Фосфорилирование белка
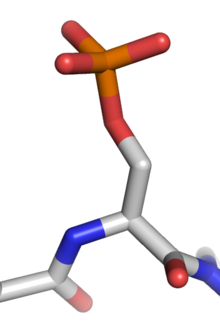

Фосфорилирование белка обратимую посттрансляционную модификацию белков , в которой аминокислотный остаток фосфорилируется протеинкиназой представляет собой путем добавления ковалентно связанной фосфатной группы. Фосфорилирование изменяет структурную конформацию белка, заставляя его стать активированным, деактивированным или иным образом модифицировать его функцию. [ 1 ] Приблизительно 13 000 белков человека имеют сайты, которые фосфорилируются. [ 2 ]
Обратная реакция фосфорилирования называется дефосфорилированием и катализируется белками фосфатаз . Протеинкиназы и фосфатазы работают независимо и в балансе, чтобы регулировать функцию белков. [ 3 ]
Аминокислоты чаще всего фосфорилируются серином , треонином , тирозином и гистидина . [ 4 ] [ 5 ] Эти фосфорилирования играют важную и хорошо охарактеризованную роль в сигнальных путях и метаболизме. Однако другие аминокислоты также могут быть фосфорилированы посттрансляционно, включая аргинин , лизин , аспарагиновую кислоту , глутаминовую кислоту и цистеин , и эти фосфорилированные аминокислоты были идентифицированы в экстрактах клеток человека и фиксированных человеческих клетках с использованием комбинации антител -Са основанный анализ (для PHI) и масс -спектрометрия (для всех других аминокислот). [ 5 ] [ 6 ] [ 7 ] [ 8 ]
Фосфорилирование белка впервые было сообщено в 1906 году Фибом Левен в Институте медицинских исследований Рокфеллера с открытием фосфорилированного вителлина . [ 9 ] Тем не менее, прошло почти 50 лет, пока не было обнаружено ферментативное фосфорилирование белков протеинкиназами. [ 10 ]
История
[ редактировать ]В 1906 году Фибус Левен в Институте медицинских исследований Рокфеллера идентифицировал фосфат в белке вителлин (фосвитин) [ 9 ] и к 1933 году обнаружил фосфосерин в казеине с Фриц Липманн. [ 11 ] Тем не менее, потребовалось еще 20 лет, прежде чем Юджин П. Кеннеди описал первое «ферментативное фосфорилирование белков». [ 10 ] Первый фермент фосфорилазы был обнаружен Карлом и Герти Кори в конце 1930 -х годов. Карл и Герти Кори обнаружили две формы гликогена фосфорилазы , которые они назвали A и B, но не правильно понимали механизм B -формы к обращению формы. Взаимосвязь фосфорилазы В к фосфорилазе А была позже описана Эдмондом Фишером и Эдвином Кребсом , а также Возилаитом и Сазерлендом , включая механизм фосфорилирования/дефосфорилирования. [ 12 ] Было обнаружено, что фермент, названный фосфорилазе-киназой и Mg-ATP, необходимы для фосфорилирования гликогена фосфорилазы, помогая в переносе γ-фосфорильной группы АТФ к сериновому остаткам на фосфорилазе B. Белковая фосфатаза 1 способна катализировать дефосфорилирование фосфорилированных ферментов путем удаления фосфатной группы. Эрл Сазерленд объяснил в 1950 году, что активность фосфорилазы была увеличена и, таким образом, гликогенолиз стимулировал, когда срезы печени инкубировали с адреналином и глюкагоном. Фосфорилирование считалось специфическим контрольным механизмом для одного метаболического пути до 1970 -х годов, когда Лестер Рид обнаружил, что митохондриальный комплекс пируватдегидрогеназы инактивирован путем фосфорилирования. Также в 1970 -х годах термин многосайт фосфорилирование было придумано в ответ на обнаружение белков, которые фосфорилируются на двух или более остатках двумя или более киназами. В 1975 году было показано, что цАМФ-зависимые белки киназы фосфорилируют сериновые остатки на специфических аминокислотных мотивах последовательности. Рэй Эриксон обнаружил, что V-SRC была киназой, и Тони Хантер обнаружил, что фосфорилированные остатки тирозина V-SRC на белках в 1970-х годах. [ 13 ] В начале 1980 года была определена аминокислотная последовательность первой протеинкиназы, которая помогла генетикам понять функции регуляторных генов. В конце 1980 -х и начале 1990 -х годов была очищена первая белковая тирозинфосфатаза (PTP1B), а также обнаружено, а также клонирование як -киназ , что привело к тому, что многие в научном сообществе назвали 1990 -е годы как десятилетие протеинкиназы каскады. [ 14 ] [ 15 ] Эдмонд Фишер и Эдвин Кребс были удостоены Нобелевской премии в 1992 году «за их открытия, касающиеся обратимого фосфорилирования белка как биологического регуляторного механизма». [ 16 ]
Избыток
[ редактировать ]Обратимое фосфорилирование белков в изобилии как в прокариотических , так и в большей степени в эукариотических организмах. [ 17 ] [ 18 ] [ 19 ] [ 20 ] Например, у бактерий 5-10% всех белков считается фосфорилированным. [ 21 ] [ 22 ] Напротив, подсчитано, что одна треть всех белков человека фосфорилируется в любой момент времени, с 230 000, 156 000 и 40 000 уникальных мест фосфорилирования, существующих у человека, мыши и дрожжей, соответственно. [ 2 ] У дрожжей около 120 киназ (из ~ 6000 белков) приводят к 8 814 известным регулируемым событиям фосфорилирования, генерируя около 3600 фосфопротеинов (около 60% всех дрожжевых белков). [ 23 ] [ 24 ] Следовательно, фосфорилирование является универсальным регуляторным механизмом, который влияет на большую часть белков. Даже если белок не фосфорилируется сам, его взаимодействие с другими белками может регулироваться фосфорилированием этих взаимодействующих белков.
