Насекомое крыло




Накрытия насекомых - это взрослые роты насекомых экзоскелета , которые позволяют насекомым летать . Они встречаются во втором и третьем грудном сегментах ( мезоторакс и метатораксах ), и две пары часто называют передними и задними крыльями , соответственно, хотя у нескольких насекомых не хватает задних крыльев, даже во прав. Крылья укрепляются рядом продольных вен, которые часто имеют перекрестные соединения, которые образуют закрытые «ячейки» в мембране (экстремальные примеры включают стрекозы и кружевные крылышки ). Паттерны, возникающие в результате слияния и перекрестного соединения вен крыла, часто являются диагностическими для различных эволюционных линий и могут использоваться для идентификации семейства или даже уровня рода во многих порядках насекомых.
Физически некоторые насекомые перемещают мышцы полета непосредственно, другие косвенно. У насекомых с прямым полетом мышцы крыла непосредственно прикрепляются к основанию крыла, так что небольшое нисходящее движение основания крыла поднимает само крыло вверх. У этих насекомых с косвенным полетом есть мышцы, которые прикрепляются к грудной клетке и деформируются, в результате чего крылья также движутся.
Крылья присутствуют только в одном поне (часто мужчине) в некоторых группах, таких как бархатные муравьи и стрепспиптера , или избирательно теряются у «работников» социальных насекомых, таких как муравьи и термиты . Редко самка крылатая, но мужчина нет, как в рис -осах . В некоторых случаях крылья производятся только в определенное время в жизненном цикле, например, в фазе рассеивания тлей . Структура крыла и окраска часто варьируются в зависимости от морфов , например, в тля , миграционные фазы саранчи и полиморфные бабочки . В состоянии покоя крылья могут удерживаться плоскими или складываемыми несколько раз по определенным узорам; В большинстве случаев, это задних крыльев, которые сложены, но в нескольких группах, таких как Vespid Wasps , это предхищения.
Эволюционное происхождение крыла насекомых обсуждается. В течение 19 -го века вопрос об эволюции крыла насекомых изначально опирался на две основные позиции. Одна позиция постулированной крылья насекомых развивалась из ранее существовавших структур, в то время как вторые предложенные крылья насекомых были совершенно новыми формациями. [ 1 ] [ 2 ] «Новая» гипотеза предполагала, что крылья насекомых формируются не из ранее существовавших предков, а скорее как рост от стены тела насекомых. [ 3 ]
Давно исследование о происхождении крыла насекомых основано на позиции «ранее существовавших структур», которая была первоначально предложена в 19 веке. [ 2 ] Недавняя литература указала на несколько наследственных структур как важных для происхождения крыльев насекомых. Среди них являются: жабры, респираторные придатки ног, а также боковые (паранотальные) и постеролатеральные проекции грудной клетки и многие другие. [ 4 ]
Согласно более современной литературе, возможные кандидаты включают в себя жаберные структуры, паранотическую долю и ракообразную тергарную плиту. Последнее основано на недавних генетических исследованиях насекомых, которые указывают на то, что насекомые представляют собой членистоногих пан-кортацев с прямым ракообразным предком и общими генетическими механизмами развития конечностей. [ 3 ] [ 5 ] [ 6 ] [ 7 ] [ 8 ]
Другими теориями происхождения крыльев насекомых являются теория паранотальной доли, теория жабры и двойная теория эволюции крыла насекомых. Эти теории постулируют, что крылья либо развивались из паранотических долей, расширения грудной территории ; [ 5 ] что они представляют собой модификации подвижных брюшных жабра которые обнаруживаются на водных наядах Мэйфлс , ; [ 5 ] Или что крылья насекомых возникли из-за слияния ранее существовавших конфиденциальных и выходящих структур, каждая из которых имела ранее существовавшую артикуляцию и трахэ. [ 9 ] [ 10 ]
Морфология
[ редактировать ]
Внутренний
[ редактировать ]Каждое из крыльев состоит из тонкой мембраны, поддерживаемой системой вен. Мембрана образуется двумя слоями покровного положения, тесно расположенной, в то время как вены образуются там, где два слоя остаются отдельными; Иногда нижняя кутикула толще и более склеротизируется под веной. В каждом из основных вен есть нерв и трахея , и, поскольку полости вен связаны с гемокоэлем , гемолимфа может течь в крылья. [ 11 ]
По мере развития крыла дорсальные и вентральные покровные слои становятся тесно обоснованными по большей части своей области, образующей мембрану крыла. Остальные области образуют каналы, будущие вены, в которых могут возникать нервы и трахей. Кутикула, окружающая вены, становится утолщенной и более склеротизирована, чтобы обеспечить прочность и жесткость для крыла. На крыльях могут возникнуть два типа волос: микротрихия, которые небольшие и нерегулярно разбросаны, а макротрихия, которые больше, гнездо, и могут быть ограничены венами. Масштабы Lepidoptera и Trichoptera являются сильно модифицированными макротрихией. [ 12 ]
Венеция
[ редактировать ]
У некоторых очень маленьких насекомых венеция может быть значительно уменьшено. Например, в хальцидоидных осах присутствуют только подкоста и часть радиуса. И наоборот, увеличение венеции может произойти благодаря разветвлению существующих вен для производства вспомогательных вен или развитием дополнительных интеркалярных вен между первоначальными, как в крыльях ортоптер (кузнечики и сверчки). Большое количество перекрестных вещей присутствует у некоторых насекомых, и они могут образовывать ретикулум, как в крыльях одонаты (драконки и самих) и у основания передних крыльев Tettigonioidea и Acridoidea (Katydids и Grasshoppers соответственно). [ 11 ]
Archedictyon - это название , данное гипотетической схеме венения крыла, предложенной для самого первого крылатого насекомого. Он основан на комбинации спекуляций и данных ископаемых. Поскольку все крылатые насекомые, как полагают, развивались от общего предка, Archedictyon представляет «шаблон», который был изменен (и обтекает) естественным отбором в течение 200 миллионов лет. Согласно нынешней догме, Archedictyon содержал 6–8 продольных вен. Эти вены (и их филиалы) названы в соответствии с системой, разработанной Джоном Комстоком и Джорджем Нидхэмом - системой Comstock -Needham : [ 13 ]
- Коста (с) - передний край крыла
- Subcosta (SC) - вторая продольная вена (позади коста), как правило, не расставленная
- Радиус (R) - третья продольная вена, от одной -пяти ветвей достигают края крыла
- СМИ (M) - Четвертая продольная вена, одна -четыре ветви достигают края крыла
- Cubitus (Cu) - Пятая продольная вена, одна -три ветви достигает края крыла
- Анальные вены (A1, A2, A3) - неразветвленные вены за кубитом
Коста (C) является ведущей маргинальной веной для большинства насекомых. Иногда выше коста, называемой Precosta, существует небольшая вена, хотя почти во всех существующих насекомых, [ 14 ] : 41–42 Precosta сливается с Costa. Коста редко ветвет, потому что он находится на переднем крае, который связан с его основанием с плечевой пластиной. Трахея костюшной вены, возможно, является ветвью подкостальной трахеи. Расположенный после Коста является третьей веной, подкоста, которая разветвляется в две отдельные вены: передние и задние. Основание подкоста связано с дистальным концом шеи первого подмышечности (см. Раздел ниже). Четвертая вена - это радиус (R), которая разветвлена на пять отдельных вен. Радиус, как правило, самая сильная вена крыла. К середине крыла он разбивается в первую неразделенную ветвь (R1) и вторую ветвь, называемую радиальным сектором (RA), которая дихотомично подразделяет на четыре дистальные ветви (R2, R3, R4, R5). В основном радиус гибко объединен с передним концом второго подмышечного (2AX). [ 15 ]
Пятая вена крыла - это СМИ. В паттерне архетипа (а) медиа в две основные ветви: передняя (мА) среда (MA), которая делится на две дистальные ветви (MA1, MA2) и средний сектор или задний (MP), который имеет четыре Терминальные ветви (M1, M2, M3, M4). У большинства современных насекомых передняя часть средств массовой информации была потеряна, а обычные «СМИ»-это четыреразветвленные средства массовой информации с общим базальным стеблем. В Ephemerida, согласно нынешней интерпретациям венения крыла, обе ветви средств массовой информации сохраняются, в то время как в Одонате сохраняющиеся среды являются примитивной передней ветвью. Стебель средств массовой информации часто объединяется с радиусом, но когда он встречается в качестве отдельного вены, его основание связано с дистальной медианой (M ') или непрерывно склеротизируется последним. Кубит, шестая вена крыла, в первую очередь двумя разветвленными. Основная развязка происходит вблизи основания крыла, образуя две основные ветви (CU1, CU2). Передняя ветвь может распаться на несколько вторичных ветвей, но обычно она разбивается в две дистальные ветви. Вторая ветвь Cubitus (Cu2) в Hymenoptera, Trichoptera и Lepidoptera была принята Comstock и Neepham за первый анал. Проксимально главный стебель кубита связан с дистальной медианной пластиной (M ') основания крыла. [ 15 ]
Postcubitus (PCU) является первым аналом системы Comstock -Needham. Посткубит, однако, имеет статус независимой крыланой вены и должен быть признан таковым. [ Цитация необходима ] У крыльев нимфы его трахея возникает между кубитальной трахей и группой Vannal Tracheae. У зрелых крыльев более обобщенного насекомых посткубит всегда ассоциируется проксимально с кубитом и никогда не тесно связан со склеритом сгибателя (3AX) основания крыла. В Neuroptera, Mecoptera и Trichoptera посткубит может быть более тесно связан с Vannal -венами, но его основание всегда свободна от последних. Посткубит обычно неразветвлен; Это примитивно два разветвленных. Вена Vannal (LV до NV) представляют собой анальные вены, которые сразу же связаны с третьим подмышечным устройством и которые напрямую влияют движение этого склерита, которое приводит к сгибанию крыльев. В числе ваннал вены различаются. с 1 до 12, в соответствии с расширением области ваннала крыла. Vannal Tracheae обычно возникает из общего ствола трахеи у нимфальных насекомых, а вены рассматриваются как ветви одной анальной вены. Дистально ваннальские вены либо просты, либо разветвлены. Югальские вены (J) Югальной доли крыла часто заняты сетью нерегулярных вен, или это может быть полностью мембранным; Но иногда он содержит одну или два различных маленьких вен, первая югальская вена или вена Аркуата, а также вторая югальская вена или вена кардиналас (2J). [ 15 ]
- C-Sc Cross-Veins -запустите между Costa и Subcosta
- R Cross-Veins -пробежать между соседними ветвями радиуса
- RM Cross-Veins -пробежать между радиусом и медиа
- M-Cu Cross-Veins -беги между средствами массовой информации и Cubitus
Все вены крыла подвергаются вторичному развящению и союзу перекрестными вещами. В некоторых приказах насекомых перекрестных веков настолько многочисленны, что весь заднациональный паттерн становится близкой сетью ветвящихся вен и перекрестных вещей. Обычно, однако, существует определенное количество перекрестных вещей, имеющих определенные места. Более постоянными перекрестными вещами являются кросс-ввеина (H) между костами и подкостами, радиальная перекрестная веща (r) между R и первой вилкой RS, секторационная перекрестная веба (ы) между двумя вилками R8, медианная перекрестная ввея (M-M) между M2 и M3 и медиокубитальная поперечная веду (M-CU) между средами и кубитом. [ 15 ]
Вены крыльев насекомых характеризуются выпуклой разместимостью, такими как те, которые наблюдаются в Mayflies (то есть, вогнутые «вниз», а выпуклость «вверх»), которые регулярно чередуются и по его триадному типу ветвления; Всякий раз, когда вена вилка всегда существует интерполированная вена противоположного положения между двумя ветвями. Вогнутая вена разворачивается в двух вогнутых венах (с выпуклой интерполированной вены) и сохраняется регулярное изменение вен. [ 16 ] Вены крыла, по -видимому, попадают в волнистый рисунок в зависимости от того, имеют ли они тенденцию складываться или вниз, когда крыло расслаблено. Базальные валы вен выпуклые, но каждая вена вилки дистально дистально в переднюю выпуклую ветвь и заднюю вогнутую ветвь. Таким образом, Коста и подкоста рассматриваются как выпуклые и вогнутые ветви первичной первой вены, RS является вогнутой ветвью радиуса, а задней среды - вогнутая ветвь среда, Cu1 и Cu2, соответственно выпуклые и вогнутые, в то время как примитивный посткабит и Первый ваннал имеет каждую переднюю выпуклую ветвь и заднюю вогнутую ветвь. Выпуклая или вогнутая природа вен использовалась в качестве доказательства при определении идентичности постоянных дистальных ветвей вен современных насекомых, но не было продемонстрировано, что он является последовательным для всех крыльев. [ 11 ] [ 15 ]
