Аллостерическая регуляция

В области биохимии и фармакологии аллостерический регулятор (или аллостерический модулятор ) — это вещество, которое связывается с участком фермента или рецептора, отличным от активного центра , что приводит к конформационным изменениям, изменяющим активность белка, либо усиливая, либо ингибируя его. функция. Напротив, вещества, которые связываются непосредственно с активным центром фермента или местом связывания эндогенного лиганда рецептора, называются ортостерическими регуляторами или модуляторами.
Сайт, с которым связывается эффектор, называется аллостерическим сайтом или регуляторным сайтом . Аллостерические сайты позволяют эффекторам связываться с белком, что часто приводит к конформационным изменениям и/или изменению динамики белка . [ 1 ] [ 2 ] Эффекторы, усиливающие активность белка, называются аллостерическими активаторами , тогда как те, которые снижают активность белка, называются аллостерическими ингибиторами .
Аллостерические регуляции являются естественным примером контуров управления, таких как обратная связь от последующих продуктов или прямая связь от вышестоящих субстратов. Аллостерия дальнего действия особенно важна для передачи сигналов в клетках . [ 3 ] Аллостерическая регуляция также особенно важна для способности клетки регулировать активность ферментов .
Термин аллостерия происходит от древнегреческого allos ( ἄλλος ), «другой», и стерео ( στερεός ), «твердое тело (объект)». Это связано с тем фактом, что регуляторный сайт аллостерического белка физически отличается от его активного сайта. Аллостерия контрастирует с презентацией субстрата , которая не требует конформационных изменений для активации фермента. Термин ортостерия происходит от древнегреческого orthós ( ὀρθός ), что означает «прямой», «вертикальный», «правильный» или «правильный».
Орто против аллостерических ингибиторов
[ редактировать ]ортостерический
[ редактировать ]- Сайт связывания: Ортостерические ингибиторы связываются непосредственно с активным центром фермента, где обычно связывается субстрат.
- Механизм действия: Занимая активный центр, эти ингибиторы предотвращают связывание субстрата, тем самым напрямую блокируя каталитическую активность фермента.
- Конкурентное ингибирование. Большинство ортостерических ингибиторов конкурируют с субстратом за активный центр, что означает, что их эффективность может снизиться, если концентрация субстрата увеличится.
Аллостерический
[ редактировать ]- Сайт связывания: аллостерические ингибиторы связываются с участком фермента, который отличается и отделен от активного сайта, известного как аллостерический сайт.
- Механизм действия: Связывание с аллостерическим сайтом вызывает конформационные изменения фермента, которые могут либо снизить сродство активного центра к субстрату, либо изменить каталитическую активность фермента. Это косвенное вмешательство может ингибировать функцию фермента, даже если субстрат присутствует.
- Неконкурентное ингибирование. Аллостерические ингибиторы часто проявляют неконкурентное ингибирование, то есть их ингибирующий эффект не зависит от концентрации субстрата.
Модели
[ редактировать ]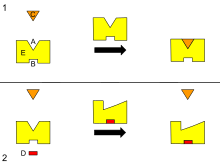
Б – Аллостерический сайт
С – Субстрат
Д – Ингибитор
Е – Фермент
Это схема аллостерической регуляции фермента.