Механизмы и функции фосфорилирования
[ редактировать ]Фосфорилирование вводит заряженную и гидрофильную группу в боковой цепи аминокислот, возможно, изменяя структуру белка, изменяя взаимодействие с близлежащими аминокислотами. Некоторые белки, такие как p53, содержат несколько сайтов фосфорилирования, облегчающие сложную многоуровневую регуляцию. Из -за легкость, с которой белки могут быть фосфорилированы и дефосфорилированы, этот тип модификации является гибким механизмом для клеток для реагирования на внешние сигналы и условия окружающей среды. [ 25 ]
Киназы фосфорилируют белки и фосфатазы дефосфорилируют белки. Многие ферменты и рецепторы переключаются «на» или «выключение» путем фосфорилирования и дефосфорилирования. Обратимое фосфорилирование приводит к конформационному изменению структуры во многих ферментах и рецепторах , что приводит к тому, что они становятся активированными или деактивированными. Фосфорилирование обычно происходит на остатках серина , треонина , тирозина и гистидина в эукариотических белках. Гистидиновое фосфорилирование эукариотических белков, по -видимому, гораздо чаще, чем фосфорилирование тирозина. [ 26 ] В прокариотических белках фосфорилирование происходит на остатках серина, треонина, тирозина, гистидина, аргинина или лизина. [ 17 ] [ 18 ] [ 26 ] [ 27 ] Добавление фосфата (PO 4 3- ) Молекула в неполярную группу R аминокислотного остатка может превратить гидрофобную часть белка в полярную и чрезвычайно гидрофильную часть молекулы. Таким образом, динамика белка может вызвать конформационное изменение структуры белка посредством долгосрочной аллостерии с другими гидрофобными и гидрофильными остатками в белке.
Одним из таких примеров регуляторной роли, которую играет фосфорилирование, является белок супрессора опухоли p53 . Белок p53 сильно регулируется [ 28 ] и содержит более 18 различных сайтов фосфорилирования. Активация p53 может привести к остановке клеточного цикла, который может быть изменен при некоторых обстоятельствах или апоптотической гибели клеток. [ 29 ] Эта активность происходит только в ситуациях, когда клетка повреждена или физиология нарушена у нормальных здоровых людей.
После деактивирующего сигнала белок снова дефосфорилируется и перестает работать. [ 30 ] [ Цитация необходима ] Это механизм во многих формах передачи сигнала , например, способ обработки входящего света в светочувствительных клетках сетчатки .
Регуляторные роли фосфорилирования включают:
- Биологическая термодинамика реквизиции энергии
- Фосфорилирование Na + /K + -Аtpase во время транспортировки натрия (NA + ) и калий (k + ) ионы через клеточную мембрану в осморегуляции для поддержания гомеостаза содержания воды организма.
- Помновает ингибирование ферментов
- Фосфорилирование фермента GSK-3 AKT ( протеинкиназа B) как часть сигнального пути инсулина. [ 31 ]
- Фосфорилирование Src (произносится «SARC») тирозинкиназы C-концевой Src-киназы (CSK), вызывает конформационное изменение фермента, что приводит к сгибе в структуре, которое маскирует его киназный домен и, таким образом, закрыт «отключен». [ 32 ]
Мембранный транспорт
[ редактировать ]- Фосфорилирование Na + /K + -Аtpase во время транспортировки натрия (NA + ) и калий (k + ) ионы через клеточную мембрану в осморегуляции для поддержания гомеостаза содержания воды организма. [ 33 ]
- АТФ-связывающий кассетный транспортер
Деградация белка
[ редактировать ]- Фосфорилирование аргинина с помощью киназы MCSB отмечает белки для деградации с помощью протеазы CLP . Система фосфорилирования аргинина, которая широко распределена по грамположительным бактериям , по-видимому, функционально аналогична эукариотической убиквитин-протеасомной системе . [ 34 ]
Регуляция фермента (активация и ингибирование)
[ редактировать ]- Первым примером регуляции белка путем фосфорилирования, которая должна быть обнаружена, была гликогеновая фосфорилаза . Эдди Фишер и Эд Кребс описали, как фосфорилирование гликогеновой фосфорилазы B превратили ее в активную гликогенфосфорилазу A. Вскоре было обнаружено, что гликогенсинтаза, другой метаболический фермент, инактивируется фосфорилированием. [ 35 ]
- Фосфорилирование фермента GSK-3 AKT (протеинкиназа B) как часть сигнального пути инсулина. [ 31 ]
- Фосфорилирование тирозинкиназы Src с помощью С-концевой Src-киназы инактивирует SRC, индуцируя конформационные изменения, которые маскируют его киназный домен. [ 32 ]
- Фосфорилирование гистонов H2AX на серине 139, в пределах двух миллионов оснований (0,03% хроматина), окружающих двойной разрыв в ДНК, необходимо для восстановления двойного разрыва. [ 36 ] Фосфорилирование метилпуриновой ДНК -гликозилазы на серине 172 необходимо для восстановления иссечения основания алкилированного повреждения основания. [ 37 ]
Белковое взаимодействие
[ редактировать ]- Фосфорилирование цитозольных компонентов NADPH-оксидазы , большого мультибелкового фермента, присутствующего в фагоцитарных клетках , играет важную роль в регуляции взаимодействия белкового белка в ферменте. [ 38 ]
- Важно в деградации белка.
- В конце 1990-х годов было признано, что фосфорилирование некоторых белков приводит к деградации АТФ-зависимого пути убиквитина /протеасомы. Эти целевые белки становятся субстратами для определенных убиквитин -лигаз E3 только при фосфорилировании.
