 ] западные сибирские племена, [ 116 ] Западная Азия (около 4% на Ближнем Востоке, а также пик с 10% в иранцах), [ 67 ] Южная Азия (около 12% в Гуджарате, самом западном штате Индии, в то время как для всей Индии его частота остается около 2% и 5% в Пакистане ), [ 67 ] и жители Ведды Шри -Ланки, где он достигает самой высокой частоты 13,33% (подклад U7A). [ 117 ] Одна треть из западно-евразийских мтдн, найденных в Индии, находятся в гаплогруппах U7 R2 и W. , Предполагается, что крупномасштабная иммиграция перенесла эти митохондриальные гаплогруппы в Индию. [ 67 ]
Подклады U7-U7A (с глубокими подкладами U7A1, U7A2, U7A2A, U7A2B) [ 118 ] и U7B. [ 118 ]
Генетический анализ лиц, связанных с поздней культурой Халлстатта из Баден-Вюртемберга Германии, считается примерами «княжеского захоронения железного века », включал гаплогруппу U7. [ 119 ] Сообщалось, что гаплогруппа U7 была обнаружена у 1200-летних человеческих останков (датируется около 834 года), у женщины, которая, как считается, была из королевского клана, который был похоронен на корабле Осеберга в Норвегии. [ 120 ] Гаплогруппа U7 была обнаружена у 1000-летних человеческих останков (датируемые около 1000-1250 г. до н. Тем не менее, U7 встречается редко среди современных этнических скандинавцев. [ 116 ]
Подклад U7A особенно распространен среди саудовцев , составляющий около 30% материнских линий в восточной провинции . [ 121 ]
Гаплогруппа U8
[ редактировать ]- Гаплогруппа U8A: басккс обладает наиболее наследственной филогениями в Европе для митохондриальной гаплогруппы U8A. Это редкая подгруппа U8, размещающая баскское происхождение этой линии в верхнем палеолите. Отсутствие линий U8A в Африке предполагает, что их предки, возможно, произошли из Западной Азии. [ 15 ]
- Гаплогруппа U8B: эта клада была найдена в Италии и Иордании. [ 15 ]
- Aplogroup U8B'K: Эта клада может быть синонимом гаплогруппы K и Haplogroup UK . [ Цитация необходима ]
Наиболее распространенным подкладом гаплогруппы U8B является гаплогруппа K , которая оценивается на сегодняшний день от 30 000 до 22 000 лет назад. [ Цитация необходима ] Гаплогруппа K составляет значительную часть линий европейских и западно -азиатских мтДНК. Теперь известно, что это на самом деле подклад гаплогруппы U8B'K, [ 15 ] и, как полагают, впервые появился в северо -восточной Италии. Haplogroup UK показывает некоторые доказательства того, что он высоко защищает от прогрессирования СПИДа. [ 34 ]
Гаплогруппа U9
[ редактировать ]Гаплогруппа U9 является редкой кладой в филогении мтДНК, характеризующейся лишь недавно в нескольких популяциях Пакистана (Quintana-Murci et al. 2004). Его присутствие в Эфиопии и Йемене , вместе с некоторыми индийскими индийскими минами в образце Йемена, указывает на поток генов вдоль побережья Аравийского моря. Гаплогруппы U9 и U4 имеют две общие мутации в корне их филогения. Интересно, что в Пакистане U9 часто встречается только среди так называемой популяции макрани . В этой конкретной популяции линии, специфичные для частей восточной Африки, встречаются до 39%, что говорит о том, что линии U9 в Пакистане могут иметь африканское происхождение (Quintana-Murci et al. 2004). Независимо от того, какое побережье Аравийского моря могло быть происхождением U9, его эфиопско -южно -южно -южно -арабский бассейн намека Гаплогруппа U4 - Восточная Европа и Западная Сибирия. [ 122 ]
Известные члены
[ редактировать ]- Доктор Санджай Гупта несет гаплогруппу U2C. [ 123 ]
- Политический аналитик Линда Чавес является членом гаплогруппы U5A1A1. [ 124 ]
- Генетический генеалогист Сеса Мур принадлежит к гаплогруппе U5B1B2. [ 125 ]
Смотрите также
[ редактировать ]- Генеалогический тест ДНК
- Генетическая генеалогия
- Митохондриальная генетика
- Генетика популяции
- Семь дочерей Евы
|
Филогенетическое дерево митохондриальной ДНК (мтДНК) человеческой ДНК (мтДНК) | |||||||||||||||||||||||||||||||||||||||
| Митохондриальная канун ( L ) | |||||||||||||||||||||||||||||||||||||||
| L0 | L1–6 | ||||||||||||||||||||||||||||||||||||||
| L1 | L2 | L3 | L4 | L5 | 16 -й | ||||||||||||||||||||||||||||||||||
| М | Не | ||||||||||||||||||||||||||||||||||||||
| Сумка | Дюймовый | И | Глин | Q. | А | А | С | Ведущий | я | В | Х | И | |||||||||||||||||||||||||||
| В | С | Беременный | Фон | R0 | до- | П | В | ||||||||||||||||||||||||||||||||
| Который | JT | K | |||||||||||||||||||||||||||||||||||||
| ЧАС | V | Дж | Т | ||||||||||||||||||||||||||||||||||||
Ссылки
[ редактировать ]- ^ Behar et al. (2012), обратите внимание на пересмотренную оценку 42–58 ключа на основе древней ДНК Hervella et al. (2016).
- ^ Hervella et al. (2016) «Лица, несущие гаплогруппу U, возможно, распространяются на запад от Западной Азии около 39–52 KY, достигнув Европы, как оказалось гаплогруппой U5, и Северная Африка, которую сигнализировали гаплогруппа U6»
- ^ Jump up to: а беременный Van Oven M, Kayser M (февраль 2009 г.). «Обновленное комплексное филогенетическое дерево глобального изменения митохондриальной ДНК человека» . Человеческая мутация . 30 (2): E386-94. doi : 10.1002/Humu.20921 . PMID 18853457 . S2CID 27566749 .