Поля
[ редактировать ]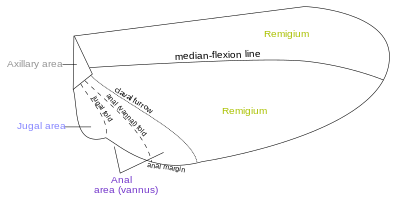
Области крыла разграничиваются и подразделяются по линии сгиба, вдоль которой крыло может складываться, и линии сгибания, вдоль которых крыло может сгибаться во время полета. Фундаментальное различие между линией сгибания и линии сгиба часто размыто, так как линии сгиба могут допустить некоторую гибкость или наоборот. Две константы, которые встречаются почти во всех крылах насекомых,-это клавал (линия сгибания) и югольные складки (или линия сгиба); формирование переменных и неудовлетворительных границ. Формирование крыльев может быть очень сложным, с поперечным складыванием, происходящим в задних крылах дермаптера и Coleoptera , а у некоторых насекомых анальная область может быть сложена как вентилятор. [ 14 ] На крыльях насекомых найдено около четырех разных полей:
- Катание на лодке
- Анальная область (Ваннус)
- Югальная зона
- Подмышечная область
- Продлить
Большинство вен и кроссвеины встречаются в передней области Remigium , который отвечает за большую часть полета, прикрепленные к грудным мышцам. Задняя часть Ремигиума иногда называют клавином ; Два других задних полей являются анал и югал Ares . [ 14 ] Когда складка Vannal имеет обычное положение перед группой анальных вен, Remigium содержит костровая, подрезок, радиальная, медиальная, кубитальная и почтовая вены. В сгибленном крыле переосмысление сзади на гибкое базальное соединение радиуса со вторым подмышечным органом, а основание медиокубитального поля сложена медиально на подмышечной области вдоль базалиса PLICA (BF) между медианными пластинами (M, M ' ) основания крыла. [ 15 ]
Ваннус граничит с складкой Ваннала, которая обычно происходит между почткубитом и первой веной. В Orthoptera это обычно имеет эту позицию. Однако в переднем крыле Blattidae единственная складка в этой части крыла лежит непосредственно перед почтком. В Plecoptera складка Vannal является задней частью почты, но проксимально она пересекает основание первой вены. В Cicada складка Vannal находится непосредственно за первой ваннал -веной (LV). Однако эти небольшие вариации в фактическом положении сгиба ваннала не влияют на единство действия ваннальных вен, контролируемых склеритом сгибателя (3AX), при сгибании крыла. В задних крылах большинства ортоптер вторичная вена дивидента образует ребро в складке ваннала. Ваннус обычно имеет треугольную форму, и его вены обычно распространяются из третьего подмышечного устройства, как ребра вентилятора. Некоторые из ваннальных вен могут быть разветвлены, а вторичные вены могут чередоваться с первичными венами. Область Ваннала, как правило, лучше всего развивается в задних крылах, в которой она может быть увеличена, чтобы сформировать устойчивую поверхность, как в Plecoptera и Orthoptera. Великие фанатные расширения задних крыльев Acrididae, безусловно, являются областями ваннала, поскольку все их вены поддерживаются на третьем подмышечных склеритах на базах крыла, хотя Мартинов (1925) приписывает большинство областей фанатов в Acrididae к Jugal Regions of the крылья. Истинный югум акридного крыла представлен только маленькой мембранной (JU) мезадом последней ваннал вены. Jugum более развит в некоторых других полинеоптерах, как в Mantidae. У большинства высших насекомых с узкими крыльями ваннус уменьшается, и складка ваннала теряется, но даже в таких случаях изгибное крыло может сгибаться вдоль линии между почткубитом и первой веной. [ 15 ]
Югальная область, или Нила, представляет собой область крыла, которая обычно представляет собой небольшую мембранную область, проксимальную к основанию Ваннуса, усиленной несколькими небольшими нерегулярными венами, похожими на вены; Но когда он хорошо развит, это отдельная часть крыла и может содержать одну или две югальные вены. Когда югальная область переднего крыла разрабатывается как свободная доля, она проецирует под углом на плече и, таким образом, служит для того, чтобы следовать два крыла вместе. В группе jugatae of Lepidoptera у него длинная доля. Джугальная область была названа Нилой («Новое крыло»), потому что она, очевидно, вторичная и недавно разработала часть крыла. [ 15 ]
Подмышечная область - это область, содержащая подмышечные склериты, имеет в целом форма скалоневого треугольника. Основание треугольника (AB) - это шарнир крыла с телом; вершина (C) является дистальным концом третьего подмышечного склерита; Более длинная сторона перед вершиной. Точка D на передней стороне треугольника отмечает артикуляцию радиальной вены вторым подмышечным склеритом. Линия между D и C - это Plica Basalis (BF) или складка крыла у основания посредника поля. [ 15 ] Терден - это внешний край крыла, между вершиной и задним или анальным углом. [ 17 ] [ 18 ]
На заднем угле основания крыла в некоторых диптерах есть пара мембранных доли (квадрат или калиптер), известная как алула. Алула хорошо развита в домашней мух. Внешняя квадрат (C) возникает из основания крыла за третьим подмышечным склеритом (3AX) и, очевидно, представляет юлюскую долю других насекомых (A, D); Большая внутренняя квадрат (D) возникает из заднего кувчарного края тергума сегмента с крыльями и образует защитный, капютный навес над Халтер. В изгибном крыле внешняя квадрат алюлы переворачивается над внутренней квадрат, последняя не подвержена влиянию движения крыла. Во многих диптерах глубокий разрез анальной области крыланой мембраны за единой ваннал -веной выходит из проксимальной доли алар, дистальной к внешней кваме алюлы. [ 15 ]
Суставы
[ редактировать ]
Различные движения крыльев, особенно у насекомых, которые сгибают крылья горизонтально над спиной, когда в состоянии покоя, требуют более сложной суставной структуры у основания крыла, чем просто шарнир крыла с телом. Каждое крыло прикреплено к телу мембранной базальной областью, но суставная мембрана содержит ряд небольших суставных склеритов, известных как птералия. Птералия включает в себя переднюю плечевую пластину у основания костюшной вены, группу подмышечных (AX), связанных с подрезок, радиальными и ванналь Медиокубитальная область. Подмышечные характеристики специально разработаны только у насекомых с смешиванием крыла, где они составляют механизм сгибателя крыла, эксплуатируемого сгибательными мышцами, возникающими на плероне. Характеристикой основания крыла также является небольшая доля на переднем крае суставной области, проксимальной к тарелке плечевой кости, которая при переднем крыте некоторых насекомых развивается в большой плоский, масштабный лоскут, тегула, перекрывая основание крыла. Сзади суставная мембрана часто образует достаточную долю между крылом и телом, и ее край обычно утолщается и гофрирован, придавая появлению связки, так называемой подмышечной шнурки, непрерывной мезами с задней краевой кутеллярной складкой тергального Тарелка с крылом. [ 15 ]
Суставные склериты, или птералия из основания крыльев сгибающих насекомых с крыльями и их отношений с телом и вен крыла, показанные диаграмма, являются следующими:
- Пластины плечевой кости
- Первый подмышечный
- Второй подмышечный
- Третий подмышечный
- Четвертый подмышечный
- Средние тарелки ( м , м ' )
Пластина плечевой кости обычно представляет собой небольшой склерит на переднем крае основания крыла, подвижным и сочлененным с основанием костюшной вены. У odonata их плетеные тарелки значительно расширены, [ 15 ] с двумя мышцами, возникающими в результате эпистерана, вставленного в плечевые пластины, и две из края эпимерона, вставленного в подмышечную пластину. [ 11 ]
Первый подмышечный склерит (слабый) - это передняя петля пластина основания крыла. Его передняя часть поддерживается на переднем процессе нотальского крыла Тергума (ANP); Его задняя часть сочленена с тергальным краем. Передний конец склерита обычно производится как тонкая рука, вершина которой (e) всегда связана с основанием подкостальной вены (SC), хотя она не объединена с последним. Тело склерита формулируется в поперечном направлении со вторым подмышечным устройством. Второй подмышечный склерит (2AX) является более изменчивым по форме, чем первый подмышечный, но его механические отношения не менее определенные. Он наклонно шарнир к внешнему краю тела первого подмышечности, а радиальная вена (R) всегда гибко прикреплена к его переднюю часть (D). Второй подмышечный характер представляет как дорсальную, так и вентральную склеротизацию в основании крыла; Его вентральная поверхность опирается на процесс фолочного крыла плерона. Следовательно, второй подмышечный, представляет собой ключевой склерик основания крыла, и он специфически манипулирует радиальной веной. [ 15 ]
Третий подмышечный склерит (3AX) лежит в задней части суставной области крыла. Его форма сильно варьируется и часто нерегулярна, но третий подмышечный, - это склерит, в который вставляется мышца сгибателя крыла (D). Месально он сочленен спереди (f) с задним концом второго подмышечного, а сзади (b) с процессом заднего крыла тергума (PNP) или с небольшим четвертым подмышечным, когда присутствует последнее. Дистально третий подмышечный, продлен в процессе, который всегда связан с основаниями группы вен в анальной области крыла, здесь называемых Vannal Vanal -венами (V). Следовательно, третий подмышечный, обычно представляет собой заднюю шарнирную пластину основания крыла и является активным склеритом механизма сгибателя, который непосредственно манипулирует венами ваннала. Сокращение мышцы сгибателей (D) вращается третьим подмышечным, на его мезальную артикуляции (B, F) и тем самым поднимает его дистальную руку; Это движение производит сгибание крыла. Четвертый подмышечный склерит не является постоянным элементом основания крыла. Когда присутствует, это обычно небольшая пластина, вмешивающаяся между третьим подмышечным и задним процессом крыла Notal, и, вероятно, является отдельным кусочком последнего. [ 15 ]
Медианные пластины (M, M ') также являются склеритами, которые не настолько определенно дифференцированы, как конкретные пластины, как и три основных подмышечных характера, но, тем не менее, они являются важными элементами аппарата сгибателя. Они лежат в срединной области основания крыла, дистально ко второму и третьему подмышкам и отделены друг от друга наклонной линией (BF), которая образует выдающуюся выпуклую складку во время сгибания крыла. Проксимальная пластина (M) обычно прикрепляется к дистальному руку третьего подмышечного жителя и, возможно, следует рассматривать как часть последнего. Дистальная пластина (M ') менее постоянно присутствует в виде отдельного склерита и может быть представлена общей склеротизацией основания посредника поля крыла. Когда вены этого региона различаются на своих основаниях, они связаны с внешней средней плитой. [ 15 ]
Мышцы
[ редактировать ]
Мышцы, которые контролируют полеты у насекомых, могут занять до 10% до 30% от общей массы тела. Мышцы, которые контролируют полеты, варьируются в зависимости от двух типов полета у насекомых: косвенные и прямые. Насекомые, которые используют сначала, косвенные, заставляют мышцы прикрепляться к территории вместо крыльев, как следует из названия. Когда мышцы сокращаются, грудная ящик искажается, передавая энергию в крыло. Есть два «пучка» мышц, которые охватывают параллельные территории, дорсолонгитудиналы и те, которые прикреплены к Тегуму и распространяются на грудину, дорсовентралы. [ 19 ] В прямой мышце соединение находится прямо от плерона (грудной стены) до отдельных склеритов, расположенных у основания крыла. Субалар и базилярные мышцы имеют прикрепления связок к субалар и базилярным склеритам. Здесь Resilin, высоко эластичный материал, образует связки, соединяющие летные мышцы с аппаратом крыла.