Сигнальные сети
[ редактировать ]Выяснение сложных событий фосфорилирования сложного сигнального пути может быть трудным. В клеточных сигнальных путях белок А фосфорилирует белок В, и В-фосфорилаты C. Однако в другом сигнальном пути белок D фосфорилирует a или фосфорилирует белок C. Глобальные подходы, такие как фосфопротеомика , изучение фосфорилированных белков, который является суб- Ветвь протеомики в сочетании с протеомикой на основе масс -спектрометрии была использована для идентификации и количественной оценки динамических изменений в фосфорилированных белках с течением времени. Эти методы становятся все более важными для систематического анализа сложных сетей фосфорилирования. [ 39 ] Они успешно использовались для выявления динамических изменений в статусе фосфорилирования более 6000 мест после стимуляции с помощью эпидермального фактора роста . [ 40 ] Другим подходом к пониманию сети фосфорилирования является измерение генетических взаимодействий между множественными фосфорилирующими белками и их мишенями. Это показывает интересные повторяющиеся модели взаимодействий - сетевые мотивы. [ 41 ] Вычислительные методы были разработаны для моделирования сетей фосфорилирования [ 42 ] [ 43 ] и предсказать их ответы при различных возмущениях. [ 44 ]
Фосфорилирование гистонов
[ редактировать ]Эукариотическая ДНК организована гистонными белками в специфических комплексах, называемых хроматином. Структура хроматина функционирует и облегчает упаковку, организацию и распределение эукариотической ДНК. Тем не менее, он оказывает негативное влияние на несколько фундаментальных биологических процессов, таких как транскрипция, репликация и репарация ДНК, путем ограничения доступности определенных ферментов и белков. Было показано, что посттрансляционная модификация гистонов, таких как фосфорилирование гистонов, модифицирует структуру хроматина путем изменения белка: ДНК или белок: белковые взаимодействия. [ 45 ] Гистоновые посттрансляционные модификации изменяют структуру хроматина. Наиболее часто связанное гистоновое фосфорилирование происходит во время клеточных реакций на повреждение ДНК, когда фосфорилированный гистон H2A разделяет большие домены хроматина вокруг сайта разрушения ДНК. [ 46 ] Исследователи исследовали, непосредственно ли влияют на модификации гистонов транскрипцию РНК -полимеразу II. Исследователи выбирают белки, которые, как известно, изменяют гистоны, чтобы проверить их влияние на транскрипцию, и обнаружили, что индуцированная стрессом киназа, MSK1, ингибирует синтез РНК. Ингибирование транскрипции с помощью MSK1 было наиболее чувствительным, когда шаблон находился в хроматине, поскольку матрицы ДНК не в хроматине были устойчивы к эффектам MSK1. Было показано, что фосфорилированный гистон H2A MSK1 на серине 1 и мутация серина 1 к аланину блокировала ингибирование транскрипции MSK1. Таким образом, результаты показали, что ацетилирование гистонов может стимулировать транскрипцию путем подавления ингибирующего фосфорилирования киназой в качестве MSK1. [ 47 ]
Киназы
[ редактировать ]Внутри белка фосфорилирование может происходить на нескольких аминокислотах . Считается, что фосфорилирование на серине является наиболее распространенным, за которым следует треонин. Фосфорилирование тирозина является относительно редким, но лежит на головке многих сигнальных путей фосфорилирования белка (например, в рецепторах, связанных с тирозинкиназой) у большинства эукариот. Фосфорилирование на аминокислотах, таких как серин, треонин и тирозин, приводит к образованию фосфопротеина, когда фосфатная группа фосфопротеина реагирует с -о -ООН, Ser, Thr или Tyr SideChain в реакции этерификации . [ 48 ] Однако, поскольку тирозин фосфорилированные белки относительно легко очищаются с использованием антител , сайты фосфорилирования тирозина относительно хорошо изучены. Фосфорилирование гистидина и аспартата происходит у прокариот в рамках двухкомпонентной передачи сигналов , а в некоторых случаях у эукариот в некоторых путях передачи сигнала. Анализ фосфорилированного гистидина с использованием стандартных биохимических и масс -спектрометрических подходов гораздо сложнее, чем у Ser, Thr или Tyr. [ 49 ] [ 7 ] [ 5 ] и [ 50 ] У прокариот, археи и некоторых нижних эукариот азот гистидина действует как нуклеофил и связывается с фосфатной группой. [ 51 ] Как только гистидин фосфорилируется регуляторным доменом регулятора ответа катализирует перенос фосфата в аспартат.
Рецепторные тирозинкиназы
[ редактировать ]
В то время как фосфорилирование тирозина обнаруживается в относительно низкой численности, оно хорошо изучается из -за простоты очистки фосфотирозина с использованием антител. Рецепторные тирозинкиназы являются важным семейством рецепторов клеточной поверхности, участвующих в трансдукции внеклеточных сигналов, таких как гормоны, факторы роста и цитокины. Связывание лиганда с мономерной рецепторной тирозинкиназой стабилизирует взаимодействия между двумя мономерами с образованием димера , после чего два связанных рецептора фосфорилируют остатки тирозина в транс . Фосфорилирование и активация рецептора активирует сигнальный путь через ферментативную активность и взаимодействия с белками адаптера. [ 52 ] Передача сигналов через рецептор эпидермального фактора роста (EGFR) , рецепторный тирозинкиназа, имеет решающее значение для развития нескольких систем органов, включая кожу, легкие, сердце и мозг. Чрезмерная передача сигналов через путь EGFR встречается при многих раке человека. [ 53 ]
Циклин-зависимые киназы
[ редактировать ]Циклин-зависимые киназы (CDK) представляют собой серин-триронинкиназы, которые регулируют прогрессирование через эукариотический клеточный цикл . CDK каталитически активны только тогда, когда связаны с регуляторным циклина . Клетки животных содержат по меньшей мере девять различных CDK, которые связываются с различными циклинами со значительной специфичностью. Ингибиторы CDK (CKIS) блокируют киназную активность в комплексе циклин-CDK, чтобы остановить клеточный цикл в G1 или в ответ на сигналы окружающей среды или повреждение ДНК. Активность различных CDK активирует сигнальные пути клеток и факторы транскрипции, которые регулируют ключевые события при митозе, таких как фазовый переход G1/S. Более ранние комплексы Cyclin-CDK обеспечивают сигнал для активации последующих комплексов Cyclin-CDK. [ 54 ]
Сайты
[ редактировать ]В данной клетке есть тысячи различных мест фосфорилирования с:
- В какой -либо конкретной клетке есть тысячи белков.
- Оценки от 1/10 до 1/2 белков фосфорилируются в некотором клеточном состоянии.
- 30–65% белков у людей и ~ 50% белков у дрожжей могут быть фосфорилированы. [ 15 ] [ 2 ]
- По оценкам, 230 000, 156 000 и 40 000 мест фосфорилирования существуют у человека, мыши и дрожжей соответственно. [ 2 ]
- Фосфорилирование часто встречается на нескольких отдельных сайтах на данном белке.