- ^ Larruga JM, Marrero P, Abu-Amero KK, Golubenko MV, Cabrera VM (май 2017). «Перевозчики митохондриальной ДНК макрохаплогруппы R колонизировали Евразию и Австралазию из ядра Юго -Восточной Азии» . BMC Эволюционная биология . 17 (1): 115. Bibcode : 2017bmcee..17..115L . doi : 10.1186/s12862-017-0964-5 . PMC 5442693 . PMID 28535779 . Марреро П., Абу-Амеро К.К., Ларруга Дж.М., Кабрера В.М. (ноябрь 2016 г.). «Перевозчики человеческой митохондриальной ДНК макрохаплогруппы M колонизировали Индию из юго -восточной Азии» . BMC Эволюционная биология . 16 (1): 246. Bibcode : 2016bmcee..16..246m . doi : 10.1186/s12862-016-0816-8 . PMC 5105315 . PMID 27832758 .
- ^ Hervella M, Svensson EM, Alberdi A, Günther T, Izagirre N, Munters AR, et al. (Май 2016 г.). «Митогеном 35 000-летних Homo Sapiens из Европы поддерживает палеолитную обратную миграцию в Африку» . Научные отчеты . 6 : 25501. BIBCODE : 2016NATSR ... 625501H . doi : 10.1038/srep25501 . PMC 4872530 . PMID 27195518 . Полем
- ^ Secher B, Fregel R, Larruga JM, Cabrera VM, Endicott P, Pestano JJ, González AM (май 2014). «История североафриканской митохондриальной ДНК -гаплогруппы U6 течет на континенты африканских, евразийских и американских» . BMC Эволюционная биология . 14 (1): 109. Bibcode : 2014bmcee..14..109S . doi : 10.1186/1471-2148-14-109 . PMC 4062890 . PMID 24885141 .
- ^ Kefi R, Hechmi M, Naouali C, Jmel H, Hsouna S, Bouzaid E, et al. (Январь 2018). «О происхождении иберамовра: новые данные, основанные на древней митохондриальной ДНК и филогенетическом анализе популяций афалу и тафоральта». Митохондриальная ДНК часть а . 29 (1): 147–157. doi : 10.1080/24701394.2016.1258406 . PMID 28034339 . S2CID 4490910 .
- ^ Schuenemann VJ, Peltzer A, Welte B, Van Pelt WP, Molak M, Wang CC, et al. (Май 2017). «Древняя египетская мумия геномы предполагают увеличение африканского происхождения к югу от Сахары в пост-римские периоды» . Природная связь . 8 : 15694. Bibcode : 2017natco ... 815694S . doi : 10.1038/ncomms15694 . PMC 5459999 . PMID 28556824 .
- ^ Loreille, O.; Ratnayake, S.; Базинет, Ал; Стоквелл, туберкулез; Соммер, DD; Rohland, N.; Mallick, S.; Джонсон, PL; Skoglund, P.; Onorato, AJ; Бергман, NH; Рейх, Д.; Ирвин, JA (2018). «Биологическое полов 4000-летней египетской мумии, чтобы оценить потенциал восстановления ядерной ДНК от наиболее поврежденных и ограниченных судебных образцов» . Гены . 9 (3): 135. doi : 10.3390/genes9030135 . PMC 5867856 . PMID 29494531 .
- ^ Rodríguez-Varela R, Günther T, Krzewińska M, Storå J, Gillingwater TH, Maccallum M, et al. (Ноябрь 2017). «Геномный анализ доевропейских завоеваний человеческих останков с Канарских островов раскрывает тесную близость к современным североафриканцам» . Текущая биология . 27 (21): 3396–3402.e5. Bibcode : 2017cbio ... 27e3396r . doi : 10.1016/j.cub.2017.09.059 . HDL : 2164/13526 . PMID 29107554 .
- ^ Jump up to: а беременный Кармин М (2005). Митохондриальная ДНК -гаплогруппа R в Индии (PDF) (диссертация M.Sc.). Университет Тарту.
- ^ Sykes B (2001). Семь дочерей Евы . Лондон; Нью -Йорк: Bantam Press. ISBN 978-0-393-02018-2 .
- ^ «Материнское происхождение» . Оксфордские предки. Архивировано из оригинала 15 июля 2017 года . Получено 7 февраля 2013 года .
- ^ Фу Q, Миттник А., Джонсон П.Л., Бос К., Лари М., Боллонгино Р. и др. (Апрель 2013). «Пересмотренная временная масштаба для эволюции человека, основанная на древних митохондриальных геномах» . Текущая биология . 23 (7): 553–559. Bibcode : 2013cbio ... 23..553f . doi : 10.1016/j.cub.2013.02.044 . PMC 5036973 . PMID 23523248 .
- ^ Jump up to: а беременный в дюймовый Гонсалес А.М., Гарсия О., Ларруга Дж. М., Кабрера В.М. (май 2006 г.). «Митохондриальная линия U8A раскрывает палеолитическое поселение в баскской стране» . BMC Genomics . 7 (1): 124. doi : 10.1186/1471-2164-7-124 . PMC 1523212 . PMID 16719915 .
- ^ Jump up to: а беременный Скотт Р. (2010). «Филогения MT-HG U3» (PDF) .
- ^ Брук, Кевин Алан (2022). Материнские генетические линии ашкеназских евреев . Академические исследования пресса. п. 100-101. ISBN 978-1-64469-984-3 .
- ^ «Проект гаплогруппы мтДНК U1» . Семейство .
- ^ Jump up to: а беременный в Бекада А., Араа Л.Р., Дебат Т., Калафелл Ф., Бенхамамуш С., Комы Д (24 сентября 2015 г.). «Генетическая гетерогенность в алжирских популяциях» . Plos один . 10 (9): E0138453. BIBCODE : 2015PLOSO..1038453B . Doi : 10.1371/journal.pone.01384453 . PMC 4581715 . PMID 26402429 .