В более полученных порядках насекомых, таких как Diptera (мухи) и гименоптера (WASP), косвенные мышцы занимают наибольший объем птероторакса и функционируют в качестве основного источника мощности для крыс. Сокращение дорсолонгитудированных мышц вызывает сильное изгибание ноту, которое подавляет крыло, в то время как сокращение дорсовентральных мышц вызывает противоположное движение Notum. Наиболее примитивные существующие летающие насекомые, эфемероптеры (Мэйфлис) и Одоната (дракозоны) используют прямые мышцы, которые ответственны за развитие необходимой силы для подъездных и вниз. [ 19 ] [ 20 ]
Мышца крыла насекомых - это строго аэробная ткань. На единицу белка он потребляет топливо и кислород со скоростью, происходящими в очень концентрированной и высокоорганизованной ткани, так что устойчивые скорости на единицу объема представляют собой абсолютную запись в биологии. Кровь, богатая топливом и кислородом, переносится в мышцы посредством диффузии, происходящей в больших количествах, для поддержания высокого уровня энергии, используемой во время полета. Многие мышцы крыльев большие и могут быть длиной 10 мм и шириной 2 мм. Более того, в некоторых диптерах волокна имеют гигантские измерения. Например, в очень активной рутилии поперечное сечение имеет длину 1800 мкм и ширину более 500 мкм. Транспортировка топлива и кислорода из окружающей среды в участки потребления и обратный транспорт диоксида углерода, следовательно, представляет собой проблему для биолога как в отношении транспортировки в жидкой фазе, так и в сложной системе воздушных труб, т.е. в трахеи система. [ 21 ]
Датчики
[ редактировать ]Несколько типов сенсорных нейронов обнаружены на крыльях насекомых: вкусовые щетинки , [ 22 ] Механоосенсорные щетинки, [ 23 ] Компаниформ Sensilla , [ 24 ] и хордотональные органы . [ 25 ] Эти датчики обеспечивают нервную систему как внешней, так и внутренней проприоцептивной обратной связи, необходимой для эффективного полета [ 26 ] и уход. [ 27 ]
Связь, складывание и другие функции
[ редактировать ]У многих видов насекомых передвижение и задних крыльев могут быть связаны друг с другом, что повышает аэродинамическую эффективность полета, соединяя передвижение и задних крыла в одно большее крыло. Наиболее распространенным механизмом связи (например, Hymenoptera и Trichoptera ) является ряд небольших крючков на переднем крае задних крыльев, или « hamuli », который фиксируется на переднем крыле, сдерживая их вместе (Hamulate Coupling). У некоторых других видов насекомых (например, Mecoptera , Lepidoptera и некоторых трихоптеров ) Югальная доля переднего крыла покрывает часть заднего крыла (Jugal Coupling), или на окраинах переднего гусени Христовые щетинки, или френулум, крючок под подпорной структурой или сетчаткой на переднем крыте. [ 14 ] : 43
В состоянии покоя крылья удерживаются над спиной у большинства насекомых, что может включать в себя продольное складывание мембраны крыла, а иногда и поперечное складывание. Складывание может иногда происходить вдоль линий сгибания. Хотя линии сгиба могут быть поперечными, как и в задних крылах жуков и ушных водорослей, они обычно радиальны к основанию крыла, что позволяет сложить соседние участки крыла или под друг другу. Самая распространенная линия сгиба - складка Jugal, расположенная сразу за третьей анальной веной, [ 12 ] Хотя у большинства неоптеров есть югальная складка сразу за вене 3А на передних крытях. Иногда это также присутствует на задних крылах. Там, где анальная область задних крыльев большая, как у ортоптер и блюттодиа, вся эта часть может быть сложена под передней частью крыла вдоль сгиба в ваннале, немного задней к борозду клавала. Кроме того, в ортоптерах и блюттодии анальная область сложена как вентилятор вдоль вен, а анальные вены выпуклые, на гребнях складок и вспомогательные вены вогнуты. Принимая во внимание, что бороздка клавала и югальная складка, вероятно, гомологичны различным видам, складка ваннала варьируется в положении в разных таксонах. Складывание вырабатывается мышцей, возникающим на плероне и вставляется в третий подмышечный склерит таким образом, что, когда он сжимается, склерит поворачивается вокруг его точек артикуляции с помощью процесса задних ноталь и второго подмышечного склерита. [ 11 ]
В результате дистальный рычаг третьего подмышечного склерита вращается вверх и внутрь, так что, наконец, его положение полностью изменено. Анальные вены сформулированы с этим склеритом таким образом, что при движении они переносятся с ним и согнуты над задней частью насекомых. Активность той же мышцы в полете влияет на выходную мощность крыла, и поэтому она также важна для управления полетом. У ортоптероидных насекомых эластичность кутикулы заставляет область ваннала крыла складываться вдоль вен. Следовательно, энергия тратит на развертывание этого региона, когда крылья перемещаются в положение полета. В целом, расширение крыла, вероятно, является результатом сокращения мышц, прикрепленных к базилярному склериту или, у некоторых насекомых, к субаларскому склериту. [ 11 ]
Полет
[ редактировать ]
Механизмы полета
[ редактировать ]Две группы относительно больших насекомых, эфемероптеры ( майки ) и одоната ( дракозоны и самих ) имеют мышцы полета, прикрепленные непосредственно к их крыльям; Крылья не могут преодолевать не так быстрее, чем скорость, с которой нервы могут посылать импульсы, чтобы командовать мышцами взбивать. [ 28 ] Все остальные живые крылатые насекомые летают, используя другой механизм, включающий непрямые летные мышцы, которые вызывают вибрацию грудной клетки; Крылья могут бить быстрее, чем скорость, с которой мышцы получают нервные импульсы. Этот механизм развивался один раз, и является определяющей особенностью ( синапоморфий ) для инфракласной неоптерии . [ 28 ]
Аэродинамика
[ редактировать ]Есть две основные аэродинамические модели полета насекомых. Большинство насекомых используют метод, который создает спирального переднего края вихрь . [ 29 ] [ 30 ] Некоторые очень маленькие насекомые используют механизм блинчиков и хлопок или Вейс-Фог, в котором крылья хлопают вместе над телом насекомого, а затем разбрасываются. Когда они открываются, воздух втягивается и создает вихрь над каждым крылом. Этот связанный вихрь затем движется через крыло и в хлопок действует как стартовый вихрь для другого крыла. Циркуляция и лифт увеличиваются по цене износа на крыльях. [ 29 ] [ 30 ]
Многие насекомые могут зависать, быстро избивая свои крылья, требуя боковой стабилизации, а также подъема. [ 31 ]
Несколько насекомых используют скользящий полет без использования тяги.