Многие аллостерические эффекты могут быть объяснены согласованной моделью MWC, предложенной Моно , Вайманом и Чанже . [ 4 ] или последовательной моделью (также известной как модель KNF), описанной Кошландом , Немети и Филмером. [ 5 ] Оба постулируют, что субъединицы белка существуют в одной из двух конформаций : напряженной (T) или расслабленной (R), и что расслабленные субъединицы легче связывают субстрат, чем те, которые находятся в напряженном состоянии. Две модели больше всего различаются в своих предположениях о взаимодействии субъединиц и предсуществовании обоих состояний. Для белков, в которых субъединицы существуют более чем в двух конформациях , модель аллостерического ландшафта, описанная Куэндетом, Вайнштейном и Левином, [ 6 ] можно использовать. Аллостерической регуляции может способствовать эволюция крупномасштабных низкоэнергетических конформационных изменений, которые обеспечивают аллостерическое взаимодействие на большие расстояния между удаленными сайтами связывания. [ 7 ]
Согласованная модель
[ редактировать ]Согласованная модель аллостерии, также называемая моделью симметрии или моделью MWC , постулирует, что субъединицы фермента связаны таким образом, что конформационное изменение в одной субъединице обязательно передается всем другим субъединицам. Таким образом, все субъединицы должны существовать в одной и той же конформации. Модель также утверждает, что в отсутствие какого-либо лиганда (субстрата или иного) равновесие благоприятствует одному из конформационных состояний, T или R. Равновесие может быть сдвинуто в состояние R или T за счет связывания одного лиганда ( аллостерический эффектор или лиганд) к сайту, отличному от активного сайта
Последовательная модель
[ редактировать ]Последовательная модель аллостерической регуляции утверждает, что субъединицы не связаны таким образом, что конформационные изменения в одной из них вызывают аналогичные изменения в других. Таким образом, все субъединицы фермента не обязательно имеют одинаковую конформацию. Более того, последовательная модель требует, чтобы молекулы субстрата связывались посредством протокола индуцированного соответствия . Хотя такое индуцированное соответствие переводит субъединицу из напряженного состояния в расслабленное состояние, оно не распространяет конформационные изменения на соседние субъединицы. Вместо этого связывание субстрата с одной субъединицей лишь незначительно изменяет структуру других субъединиц, так что их сайты связывания становятся более восприимчивыми к субстрату. Подводя итог:
- субъединицы не обязательно должны существовать в одной и той же конформации
- молекулы субстрата связываются по протоколу индуцированной подгонки
- конформационные изменения не распространяются на все субъединицы
Модель Морфеи
[ редактировать ]Морфеиновая . модель аллостерической регуляции представляет собой диссоциативную согласованную модель [ 8 ]
Морфеин представляет собой гомоолигомерную структуру, которая может существовать как ансамбль физиологически значимых и функционально различных альтернативных четвертичных ансамблей. Переходы между альтернативными сборками морфеина включают диссоциацию олигомеров, конформационные изменения в диссоциированном состоянии и повторную сборку в другой олигомер. Необходимый этап разборки олигомера отличает морфеиновую модель аллостерической регуляции от классических моделей MWC и KNF.
Порфобилиногенсинтаза (PBGS) является прототипом морфеина.
Модели ансамбля
[ редактировать ]аллостерической системы Ансамблевые модели аллостерической регуляции перечисляют статистический ансамбль как функцию ее функции потенциальной энергии , а затем связывают конкретные статистические измерения аллостерии с конкретными энергетическими членами энергетической функции (например, межмолекулярным солевым мостиком между двумя доменами). [ 9 ] Ансамблевые модели, такие как аллостерическая модель ансамбля. [ 10 ] и аллостерическая модель Изинга [ 11 ] предположим, что каждая область системы может принимать два состояния, аналогично модели MWC. Модель аллостерического ландшафта, предложенная Кюэнде, Вайнштейном и ЛеВином. [ 6 ] позволяет доменам иметь любое количество состояний, а вклад конкретного молекулярного взаимодействия в данную аллостерическую связь можно оценить с помощью строгого набора правил. Моделирование молекулярной динамики можно использовать для оценки статистического ансамбля системы, чтобы его можно было проанализировать с помощью модели аллостерического ландшафта.
Аллостерическая модуляция
[ редактировать ]Аллостерическая модуляция используется для изменения активности молекул и ферментов в биохимии и фармакологии. Для сравнения: обычное лекарство связывается с активным центром фермента, что, таким образом, запрещает связывание субстрата с этим ферментом, вызывая снижение активности фермента. Аллостерическая модуляция происходит, когда эффектор связывается с аллостерическим сайтом (также известным как регуляторный сайт) фермента и изменяет активность фермента. Аллостерические модуляторы разработаны таким образом, чтобы соответствовать аллостерическому сайту и вызывать конформационные изменения фермента, в частности, изменение формы активного сайта, что затем вызывает изменение его активности. В отличие от типичных лекарств модуляторы не являются конкурентными ингибиторами . Они могут быть положительными (активирующими), вызывающими повышение активности фермента, или отрицательными (ингибирующими), вызывающими снижение активности фермента. Использование аллостерической модуляции позволяет контролировать эффекты активности специфических ферментов; в результате аллостерические модуляторы очень эффективны в фармакологии. [ 12 ] В биологической системе аллостерическую модуляцию бывает трудно отличить от модуляции по представлению субстрата .