- ^ Jump up to: а беременный Molto JE, Loreille O, Mallott EK, Malhi RS, Fast S, Daniels-Higginbotham J, et al. (Октябрь 2017). «Полное митохондриальное секвенирование погребения с римско-христианского кладбища в Оазисе Дахле, Египет: предварительные признаки» . Гены . 8 (10): 262. doi : 10.3390/genes8100262 . PMC 5664112 . PMID 28984839 .
- ^ Сирак К., Френанд Д., Новак М., Ван Герсен Д., Пинхаси Р. (2016). «Абстрактная книга IUAES Inter-Congress 2016- сообщество, разделенное? Раскрытие генома сообщества средневекового кулубнарти с использованием секвенирования следующего поколения» . Абстрактная книга IUAES Intercongress 2016 . IUAES: 115.
- ^ Jump up to: а беременный Рай Н., Татье Н., Сингх М., Чауби Г., Джа А.Н., Сингх Л., Тангара К (январь 2014 г.). «Реликвия раскопана в западной Индии, вероятно, из грузинской королевы Кетевана» Митохондрион 14 (1): 1–6 Doi : 10.1016/ j.mito.2013.12.0 24355295PMID
- ^ "Гаплогруппа U5" . haplogroup.org . 2016-06-06. Архивировано из оригинала 2017-07-03 . Получено 2017-07-13 .
30 248,3 ± 5,330,5; CI = 95% (Behar et al., 2012)
- ^ Brandstätter, Anita; Циммерманн, Беттина; Вагнер, Джанин; Гёбель, Танджа; Рёк, Александр В.; Салас, Антонио; Carrecedo, ангел; Parson, Walther (2008-07-04). «Время и расшифровка митохондриальной ДНК макро-ха-плагруппы R0 изменчивость в центральной Европе и на Ближнем Востоке» . BMC Эволюционная биология . 8 (1): 191. Bibcode : 2008bmcee ... 8..191b . doi : 10.1186/1471-2148-8-191 . ISSN 1471-2148 . PMC 2491632 . PMID 18601722 .
- ^ Soares, Pedro; Ахилли, Алессандро; Semino, Ornella; Дэвис, Уильям; Маколей, Винсент; Бандельт, Ханс-Юрген; Торрони, Антонио; Ричардс, Мартин Б. (2010-02-23). «Археогенетика Европы» . Текущая биология . 20 (4): R174 - R183. Bibcode : 2010cbio ... 20.r174s . doi : 10.1016/j.cub.2009.11.054 . PMID 20178764 .
- ^ British B, Thomas Mg, Hak W, Unterlaender M, Jores P, Tambedts K, et al. (Октябрь 2009 г.). «Генетический разрыв между местными охотниками и первыми фермерами в Центральной Европе » Наука . 326 (5949): 137–4 Bibcode : 2009sci ... 326..137b Doi : 10.1126/ science.1 19729620PMID S2CID 206521424
- ^ Jump up to: а беременный Malmström H, Gilbert MT, Thomas MG, Brandström M, Storå J, Molnar P, et al. (Ноябрь 2009 г.). «Древняя ДНК раскрывает отсутствие преемственности между неолитическими охотниками-собирателями и современными скандинавтами» . Текущая биология . 19 (20): 1758–62. Bibcode : 2009cbio ... 19.1758m . doi : 10.1016/j.cub.2009.09.017 . PMID 19781941 . S2CID 9487217 .
- ^ Deguilloux MF, Soler L, Pemonge MH, Scarre C, Joussaume R, Laporte L (январь 2011 г.). «Новости с Запада: древняя ДНК из французской мегалитической захоронения». Американский журнал физической антропологии . 144 (1): 108–18. doi : 10.1002/ajpa.21376 . PMID 20717990 .
- ^ Sánchez-Quinto F, Schroeder H, Ramirez O, Avila-Arcos MC, Pybus M, Olalde I, et al. (Август 2012 г.). «Геномное сродство двух 7000-летних иберийских охотников-собирателей» (PDF) . Текущая биология . 22 (16): 1494–9. Bibcode : 2012cbio ... 22.1494S . doi : 10.1016/j.cub.2012.06.005 . PMID 22748318 . S2CID 14913530 .
- ^ Lacan M, Keyser C, Ricaut FX, Brucato N, Tarrús J, Bosch A, et al. (Ноябрь 2011). «Древняя ДНК предполагает ведущую роль, которую играют мужчины в неолитическом распространении» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (45): 18255–9. BibCode : 2011pnas..10818255L . doi : 10.1073/pnas.1113061108 . PMC 3215063 . PMID 22042855 .
- ^ "Гаплогруппа U5" . Епипедия .
- ^ «Басская генетика: рефераты и резюме» . Кажария .
- ^ Jump up to: а беременный «Генографический проект» . National Geographic . Архивировано из оригинала 13 апреля 2005 года.
- ^ Jump up to: а беременный Hendrickson SL, Hutcheson HB, Ruiz-Pesini E, Poole JC, Lautenberger J, Sezgin E, et al. (Ноябрь 2008 г.). «Митохондриальные ДНК -гаплогруппы влияют на прогрессирование СПИДа» . СПИД . 22 (18): 2429–39. doi : 10.1097/QAD.0B013E32831940BB . PMC 2699618 . PMID 19005266 .
- ^ Антонио, Маргарет Л.; Гао, Ziyue; M. Moots, Hannah (2019). «Древний Рим: генетический перекресток Европы и Средиземного моря» . Наука . 366 (6466). Вашингтон, округ Колумбия: Американская ассоциация развития науки (опубликовано 8 ноября 2019 г.): 708–714. Bibcode : 2019sci ... 366..708a . doi : 10.1126/science.aay6826 . PMC 7093155 . PMID 31699931 .