Поскольку фосфорилирование любого сайта на данном белке может изменить функцию или локализацию этого белка, понимание «состояния» клетки требует знания состояния фосфорилирования его белков. Например, как правило, если аминокислотный серин-473 в белке AKT фосфорилируется, AKT функционально активен в качестве киназы, и если он не фосфорилируется, AKT является неактивной киназой.
Сайты фосфорилирования имеют решающее значение для белков и их транспортировки и функций. Они представляют собой ковалентную модификацию белков посредством обратимого фосфорилирования. Это позволяет белкам оставаться входящими в клетке, поскольку отрицательный фосфорилированный сайт рассеивает свою проницаемость через клеточную мембрану. Дефосфорилирование белка позволяет клетке воспроизвести фосфаты посредством высвобождения пирофосфатов , что экономит использование АТФ в клетке. [ 55 ] Пример фосфорилирующего фермента обнаружен в бактериях E. coli . Он обладает щелочной фосфатазой в периплазматической области его мембраны. Самая внешняя мембрана проницаема для фосфорилированных молекул, однако внутренняя цитоплазматическая мембрана является непроницаемой из -за больших отрицательных зарядов. [ 56 ] Таким образом, бактерии E. coli хранят белки и пирофосфаты в ее периплазматической мембране, пока ни одна из них не будет необходимы в клетке.
Недавнее продвижение в фосфопротеомной идентификации привело к открытию бесчисленных сайтов фосфорилирования в белках. Это требовало интегративной среды для доступных данных, в которых организованы известные сайты фосфорилирования белков. Была создана кураторская база данных DBPAF, содержащая известные сайты фосфорилирования у H. sapiens , M. musculus , R. Norvegicus , D. Melanogaster , C. Elegans , S. Pombe и S. cerevisiae . В настоящее время база данных занимает 294 370 не избыточных мест фосфорилирования 40 432 белка. [ 57 ] Другие инструменты прогнозирования фосфорилирования в белках включают Netphos [ 58 ] Для эукариот, netphosbac [ 58 ] для бактерий и вируса [ 59 ] для вирусов.
Серин и треонин
[ редактировать ]Существует большое разнообразие сериновых остатков, и фосфорилирование каждого остатка может привести к различным метаболическим последствиям.
- Протеинкиназа N1 отвечает за фосфорилирование фактора, ассоциированного с рецептором TNF (TRAF1) на серине 139 в специфических условиях. Мышиный TRAF1 также фосфорилируется той же киназой, что приводит к молчанию активности IKK/NF-κB. Устранение фосфорилирования на серине 139 может быть достигнуто путем замены TRAF1 аланиновым остатком, что, следовательно, приводит к улучшению рекрутирования TBK1. [ 60 ]
- На остатках серина 789 FGFR1 фосфорилируется RSK2, когда киназа находится в его активной форме. Возможности сигнализации FGFR1 на сайте Serine 777 могут быть ослаблены путем фосфорилирования. Serine 1047 и Serine 1048 были связаны с уменьшенной аффинностью связывания убиквитин лигазы C-CBL с EFGR, когда они фосфорилируются. [ 61 ]
- Когда серин 349 фосфорилируется, аффинность связывания между белковым комплексом p62 и белком Keap1 усиливается, что связано с реакцией на стресс. [ 62 ]
- Когда серин 337 фосфорилируется протеинкиназой A in vitro, эффективность связывания ДНК значительно увеличивается. [ 63 ]
Известно, что фосфорилирование сериновых и треониновых остатков переправляется с модификацией O -glcnac остатков серина и треонина.
Тирозин
[ редактировать ]Фосфорилирование тирозина является быстрой, обратимой реакцией и одним из основных регуляторных механизмов в передаче сигнала . Рост клеток , дифференцировка , миграция и метаболический гомеостаз - это клеточные процессы, поддерживаемые фосфорилированием тирозина. Функция белкового тирозинкиназы и белко-тирозинфосфатазы уравновешивает уровень фосфотирозина на любом белке. Неисправность специфических цепей белковых тирозинкиназ и белковой тирозинфосфатазы было связано с множественными заболеваниями человека, такими как ожирение , резистентность к инсулину и сахарный диабет 2 типа . [ 64 ] Фосфорилирование на тирозине происходит у эукариот, избранных видов бактерий и присутствует среди прокариот. Фосфорилирование на тирозине поддерживает клеточную регуляцию у бактерий, сходных с его функцией у эукариот. [ 65 ]
Аргинин
[ редактировать ]Фосфорилирование аргинина во многих грамположительных бактериях отмечает белки для деградации с помощью протеазы CLP . [ 34 ]
Неканоническое фосфорилирование на его, ASP, Cys, Glu, Arg и Lys в клетках человека
[ редактировать ]Широко распространенное фосфорилирование белка человека происходит на множественных неканонических аминокислотах, включая мотивы, содержащие фосфорилированный гистидин (1 и 3 положения), аспартат, цистеин, глутамат, аргинин и лизин в экстрактах клеток HeLa. Из -за химической и термической лабилизации этих фосфорилированных остатков, для сохранения необходимы специальные процедуры и методы разделения наряду с тепловым фосфорилированием THR и TYR. [ 66 ]
Обнаружение и характеристика
[ редактировать ]Антитела могут использоваться в качестве мощного инструмента для выявления того, фосфорилируется ли белок на определенном месте. Антитела связываются и обнаруживают, вызванные фосфорилированием конформационные изменения в белке. Такие антитела называются фосфо-специфическими антителами; Сотни таких антител теперь доступны. Они становятся критическими реагентами как для базовых исследований, так и для клинической диагностики.
Изоформы посттрансляционной модификации (PTM) легко обнаруживаются на 2D-гелях . Действительно, фосфорилирование заменяет нейтральные гидроксильные группы на серинах, треонинах или тирозинах с отрицательно заряженными фосфатами с PK около 1,2 и 6,5. Таким образом, ниже рН 5,5 фосфаты добавляют один отрицательный заряд; Рядом с рН 6,5 они добавляют 1,5 отрицательных заряда; Выше рН 7,5, они добавляют 2 отрицательных заряда. Относительное количество каждой изоформы также может легко и быстро определяться по интенсивности окрашивания на 2D -гелях.