Эволюция
[ редактировать ]Когда -то в каменноугольный период , около 350 миллионов лет назад, когда было только две основные земельные массы, насекомые начали летать. Как и почему развивались крылья насекомых, однако, недостаточно понятно, в значительной степени из -за нехватки соответствующих окаменелостей из периода их развития в нижнем углероде. Три основные теории о происхождении полета насекомых заключаются в том, что крылья, разработанные из паранотических дол, расширения грудной территории ; что они представляют собой модификации подвижных брюшных жабра которые обнаруживаются на водных наядах Мэйфлс , ; или что они разработали от грудных выступов, используемых в качестве радиаторов . [ 32 ]
Окаменелости
[ редактировать ]
Окаменелости от Девонца (400 миллионов лет назад) - все без крыла, но по карбоне (320 миллионов лет назад) более 10 различных родов насекомых имели полностью функциональные крылья. Между двумя периодами мало сохранения переходных форм. Самые ранние крылатые насекомые-это период времени ( Pterygota ), включая Blattoptera , Caloneurodea стволовой группы , примитивные эфемероптеры , Orthoptera и Palaeodictyopteroidea . Очень ранние блюттоптераны (во время каменноугольника) имели очень большую дискоидную переднеспинку и кориасовые ветки с отчетливой чашкой (неразветвленная вена крыла, лежащая возле клавала и достигая заднего края крыла). [ 33 ] : 399 Несмотря на то, что самым старым возможным ископаемым насекомым является девонский риниогнатха Хирсти , оцениваемый в 396–407 миллионов лет, он обладал дикондиными нижней частью, особенность, связанная с крылатыми насекомыми, [ 34 ] Хотя позже это считается как можно более мирипод . [ 35 ]
Во время перми дракозонами . ( одоната ) были доминирующим воздушным хищником и, вероятно, доминировали и преобладали хищничество на наземных насекомых Истинная Одоната появилась на пермском [ 36 ] [ 37 ] И все являются амфибийными . Их прототипы - самые старые крылатые окаменелости, [ 38 ] Вернитесь к девонцам и во всех отношениях отличаются от других крыльев. [ 39 ] Их прототипы, возможно, имели начало многих современных атрибутов даже по позднему каменноугольнику , и возможно, что они даже захватили мелких позвоночных, поскольку у некоторых видов был промежуток крыльев 71 см. [ 37 ] Самые ранние виды, похожие на жука во время пермского, имели заостренные кожа, похожие на кожу с клетками и ямами. Hemiptera , или настоящие ошибки появились в форме Arctiniscytina и Paraknightia, которые имели напряженность с необычным венчением, возможно, расходящиеся от Blattoptera . [ 33 ] : 186
Одно большое крыло от вида диптеры в триасе (10 мм вместо обычного 2–6 мм) было обнаружено в Австралии (Mt. Crosby). Это семейство Tilliardipteridae, несмотря на многочисленные «синулоидные» особенности, следует включить в Psychodomorpha. Sensu hennig из -за потери выпуклого дистального 1A, достигающего края крыла и образования анальной петли. [ 40 ]
Гипотезы
[ редактировать ]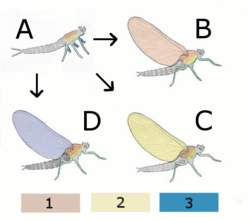
Гипотетический предок без крыла
B Паранотическая теория:
Гипотетическое насекомое с крыльями со спины (notum)
C Гипотетическое насекомое с крыльями из плерума
D Эпиксальская теория
Гипотетическое насекомое с крыльями из приложения ног
1 notum (назад)
2 плерума
3 Выход (Внешние прикрепления ног)
- Паранотическая гипотеза : эта гипотеза предполагает, что крылья насекомого развивались из паранотических дол, предварительная предварительная опция, обнаруженная у окаменелостей насекомых , которые, как полагают, помогали стабилизации при прыжке или падении. В пользу этой гипотезы является тенденция большинства насекомых, когда они поражаются во время подъема на ветви, сбежать, упав на землю. Такие доли служили парашютами и позволяли насекомому приземлиться более мягко. Теория предполагает, что эти доли постепенно росли больше, а на более поздней стадии разработал сустав с грудной клеткой. Даже позже появится мышцы, чтобы переместить эти сырые крылья. Эта модель подразумевает прогрессивное повышение эффективности крыльев, начиная с парашютирования , затем скользящего и, наконец, активного полета . Тем не менее, отсутствие существенных ископаемых доказательств развития крыльев и мышц представляет собой серьезную трудности для теории, как и, казалось бы, спонтанное развитие артикуляции и венеции. [ 32 ]
- Эпиксальская гипотеза : эта теория, впервые предложенная в 1870 году Карлом Гегенбауром, предположила, что возможным происхождением для крыльев насекомых могло быть подвижные брюшные жабра, обнаруженные у многих водных насекомых, таких как на Naiads of Mayflies . [ 41 ] Согласно этой теории, эти трахеальные жабры, которые начались в качестве выхода дыхательной системы и со временем были изменены в локомотивные цели, в конечном итоге превратились в крылья. Трахеальные жабры оснащены маленькими крыльями, которые постоянно вибрируют и имеют свои крошечные прямые мышцы. [ 32 ]
- Гипотеза Endite-Exit : эта гипотеза связана с адаптацией эндийных и выпускных, придатков к соответствующим внутренним и внешним аспектам примитивной конечности членистоногих. Это было продвинуто Trueman [ 42 ] Основываясь на исследовании Goldschmidt в 1945 году на Drosophila melanogaster , в котором вариация POD демонстрировала мутацию, трансформирующую нормальные крылья в то, что интерпретировалось как трехместное расположение ног с некоторыми дополнительными придатками, но не хватало лавша, где обычно можно было быть. Эта мутация была переосмыслена как убедительные доказательства дорсального выхода и экинтного слияния, а не ноги, причем придатки гораздо лучше подходили к этой гипотезе. Иннервация, артикуляция и мускулатура, необходимые для эволюции крыльев, уже присутствуют в подомерах. [ 32 ]
- Гипотеза рекрутирования гена Paranota Plus (также известная как гипотеза двойного происхождения) : ископаемые личинки Coxoplectoptera дали важные новые подсказки для оспариваемого вопроса эволюционного происхождения крыльев насекомых. Перед обнаружением ископаемых личинок паранотальная гипотеза и гипотеза с ногами считались несовместимыми альтернативными объяснениями, которые были подтверждены набором доказательств из ископаемого записи, сравнительной морфологии , биологии развития и генетики . Экспрессия генов ног в онтогенезе крыла насекомых повсеместно рассматривалась как убедительные доказательства в пользу гипотезы о ногах, которая предлагает, чтобы крылья насекомых были получены из придаток мобильных ног (Exceents). Тем не менее, личинки Coxoplectoptera показывают, что брюшные жабра от майфей и их предков, которые обычно считаются соответствующими структурами для насекомых крыльев, сформулированных в дорсальных тергитовых пластинах. Это нельзя увидеть у современных личинок Мэйфля, потому что их брюшные тергиты и стерниты сливаются с кольцами, без каких -либо следов, оставшихся даже при эмбриональном развитии. Если личиночные жабры и крылья являются соответствующими («серийными гомологичными») структурами и, таким образом, имеют одно и то же эволюционное происхождение, новые результаты Coxoplectoptera демонстрируют, что крылья также имеют тергальное происхождение, что предлагается классической паранотальной гипотезой. Staniczek, Bechly & Godunko (2011) [ 32 ] [ 43 ] Поэтому предположила новую гипотезу, которая могла бы согласовать, по -видимому, противоречивые доказательства из палеонтологии и генетики развития : крылья впервые возникли как жесткие начальны тергальных пластин ( паранота ), и только позже в эволюции стали подвижными, сформулированными приложениями посредством вторичного набора генов ног. [ 32 ] Более поздний анализ ископаемых палеозойских нимф -крыльев обеспечивает дополнительную поддержку для слияния элементов параноты и генов Arthopodan ног. [ 44 ]
Были сделаны предложения, что крылья могли изначально развиваться для плавания на поверхности воды, как видно в некоторых каменных светах . [ 45 ] Альтернативная идея заключается в том, что она происходит от направленного воздушного скользящего спуска - явлений с предварительной полетом, найденными в какой -то Аптериготе , безвесе -таксонах сестринских сестринских таксонов для крылатых насекомых. [ 46 ] Самые ранние листовки были похожи на стрекоз с двумя наборами крыльев, прямыми мышцами полета и отсутствием способности складывать крылья над их животом . Большинство насекомых сегодня, которые развивались от этих первых летчиков, упростились до одной пары крыльев или двух пар, функционирующих в виде одной пары, и используя систему непрямых мышц полета. [ 32 ]
Естественный отбор сыграл огромную роль в совершенствовании крыльев, контрольных и сенсорных систем , а также всего, что поражает аэродинамику или кинематику . Одна примечательная черта - это кручение крыла. Большинство крыльев насекомых скручены, как и лопасти вертолета, с более высоким углом атаки у основания. Твист, как правило, составляет от 10 до 20 градусов. В дополнение к этому повороту, поверхности крыла не обязательно являются плоскими или безличными; Большинство больших насекомых имеют крыловые мембраны, искаженные и наклоненные между венами таким образом, что поперечное сечение крыльев приближается к аэродинамическому профилю . Таким образом, основная форма крыла уже способна генерировать небольшое количество подъема под нулевым углом атаки . Большинство насекомых контролируют свои крылья, регулируя наклон, жесткость и частоту хлопья крыльев с крошечными мышцами в грудной клетке (ниже). Некоторые насекомые развивали другие функции крыла, которые не выгодны для полета, но играют роль в чем -то другом, таких как спаривание или защита . [ 32 ]
| Эволюция того, как крылья отдыхают в теле для создания тела | ||
| Крылья не складываются назад (Недавние археоптера) |
Распространение поперечно (большие пузырьки) | |
| над спиной друг против друга (Damselflies, Mayflies) | ||
| Складывание (Неоптер) | ||
| Крылья не складываются (например, Stoneflies) | ||
| Складывание | фанат (например, передние крылья ос) | |
| Перекрестная складка (например, заднее крыло жука) | ||
| Складывание субъектов (например, заднее крыло ушных) | ||
Некоторые насекомые, занимающие биологические ниши, которые они делают, должны быть невероятно маневренными. Они должны найти свою еду в жестких пространствах и быть способными избежать больших хищников - или они сами могут быть хищниками, и им необходимо захватить добычу. Их маневренность с аэродинамической точки зрения обеспечивается силами высокого подъема и тяги. Типичные листовки насекомых могут достичь сил подъема в три раза их веса и горизонтального тяги в пять раз их вес. Существуют два существенно разных механизма полета насекомых, и каждый из них имеет свои собственные преимущества и недостатки - только то, что одонаты имеют более примитивный механизм полета, не означает, что они менее способны летчикам; Они определенными способами более гибкие, чем все, что развивалось впоследствии. [ 32 ]
Морфогенез
[ редактировать ]В то время как развитие крыльев у насекомых четко определено у тех, кто является членами Endopterygota , которые подвергаются полной метаморфозе ; У этих видов крыло развивается, находясь на стадии жизненного цикла насекомых насекомых. Тем не менее, насекомые, которые подвергаются неполной метаморфозе , не имеют стадии куколки, поэтому они должны иметь различный морфогенез крыла . Насекомые, такие как те, которые являются гемиметаболическими, имеют крылья, которые начинаются как починки, которые находятся под экзоскелетом, и не подвергаются воздействию до последнего возраста нимфы . [ 47 ]
Первым показателем крыловых почек является утолщение гиподермы, которое можно наблюдать у видов насекомых как на ранних стадиях эмбриона и на самых ранних стадиях жизненного цикла. Во время развития морфологических особенностей в эмбрионе или эмбриогенеза кластер клеток растут под эктодермой , которая впоследствии в развитии после того, как боковая эктодерма выросла дорсально с образованием ветряного изображного диска. Пример развития бутона крыла у личинок можно увидеть в белых бабочках ( Pieris ). Во втором возрасте гистоласт становится более заметной, что теперь образует карманную структуру. По состоянию на третьем и четвертого возраста гистоласт становится более удлиненной. Это значительно расширенное и евагированное или выступающее, часть становится тем, что становится крылом. К концу последнего возраста или пятого крыла выталкивается из кармана крыла, хотя продолжается лежать под старой личиночной кутикулой, находясь на стадии преодоления. Только после того, как бабочка находится на стадии куколки Eclosion , крыло начинает расширять и формировать свою окончательную форму. [ 47 ]
Развитие трахей крыльев начинается до формы гистоластов крыла, так как важно отметить, что они развиваются около большой трахеи . Во время четвертого возраста клетки из эпителия этой трахеи становятся значительно увеличенными, простирающиеся в полость крылового зачатка, причем каждая клетка развивает тщательно спиральную трахеолу . Каждый трахкол имеет одноклеточное происхождение и сначала находится внутриклеточной в положении; в то время как трахеи имеют многоклеточное происхождение, а просвет каждого - межклеточное положение. Развитие трахеолей, каждая из которых свернула в одной ячейке эпителия трахеи, и последующее открытие связи между трахеолями и просветом трахеи, а также рассеяние и растяжение из трахеолей, так что они достигают всех частей крыла. [ 47 ]
На более ранних этапах его развития крыланая бута не предоставляется особыми органами дыхания, такими как трахеация, как в этом отношении напоминает другие части гиподермы, из которых он все еще является частью. Гистобласт развивается вблизи большой трахеи, поперечное сечение которого показано, которое представляет участки этих частей первого, второго, третьего и четвертого возраста соответственно. В то же время трахеолы раздвигаются и простираются в пучках в формирующихся венщиках На молте, которое отмечает начало стадии куколки, они становятся функциональными. В то же время личиночные трахеолы дегенерируют; Их функция была заменена крылами Tracheae. [ 47 ]
Номенклатура
[ редактировать ]Большая часть номенклатуры орденов насекомых основана на древнегреческом словом для крыла, πτερόν ( pteron ), как суффикс -ptera .