- ^ Возраст: 1398,1 ± 1,872,3; Ci = 95% (Behar et al., 2012) haplogroup.org [ Постоянная мертвая ссылка ]
- ^ Palencia-Madrid L, Cardo S, Keyser C, López-Quintana JC, с Pancorbo MM (май 2017 г.). «Древние митохондриальные линии подтверждают доисторический материнский корень баски на северном иберийском полуострове » Европейский журнал человеческой генетики 25 (5): 631–6 Doi : 10.1038/ ejhg.2017.2 5437903PMC PMID 28272540
- ^ 22 794,0 ± 3590,3; CI = 95% (Behar et al., 2012) Haplogroup.org Archived 2017-10-11 на машине Wayback
- ^ Coudray C, Olivieri A, Achilli A, Pala M, Melhaoui M, Cherkaoui M, et al. (Март 2009 г.). «Сложный и диверсифицированный митохондриальный генный пул берберских популяций». Анналы человеческой генетики . 73 (2): 196–214. doi : 10.1111/j.1469-1809.2008.00493.x . PMID 19053990 . S2CID 21826485 .
- ^ Возраст: 15 529,7 ± 4889,9; CI = 95% "МтДНК Гаплогруппа U5B1" . haplogroup.org . 2016-06-06. Архивировано из оригинала 2017-03-22 . Получено 2017-07-13 .
- ^ Роза А., Орнелас С., Джоблс М.А., Брем А., Виллемс Р. (июль 2007 г.). «Y-хромосомное разнообразие в популяции Гвинеи-Бисау: многонациональная перспектива» . BMC Эволюционная биология . 7 (1): 124. Bibcode : 2007bmcee ... 7..124r . doi : 10.1186/1471-2148-7-124 . PMC 1976131 . PMID 17662131 .
- ^ Achilli A, Rengo C, Battaglia V, Pala M, Olivieri A, Fornarino S, et al. (Май 2005 г.). «Саами и Берберс-неожиданная митохондриальная связь ДНК» . Американский журнал человеческой генетики . 76 (5): 883–6. doi : 10.1086/430073 . PMC 1199377 . PMID 15791543 .
- ^ Héctor DZ, María NC, Juan MC (апрель 2017 г.). «В основном обход средиземноморского происхождения для западно -евразийских и североафриканских мтдн в Пуэрто -Рико с сильным вкладом Канарских островов и Западной Африки». Человеческая биология . 89 (2): 125–155. doi : 10.13110/HumanBiology.89.2.04 . PMID 29299964 . S2CID 21941155 .
- ^ Jump up to: а беременный в дюймовый Malyarchuk B, Derenko M, Grzybowski T, Perkova M, Rogalla U, Vanecek T, et al. (Апрель 2010). Гилберт (ред.). «Народ Европы с точки зрения митохондриальной гаплогруппы U5» . Plos один . 5 (4): E10285. BIBCODE : 2010PLOSO ... 510285M . doi : 10.1371/journal.pone.0010285 . PMC 2858207 . PMID 20422015 .
Субкластер U5B2A характеризуется преимущественно центральным европейским распределением, поскольку большое количество образцов U5B из Польши, Словакии и Чешской Республики попадают в этот субкластер. Например, субкластер U5B2A2 часто встречается в Центральной Европе (с самой высокой частотой его подкластерного U5B2A2A1 на полюсах) и датируется как возникающая между 12–18 KYA [...] также примечательно, что в US5B2A1A среди серединераранского ветви Subcluster U5B2A1A1. , что характерно для центральной и восточной европейской популяции (рис. 1). Другой подкластер U5B, U5B3, имеет свою наиболее вероятную родину на итальянском полуострове, откуда он расширился в голоцене вдоль средиземноморских побережья [11]. Следовательно, в целом можно сделать вывод, что первоначальная диверсификация U5B произошла в южной и центральной Европе, после чего последовало распространение конкретных подкластеров U5B в Восточную Европу.
- ^ 20,040,2 ± 3208,6; CI = 95% (Behar et al., 2012) Haplogroup.org Archived 2017-06-17 на машине Wayback
- ^ 14,938,0 ± 3656,4; CI = 95% (Behar et al., 2012) Haplogroup.org Archived 2017-06-17 на машине Wayback
- ^ Возраст: 14 676,9 ± 2716,6; CI = 95% (Behar et al., 2012) haplogroup.org Archived 2017-07-27 на машине Wayback
- ^ Меткалф, Том (14 декабря 2021 г.). «Самая ранняя современная женщина -младенец, найдено в Европе» . National Geographic . Архивировано из оригинала 14 декабря 2021 года . Получено 16 декабря 2021 года .
- ^ Ходжкинс, Джейми (2021). «Похороны младенца из Арма Вейраны в северо -западной Италии дают представление о похоронных практиках и личности женской в ранней мезолитической Европе» . Научные отчеты . 11 (1): 23735. Bibcode : 2021natsr..1123735H . doi : 10.1038/s41598-021-02804-z . PMC 8671481 . PMID 34907203 .
- ^ Возраст: 12 651,1 ± 5668,2; CI = 95% (Behar et al., 2012) haplogroup.org Archived 2017-12-28 на машине Wayback
- ^ 4015,9 ± 3898,7; CI = 95% (Behar et al., 2012) haplogroup.org Archived 2017-09-06 на машине Wayback
- ^ Matisoo-Smith EA, Gosling AL, Boocock J, Kardailsky O, Kurumilian Y, Roudesli-Chebbi S, et al. Matisoo-Smith EA, Gosling AL, Boocock J, Kardailsky O, Kurumilian Y, Roudesli-Chebbi S, et al. (25 мая 2016 г.). «Европейский митохондриальный гаплотип, идентифицированный в древних финикийских останках из Карфагена, Северная Африка» . Plos один . 11 (5): E0155046. BIBCODE : 2016PLOSO..1155046M . doi : 10.1371/journal.pone.0155046 . PMC 4880306 . PMID 27224451 .