В некоторых очень специфических случаях обнаружение фосфорилирования в качестве сдвига в электрофоретической мобильности белка возможно на простом одномерном SDS-PAGE Gels, как описано, например, для транскрипционного коактиватора Kovacs et al. [ 67 ] Считается, что сильные конформационные изменения, связанные с фосфорилированием (которые сохраняются в решениях, содержащих моющие средства), лежат в основе этого явления. Большинство сайтов фосфорилирования, для которых был описан такой сдвиг подвижности, попадает в категорию сайтов SP и TP (то есть остаток пролина следует за фосфорилированным остатком серина или треонина).
Крупномасштабные анализы масс-спектрометрии использовались для определения сайтов фосфорилирования белка. Были опубликованы десятки исследований, каждый из которых выявил тысячи сайтов, многие из которых были ранее неписаны. [ 68 ] [ 69 ] Масс -спектрометрия идеально подходит для таких анализов с использованием фрагментации HCD или ETD , поскольку добавление фосфорилирования приводит к увеличению массы белка и фосфорилированного остатка. Для этих исследований необходимы расширенные, высоко точные масс-спектрометры, ограничивая технологию лабораториями высококачественными масс-спектрометрами. Однако анализ фосфорилированных пептидов с помощью масс -спектрометрии по -прежнему не так прост, как «обычные», немодифицированные пептиды. ETHCD был разработан, объединяющий электронную передачу и диссоциацию с более высокой энергией. По сравнению с обычными методами фрагментации, схема ETHCD обеспечивает более информативные спектры MS/MS для однозначной фосфозийной локализации. [ 70 ]
Подробная характеристика сайтов фосфорилирования очень сложна, и количественное определение фосфорилирования белка с помощью масс -спектрометрии требует изотопных подходов внутреннего стандарта. [ 71 ] Относительное количественное определение может быть получено с помощью различных технологий дифференциальной изотопной маркировки. [ 72 ] Существует также несколько методов количественного фосфорилирования белка, включая флуоресцентные иммуноанализа, микромасштабный термофорез , FRET , TRF, поляризация флуоресценции, флуоресцентное сдвиг, сдвиг подвижности, обнаружение на основе шариков и клетки. [ 73 ] [ 74 ]
Эволюция
[ редактировать ]Фосфорилирование белка распространено среди всех клад жизни, включая всех животных, растений, грибов, бактерий и археи. Происхождение механизмов фосфорилирования белка является наследственным и сильно расходилось между различными видами. У эукариот считается, что от 30 до 65% всех белков могут быть фосфорилированы, с десятками или даже сотнями тысяч различных сайтов фосфорилирования. [ 75 ] [ 2 ] Некоторые сайты фосфорилирования, по -видимому, эволюционировали в виде условных «выключателей», блокируя активный сайт фермента, например, в прокариотическом метаболическом ферменте изоцитратдегидрогеназу. Однако в случае белков, которые должны быть фосфорилированы, чтобы быть активными, менее ясно, как они могли возникнуть у нефосфорилированных предков. Было показано, что подмножество сериновых фосфоситов часто заменяется кислыми остатками, такими как аспартат и глутамат между различными видами. Эти анионные остатки могут взаимодействовать с катионными остатками, такими как лизин и аргинин, образуя солевые мосты , стабильные нековалентные взаимодействия, которые изменяют структуру белка. Эти фосфозиты часто участвуют в солевых мостах, что позволяет предположить, что некоторые сайты фосфорилирования развивались как условные »на« переключателях для солевых мостов, что позволяет этим белкам принимать активную конформацию только в ответ на конкретный сигнал. [ 76 ]
Существует около 600 известных эукариотических протеинкиназ, что делает их одним из крупнейших семейств эукариотических генов. Большая часть фосфорилирования осуществляется одним суперсемейством протеинкиназ, которые имеют консервативный домен киназы. Фосфорилирование белка высококонсервативное в пути, центральное для выживания клеток, таких как прогрессирование клеточного цикла, полагаясь на циклин-зависимые киназы (CDK), но отдельные сайты фосфорилирования часто являются гибкими. Цели фосфорилирования CDK часто имеют фосфоситы в беспорядочных сегментах , которые обнаружены в неидентичных местах даже у близких видов. И наоборот, цели фосфорилирования CDK в структурно определенных областях более высоко консервативны. В то время как активность CDK имеет решающее значение для роста и выживания клеток во всех эукариотах, лишь очень немногие фосфоситы демонстрируют сильное сохранение их точных положений. Позиционирование, вероятно, будет очень важным для фосфатов, которые аллостерически регулируют структуру белка, но гораздо более гибкий для фосфатов, которые взаимодействуют с фосфопептидсвязывающими доменами для рекрутирования регуляторных белков. [ 77 ]
Сравнения между эукариотами и прокариотами
[ редактировать ]Фосфорилирование белка является обратимой посттрансляционной модификацией белков. У эукариот функции фосфорилирования белка в клеточной передаче сигналов, экспрессии генов и дифференцировки. Он также участвует в репликации ДНК во время клеточного цикла и механизмов, которые справляются с индуцированными стрессом блоками репликации. По сравнению с эукариотами, прокариоты используют киназы типа Hanks и фосфатазы для передачи сигнала. Независимо от того, может ли фосфорилирование белков в бактериях также регулировать такие процессы, как репарация или репликация ДНК, по -прежнему остается неясным. [ 78 ]
По сравнению с фосфорилированием белка прокариот, исследования фосфорилирования белка у эукариот от дрожжей к клеткам человека были довольно обширными. Известно, что эукариоты полагаются на фосфорилирование гидроксильной группы на боковых цепях серина, треонина и тирозина для передачи сигналов клеток. Это основные регуляторные посттрансляционные модификации в эукариотических клетках, но фосфорилирование белка прокариот менее интенсивно изучено. В то время как серин, треонин и тирозин фосфорилируются у эукариот, гистидин и аспартат фосфорилируются в прокариотах и эукариотах. У бактерий фосфорилирование гистидина происходит в фосфоенолпируват-зависимых системах фосфотрансферазы (PTSS), которые участвуют в процессе интернализации, а также фосфорилировании сахаров. [ 79 ]
Фосфорилирование белка протеинкиназой было впервые показано в E. coli и сальмонелле тифимурий и с тех пор было продемонстрировано во многих других бактериальных клетках. [ 80 ] Было обнаружено, что бактерии используют гистидин и аспартатное фосфорилирование в качестве модели для передачи передачи сигналов бактерий. Фосфорилирование серина, треонина и тирозина также присутствует в бактериях. Бактерии несут киназы и фосфатазы, сходные с их эукариотическими эквивалентами, а также разработали уникальные киназы и фосфатазы, не обнаруженные у эукариот. [ 79 ]
Патология
[ редактировать ]Аномальное фосфорилирование белка участвовало в ряде заболеваний, включая рак , болезнь Альцгеймера , болезнь Паркинсона и другие дегенеративные расстройства .