Модель измерения энергии
[ редактировать ]Примером этой модели является микобактерия туберкулеза , бактерия , которая идеально приспособлена к жизни в макрофагах человека. Сайты фермента служат связующим звеном между различными субстратами. В частности, между AMP и G6P . Подобные сайты также служат механизмом определения производительности фермента. [ 13 ]
Положительная модуляция
[ редактировать ]Положительная аллостерическая модуляция (также известная как аллостерическая активация ) происходит, когда связывание одного лиганда усиливает притяжение между молекулами субстрата и другими сайтами связывания. Примером является связывание молекул кислорода с гемоглобином , где кислород эффективно является одновременно субстратом и эффектором. Аллостерический, или «другой», сайт — это активный сайт примыкающей белковой субъединицы . Связывание кислорода с одной субъединицей вызывает конформационные изменения в этой субъединице, которые взаимодействуют с остальными активными центрами, повышая их сродство к кислороду. Другой пример аллостерической активации наблюдается в цитозольной IMP-GMP-специфичной 5'-нуклеотидазе II (cN-II), где сродство к субстрату GMP увеличивается при связывании GTP на границе раздела димеров.
Отрицательная модуляция
[ редактировать ]Отрицательная аллостерическая модуляция (также известная как аллостерическое ингибирование ) происходит, когда связывание одного лиганда снижает сродство к субстрату в других активных центрах. Например, когда 2,3-БФГ связывается с аллостерическим участком гемоглобина, сродство к кислороду всех субъединиц снижается. Это когда регулятор отсутствует в месте связывания.
Прямые ингибиторы тромбина представляют собой прекрасный пример отрицательной аллостерической модуляции. аллостерические ингибиторы Обнаружены тромбина, которые потенциально могут использоваться в качестве антикоагулянтов.
Другой пример — стрихнин , судорожный яд, который действует как аллостерический ингибитор рецептора глицина . Глицин является основным постсинаптическим тормозным нейромедиатором в спинном млекопитающих мозге и стволе головного мозга . Стрихнин действует на отдельный сайт связывания на глициновом рецепторе аллостерическим образом; т.е. его связывание снижает сродство глицинового рецептора к глицину. Таким образом, стрихнин подавляет действие тормозного медиатора, приводящее к судорогам.
Другой случай, когда можно увидеть отрицательную аллостерическую модуляцию, - это между АТФ и ферментом фосфофруктокиназой в петле отрицательной обратной связи , которая регулирует гликолиз . Фосфофруктокиназа (обычно называемая ПФК ) представляет собой фермент, который катализирует третий этап гликолиза: фосфорилирование фруктозо -6-фосфата во фруктозо-1,6-бисфосфат . ПФК может аллостерически ингибироваться высокими уровнями АТФ внутри клетки. Когда уровни АТФ высоки, АТФ связывается с аллостерическим участком фосфофруктокиназы , вызывая изменение трехмерной формы фермента. Это изменение приводит к снижению его сродства к субстрату ( фруктозо-6-фосфату и АТФ ) в активном центре, и фермент считается неактивным. Это приводит к остановке гликолиза в организме при высоких уровнях АТФ, сохраняя таким образом глюкозу и поддерживая сбалансированный уровень клеточного АТФ. Таким образом, АТФ служит отрицательным аллостерическим модулятором ПФК, несмотря на то, что он также является субстратом фермента.
Типы
[ редактировать ]Гомотропный
[ редактировать ]Гомотропный аллостерический модулятор является субстратом для своего белка- мишени , а также регуляторной молекулой активности белка. Обычно это активатор белка. [ 14 ] Например, О 2 и СО являются гомотропными аллостерическими модуляторами гемоглобина. Аналогично, в 5'-нуклеотидазе, специфичной для IMP/GMP, связывание одной молекулы GMP с одной субъединицей тетрамерного фермента приводит к повышенному сродству к GMP со стороны последующих субъединиц, как показывают сигмоидальные графики зависимости субстрата от скорости. [ 14 ]
Гетеротропный
[ редактировать ]Гетеротропный аллостерический модулятор представляет собой регуляторную молекулу, не являющуюся субстратом фермента. Это может быть либо активатор, либо ингибитор фермента. Например, Х + , CO 2 и 2,3-бисфосфоглицерат являются гетеротропными аллостерическими модуляторами гемоглобина. [ 15 ] И снова, в IMP/GMP-специфической 5'-нуклеотидазе связывание молекулы GTP на границе раздела димеров в тетрамерном ферменте приводит к повышенному сродству к субстрату GMP в активном центре, что указывает на гетеротропную аллостерическую активацию K-типа. [ 14 ]
Как было подробно подчеркнуто выше, некоторые аллостерические белки могут регулироваться как их субстратами, так и другими молекулами. Такие белки способны как к гомотропным, так и к гетеротропным взаимодействиям. [ 14 ]
Основные активаторы
[ редактировать ]Некоторые аллостерические активаторы называются «незаменимыми» или «облигатными» активаторами в том смысле, что в их отсутствие активность их целевого фермента очень низкая или незначительная, как в случае с активностью N-ацетилглутамата в отношении карбамоилфосфата. синтетаза I, например. [ 16 ] [ 17 ]
Нерегуляторная аллостерия
[ редактировать ]Нерегуляторный аллостерический сайт — это любой нерегуляторный компонент фермента (или любого белка), который сам по себе не является аминокислотой. Например, многим ферментам для обеспечения правильного функционирования требуется связывание натрия. Однако натрий не обязательно действует как регуляторная субъединица; натрий присутствует всегда, и не существует известных биологических процессов добавления/удаления натрия для регулирования активности ферментов. Нерегуляторная аллостерия может включать любые другие ионы, кроме натрия (кальция, магния, цинка), а также другие химические вещества и, возможно, витамины.