| Научное название | лингвистический корень | Перевод научного названия | Английское имя |
|---|---|---|---|
| Анизопера | Антизо- ( анизо- ) | Неравные крылья | Дракона |
| Аптера | ἀ- ( a- ) не | Бессмысленный | Аптериготаны , теперь устарели |
| Аптеригота | Лоскут ( Pterygion маленькое крыло) [ Цитация необходима ] ἀ- ( a- ) не |
Бессмысленный | Аптериготаны |
| Coleoptera | Колледж ( Koleos , оболочка) | Закаленные крылья | Жуки |
| Дермаптер | Кожа ( дерма , кожа, кожа) | Кожаные крылья | Ушной |
| Diaphanopterodea | Прозрачный ( диафан , прозрачные или полупрозрачные) | С прозрачными крыльями | Диафаноптероиды |
| Диктиоптера | Сеть ( Diktyon , сеть) | Крылья с сеткой | Тараканы, борьбы и термиты |
| Diptera | Два- (dyo-, два ) | Два крыла | Мухи |
| Эмбиоптера | ἐ- ( en , внутри; Life Bios , жизнь) | Внутренние живые крылатые насекомые | WebSpinners |
| Эндоптеригота | В ( эндос , внутри; лоскут, маленькое крыло) | Внутри крылья | Голометаболистые насекомые |
| Эфемероптера | ἐφήμερος ( эфемерос около одного дня) | Короткие живые крылатые насекомые | Мэйфлис |
| Exopterygota | Out ( Exo , внешний) | Внешние крылья | Насекомые, которые подвергаются неполной метаморфозе (и, таким образом, имеют внешние починки крыла в качестве нимф) |
| Hemiptera | ἡμι- ( полу- , половина) | Полупавшие насекомые | Hemiptera (истинные жуки, листовины, тля и т. Д.) |
| Гетероптер | Гетеро- ( гетеро- , разные) | Разное крылатое | Истинные ошибки |
| Гомоптера | ὅμο- ( гомо- , аналогичный) | Тот же крылатый | Теперь устарел |
| Hymenoptera | О ( гименион , мембрана) | Насекомые с крыльями тонких мембран | Пчелы, осы, муравьи и т. Д. |
| Изоптера | на ( ison , равных) | Тот же крылатый | Термиты |
| Лепидоптера | Леп ( лепис , масштаб) | Масштабированные крылья | Бабочки и мотыльки |
| Lonchopteridae | Логотипы ( Lonche , Lance) | Ланс Крылья | Ланс летит |
| Мекоптера | Mikos ( Mekos , длина) | Длинные крылья | Змеиные мухи и т. Д. |
| Мегалоптера | Великолепный ( мегало- , большой) | Большие крылья | Dobsonflies, Fishflies |
| Нейроптер | Мудрость ( нейрон , вена) | VEINED WING | Кружеватели, Оулфлис, муравьев и т. Д. |
| Неоптерий | Новый ( Neos , New, Young) | Новые крылья | Включает все в настоящее время живые заказы летающих насекомых, кроме Mayflies и Dragonflies |
| Oligoneoptera | Маленький ( олигон- , мало) Новости ( NEO или новое) |
Новый с маленькими венами | Неоптер |
| Ортоптер | ὀρθο ( орто- , прямой) | Прямые крылья | Кузнечики, катидиды и сверчки |
| Palaeodictyoptera | Старый ( палайос , старый) Сеть ( диктион, означающая сеть) |
Старые крылья | Примитивные палеозоические палеоптероучные насекомые |
| Палеоптера | Старый ( палаи , старый) | Старые крылья | Mayflies, Dragonflies и несколько ископаемых заказов |
| Паранеоптера | Пара- ( пара- ) молодой ( neos , новый) |
Часть неоптери, в основном с пронзительными полосами | Истинные жуки, всы, Баркличе, трипсы |
| Phthiraptra | Φθείρ ( Phtheir , Lice) ἀ, а , а не |
Вши без крыльев | Животные всы |
| Plecoptera | Plekein ( Plekein , Fold) | Сложенные крылья | Стоунфи |
| Полиоптера | Много ( Полис , многие Новый Neos new) |
Много крыльев | Неоптер с гемиметаболизованным развитием |
| Psocoptera | Ώώχω ( psocho , чтобы втирать) | Потирать крылья | Barklice, Booklice |
| Птеригота | Лоскут ( птеригион , крыло) | Крылатые насекомые | В классе, в отличие от Apterygota, в том числе крылатые и бесцветные вторичные системы |
| Raphidioptera | ῥαφίς ( Rhaphis , игла) | Игольчатые крылья | Закуски |
| Siphonaptera | Сифон ( сифон , трубка) ἀ- или без |
Бесполучный сифон | Блох |
| Strepsiptera | Твист ( Strepsis , чтобы обернуться) | Вращающиеся или скрученные крылья | Скрученные паразиты |
| Тисаноптера | Thyssers ( thysanoi , бахромы) | Окрашенная крылатая | Трипсы |
| Трихоптера | Волосы ( трихома , волосы) | Илохлые крылья | Caddisflies |
| Зораптер | Ζωρός ( Zōros означает сильный) | Сильные крылья | Zorapterans |
| Zygoptera | Зевс ( Зего, означающая пара) | Парные крылья | Самостоятельные |
Адаптация
[ редактировать ]Вариация
[ редактировать ]Накрытия насекомых имеют основополагающее значение для выявления и классификации видов, так как нет другого набора структур при изучении насекомых более значимых. Каждый заказ и семейство насекомых имеют отличительные формы и особенности крыла. Во многих случаях даже виды можно отличить друг от друга различиями цвета и рисунка. Например, только по позиции можно идентифицировать виды, хотя и в гораздо меньшей степени. Хотя большинство насекомых складывают свои крылья, когда они находятся в состоянии покоя, дракозоны и некоторые самих отдыхают, когда их крылья распространяются горизонтально, в то время как такие группы, как Caddisflies , Stoneflies , Alderflies и кружевные крылья , держат свои крылья наклоненными на крыше над спиной. Несколько мотыльков обертывают крылья вокруг их тела, в то время как многие мухи и большинство бабочек закрывают свои крылья вместе прямо над спиной. [ 48 ]
Много раз форма крыльев коррелирует с типом полета насекомых. Лучшие насекомые, как правило, имеют длинные, тонкие крылья. У многих видов Sphingidae (Sphinx Moths) Другая, возможно, более важная корреляция, - это размер и сила мышц до скорости и силы полета. У мощно летающих насекомых крылья наиболее адаптированы для напряжений и аэродинамики полета. Вены более толстые, сильнее и ближе друг к другу к переднему краю (или «передний край») и более тонкие, но гибкие к заднему краю (или «зацепленный край»). Это делает крыло насекомых превосходно построенным аэродинамическим профилем, способным проявлять как движение , так и подъем при минимизации сопротивления . [ 48 ]
Вариация крыла может также возникнуть не только среди разных видов, но и даже среди людей в разное время. В целом, частота зависит от отношения между силой мышц крыла и сопротивлением нагрузки. Большие крылья, бабочки с легким телом могут иметь частоту ударов крыла 4–20 в секунду, тогда как мухи с маленьким крылом, и пчелы избивают свои крылья более 100 раз в секунду, а комары могут победить до 988–1046 раз второй. То же самое касается полета; Хотя, как правило, трудно оценить скорость насекомых в полете, большинство насекомых, вероятно, могут летать быстрее по своей природе, чем в контролируемых экспериментах. [ 48 ]
Coleoptera
[ редактировать ]У видов Coleoptera ( жуков ) единственными функциональными крыльями являются задние крылья. Задесчиевины длиннее, чем элитра , сложенные продольно и поперечно под элитрой. Крыло вращается вперед на основе в положение полета. Это действие распространило крыло и разворачивалось продольно и поперечно. В структуре крыла есть пружинный механизм, иногда с помощью движения живота, чтобы держать крыло в сложенном положении. Венеция крыла жука уменьшается и модифицировано из -за структуры складывания, которая включает в себя: [ 49 ]
| Перекрестная складывание на крыльях жуков | |

|
Хиндвинг, разбросан: складывающимися линии, он разделен на пять полей, которые заполняются каждое к задней части. |

|
То же крыло, наполовину сложенное: два соединения поперечного склада образуют тупой угол. Право уже в крыльях, сложенных в трех слоях. С большим разрешением, видно третья арка края крыла в первом и втором. Слева от пятой арки появляется в четвертом. |

|
То же крыло, сложенное полностью: пять полей выровнены (Elytra были удалены). |
- Коста (C) , подзажек (SCP) - на ведущем маргинальном крыле, слитый большую часть длины.
- Радиус передний (ra) - разделен на две ветви за серединой крыла.
- Радиус задний (RP) - базальная связь теряется.
- СМИ задний (MP) - ветви, длинная и сильная вена.
- Локоть спереди (CUA)
- Анальные вены (AA, AP) - вены за кубитом, разделенные анальной складкой.
У большинства видов жуков передняя пара крыльев модифицирована и склеротизируется (закалена) для формирования элитры , и они защищают деликатные задних крыльев, которые сложены под ним. [ 33 ] Элитра связана с птератораксом; называться как таковой, потому что именно там крылья связаны ( птерон, означающий «крыло» на греческом). Элитра не используется для полета , но, как правило, покрывают заднюю часть тела и защищают вторую пару крыльев ( Alae ). Элитра должна быть поднята, чтобы переместить задние крылья полета. Крылья полета жука пересекаются с венами и сложены после посадки, часто вдоль этих вен и хранятся под элитрой. У некоторых жуков способность летать была потеряна. К ним относятся несколько наземных жуков (Family Carabidae) и некоторые «настоящие долгоносики» (Family Curculionidae ), а также некоторые виды пустыни и певородов других семей. Многие из этих видов имеют два Elytra, слитые вместе, образуя твердый щит над животом. В нескольких семьях как способность летать, так и Элитра были потеряны, причем самым известным примером являются светящиеся червя семейства Phengodidae , в которых женщины имеют личистость на протяжении всей своей жизни. [ 13 ] [ 33 ]
Лепидоптера
[ редактировать ]Две пары крыльев находятся в среднем и третьем сегменте, или мезоторс и метаторакса соответственно. В более поздних родах крылья второго сегмента гораздо более выражены, однако некоторые более примитивные формы имеют одинаковый размер крыльев обоих сегментов. Крылья покрыты весами, расположенными как черепица, образуя необычайное разнообразие, которое можно увидеть в цвете. Мезоторакс развивается, чтобы иметь более мощные мышцы, чтобы продвигать мотыльку или бабочку через воздух, при этом крыло упомянутого сегмента имеет более сильную структуру вен. [ 33 ] : 560 Крупнейшее суперсемейство, Noctuidae , имеет модифицированные крылья, чтобы выступать в роли баллов или органов слуха [ 50 ] Модификации в Venation включают: [ 49 ]
- Коста (C) - не найдено в бабочках.