Белок тау принадлежит группе белков, связанных с микротрубочками (карты), которые помогают стабилизировать микротрубочки в клетках, включая нейроны. [ 81 ] Ассоциация и стабилизирующая активность белка тау зависит от его фосфорилированного состояния. При болезни Альцгеймера из -за неправильных складок и аномальных конформационных изменений в структуре белка тау он делает неэффективным при связывании с микротрубочками и неспособна сохранить нейронную цитоскелетную структуру, организованной во время нервных процессов. Аномальный тау ингибирует и нарушает организацию микротрубочек и отключает нормальный тау от микротрубочек в цитозольную фазу. [ 82 ] Неправильные складывания приводят к ненормальной агрегации в фибриллярные клубки внутри нейронов. Белок тау должен быть фосфорилирован для функционирования, но гиперфосфорилирование белка тау является одним из основных влияний на его неспособность к ассоциации. [ 82 ] Фосфатазы PP1, PP2A, PP2B и PP2C дефосфорилируют белок тау in vitro , и их активность снижается в областях мозга у пациентов с болезнью Альцгеймера. [ 82 ] [ 83 ] Тау-фосфопротеин состоит из гиперфосфорилированного у пациента с болезнью Альцгеймера по сравнению со старым неафселированным человеком. Альцгеймерная болезнь тау, по -видимому, удаляет MAP1 и MAP2 (два других основных связанных белка) из микротрубочек, и этот вредный эффект изменяется, когда выполняется дефосфорилирование, подтверждая гиперфосфорилирование в качестве единственной причины активности каропки. [ 82 ]
Болезнь Паркинсона
[ редактировать ]α-синуклеин -это белок, который связан с болезнью Паркинсона. [ 84 ] У людей этот белок кодируется геном SNCA . [ 85 ] α-синуклеин участвует в рециркуляции синаптических везикул, которые несут нейротрансмиттеры и естественно встречаются в развернутой форме. Повышенные уровни α-синуклеина обнаруживаются у пациентов с болезнью Паркинсона. Существует корреляция между концентрацией нефосфорилированного α-синуклеина, присутствующего у пациента, и тяжестью болезни Паркинсона. [ 86 ] В частности, фосфорилирование Ser129 в α-синуклеине оказывает влияние на тяжесть. У здоровых пациентов более высокий уровень нефосфорилированного α-синуклеина, чем пациенты с болезнью Паркинсона. Измерение изменения отношения концентраций фосфорилированного α-синуклеина к нефосфорилированному α-синуклеину в пациенте может быть маркером прогрессирования заболевания. Антитела, которые нацелены на α-синуклеин в фосфорилированном Ser129, используются для изучения молекулярных аспектов синуклеинопатий. [ 87 ] [ 88 ]
Фосфорилирование Ser129 связано с агрегацией белка и дальнейшим повреждением нервной системы. Агрегация фосфорилированного α-синуклеина может быть усилена, если в недостаточных количествах присутствует пресинаптический белок каркаса, SEPT4. Прямое взаимодействие α-синуклеина с SEPT4 ингибирует фосфорилирование SER129. [ 89 ] [ 90 ] [ 91 ] Однако фосфорилирование Ser129 может наблюдаться без агрегации синуклеина в условиях сверхэкспрессии. [ 92 ]
Ссылки
[ редактировать ]- ^ Коэн, Филип (2002-05-01). «Происхождение фосфорилирования белка». Природная клеточная биология . 4 (5): E127–130. doi : 10.1038/ncb0502-e127 . ISSN 1465-7392 . PMID 11988757 . S2CID 29601670 .
- ^ Jump up to: а беременный в дюймовый и Vlastaridis, panayotis; Кириакиду, Пелагия; Чалиотис, Анаргирос; Ван де Пир, Ив; Оливер, Стивен Дж.; Amoutzias, Grigoris D. (2017-02-01). «Оценка общего количества фосфопротеинов и сайтов фосфорилирования у эукариотических протеомов» . Гигаскаука . 6 (2): 1–11. doi : 10.1093/gigascience/giw015 . PMC 5466708 . PMID 28327990 .
- ^ Илан Смоли, Нетта Шемеш, Михал Зив-Укельсон, Анат Бен-Зви, Эсти Йегер-Лотем (январь 2017 г.). «Асимметрически сбалансированная организация киназ по сравнению с фосфатазами в эукариотах определяет их четкие воздействия» . PLOS Computational Biology . 13 (1): E1005221. BIBCODE : 2017PLSCB..13E5221S . doi : 10.1371/journal.pcbi.1005221 . PMC 5279721 . PMID 28135269 .
{{cite journal}}: Cs1 maint: несколько имен: список авторов ( ссылка ) - ^ Potel, Clement M.; Лин, Миао-Хсиа; Черт, Альберт -младший; Lemeer, Simone (март 2018 г.). «Широко распространенное фосфорилирование бактериального белка, выявленное протеомикой на основе масс-спектрометрии» . Природные методы . 15 (3): 187–190. doi : 10.1038/nmeth.4580 . HDL : 1874/362159 . ISSN 1548-7105 . PMID 29377012 . S2CID 3367416 .
- ^ Jump up to: а беременный в Fuhs SR, Meisenhelder J, Aslanian A, Ma L, Zagorska A, Stankova M, Binnie A, Al-Obeidi F, Mauger J, Lemke G, Yates Jr 3rd, Hunter T (2015). «Моноклональные 1- и 3-фосфогистидиновые антитела: новые инструменты для изучения фосфорилирования гистидина» . Клетка . 162 (1): 198–210. doi : 10.1016/j.cell.2015.05.046 . PMC 4491144 . PMID 26140597 .