Молодой человек из образца Бирсы датируется концом 6 -го века до нашей эры, и, как полагают, его происхождение представляет ранний поток генов из Иберии к Магрибу.
- ^ Пала М., Ахилли А., Оливьери А., Хошиар Кашани Б., Перего Уа, Санна Д. и др. (Июнь 2009 г.). «Митохондриальная гаплогруппа U5B3: отдаленное эхо эпипалеолита в Италии и наследие ранних сардинцев» (PDF) . Американский журнал человеческой генетики . 84 (6): 814–21. doi : 10.1016/j.ajhg.2009.05.004 . PMC 2694970 . PMID 19500771 . Архивировано из оригинала (PDF) 2018-07-22 . Получено 2009-09-02 .
- ^ Farragut SJ (июнь 2017 г.). отцовская и материя Генетическое наследие сефардных сефардных евреев : Университет Балеарских островов (тезис).
- ^ Пала М., Ахилли А., Оливьери А., Хошиар Кашани Б., Перего Уа, Санна Д. и др. (Июнь 2009 г.). «Митохондриальная гаплогруппа U5B3: отдаленное эхо эпипалеолита в Италии и наследие ранних сардинцев» (PDF) . Американский журнал человеческой генетики . 84 (6): 814–21. doi : 10.1016/j.ajhg.2009.05.004 . PMC 2694970 . PMID 19500771 .
- ^ Soficaru A, Dobos A, Trinkaus E (ноябрь 2006 г.). «Ранние современные люди из Пестера Муерия, Байя де Фрир, Румыния» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (46): 17196–201. Bibcode : 2006pnas..10317196S . doi : 10.1073/pnas.0608443103 . PMC 1859909 . PMID 17085588 .
- ^ Hervella M, Svensson EM, Alberdi A, Günther T, Izagirre N, Munters AR, et al. (Май 2016 г.). «Митогеном 35 000-летних Homo Sapiens из Европы поддерживает палеолитную обратную миграцию в Африку» . Научные отчеты . 6 : 25501. BIBCODE : 2016NATSR ... 625501H . doi : 10.1038/srep25501 . PMC 4872530 . PMID 27195518 . "Люди, несущие гаплогруппу, возможно, распространяются на запад от Западной Азии около 39–52 тыс. Кентукки, достигая Европы, как оказалась гаплогруппой U5, и Северная Африка, о которой говорится гаплогруппа U6, которая, вероятно, представляет собой генетический сигнал возвращения eup от европий на север Африка. линии U6 обратно до 49,6 KY BP (95% HPD: 42–58 KY) "
- ^ Plaza S, Calafell F, Helal A, Bouzerna N, Lefranc G, Bertranpetit J, Comas D (июль 2003 г.). «Присоединение к столбам Геркулеса: последовательности мтДНК показывают многонаправленный поток генов в западном Средиземноморье» (PDF) . Анналы человеческой генетики . 67 (Pt 4): 312–28. doi : 10.1046/j.1469-1809.2003.00039.x . PMID 12914566 . S2CID 11201992 .
- ^ Jump up to: а беременный в Maca-Meyer N, González AM, Pestano J, Flores C, Larruga JM, Cabrera VM (октябрь 2003 г.). «Митохондриальный ДНК -транзит между Западной Азией и Северной Африкой выведен из филогеографии U6» . BMC Genetics . 4 (1): 15. doi : 10.1186/1471-2156-4-15 . PMC 270091 . PMID 14563219 .
- ^ Cerezo M, černý V, Carredcodo A, Salas A (апрель 2011 г.). «Новое понимание структуры популяции бассейна озера Чад, выявленное с помощью высокопроизводительного генотипирования митохондриальной ДНК, кодирующих SNP» . Plos один . 6 (4): E18682. BIBCODE : 2011PLOSO ... 618682C . Citeseerx 10.1.1.291.8871 . doi : 10.1371/journal.pone.0018682 . PMC 3080428 . PMID 21533064 .
- ^ Jump up to: а беременный Secher B, Fregel R, Larruga JM, Cabrera VM, Endicott P, Pestano JJ, et al. (Май 2014). «История североафриканской митохондриальной ДНК -гаплогруппы U6 течет на континенты африканских, евразийских и американских» . BMC Эволюционная биология . 14 (1): 109. Bibcode : 2014bmcee..14..109S . doi : 10.1186/1471-2148-14-109 . PMC 4062890 . PMID 24885141 .
- ^ Hernández CL, Soares P, Dugoujon JM, Novelletto A, Rodríguez JN, Rito T, et al. (2015). «Ранние голоценические и исторические африканские признаки мтДНК на иберийском полуострове: Андалузский регион как парадигма» . Plos один . 10 (10): E0139784. BIBCODE : 2015PLOSO..1039784H . doi : 10.1371/journal.pone.0139784 . PMC 4624789 . PMID 26509580 .
- ^ Van de Loosdrecht M, Bouzouggar A, Humphrey L, Poth C, Barton N, Aximu-Petri A, et al. (Май 2018). «Геномы плейстоцена в североафриканском языке связывают ближневосточный и странный африканский человеческий популяции африканского и субзара» . Наука . 360 (6388): 548–552. Bibcode : 2018sci ... 360..548v . doi : 10.1126/science.aar8380 . PMID 29545507 .
- ^ Fregel R, Méndez FL, Bokbot Y, Martín-Socas D, Camalich-Massieu MD, Santana J, et al. (Июнь 2018 г.). «Древние геномы из Северной Африки свидетельствуют о доисторических миграциях в Магриб из Леванта и Европы» . Труды Национальной академии наук Соединенных Штатов Америки . 115 (26): 6774–6779. Bibcode : 2018pnas..115.6774f . doi : 10.1073/pnas.1800851115 . PMC 6042094 . PMID 29895688 .