- Subcosta (SC) + Radius 1 (SC + R1) - на ведущем краевом крыле, слитом или очень близком для большей части длины, в хэндвинге, слитых и хорошо развитых в области плеча, Subcosta никогда не разветвляется в бабочке.
- Радиус (R2-R5) -Радиус делится на ветви за серединой крыла до пяти ветвей в Papilionidae. На переднем крыте, последний r преследуется во всех бабочках, за исключением того, что Hesperiidae отделяется.
- Радиус -сектор (RS) - в задних крылах.
- СМИ (M1-M3) -базальная секция была потеряна.
- Кубит передний (CUA1-CUA2) -секция чашки была потеряна.
- Анальные вены (A, 1a+2a, 3a) - либо одна вена A, либо две вены 1a+2a, 3a.
- Вена плечевой кости - задним ход большинства бабочек имеет плечевую вену, за исключением Lycaenidae, существует расширение области плечевой области, которая перекрывается с передним крылом. Вена плечевой кости укрепила перекрывающуюся площадь задних крыльев так, чтобы два крыла были лучше.
Крылья, части головки грудной клетки и брюшной полости Лепидоптера покрыты мельчайшими масштабами , из которых функционирует порядок «Lepidoptera» вызывает свои имена, слово «Lepteron» в древнегреческом значении «шкала». Большинство масштабов являются пластинчарными или похожими на лезвие и прикреплены с помощью цветоножки, в то время как другие формы могут быть похожими на волосы или специализированы как вторичные сексуальные характеристики. [ 51 ] Просвет или поверхность ламеллы, имеет сложную структуру. Он дает цвет либо из-за пигментных цветов, содержащихся в ее трехмерной структуре . [ 52 ] Шкалы предоставляют ряд функций, которые включают изоляцию, терморегуляцию, помощь в скольжении, среди прочего, наиболее важным из которых является большое разнообразие ярких или нечетких моделей, которые они предоставляют, которые помогают организму защитить себя путем камуфляжа , мимики и искать товарищи. [ 51 ]
Одоната
[ редактировать ]Виды Odonata (Sewhe Selflies и Dragonflies) имеют две пары крыльев, которые примерно равны по размеру и форме и имеют четкие по цвету. Есть пять, если R+M считается 1, основная вена стебли на Dragonfly и Damselfly Wings, а крыловые вены сливаются на их основаниях, а крылья не могут быть сложены над телом в состоянии покоя, что также включает в себя: [ 49 ]
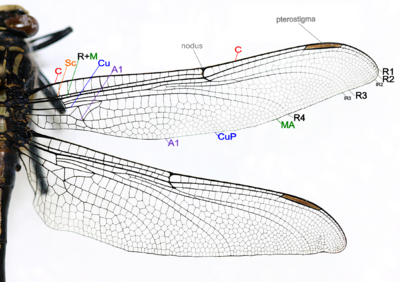

- Коста (C) - на переднем крае крыла, сильного и маргинального, распространяется на вершину крыла.
- Subcosta (SC) - вторая продольная вена, она неразветвлена, соединяет C в Nodus.
- Радиус и среда (R+M) -третья и четвертая продольная вена, самая сильная вена на крыле, с ветвями, R1-R4, достигают края крыла, передняя среда (MA) также достигает края крыла. IR2 и IR3 являются интеркалярными венами за R2 и R3 соответственно.
- Cubitus (Cu) - Пятая продольная вена, задний кубит (чашка) не расстановлен и достигает края крыла.
- Анальные вены (A1) - неразветвленные вены за кубиком.
- Сформируется узд , где вторая главная вена встречается с передним краем крыла. Черная птеростигма переносится возле кончика крыла.
Основные вены и Crossveins образуют шаблон венения крыла. Схемы венеции разные у разных видов. Там может быть очень многочисленные Crossveins или, скорее, мало. Австралийские крылья Damselfly's Plattling - один из немногих моделей вен. Схема венеции полезна для идентификации видов. [ 49 ] Почти все анизоптера оседают с крыльями, проведенными вбок или слегка вниз, однако большинство зигоптеров оседают с крыльями, удерживаемыми вместе, дорсальными поверхностями. Грумовая клетка Zygoptera настолько наклонена, что, когда он держатся таким образом, крылья аккуратно подходят вдоль верхней части живота. Они, кажется, не удерживаются прямо, как у бабочек или майков. В нескольких зигоптеранах крылья удерживаются горизонтально в состоянии покоя, а в одном роде анизоптера (например, Cordulephya , Corduliidae ) крылья удерживаются в типичной плодоподобившей позиции. Взрослые виды обладают двумя парами равных или подравленных крыльев. Похоже, есть только пять основных стеблей вен. Сформируется узд , где вторая главная вена (подкоста) соответствует переднему краю крыла. В большинстве семей у кончика крыла переносится заметная птеростигма. Идентификация как одоната может быть основана на венеции. Единственная, вероятно, путаница - с некоторыми кружевными крышками (порядок нейроптер), которые имеют много перекрестных вещей в крыльях. До первых лет 20 -го века Одоната часто рассматривались как связанные с кружевами и получили порядковое название Paraneuroptera, но любое сходство между этими двумя порядками является полностью поверхностным. В Anisoptera задних крыльев шире, чем передвижение, и в обоих крыльях кроссвийн делит дискоидальную клетку на треугольник и супертрианг. [ 53 ]
Ортоптер
[ редактировать ]
Виды ортоптер (кузнечики и сверчки) имеют передние уколы, которые являются жесткими непрозрачными тегминами, узкими, которые обычно покрывают задних крыльев и живот в состоянии покоя. Заджественные крышки мембраны и свернуты в фанатах, которые включают в себя следующее венеция: [ 49 ]
- Коста (C) - на ведущем маргинальном положении переднего крыла и задних крыльев, неразветвленный.
- Подкоста (SC) - Вторая продольная вена, неразветвленная.
- Радиус (R) - третья продольная вена, разветвленная до рупий в лесорубке и задних крылах.
- Среда передняя (MA) - Четвертая продольная вена, разветвленная в базальной части в качестве задней среды (MP).
- Cubitus (Cu) - Пятая продольная вена, на переднем крыле и задних крылах, разделившихся возле основания крыла на разветвленную CUA и неразветвленную чашку.
- Анальные вены (а) - вены за кубиком, неразветвленные, две в лесорубке, многие в Хиндвинге.
Фасматодея
[ редактировать ]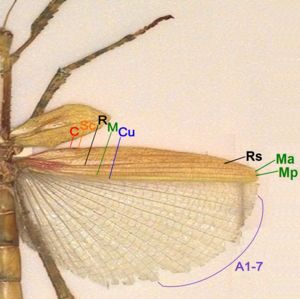
- Коста (C) - на ведущем маргинальном положении задних крыльев, неразветвленных, отсутствующих при переднем крылью.
- Подкоста (SC) - Вторая продольная вена, неразветвленная.
- Радиус (R) - третья продольная вена, разветвленная до рупий в задних крылах, не распределенная при переднем крыле.
- Среда передняя (MA) - Четвертая продольная вена, разветвленная в базальной части в качестве задней среды (MP).
- Локоть (Cu) - Пятая продольная вена, неразветвленная.
- Анальные вены (а) -вены за кубиком, неразветвленные, две в лесорубке, многие из Хиндвинг 1A-7A в одной группе, а остальные в другой группе.
У насекомых у насекомых есть жесткие, непрозрачные тегмины, короткие и покрывают только базовую часть задних крыльев. Задеские вины от Коста до Кубитуса жесткие и непрозрачные, как и передние крылья. Большая анальная область мембрана и сложена по вентиляторам. В жилах насекомых нет или очень мало ветвящихся вен. [ 49 ]
Дермаптер
[ редактировать ]| Развертывание крыла уш | |

|
Передние и задние крылья в состоянии покоя: переднее крыло покрывает большую часть задних крыльев, и только совместные проекты в виде четверти круга вперед с центральным белым пятном под передним крылом. На правой части переднего крыла открывается справа (синяя стрелка), которая с этой точки зрения кажется более узкой, чем с задним крылом, все еще полностью сложенным. Полем |

|
Переднее крыло открыто слева (синяя стрелка) с удаленной правой стороной переднего крыла; Хиндвинг наполовину открыт. С большим разрешением показано множественное складывание, напоминающее вентилятор, который параллельно линиям B и C. Стрелка указывает на точку E, где вентилятор снова закрыт, сложив на 180 °. Полем |
такие как дермаптера ( ушной писатели ), ортоптеры ( кузнечики , сверчки , мантодея ( мантида ) и блюттодиа ( ) кожа ) тараканы имеют Другие приказы , жесткую молитва Элитра ) , или псевдоэлитрон . [ 13 ]
Hemiptera
[ редактировать ]В Hemiptera (истинные ошибки) передние крылья могут быть закалены, хотя и в меньшей степени, чем в жуках. Например, передняя часть передних крыльев вонючих жуков затвердевает, в то время как задняя часть мембрана. Они называются Hemelytron (пл. Хемэлитра ). Они встречаются только в подряд -гетероптерах ; Крылья гомоптеры , такие как цикада , как правило, полностью мембраны. И Большинство видов похожи на стекло, хотя некоторые непрозрачны. Цикады не хорошие листовки, и большинство летают всего несколько секунд. При лете, передние крылья и задних крыльев соединяются с помощью палочной муфты вдоль задних костей и края переднего крыла. Большинство видов имеют базовое венецию, как показано на следующей картине. [ 49 ]
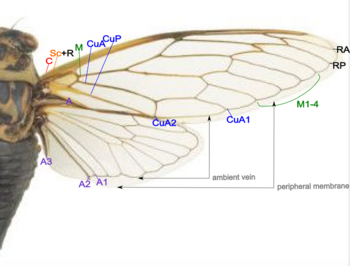
- Коста (C) - На ведущем крае крыла в лесу распространяется узел и лежит близко к SC+R.
- Subcosta + Radius (SC + R) - в переднем обонялке SC и R, объединенные вместе с узлом. Радиальный сектор (RS) возникает вблизи узла и небрызги.
- Радиус передний (ra)
- Радиус задний (RP)
- СМИ (M) - филиалы до M1 до M4.