Фармакология
[ редактировать ]Аллостерическая модуляция рецептора возникает в результате связывания аллостерических модуляторов в другом сайте (« регуляторный сайт »), отличном от сайта эндогенного лиганда (« активный сайт »), и усиливает или ингибирует эффекты эндогенного лиганда. В нормальных условиях он действует, вызывая конформационные изменения в молекуле рецептора, что приводит к изменению аффинности связывания лиганда. Таким образом, аллостерический лиганд модулирует активацию рецептора своим первичным ортостерическим лигандом, и можно считать, что он действует как регулятор яркости в электрической цепи, регулируя интенсивность ответа.
Например, ГАМК А рецептор имеет два активных сайта, с которыми связывается нейротрансмиттер гамма-аминомасляная кислота (ГАМК), но также имеет бензодиазепина и общего анестетика сайты регуляторного связывания . Каждый из этих регуляторных сайтов может вызывать положительную аллостерическую модуляцию, усиливая активность ГАМК. Диазепам является положительным аллостерическим модулятором регуляторного участка бензодиазепина, а его антидот флумазенил является антагонистом рецептора .
Более поздние примеры препаратов, которые аллостерически модулируют свои цели, включают цинакальцет для лечения ВИЧ , имитирующий кальций, и маравирок .
Аллостерические сайты как мишени для лекарств
[ редактировать ]Аллостерические белки участвуют и играют центральную роль во многих заболеваниях. [ 18 ] [ 19 ] и аллостерические сайты могут представлять собой новую мишень для лекарств . Использование аллостерических модуляторов в качестве предпочтительных терапевтических средств имеет ряд преимуществ по сравнению с классическими ортостерическими лигандами. Например, аллостерические сайты связывания рецептора, связанного с G-белком (GPCR), не подвергались такому же эволюционному давлению, как ортостерические сайты, для размещения эндогенного лиганда, поэтому они более разнообразны. [ 20 ] Следовательно, большая селективность GPCR может быть достигнута путем нацеливания на аллостерические сайты. [ 20 ] Это особенно полезно для GPCR, где селективная ортостерическая терапия затруднена из-за консервативности последовательности ортостерического сайта среди подтипов рецепторов. [ 21 ] Кроме того, эти модуляторы имеют пониженный потенциал токсического действия, поскольку модуляторы с ограниченной кооперативностью будут иметь максимальный уровень своего эффекта, независимо от введенной дозы. [ 20 ] Другой тип фармакологической селективности, уникальный для аллостерических модуляторов, основан на кооперативности. Аллостерический модулятор может проявлять нейтральную кооперативность с ортостерическим лигандом для всех подтипов данного рецептора, кроме интересующего подтипа, который называется «абсолютной селективностью подтипа». [ 21 ] Если аллостерический модулятор не обладает заметной эффективностью, он может обеспечить еще одно мощное терапевтическое преимущество перед ортостерическими лигандами, а именно способность избирательно повышать или понижать тканевые реакции только при наличии эндогенного агониста. [ 21 ] Олигомер-специфичные сайты связывания малых молекул являются мишенями для медицински значимых морфеинов . [ 22 ]
Синтетические аллостерические системы
[ редактировать ]Существует множество синтетических соединений, содержащих несколько сайтов нековалентного связывания, конформационные изменения которых происходят при занятии одного сайта. Кооперативность между отдельными вкладами связывания в таких супрамолекулярных системах положительна, если занятие одного сайта связывания увеличивает сродство ΔG ко второму сайту, и отрицательна, если сродство не повышается. Большинство синтетических аллостерических комплексов основаны на конформационной реорганизации при связывании одного эффекторного лиганда, что затем приводит либо к усилению, либо к ослаблению ассоциации второго лиганда в другом сайте связывания. [ 23 ] [ 24 ] [ 25 ] Конформационная связь между несколькими сайтами связывания в искусственных системах обычно намного больше, чем в белках с их обычно большей гибкостью. Параметром, определяющим эффективность (измеряемым соотношением констант равновесия Krel = KA(E)/KA в присутствии и в отсутствие эффектора E), является конформационная энергия, необходимая для принятия закрытой или напряженной конформации для связывания лиганда. А. [ 26 ]
Во многих мультивалентных супрамолекулярных системах [ 27 ] может произойти прямое взаимодействие между связанными лигандами, что может привести к большим кооперативам. Наиболее распространенным является такое прямое взаимодействие ионов в рецепторах для ионных пар. [ 28 ] [ 29 ] Эту кооперативность часто также называют аллостерией, хотя конформационные изменения здесь не обязательно запускают события связывания.