- ^ Мохамед Хи. Генетические закономерности Y-хромосомы и митохондриальной ДНК-вариации, что является последствиями для людей Судана (диссертация доктора философии). Университет Хартума. Архивировано из оригинала (PDF) 10 ноября 2020 года . Получено 17 апреля 2016 года .
- ^ Сикора, Мартин; и др. (2019). «История населения северо -восточной Сибири со времен плейстоцена» (PDF) . Природа . 570 (7760): 182–188. Bibcode : 2019natur.570..182S . Biorxiv 10.1101/448829 . doi : 10.1038/s41586-019-1279-z . PMID 31168093 . S2CID 174809069 .
- ^ Jump up to: а беременный в дюймовый и фон глин Metspalu M, Kivisild T, Metspalu E, Parik J, Hudjashov G, Kaldma K, et al. (Август 2004 г.). «Большинство существующих границ мтДНК в Южной и Юго -Западной Азии, вероятно, были сформированы во время первоначального поселения Евразии анатомически современными людьми» . BMC Genetics . 5 : 26. doi : 10.1186/1471-2156-5-26 . PMC 516768 . PMID 15339343 .
- ^ Jump up to: а беременный Маджи С., Критика С., Васулу Т.С. (март 2008 г.). В Индии. (PDF ) Международный журнал генетики человека 8 (1–2): 85–96. doi : 10.1080/ 0972375.2008.1186022 14231815S2CID
- ^ Shinde, V; Narasimhan, VM; Rohland, N; Mallick, S; Mah, M; Lipson, M; Nakatsuka, N; Adamski, N; Broomandkhoshbacht, N; Ferry, M; Lawson, AM; Michel, M; Oppenheimer, J; Stewardson, K; Jadhav, N; Kim, YJ; Chatterjee, M; Munshi, A; Panyam, A; Waghmare, P; Yadav, Y; Patel, H; Kaushik, A; Thangaraj, K; Meyer, M; Patterson, N; Rai, N; Reich, D (17 October 2019). "An Ancient Harappan Genome Lacks Ancestry from Steppe Pastoralists or Iranian Farmers" (PDF). Cell. 179 (3): 729–735.e10. doi:10.1016/j.cell.2019.08.048. PMC 6800651. PMID 31495572.
- ^ Bingusheva MA, Korshunova Ti, Kubora, Villems R, Kusutuntinova EK (2004). «[Филогеографический анализ митохондриальных ДНК -ногай: высокий уровень смеси материнских линий из восточной и западной Евразии]». Molecularnaia Biologia 38 (4): 617–2 PMID 15456133 .
- ^ Jump up to: а беременный Derbeneva OA, Staricovskaia EB, Volod'ko NV, Wallace DC, Sukernik Ri (ноябрь 2002 г.). «[Изменение митохондриальной ДНК в Кетах и Анганах и ранних народах Северной Евразии]» генетика 38 (11): 1554–6 PMID 1250682
- ^ Скотт Р. (2009). «Филогения MT-HG U2A» (PDF) .
- ^ Скотт Р. (2010). «Филогения MT-HG U2B» (PDF) .
- ^ Скотт Р. (2009). «Филогения MT-HG U2C» (PDF) .
- ^ Скотт Р. (2009). «Филогения MT-HG U2D» (PDF) .
- ^ Скотт Р. (2010). «Филогения MT-HG U2E» (PDF) .
- ^ Malyarchuk B, Derenko M, Perkova M, Vanecek T (октябрь 2008 г.). «Митохондриальная гаплогруппа U2D Филогения и распределение». Человеческая биология . 80 (5): 565–71. doi : 10.3378/1534-6617-80.5.565 . PMID 19341323 . S2CID 9447306 .
- ^ А. Сегин-Орландо; и др. (6 ноября 2014 г.). «Геномная структура у европейцев, начиная с не менее 36 200 лет» . Наука . 346 (6213): 1113–1118. Bibcode : 2014sci ... 346.1113S . doi : 10.1126/science.aaa0114 . PMID 25378462 . S2CID 206632421 .
- ^ «ДНК проанализирована с ранней европейской» . BBC News . 2010 год.
- ^ Ли Э.Дж., Макаревич С., Реннеберг Р., Хардер М., Краузе-Киора Б., Мюллер С., Остриц С., Ферен-Шмитз Л., Шрайбер С., Мюллер Дж., Фон Вурмб-Шквар Н., Небель А (август 2012 г.). «Новые генетические паттерны европейского неолита: перспективы от покойного неолитического места захоронения колокола в Германии» . Американский журнал физической антропологии . 148 (4): 571–9. doi : 10.1002/ajpa.22074 . PMID 22552938 .
- ^ Мелхиор Л., Гилберт М.Т., Кивизилд Т., Линнерп Н., Диссергинг Дж (февраль 2008 г.). «Редкие гаплогруппы мтДНК и генетические различия в богатых и бедных деревнях датских железных веков». Американский журнал физической антропологии . 135 (2): 206–15. doi : 10.1002/ajpa.20721 . PMID 18046774 .
- ^ Ким К., Бреннер Ч., Мейр В.Х., Ли К.Х., Ким Дж.Х., Гелегдорж Э. и др. (Июль 2010). «Западный евразийский мужчина встречается на 2000-летнем элите Сюнгу-кладбище на северо-восточной Монголии» (PDF) . Американский журнал физической антропологии . 142 (3): 429–40. doi : 10.1002/ajpa.21242 . PMID 20091844 .
- ^ Jump up to: а беременный Скотт Р. (2010). «Филогения MT-HG U3» (PDF) .
- ^ Derenko M, Malyarchuk B, Bahmanimehr A, Denisova G, Perkova M, Farjadian S, et al. (2013). «Полное разнообразие митохондриальной ДНК у иранцев» . Plos один . 8 (11): E80673. Bibcode : 2013ploso ... 880673d . doi : 10.1371/journal.pone.0080673 . PMC 3828245 . PMID 24244704 .