- Предыдущий Cubitus (CUA) - ветви к Qua1 и Qua2.
- Локоть задний (чашка) - Unbranches.
- Анальные вены (A) - вены за кубитом, 1a и 2a слиты в переднем крыле, чашка и 2a складываются.
Также обратите внимание, что есть окружающие вены и периферические мембраны на краю обоих крыльев.
Diptera
[ редактировать ]В Diptera ( истинные мухи ) есть только одна пара функциональных крыльев, причем задняя пара крыльев сводится к заповедникам , которые помогают муху ощутить его ориентацию и движение, а также улучшать баланс, действуя аналогично гироскопам Полем У Calyptratae самая задняя часть крыльев модифицирована в несколько утолщенных клапанов, называемых калиптерами , которые покрывают заповедники. [ 49 ]

- Коста (C) - не найдено в Diptera.
- Subcosta (SC) - стала ведущей крыломной веной, неразветвленной.
- Радиус (R) -разветвлен до R1-R5.
- СМИ (M) -разветвляется до M1-M4.
- Кубит передний (CUA) - неразветвленная чашка уменьшается в Diptera. Некоторые виды CuA и 1a разделены, некоторые виды встречаются при достижении края крыла, некоторые виды слиты.
- Анальные вены (A) - присутствуют только две анальные вены 1a и 2a, 2a не отличается у некоторых видов.
- Дисковая клетка (DC) - четко определенная у большинства видов.
Блюттодея
[ редактировать ]Виды блюттодии ( тараканы ) имеют передние крылья, также известны как тегмен, которая более или менее склеротизирована. Он используется в полете, а также в форме защиты мембранных задних крыльев. Вены задних крыльев примерно так же, как переднее крыло, но с большой анальной долей, сложенной в покое между чашкой и 1А. Анальная доля обычно сложена по вентилятором образом. [ 49 ]

- Коста (C) - на переднем крае крыла.
- Подкоста (SC) - вторая продольная вена, она относительно короткая.
- Радиус (R) - третья продольная вена, со многими пектинационными ветвями.
- СМИ (M) - Четвертая продольная вена, достичь края крыла.
- Кубит передний (CUA) - Пятая продольная вена, с дихотомическими ветвями занимают большую часть тегменов.
- Cubitus задний (чашка) - неразветвленный, изогнутый и достигать края крыла.
- Анальные вены (а) - вены за кубитом.
Hymenoptera
[ редактировать ]| Пример продольного складывания в ос (Vespidae) | |

|
Основная линия сгиба переднего крыла рассматривалась на полпути как яркая горизонтальная линия. Часть крыла, которая стоит за этой линией, повернута назад. Узкая полоса на переднем крае крыла находится перед первым прочным проводом, сложенным вперед и вниз. |

|
Таким образом, в положении покоя внешняя подкладка образует жесткий внешний край крыла, который защищает стороны живота как амортизатор. Заднее крыло покрыто в значительной степени передвижением. |
Взрослые Hymenoptera , в том числе пилаты, осы, пчел и муравья, не работающие, имеют две пары мембранных крыльев. [ 49 ]
- Коста (C) - не найдено в Hymenoptera.
- Подкоста (SC) - неразветвленный.
- Радиус (R) -разветвлен до R1-R5.
- СМИ (M) - M неразветвлен, в переднем состоянии M сливается с RS для части ее длины.
- Cubitus (CUA) - неразветвленная, чашка отсутствует в Hymenoptera.
- Анальные вены (A) - присутствуют только две анальные вены 1a и 2a, 2a не отличается у некоторых видов.
- Крылополирование -ряд крючков на переднем крае задних крыльев задействует задним краем переднего крыла, сильно соединяет крылья в полете.
- Линия складывания крыла - некоторые виды, включая Vespidae, передние крылья продольно сложены вдоль «линии складывания крыла» в состоянии покоя.
- Птеростигма - присутствует для некоторых видов.
Передний запас задних крышек имеет несколько подключенных щетинок, или « хамули », которые фиксируются на переднем крыте, держа их вместе. Меньшие виды могут иметь только два или три хамули с каждой стороны, но самые большие осы могут иметь значительное число, что удерживает крылья, особенно плотно. У крыльев гименоптерана относительно мало вен по сравнению со многими другими насекомыми, особенно у меньших видов. [ 13 ]
Другие семьи
[ редактировать ]Термиты являются относительно плохими листовками и легко взорваны по ветру с скоростью ветра менее 2 км/ч, проливая свои крылья вскоре после приземления на приемлемого участка, где они спариваются и пытаются сформировать гнездо во влажной древесине или земле. [ 54 ] Крылья большинства термитов имеют три тяжелые вены вдоль базальной части переднего края переднего крыла, а кроссвеины возле кончика крыла имеют угловые, изготовляя трапециевидные клетки. Хотя подземные термитные крылья имеют только две основные вены вдоль переднего края переднего крыла, а поперечные вены в направлении крыла перпендикулярно этим венам, делая квадратные и прямоугольные клетки. [ 55 ]
Виды тисаноптеров ( трипс ), Ptiliidae и других летающих микроинсектов имеют стройную переднюю и задних крыльев с длинными полосами волос, называемых полосами, также называемыми Ptiloptery. [ 56 ] В то время как виды трихоптера ( Caddisfly ) имеют волосатые крылья с передними и задними крыльями, одетыми с щетинками. [ 13 ]
У мужчин Strepsiptera также есть заповедники , которые развивались от передних крыльев вместо задних крыльев. Это означает, что только их задних крыльев функционируют при полете, в отличие от Diptera , которые имеют функциональные передники и недостатки для задних крыльев. Кроме того, задних крыльев у мужчин Coccidae снижаются до заповедников (или отсутствуют). [ 57 ]
Смотрите также
[ редактировать ]- Придаток
- Comstock-Needham System
- Крыло
- Роботы, вдохновленные насекомыми: робоби , делли
Примечания
[ редактировать ]- ^ Крэмптон, Г. (1916). «Филогенетическое происхождение и природа крыльев насекомых в соответствии с паранотальной теорией». Журнал нью -йоркского энтомологического общества . 24 (1): 1–39. JSTOR 25003692 .
- ^ Jump up to: а беременный Росс, Эндрю (2017). «Эволюция насекомых: происхождение крыльев» . Текущая биология . 27 (3): R113 - R115. doi : 10.1016/j.cub.2016.12.014 . PMID 28171756 - через Web of Science.
- ^ Jump up to: а беременный Авероф, Михалис и С.М. Коэн. (1997). «Эволюционное происхождение крыльев насекомых из наследственных жабра». Природа . 385 (6617): 627–630. Bibcode : 1997natur.385..627a . doi : 10.1038/385627A0 . PMID 9024659 . S2CID 4257270 - через Web of Science.
{{cite journal}}: Cs1 maint: несколько имен: список авторов ( ссылка ) - ^ Grodnitsky, Dmitry, L. (1999). Форма и функция крыльев насекомых: эволюция биологических структур . С. 82–83.
{{cite book}}: Cs1 maint: несколько имен: список авторов ( ссылка ) - ^ Jump up to: а беременный в Александр, Дэвид, Э. (2015). На крыле: насекомые, птерозавры, птицы, летучие мыши и эволюция полета животных . Издательство Оксфордского университета. С. 74–101.
{{cite book}}: Cs1 maint: несколько имен: список авторов ( ссылка ) - ^ Haug, Joachim, C. Haug., И RJ Garwood. (2016). «Эволюция крыльев насекомых и развития - новые детали от палеозоических нимф» . Биологические обзоры . 91 (1): 53–69. doi : 10.1111/brv.12159 . PMID 25400084 . S2CID 21031689 .
{{cite journal}}: Cs1 maint: несколько имен: список авторов ( ссылка ) - ^ Альмуди, Изабель; Визуэта, Джоэл; Уайетт, Кристофер доктор; де Мендоса, Алекс; Марлетаз, Фердинанд; FIRBAS, Panos N.; Феуда, Роберто; Масеро, Джулио; Медина, Патриция; Alcaina-Caro, Ana; Круз, Фернандо (2020). «Геномная адаптация к водной и воздушной жизни у Мэйфлс и происхождение крыльев насекомых» . Природная связь . 11 (1): 2631. Bibcode : 2020ntco..11.2631a . doi : 10.1038/s41467-020-16284-8 . ISSN 2041-1723 . PMC 7250882 . PMID 32457347 .
- ^ Брюс, Хизер, Н.Х. Патель. (2020). «Нокаут генов с паттерном ракообразных ног предполагает, что крылья насекомых и стены тела развивались из древних сегментов ног». Природа экология и эволюция . 4 (12): 1703–1712. Bibcode : 2020natee ... 4.1703b . doi : 10.1038/s41559-020-01349-0 . PMID 33262517 . S2CID 227253368 .
{{cite journal}}: Cs1 maint: несколько имен: список авторов ( ссылка ) - ^ Кларк-Хатчел, Кортни (2013). «Понимание происхождения крыла насекомых, предоставленное функциональным анализом рудимента в красно -муке, Tribolium Castaneum» . Труды Национальной академии наук Соединенных Штатов Америки . 110 (42): 16951–16956. BIBCODE : 2013PNAS..11016951C . doi : 10.1073/pnas.1304332110 . PMC 3801059 . PMID 24085843 .
- ^ Prokop, Jakub, Pecharová, M., Nel, A., Hörnschemeyer, T., Krzemińska, E., Krzemiński, W. & Engel, M (2017). «Палеозойские нимфальные подушки поддерживают двойную модель происхождения крыла насекомых» . Текущая биология . 27 (2): 263–269. doi : 10.1016/j.cub.2016.11.021 . PMID 28089512 .
{{cite journal}}: Cs1 maint: несколько имен: список авторов ( ссылка ) - ^ Jump up to: а беременный в дюймовый и фон Чепмен, RF (1998). Насекомые: структура и функция (4 -е изд.). Кембридж, Нью -Йорк: издательство Кембриджского университета. ISBN 0-521-57048-4 .
- ^ Jump up to: а беременный Гиллиот, Седрик (август 1995 г.). Энтомология (2 изд.). Springer-Verlag New York, LLC. ISBN 0-306-44967-6 .
- ^ Jump up to: а беременный в дюймовый и Мейер, Джон Р. (5 января 2007 г.). «Внешняя анатомия: крылья» . Кафедра энтомологии, Университет штата Северная Каролина. Архивировано из оригинала 16 июля 2011 года . Получено 2011-03-21 .
- ^ Jump up to: а беременный в дюймовый Гуллан, PJ; Cranston, PS (2004). Насекомые: схема энтомологии Великобритания: Blackwell Publishing. П. 42 ISBN 1-4051-1113-5 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л м не а Snodgrass, Re (декабрь 1993 г.). Принципы морфологии насекомых . Корнелл Univ Press. ISBN 0-8014-8125-2 .
- ^ Spieth, HT (1932). «Новый метод изучения крыльев вен майки и некоторых результатов от него (эфемерида)» (PDF) . Энтомологические новости . Архивировано из оригинала (PDF) 2011-09-30.