Интернет-ресурсы
[ редактировать ]Аллостерическая база данных
[ редактировать ]Аллостерия — это прямой и эффективный способ регуляции биологической функции макромолекулы, вызываемый связыванием лиганда в аллостерическом сайте, топографически отличном от ортостерического сайта. Ожидается, что из-за часто высокой селективности рецепторов и более низкой целевой токсичности аллостерическая регуляция будет играть все большую роль в открытии лекарств и биоинженерии. Аллостерическая база данных (ASD) [ 30 ] предоставляет центральный ресурс для отображения, поиска и анализа структуры, функций и связанных с ними аннотаций для аллостерических молекул. В настоящее время РАС содержит аллостерические белки более чем 100 видов и модуляторы трех категорий (активаторы, ингибиторы и регуляторы). Каждый белок снабжен подробным описанием аллостерии, биологического процесса и связанных с ним заболеваний, а каждый модулятор - аффинностью связывания, физико-химическими свойствами и терапевтической областью. Интеграция информации об аллостерических белках при РАС должна позволить прогнозировать аллостерию неизвестных белков с последующей экспериментальной проверкой. Кроме того, модуляторы, курируемые в ASD, могут использоваться для исследования потенциальных аллостерических мишеней для исследуемого соединения и могут помочь химикам реализовать модификации структуры для разработки новых аллостерических лекарств.
Аллостерические остатки и их предсказание
[ редактировать ]Не все белковые остатки играют одинаково важную роль в аллостерической регуляции. Идентификация остатков, которые необходимы для аллостерии (так называемых «аллостерических остатков»), была в центре внимания многих исследований, особенно в последнее десятилетие. [ 31 ] [ 32 ] [ 33 ] [ 34 ] [ 35 ] [ 36 ] [ 37 ] [ 38 ] Частично этот растущий интерес является результатом их общей важности в науке о белках, а также потому, что аллостерические остатки могут использоваться в биомедицинском контексте . Фармакологически важные белки с труднодоступными сайтами могут уступить место подходам, в которых альтернативно нацеливаются на более легкодоступные остатки, которые способны аллостерически регулировать первичный интересующий сайт. [ 39 ] Эти остатки можно в широком смысле классифицировать как поверхностные и внутренние аллостерические аминокислоты. Аллостерические сайты на поверхности обычно играют регуляторную роль, фундаментально отличную от той, что находится внутри; поверхностные остатки могут служить рецепторами или эффекторными участками при аллостерической передаче сигналов, тогда как остатки внутри могут передавать такие сигналы. [ 40 ] [ 41 ]
См. также
[ редактировать ]- база данных АСД
- Ангармонизм
- Конкурентное ингибирование
- Кооперативная привязка
- Кинетика ферментов
- Динамика белка
- Теория рецепторов
Ссылки
[ редактировать ]- ^ Купер А., Драйден Д.Т. (октябрь 1984 г.). «Аллостерия без конформационных изменений. Правдоподобная модель». Европейский биофизический журнал . 11 (2): 103–109. дои : 10.1007/BF00276625 . ПМИД 6544679 . S2CID 12591175 .
- ^ Лю Дж., Нусинов Р. (июнь 2016 г.). «Аллостерия: обзор ее истории, концепций, методов и приложений» . PLOS Вычислительная биология . 12 (6): e1004966. Бибкод : 2016PLSCB..12E4966L . дои : 10.1371/journal.pcbi.1004966 . ПМЦ 4890769 . ПМИД 27253437 . S2CID 3610740 .
- ^ Бу Зи, Callaway DJ (2011). «Белки движутся! Динамика белков и дальняя аллостерия в передаче сигналов в клетках». Структура белка и болезни . Достижения в области химии белков и структурной биологии. Том. 83. стр. 163–221. дои : 10.1016/B978-0-12-381262-9.00005-7 . ISBN 9780123812629 . ПМИД 21570668 .