- ^ Jump up to: а беременный Ричардс М., Маколей В., Хикки Е., Вега Е., Сайкс Б., Гуида В. и др. (Ноябрь 2000). «Отслеживание европейских линий основателей в ближневосточном пуле мтДНК» (PDF) . Американский журнал человеческой генетики . 67 (5): 1251–76. doi : 10.1016/s0002-9297 (07) 62954-1 . PMC 1288566 . PMID 11032788 . Архивировано из оригинала (PDF) на 2009-03-04 . Получено 2007-03-19 .
- ^ Jump up to: а беременный Quintana-Murci L, Chaix R, Wells RS, Behar DM, Sayar H, Scozzari R, et al. (Май 2004 г.). «Где Запад встречается на востоке: сложный ландшафт мтДНК юго -западного и центрального азиатского коридора» . Американский журнал человеческой генетики . 74 (5): 827–45. doi : 10.1086/383236 . PMC 1181978 . PMID 15077202 .
- ^ Malyarchuk BA, Grzybowski T, Derenko MV, Czarny J, Miścicka-Sliwka D (март 2006 г.). «Митохондриальное разнообразие ДНК в польском роме». Анналы человеческой генетики . 70 (Pt 2): 195–206. doi : 10.1111/j.1529-8817.2005.00222.x . PMID 16626330 . S2CID 662278 .
- ^ Mendizabal I, Valente C, Gusmão A, Alves C, Gomes V, Goios A, et al. (Январь 2011). «Реконструкция индийского происхождения и рассеяния европейской цыгане: материнская генетическая перспектива» . Plos один . 6 (1): E15988. Bibcode : 2011ploso ... 615988m . doi : 10.1371/journal.pone.0015988 . PMC 3018485 . PMID 21264345 .
- ^ Kujanovaá M, Perseira L, Fernandes V, Pereira JB, Cerný V (октябрь 2009 г.). Западные пустыни Журнал физической анторологии 140 (2): 336–4 doi : 10.1002/ajp . PMID 19425100 .
- ^ Stevanovitch A, Gilles A, Bouzaid E, Kefi R, Paris F, Gayraud RP, et al. (Январь 2004 г.). «Разнообразие последовательности митохондриальных последовательности ДНК в сидячей популяции из Египта». Анналы человеческой генетики . 68 (Pt 1): 23–39. doi : 10.1046/j.1529-8817.2003.00057.x . PMID 14748828 . S2CID 44901197 .
- ^ Pereira L, Cerný V, Cerezo M, Silva NM, Hájek M, Vasíková A, et al. (Август 2010 г.). «Связывая генные бассейны к югу и западному евразии: материнское и отцовское наследие кочевников Туарега из африканского Сахеля» . Европейский журнал человеческой генетики . 18 (8): 915–23. doi : 10.1038/ejhg.2010.21 . PMC 2987384 . PMID 20234393 .
- ^ «Судебные и филогеографические характеристики линий мтДНК из Сомали» .
- ^ Jump up to: а беременный Саламон М., Цур С., Цур С., Аренсбург Б., Зиас Дж., Боаретто Е (январь 2010 г.). « PDF (PDF) человека» . Археология 10 (2): 1–4. Bibcode : 2010maa .... 10 ... 10 ... 1s Архивировано с 2016-03-03 .
- ^ Буджа М (декабрь 2013 г.). «Неолитические горшки и гончары в Европе: конец миграционной модели« Демической диффузии » . Documenta Praehistorica . 40 : 38–56. doi : 10.4312/dp.40.5 .
- ^ Der Sarkissian C, Balanovsky O, Brandt G, Khartanovich V, Buzhilova A, Koshel S, et al. (2013). «Древняя ДНК выявляет доисторический ген-поток из Сибири в сложной истории человеческой популяции северо-восточной Европы» . PLOS Genetics . 9 (2): E1003296. doi : 10.1371/journal.pgen.1003296 . PMC 3573127 . PMID 23459685 .
- ^ Haak W, Balanovsky O, Sanchez JJ, Koshel S, Zaporozhchenko V, Adler CJ, et al. (Ноябрь 2010). «Древняя ДНК от европейских ранних неолитических фермеров раскрывает их ближневосточную аффинность» . PLOS Биология . 8 (11): E1000536. doi : 10.1371/journal.pbio.1000536 . PMC 2976717 . PMID 21085689 .
- ^ Jump up to: а беременный в Derbeneva OA, Starikovskaya EB, Wallace DC, Sukernik Ri (апрель 2002 г.). «Следы ранних евразийцев в манси Северо -Западной Сибири выявлены с помощью анализа митохондриальной ДНК» . Американский журнал человеческой генетики . 70 (4): 1009–14. doi : 10.1086/339524 . PMC 379094 . PMID 11845409 .
- ^ Volodko NV, Starikovskaya EB, Mazunin IO, Eltsov NP, Naidenko PV, Wallace DC, et al. (Май 2008 г.). «Разнообразие митохондриального генома у арктических сибирцев, с особым ссылкой на эволюционную историю Берингии и плейстоценической народы Америки» . Американский журнал человеческой генетики . 82 (5): 1084–100. doi : 10.1016/j.ajhg.2008.03.019 . PMC 2427195 . PMID 18452887 .
- ^ «Клан Ульрике» . Материнское происхождение . Оксфордские предки. 2013. Архивировано из оригинала 2017-07-15 . Получено 2013-02-07 .
- ^ «Материнское происхождение» . Oxford Acestors Ltd. Архивировано с оригинала 2017-07-15 . Получено 2013-02-07 .
- ^ Башир М (2012). Полем Пакистан сегодня .
- ^ Логан I (2014). "U4A1 GenBank Peseecens" .
- ^ "Калаш" . Блог, посвященный гаплогруппе U4 .
- ^ Скотт Р. (2011). «Филогения MT-HG U4A» (PDF) .
- ^ Скотт Р. (2011). «Филогения MT-HG U4B» (PDF) .
- ^ Скотт Р. (2011). «Филогения MT-HG U4C» (PDF) .