- ^ Хардман Г., Перкинс С., Браунридж П.Дж., Кларк С.Дж., Бирн Д.П., Кэмпбелл А.Е., Калиужни А., Майалл А., Эйерс П.А., Джонс А.Р., Эйерс С.Е. (2019). «Сильная анионная обменная фосфопротеомика выявляет обширное неканоническое фосфорилирование человека» . Embo j . 38 (21): E100847. doi : 10.15252/embj.2018100847 . PMC 6826212 . PMID 31433507 .
- ^ Jump up to: а беременный Fuhs Sr, Hunter T (2017). «Фисфорилирование: появление фосфорилирования гистидина в качестве обратимой регуляторной модификации» . Curr Mind Cell Biol . 45 : 8–16. doi : 10.1016/j.ceb.2016.12.010 . PMC 5482761 . PMID 28129587 .
- ^ Cieśla J; Frączyk t; Rode W (2011). «Фосфорилирование основных аминокислотных остатков в белках: важно, но легко пропустить» (PDF) . Acta Biochimica Polonica . 58 (2): 137–147. doi : 10.18388/abp.2011_2258 . PMID 21623415 .
- ^ Jump up to: а беременный Левен П.А.; Алсберг Кл (1906). «Продукты расщепления вителлина» . Дж. Биол. Химический 2 (1): 127–133. doi : 10.1016/s0021-9258 (17) 46054-6 .
- ^ Jump up to: а беременный Бернетт G; Кеннеди Эп (декабрь 1954 г.). «Ферментативное фосфорилирование белков» . Дж. Биол. Химический 211 (2): 969–80. doi : 10.1016/s0021-9258 (18) 71184-8 . PMID 13221602 .
- ^ Липманн Ф.А.; Левен П.А. (октябрь 1932). «Серинфосфорная кислота, полученная при гидролизе вителлиновой кислоты» . Дж. Биол. Химический 98 (1): 109–114. doi : 10.1016/s0021-9258 (18) 76142-5 .
- ^ Кресге, Николь; Симони, Роберт Д.; Хилл, Роберт Л. (2011-01-21). «Процесс обратимого фосфорилирования: работа Эдмонда Х. Фишера» . Журнал биологической химии . 286 (3): E1 - E2. doi : 10.1074/jbc.o110.000242 . ISSN 0021-9258 . PMC 3023531 . PMID 21294299 .
- ^ Хантер, Тони (2015-06-30). «Обнаружение первой тирозинкиназы» . Труды Национальной академии наук . 112 (26): 7877–7882. Bibcode : 2015pnas..112.7877h . doi : 10.1073/pnas.1508223112 . ISSN 0027-8424 . PMC 4491733 . PMID 26130799 .
- ^ Фишер, Эдмонд Х. (2010). «Фосфорилаза и происхождение обратимого фосфорилирования белка». Биологическая химия . 391 (2/3): 131–7. doi : 10.1515/bc.2010.011 . PMID 20030590 . S2CID 29724939 .
- ^ Jump up to: а беременный Коэн, Филип (2002-05-01). «Происхождение фосфорилирования белка». Природная клеточная биология . 4 (5): E127–130. doi : 10.1038/ncb0502-e127 . ISSN 1465-7392 . PMID 11988757 . S2CID 29601670 .
- ^ «Нобелевская премия по физиологии или медицине 1992» . www.nobelprize.org . Получено 2016-05-19 .
- ^ Jump up to: а беременный Коззоне AJ (1988). «Фосфорилирование белка у прокариот». Анну. Rev. Microbiol . 42 : 97–125. doi : 10.1146/annurev.mi.42.100188.000525 . PMID 2849375 .
- ^ Jump up to: а беременный Сток JB; Ninfa AJ; Сток AM (декабрь 1989 г.). «Фосфорилирование белка и регуляция адаптивных реакций у бактерий» . Микробиол. Преподобный 53 (4): 450–90. doi : 10.1128/mmbr.53.4.450-490.1989 . PMC 372749 . PMID 2556636 .
- ^ Чанг C; Стюарт RC (июль 1998 г.). «Двухкомпонентная система. Регуляция разнообразных сигнальных путей у прокариот и эукариот» . Plant Physiol . 117 (3): 723–31. doi : 10.1104/pp.117.3.723 . PMC 1539182 . PMID 9662515 .
- ^ Барфорд D; Дас Ак; Egloff MP (1998). «Структура и механизм белковых фосфатаз: понимание катализа и регуляции». Анну. Преподобный биофиз. Биомол. Структура 27 : 133–64. doi : 10.1146/annurev.biophys.27.1.133 . PMID 9646865 . S2CID 12138601 .
- ^ ПИТАК, Нико; Бехер, Дёрте; Шмидл, Себастьян Р.; Saier, Milton H.; Хеккер, Майкл; Commichau, Fabian M.; Stülke, Jörg (2010). «Фосфорилирование in vitro ключевых метаболических ферментов из Bacillus subtilis: PRKC фосфорилирует ферменты из разных ветвей основного метаболизма» . Журнал молекулярной микробиологии и биотехнологии . 18 (3): 129–140. doi : 10.1159/000308512 . ISSN 1660-2412 . PMID 20389117 . S2CID 19535600 .
- ^ Шмидл, Себастьян Р.; Гронау, Катрин; ПИТАК, Нико; Хеккер, Майкл; Бехер, Дёрте; Stülke, Jörg (июнь 2010 г.). «Фосфопротеом минимальной бактерий Mycoplasma pneumoniae: анализ полного известного кинома Ser/Thr предполагает существование новых киназ» . Молекулярная и клеточная протеомика . 9 (6): 1228–1242. doi : 10.1074/mcp.m900267-mcp200 . ISSN 1535-9484 . PMC 2877983 . PMID 20097688 .