- ^ Моно Дж., Вайман Дж., Чанже Дж. П. (май 1965 г.). «О природе аллостерических переходов: правдоподобная модель». Журнал молекулярной биологии . 12 : 88–118. дои : 10.1016/s0022-2836(65)80285-6 . ПМИД 14343300 .
- ^ Кошланд Д.Е., Немети Дж., Филмер Д. (январь 1966 г.). «Сравнение экспериментальных данных связывания и теоретических моделей в белках, содержащих субъединицы». Биохимия . 5 (1): 365–85. дои : 10.1021/bi00865a047 . ПМИД 5938952 .
- ^ Перейти обратно: а б Куендет М.А., Вайнштейн Х., Левин М.В. (декабрь 2016 г.). «Аллостерический ландшафт: количественная оценка термодинамических связей в биомолекулярных системах» . Журнал химической теории и вычислений . 12 (12): 5758–5767. дои : 10.1021/acs.jctc.6b00841 . ПМК 5156960 . ПМИД 27766843 .
- ^ Экманн Дж. П., Ружмон Дж., Тлусти Т. (30 июля 2019 г.). «Коллоквиум: Белки: физика аморфной развивающейся материи» . Обзоры современной физики . 91 (3): 031001. arXiv : 1907.13371 . Бибкод : 2019RvMP...91c1001E . дои : 10.1103/RevModPhys.91.031001 . ISSN 0034-6861 . S2CID 199001124 .
- ^ Яффе ЭК (сентябрь 2005 г.). «Морфеины - новая структурная парадигма аллостерической регуляции». Тенденции биохимических наук . 30 (9): 490–7. дои : 10.1016/j.tibs.2005.07.003 . ПМИД 16023348 .
- ^ Мотлах Х.Н., Врабль Дж.О., Ли Дж., Хильзер В.Дж. (апрель 2014 г.). «Ансамблевая природа аллостерии» . Природа . 508 (7496): 331–9. Бибкод : 2014Natur.508..331M . дои : 10.1038/nature13001 . ПМК 4224315 . ПМИД 24740064 .
- ^ Хильзер В.Дж., Врабль Дж.О., Мотлаг Х.Н. (2012). «Структурно-энергетическая основа аллостерии» . Ежегодный обзор биофизики . 41 : 585–609. doi : 10.1146/annurev-biophys-050511-102319 . ПМЦ 3935618 . ПМИД 22577828 .
- ^ ЛеВайн М.В., Вайнштейн Х. (май 2015 г.). «AIM для аллостерии: использование модели Изинга для понимания обработки и передачи информации в аллостерических биомолекулярных системах» . Энтропия . 17 (5): 2895–2918. Бибкод : 2015Entrp..17.2895L . дои : 10.3390/e17052895 . ПМЦ 4652859 . ПМИД 26594108 .
- ^ Абдель-Магид А.Ф. (февраль 2015 г.). «Аллостерические модуляторы: новая концепция открытия лекарств» . Письма ACS по медицинской химии . 6 (2): 104–7. дои : 10.1021/ml5005365 . ПМЦ 4329591 . ПМИД 25699154 .
- ^ Чжун В., Цуй Л., Го Б.С., Цай К., Хо П., Чион Ю.Х. и др. (декабрь 2017 г.). «Логические ворота на основе аллостерической пируваткиназы синергетически определяют уровень энергии и сахара в микобактериях туберкулеза» . Природные коммуникации . 8 (1): 1986. Бибкод : 2017NatCo...8.1986Z . дои : 10.1038/s41467-017-02086-y . ПМЦ 5719368 . ПМИД 29215013 .
- ^ Перейти обратно: а б с д Сринивасан Б., Форухар Ф., Шукла А., Сампанги С., Кулкарни С., Абашидзе М. и др. (март 2014 г.). «Аллостерическая регуляция и активация субстрата цитозольной нуклеотидазы II Legionella pneumophila» . Журнал ФЭБС . 281 (6): 1613–1628. дои : 10.1111/февраль 12727 . ПМЦ 3982195 . ПМИД 24456211 .
- ^ Эдельштейн С.Дж. (1975). «Кооперативные взаимодействия гемоглобина». Ежегодный обзор биохимии . 44 : 209–32. дои : 10.1146/annurev.bi.44.070175.001233 . ПМИД 237460 .
- ^ Ши Д., Аллевелл Н.М., Тачман М. (июнь 2015 г.). «Семейство N-ацетилглутаматсинтазы: структуры, функции и механизмы» . Международный журнал молекулярных наук . 16 (6): 13004–22. дои : 10.3390/ijms160613004 . ПМЦ 4490483 . ПМИД 26068232 .