- ^ Скотт Р. (2010). «Филогения MT-HG U4D» (PDF) .
- ^ «Доисторическая европейская ДНК -мтДНК и гаплогруппа частоты Y -ДНК по периоду» . Епипедия .
- ^ Jump up to: а беременный Мелхиор Л., Линнерп Н., Зигисминд Х.Р., Кивизилд Т., Диссергинг Дж (июль 2010 г.). «Генетическое разнообразие среди древних скандинавских популяций» . Plos один . 5 (7): E11898. Bibcode : 2010ploso ... 511898m . doi : 10.1371/journal.pone.0011898 . PMC 2912848 . PMID 20689597 .
- ^ Malyarchuk B, Grzybowski T, Derenko M, Perkova M, Vanecek T, Lazur J, Gomolcak P, Tsybovsky I (август 2008 г.). «Митохондриальная ДНК -филогения у восточных и западных славян» . Молекулярная биология и эволюция . 25 (8): 1651–8. doi : 10.1093/molbev/msn114 . PMID 18477584 .
- ^ Манко Дж. (2009). «Древняя западная Евразия ДНК медного и бронзового возраста» . Наследственные путешествия . Архивировано из оригинала 2017-01-22.
- ^ Скотт Р. (2010). «Комплексный список мутаций HG U» (PDF) .
- ^ «Славицизация уральских динамиков Volga-Oka Interfluve. Языки и гены от железного века до славян» . 22 января 2023 года . Получено 2023-08-31 -через YouTube .
- ^ PDF . Культурный музей . Университет Осло. [ Постоянная мертвая ссылка ]
- ^ Jump up to: а беременный «U7 Гаплогруппа Митохондриальная ДНК проект» . Архивировано с оригинала 2 июня 2014 года.
- ^ Jump up to: а беременный Rudbeck L, Gilbert MT, Willerslev E, Hansen AJ, Lynnerup N, Christensen T, Discing J (октябрь 2005 г.). «Анализ мтДНК человеческих останков с раннего датского христианского кладбища». Американский журнал физической антропологии . 128 (2): 424–9. doi : 10.1002/ajpa.20294 . PMID 15838837 .
- ^ Ranaweera L, Kaewsutthi S, Win Tun A, Boonyarit H, Poolsuwan S, Lertrit P (январь 2014 г.). «Митохондриальная ДНК История шри -ланкийских этнических людей: их отношения на острове и индийские субконтинентальные популяции» . Журнал человеческой генетики . 59 (1): 28–36. doi : 10.1038/jhg.2013.112 . PMID 24196378 . S2CID 41185629 .
- ^ Jump up to: а беременный Скотт Р. (2010). «Филогения MT-HG U7» (PDF) .
- ^ Ли Э.Дж., Штеффен С., Хардер М., Краузе-Киора Б., фон Вурмб-Шшварк Н., Небель А (январь 2012 г.). «Древняя перспектива ДНК на железный век" княжеских захоронения "из Баден-Вииртберга, Германия» . Американский журнал физической антропологии . 147 : 190. doi : 10.1002/ajpa.22033 .
- ^ Берглунд N (26 марта 2007 г.). «Женщина -викинги имела корни возле Черного моря» . Aftenposten - Новости на английском языке . Aftenposten.no. Архивировано из оригинала 2010-08-26 . Получено 11 апреля 2010 года .
- ^ Фаррелл Дж.Дж., Аль-Али А.К., Фаррер Л.А., Аль-Нафай А.Н., Аль-Рубайш А.М., Мелиста Е. и др. «Геном Саудовской Аравии выявляет двухэтапный миграцию вне Африки» . Американское общество человеческой генетики (Ашг). Архивировано с оригинала 2016-09-13 . Получено 2016-05-12 .
- ^ Kivisild T, Reidla M, Metspalu E, Rosa A, Break and Pennarun E, et al. (Ноябрь 2004 г.). «Эфиопская митохондриальная ДНК наследие: отслеживание гена течет ворота слез » Американский журнал человеческой генетики 75 (5): 752–7 Doi : 10.1086/ 4 1182106PMC 15457403PMID
- ^ Найти ваши корни . Сезон 1. Эпизод 8. 2012-05-06. Пбс.
- ^ Надин Эпштейн (сентябрь -октябрь 2012 года). «Moment Magazine Magazine Great DNA Experiment» . Moment Magazine . п. 45 Получено 2024-03-07 .
- ^ Мур, Cece (2010-12-31). «Моя полная митохондриальная последовательность, гетероплазмия и сравнение обозначений мтДНК на ftdna и 23andme» . Ваш генетический генеалог .
Дальнейшее чтение
[ редактировать ]- Бехар Д.М., Ван Очев М., Россет С., Метпалу М., Лугвели Э.Л., Сильва Н.М., Кивизилд Т., Торрони А., Виллемы Р. (апрель 2012 г.). «Коперник» переоценка человеческого дерева митохондриальной ДНК из его корня » . Американский журнал человеческой генетики . 90 (4): 675–84. doi : 10.1016/j.ajhg.2012.03.002 . PMC 3322232 . PMID 22482806 .
Внешние ссылки
[ редактировать ]- Общий
- Яна Логана Сайт митохондриальной ДНК
- духовки Манни из филотрии
- Гаплогруппа u
- Yfull Mtree's Aplogroup u
- Митомап гаплогруппа u
- Распространение гаплогруппы U , из National Geographic
- Проект мтДНК U1 в ДНК семейного дерева
- U3 Mtdna Haplogroup Project в ДНК семейного дерева
- U4 Mtdna Haplogroup Project в ДНК семейного дерева
- Проект U5 мтДНК в ДНК семейного дерева
- Проект U6 мтДНК в ДНК семейного дерева
- U7 MtDna Гаплогруппа в ДНК семейного древа
- U8 Mtdna Гаплогруппа в ДНК семейного древа
- Проект U9 мтДНК в ДНК семейного дерева