- ^ «Объяснение терминов, используемых в энтомологии» . www.gutenberg.org . Получено 2024-01-08 .
- ^ «Определение термина» . www.merriam-webster.com . Получено 2024-01-08 .
- ^ Jump up to: а беременный Knospe, Carl R. (осень 1998). «Механизмы полета насекомых: анатомия и кинематика» (PDF) . Механическая и аэрокосмическая инженерия, Университет Вирджинии.
- ^ Вейс-Фог, Т (июль 1963 г.). «Диффузия в мышцах крыла насекомых, наиболее активной ткани» . J Exp Biol . 41 (2): 229–256. doi : 10.1242/jeb.41.2.229 . PMID 14187297 .
- ^ Тигс, Оу (февраль 1954 г.). «Летные мышцы анатомии и гистологии насекомых-с некоторыми наблюдениями о структуре полосатых мышц в целом» . Философские транзакции Королевского общества Лондона. Серия B, биологические науки . 238 (656): 221–348. Bibcode : 1955rsptb.238..221t . doi : 10.1098/rstb.1955.0001 . JSTOR 3739600 .
- ^ Он, Чжэ; Ло, Йихен; Шан, Xueying; Солнце, Дженнифер с.; Карлсон, Джон Р. (21 мая 2019 г.). «Хемосенсорная сенсорная сенсация крыла Drosophila экспрессирует рецептор -ионотроп -феромон -рецептор -кандидата» . PLOS Биология . 17 (5): E2006619. doi : 10.1371/journal.pbio.2006619 .
- ^ Valmalette, JC, Raad, H., Qiu, N., Ohara, S., Capovilla, M. and Robichon, A., 2015. Нано-архитектура вкусовых хемосенсорных щетин и трахеи у Drosophila Wings. Scientific Reports, 5 (1), pp.1-11.
- ^ Dinges, Gf, Chockley, AS, Bockemühl, T., Ito, K., Blanke, A. и Büschges, A., 2021. Расположение и расположение Campaniform Sensilla у Drosophila Melanogaster. Журнал сравнительной неврологии, 529 (4), стр.905-925.
- ^ Field, LH and Matheson, T., 1998. Хордотональные органы насекомых. В достижениях в области физиологии насекомых (том 27, с. 1-228). Академическая пресса.
- ^ Wolf, H., 1993. The Lacust Tegula: значение для генерации ритма полета, управления движением крыла и аэродинамической силы. Журнал экспериментальной биологии, 182 (1), с.229-253.
- ^ Zhang, N. и Simpson, JH, 2022. Пара комиссуральных командных нейронов индуцирует груминг Drosophila Wing. Iscience, 25 (2), p.103792.
- ^ Jump up to: а беременный Чепмен, AD (2006). Количество живых видов в Австралии и мире . Канберра: Австралийское исследование биологических ресурсов . С. 60pp. ISBN 978-0-642-56850-2 Полем Архивировано с оригинала на 2009-05-19 . Получено 2012-06-18 .
- ^ Jump up to: а беременный Ван, З. Джейн (2005). «Расширение полета насекомых» (PDF) . Ежегодный обзор механики жидкости . 37 (1). Ежегодные обзоры: 183–210. BIBCODE : 2005ANRFM..37..183W . doi : 10.1146/annurev.fluid.36.050802.121940 .
- ^ Jump up to: а беременный Сэйн, Санджай П. (2003). «Аэродинамика полета насекомых» (PDF) . Журнал экспериментальной биологии . 206 (23): 4191–4208. doi : 10.1242/jeb.00663 . PMID 14581590 . S2CID 17453426 .
- ^ Давидовит, Пол (2008). Физика в биологии и медицине . Академическая пресса. С. 78–79. ISBN 978-0-12-369411-9 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Гримальди, Дэвид; Энгель, Майкл С. (2005). Эволюция насекомых . Нью -Йорк, Нью -Йорк: издательство Кембриджского университета.
- ^ Jump up to: а беременный в дюймовый и Пауэлл, Джерри А. (2009). "Coleoptera" . В Реш, Винсент Х.; Cardé, Ring T. (ред.). Энциклопедия насекомых (2 (иллюстрировано) изд.). Академическая пресса. п. 1132. ISBN 978-0-12-374144-8 .
- ^ Майкл С. Энгель; Дэвид А. Гримальди (2004). «Новый свет проливает на самого старого насекомого». Природа . 427 (6975): 627–630. Bibcode : 2004natur.427..627e . doi : 10.1038/nature02291 . PMID 14961119 . S2CID 4431205 .
- ^ Хауг, Каролин; Haug, Joachim T. (2017-05-30). "Предполагаемое старое летающее насекомое: скорее всего, бесчисленное множество?" Полем ПЕРЕЙ . 5 : E3402. doi : 10.7717/peerj.3402 . ISSN 2167-8359 . PMC 5452959 . PMID 28584727 .
- ^ Grzimek HC Bernhard (1975) Грзимек Энциклопедия Живота Грзимека Том 22 насекомых. Ван Ностренд Рейнхольд Ко. Нью -Йорк.
- ^ Jump up to: а беременный Рик Эф Кукалова-Пек Дж. (1984). «Новая интерпретация венения дракона, основанная на ранних верхних карбоновых окаменелостях из Аргентины (Insecta: Odonatoida и основные состояния характера в крылах Pterygote.)». Может. J. Zool . 62 (6): 1150–1160. doi : 10.1139/Z84-166 .
- ^ Wakeling JM Ellington CP (1997). «Драконфляйская полет III поднятия и требования к мощности» . Журнал экспериментальной биологии . 200 (Pt 3): 583–600 (589). doi : 10.1242/jeb.200.3.583 . PMID 9318294 .
- ^ Мацуда Р. (1970). «Морфология и эволюция грудной клетки насекомых». Мем Энтомол. Соц Может . 102 (76): 1–431. doi : 10.4039/entm10276fv .
- ^ VA Blogerov; Эд Лукашевич; MB Mostovski (2002). "Заказ Diptera Linné, 1758. Истинные летат" в AP Rassyn; DLJ Quicke (ред.). История насекомых Kluwer Academic Bublishers ISBN 1-4020-0026-х .
- ^ Гегенбаур, Карл (1870). Основные особенности сравнительной анатомии . В. Энгельманн.
- ^ Trueman JWH (1990), Комментарий: Эволюция крыльев насекомых: Exte Excite Plus модель канадского журнала зоологии
- ^ Staniczek, Ah; Bechly, G. & Godunko, RJ (2011). «Coxoplectoptera, новый ископаемый порядок палеоптеров (членистоногие: инсекта), с комментариями о филогениях группы маяфей ствола (эфемероптера)» (PDF) . Систематика насекомых и эволюция . 42 (2): 101–138. doi : 10.1163/187631211x578406 . S2CID 4986911 . Архивировано из оригинала (PDF) 2011-10-03.
{{cite journal}}: Cs1 maint: несколько имен: список авторов ( ссылка ) - ^ Прокоп, Джакуб; Печарова, Мартина; Нел, Андре; Hörnschemeyer, Thomas; Креминьска, Эва; Креминьски, Вислав; Энгель, Майкл С. (январь 2017 г.). «Палеозойские нимфальные подушки поддерживают двойную модель происхождения крыла насекомых» . Текущая биология . 27 (2): 263–269. doi : 10.1016/j.cub.2016.11.021 . PMID 28089512 .
- ^ Адриан Л.Р. Томас; Р. Оке Норберг (1996). «Смириться с поверхностью - происхождение полета у насекомых?». Тенденции в экологии и эволюции . 11 (5): 187–188. doi : 10.1016/0169-5347 (96) 30022-0 . PMID 21237803 .
- ^ Yanoviak SP, Kaspari M, Dudley R (2009). «Скоростные гексапод и происхождение воздушного поведения насекомых» . Биологические письма . 5 (4): 510–2. doi : 10.1098/rsbl.2009.0029 . PMC 2781901 . PMID 19324632 .
- ^ Jump up to: а беременный в дюймовый Х. Комсток, Генри (1918). Крылья насекомых . Итака, Нью -Йорк: издательская компания Comstack. п. 114
- ^ Jump up to: а беременный в «Обсекие крылья в целом» . Аэродинамика насекомых . Cislunar Aerospace. 1997 . Получено 28 марта 2011 года .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж Чу, Питер (9 мая 2009 г.). «Крылья насекомых» . Брисбенские насекомые и пауки . Получено 2011-03-21 .
- ^ Скобл, MJ. (1992). Lepidoptera: форма, функция и разнообразие . Оксфордский Univ. Нажимать. ISBN 978-1-4020-6242-1 .
- ^ Jump up to: а беременный Скобл (1995). сечения Шкалы , (стр. 63 - 66).
- ^ Vukusic, P. (2006). «Структурный цвет в lepidoptera» (PDF) . Текущая биология . 16 (16): R621–3. doi : 10.1016/j.cub.2006.07.040 . PMID 16920604 . S2CID 52828850 . Получено 11 ноября 2010 года .
- ^ Trueman, John WH; Ричард Дж. Роу (16 октября 2009 г.). «Одоната. Драконы и Самодельные» . Tolweb.org . Получено 2011-03-21 .
- ^ Абэ Т., Бигнелл де; Хигаши М. (2000). Термиты: эволюция, социальность, симбиозы, экология, эколаб . Kluwer Academic Publishers. ISBN 0-7923-6361-2 .
- ^ "Термит" . Техасская служба расширения агроллы. Архивировано из оригинала 2011-04-13.
- ^ Полилов, Алексей А.; Решетникова, Наталья I.; Петров, Pyotr N.; Фарисенков, Сергей Э. (январь 2019 г.). «Морфология крыла у Featherwing Beetles (Coleoptera: Ptiliidae): особенности, связанные с миниатюризацией и анализом функционального масштабирования» . Членистоногие структура и развитие . Специальный выпуск: миниатюризация в Панартроподе. 48 : 56–70. Bibcode : 2019artsd..48 ... 56p . doi : 10.1016/j.asd.2019.01.003 .
- ^ Камачо, Эрнесто Робайо; Чонг, Джуан-Хорнг (2015). «Общая биология и текущие подходы к лечению вредителей мягкого масштаба (Hemiptera: Coccidae)» (PDF) . Журнал интегрированного управления вредителями . 6 (1): 17. doi : 10.1093/jipm/pmv016 . ISSN 2155-7470 . PMC 4725186 . PMID 26823990 .
Ссылки
[ редактировать ]- Триплхорн, Чарльз А.; Джонсон Норман Ф. (2005). Боррор и Делонг введение в изучение насекомых (7 -е изд.). Томсон Брукс/Коул. ISBN 0-03-096835-6 .
Внешние ссылки
[ редактировать ]- Курс университета Брисбена по крылам насекомых
- Курс Университета штата Северная Каролина по крылам насекомых
- Рисунки крыла насекомых
| До |  | |
|---|---|---|
| Конечности | ||
| Крылья | ||
| Эволюция | ||
| Связанный | ||