- ^ де Сима С., Поло Л.М., Диес-Фернандес С., Мартинес А.И., Сервера Х., Фита И. и др. (ноябрь 2015 г.). «Структура карбамоилфосфатсинтетазы человека: расшифровка включения/выключения уреагенеза человека» . Научные отчеты . 5 (1): 16950. Бибкод : 2015NatSR...516950D . дои : 10.1038/srep16950 . ПМЦ 4655335 . ПМИД 26592762 .
- ^ Нусинов Р., Цай С. (2013). «Аллостерия в болезнях и в открытии лекарств» . Клетка . 153 (2): 293–305. дои : 10.1016/j.cell.2013.03.034 . ПМИД 23582321 .
- ^ Абрусан Г., Ашер Д.Б., Иноуе М. (2022). «Известные аллостерические белки играют центральную роль в генетических заболеваниях» . PLOS Вычислительная биология . 18 (2): e1009806. arXiv : 2107.04318 . Бибкод : 2022PLSCB..18E9806A . дои : 10.1371/journal.pcbi.1009806 . ПМИД 10138267 .
- ^ Перейти обратно: а б с Христопулос А., Мэй Л.Т., Авлани В.А., Секстон П.М. (ноябрь 2004 г.). «Аллостеризм рецепторов, связанных с G-белком: обещания и проблемы». Труды Биохимического общества . 32 (Часть 5): 873–7. дои : 10.1042/BST0320873 . ПМИД 15494038 .
- ^ Перейти обратно: а б с Мэй Л.Т., Лич К., Секстон П.М., Христопулос А. (2007). «Аллостерическая модуляция рецепторов, связанных с G-белком». Ежегодный обзор фармакологии и токсикологии . 47 : 1–51. doi : 10.1146/annurev.pharmtox.47.120505.105159 . ПМИД 17009927 .
- ^ Яффе ЭК (2010). «Морфеины – новый путь к открытию аллостерических лекарств~!12 февраля 2010 г.~!21 мая 2010 г.~!08 июня 2010 г.~!» . Журнал материалов открытой конференции . 1 : 1–6. doi : 10.2174/2210289201001010001 (неактивен 11 марта 2024 г.). ПМК 3107518 . ПМИД 21643557 .
{{cite journal}}: CS1 maint: DOI неактивен по состоянию на март 2024 г. ( ссылка ) - ^ Такеучи М., Икеда М., Сугасаки А., Синкай С. (ноябрь 2001 г.). «Молекулярный дизайн искусственных систем распознавания молекул и ионов с аллостерическими ответами гостя». Отчеты о химических исследованиях . 34 (11): 865–73. дои : 10.1021/ar0000410 . ПМИД 11714258 .
- ^ Кремер С., Лютцен А. (май 2013 г.). «Искусственные аллостерические рецепторы». Химия: Европейский журнал . 19 (20): 6162–96. дои : 10.1002/chem.201203814 . ПМИД 23463705 .
- ^ Ковбасюк Л., Кремер Р. (июнь 2004 г.). «Аллостерические супрамолекулярные рецепторы и катализаторы». Химические обзоры . 104 (6): 3161–87. дои : 10.1021/cr030673a . ПМИД 15186190 .
- ^ Шнайдер HJ (сентябрь 2016 г.). «Параметры эффективности искусственных аллостерических систем». Органическая и биомолекулярная химия . 14 (34): 7994–8001. дои : 10.1039/c6ob01303a . ПМИД 27431438 .
- ^ Баджич Дж.Д., Нельсон А., Кантрилл С.Дж., Тернбулл В.Б., Стоддарт Дж.Ф. (сентябрь 2005 г.). «Мультивалентность и кооперативность в супрамолекулярной химии». Отчеты о химических исследованиях . 38 (9): 723–32. дои : 10.1021/ar040223k . ПМИД 16171315 .
- ^ Ким С.К., Сесслер Дж.Л. (октябрь 2010 г.). «Рецепторы ионных пар» . Обзоры химического общества . 39 (10): 3784–809. дои : 10.1039/c002694h . ПМК 3016456 . ПМИД 20737073 .
- ^ МакКоннелл А.Дж., Бир PD (май 2012 г.). «Гетеродитопические рецепторы для распознавания ионных пар». Ангеванде Хеми . 51 (21): 5052–61. дои : 10.1002/anie.201107244 . ПМИД 22419667 .
- ^ Хуан З, Чжу Л, Цао Ю, Ву Г, Лю С, Чэнь Ю и др. (январь 2011 г.). «ASD: обширная база данных аллостерических белков и модуляторов» . Исследования нуклеиновых кислот . 39 (Проблема с базой данных): D663–9. дои : 10.1093/нар/gkq1022 . ПМК 3013650 . ПМИД 21051350 .
- ^ Панькович А, Даура X (октябрь 2012 г.). «Использование гибкости белка для прогнозирования местоположения аллостерических сайтов» . БМК Биоинформатика . 13 : 273. дои : 10.1186/1471-2105-13-273 . ПМК 3562710 . ПМИД 23095452 .
- ^ Суэль GM, Lockless SW, Wall MA, Ranganathan R (январь 2003 г.). «Эволюционно консервативные сети остатков опосредуют аллостерическую связь в белках». Структурная биология природы . 10 (1): 59–69. дои : 10.1038/nsb881 . ПМИД 12483203 . S2CID 67749580 .
- ^ Миттернахт С., Березовский И.Н. (сентябрь 2011 г.). «Связывающий рычаг как молекулярная основа аллостерической регуляции» . PLOS Вычислительная биология . 7 (9): e1002148. Бибкод : 2011PLSCB...7E2148M . дои : 10.1371/journal.pcbi.1002148 . ПМК 3174156 . ПМИД 21935347 .
- ^ Гаспер П.М., Фуглестад Б., Комивес Е.А., Марквик П.Р., Маккаммон Дж.А. (декабрь 2012 г.). «Аллостерические сети тромбина различают прокоагулянтную и антикоагулянтную активность» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (52): 21216–22. дои : 10.1073/pnas.1218414109 . ПМЦ 3535651 . ПМИД 23197839 .
- ^ Гош А., Вишвешвара С. (ноябрь 2008 г.). «Вариации в моделях клик и сообществ в белковых структурах во время аллостерической коммуникации: исследование динамически уравновешенных структур комплексов метионил-тРНК-синтетазы». Биохимия . 47 (44): 11398–407. дои : 10.1021/bi8007559 . ПМИД 18842003 .
- ^ Сети А., Эргл Дж., Блэк А.А., Люти-Шультен З. (апрель 2009 г.). «Динамические сети в комплексах тРНК:белок» . Труды Национальной академии наук Соединенных Штатов Америки . 106 (16): 6620–5. Бибкод : 2009PNAS..106.6620S . дои : 10.1073/pnas.0810961106 . ПМЦ 2672494 . ПМИД 19351898 .
- ^ Ванварт А.Т., Эргл Дж., Люти-Шультен З., Амаро Р.Э. (август 2012 г.). «Изучение вклада остаточных компонентов в динамические сетевые модели аллостерии» . Журнал химической теории и вычислений . 8 (8): 2949–2961. дои : 10.1021/ct300377a . ПМЦ 3489502 . ПМИД 23139645 .
- ^ Ривальта I, Султан ММ, Ли Н.С., Мэнли Г.А., Лория Дж.П., Батиста В.С. (май 2012 г.). «Аллостерические пути имидазолглицеринфосфатсинтазы» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (22): E1428–36. дои : 10.1073/pnas.1120536109 . ПМК 3365145 . ПМИД 22586084 .
- ^ Негре К.Ф., Морзан У.Н., Хендриксон Х.П., Пал Р., Лиси Г.П., Лория Дж.П. и др. (декабрь 2018 г.). «Центральность собственного вектора для характеристики аллостерических путей белков» . Труды Национальной академии наук Соединенных Штатов Америки . 115 (52): Е12201–Е12208. arXiv : 1706.02327 . Бибкод : 2018PNAS..11512201N . дои : 10.1073/pnas.1810452115 . ПМК 6310864 . ПМИД 30530700 .
- ^ Кларк Д., Сетхи А., Ли С., Кумар С., Чанг Р.В., Чен Дж. и др. (май 2016 г.). «Выявление аллостерических горячих точек с помощью динамики: применение к меж- и внутривидовой сохранению» . Структура . 24 (5): 826–837. дои : 10.1016/j.str.2016.03.008 . ПМЦ 4883016 . ПМИД 27066750 .
- ^ Дутта С., Экманн Дж.П., Либхабер А., Тлусти Т. (май 2018 г.). «Зеленая функция коррелирующих генов в минимальной механической модели эволюции белка» . Труды Национальной академии наук Соединенных Штатов Америки . 115 (20): Е4559–Е4568. arXiv : 1801.03681 . Бибкод : 2018PNAS..115E4559D . дои : 10.1073/pnas.1716215115 . ПМК 5960285 . ПМИД 29712824 .
Внешние ссылки
[ редактировать ]- Мгновенное представление о системе классификации механизмов аллостерии белков от Королевского химического общества.