- ^ Боденмиллер, Бернд; Ванка, Стефани; Крафт, Клодин; Urban, Jörg; Кэмпбелл, Дэвид; Педриоли, Патрик Г.; Герритс, Бертран; Пикотти, Паола ; Лам, Генри; Vitek, Olga ; Brusniak, Mi-Youn (2010-12-21). «Фосфопротеомный анализ выявляет взаимосвязанные реакции общеготь системы на возмущения киназ и фосфатаз в дрожжах» . Наука сигнализация . 3 (153): RS4. doi : 10.1126/Scisignal.2001182 . ISSN 1937-9145 . PMC 3072779 . PMID 21177495 .
- ^ Яхи, Нозому; Сайто, Ринтаро; Сугияма, Наюки; Томита, Масару; Ишихама, Ясуши (2011-01-27). «Интегративные особенности карты взаимодействия дрожжевого фосфопротеома и белкового белка» . PLOS Computational Biology . 7 (1): E1001064. BIBCODE : 2011PLSCB ... 7E1064Y . doi : 10.1371/journal.pcbi.1001064 . ISSN 1553-7358 . PMC 3029238 . PMID 21298081 .
- ^ Джонсон Л.Н., Барфорд Д. (1993). «Влияние фосфорилирования на структуру и функцию белков [J]». Ежегодный обзор биофизики и биомолекулярной структуры . 22 (1): 199–232. doi : 10.1146/annurev.bb.22.060193.001215 . PMID 8347989 .
- ^ Jump up to: а беременный Ciesla J; Fraczyk T; Rode W (2011). «Фосфорилирование основных аминокислотных остатков в белках: важно, но легко пропустить» . Acta Biochim. Поли 58 (2): 137–47. doi : 10.18388/abp.2011_2258 . PMID 21623415 .
- ^ Deutscher, J.; Saier, J. (2005). «Фосфорилирование белка Ser/Thr/Tyr в бактериях - в течение долгого времени пренебрегает, теперь хорошо установлена». Журнал молекулярной микробиологии и биотехнологии . 9 (3–4): 125–131. doi : 10.1159/000089641 . PMID 16415586 . S2CID 13093867 .
- ^ Эшкрофт М; Kubbutat MH; Vousden KH (март 1999 г.). «Регуляция функции p53 и стабильности путем фосфорилирования» . Мол Клетка. Биол . 19 (3): 1751–8. doi : 10.1128/mcb.19.3.1751 . PMC 83968 . PMID 10022862 .
- ^ Бейтс с; Vousden KH (февраль 1996 г.). «P53 в сигнальной контрольной точке остановки или апоптоза» . Карт Мнение Генет. Девчонка 6 (1): 12–8. doi : 10.1016/s0959-437x (96) 90004-0 . PMID 8791489 .
- ^ Learnwithalbert (2016-09-16). «В чем разница между фосфорилированием и дефосфорилированием?» Полем Блог Альберта . Получено 2019-02-01 .
- ^ Jump up to: а беременный Van Weeren PC; де Брюн К.М.; De Vries-Smits Am; Ван Линт J; Бургеровка BM (май 1998). «Основная роль протеинкиназы B (PKB) в инсулино-индуцированной гликогенсинтазы киназе 3 инактивация. Характеристика доминантно-негативного мутанта PKB» . Дж. Биол. Химический 273 (21): 13150–6. doi : 10.1074/jbc.273.21.13150 . PMID 9582355 .
- ^ Jump up to: а беременный Коул Па; Шен К; Qiao y; Ван Д (октябрь 2003 г.). «Протеиновые тирозинкиназы SRC и CSK: сказка хвоста». Curr Smink Chem Biol . 7 (5): 580–5. doi : 10.1016/j.cbpa.2003.08.009 . PMID 14580561 .
- ^ Zubareva, VM; Лапашина, как; Shugaeva, Te; Литвин, Ав; Feniouk, Ba (декабрь 2020 г.). «Ротари-ионно-транслокационные АТФазы/АТФ-синтазы: разнообразие, сходства и различия» . Биохимия. Biohhimiia . 85 (12): 1613–1630. doi : 10.1134/s0006297920120135 . ISSN 1608-3040 . PMID 33705299 . S2CID 229701146 .
- ^ Jump up to: а беременный Broch Trentini, Débora (2016). «Фосфорилирование аргинина отмечает белки для деградации протеазой CLP» . Природа . 539 (7627): 48–53. Bibcode : 2016natur.539 ... 48t . doi : 10.1038/nature20122 . PMC 6640040 . PMID 27749819 .
- ^ Джонсон, Луиза Н. (2009-08-01). «Регуляция фосфорилирования белка». Биохимическое общество транзакций . 37 (Pt 4): 627–641. doi : 10.1042/bst0370627 . ISSN 1470-8752 . PMID 19614568 .
- ^ Rogakou EP, Pilch Dr, Orr AH, Ivanova VS, Bonner Wm (март 1998 г.). «Двойные разрывы ДНК индуцируют фосфорилирование гистона H2AX на серине 139» . Дж. Биол. Химический 273 (10): 5858–68. doi : 10.1074/jbc.273.10.5858 . PMID 9488723 .
- ^ Картер Р.Дж., Парсонс Дж.Л. (май 2016 г.). «Ремонт базового удаления, путь, регулируемый посттрансляционными модификациями» . Мол Клетка. Биол . 36 (10): 1426–37. doi : 10.1128/mcb.00030-16 . PMC 4859697 . PMID 26976642 .
- ^ Babior BM (март 1999 г.). «NADPH Oxidase: обновление». Кровь . 93 (5): 1464–76. doi : 10.1182/blood.v93.5.1464 . PMID 10029572 .
- ^ Олсен СП; Благейв Б; Гнад Ф; Macek B; Кумар С; Мортенсен P; Манн М (ноябрь 2006 г.). «Глобальная динамика фосфорилирования в Global, in vivo и сайт в сигнальных сетях» . Клетка . 127 (3): 635–48. doi : 10.1016/j.cell.2006.09.026 . PMID 17081983 . S2CID 7827573 .
- ^ Li-rong y; Issaq HJ; Veenstra TD (2007). «Фосфопротеомика для открытия киназ в качестве биомаркеров рака и лекарственных целей». Протеомика: клинические применения . 1 (9): 1042–1057. doi : 10.1002/prca.200700102 . PMID 21136756 . S2CID 33999702 .
- ^ Fiedler D, Braberg H, Mehta M, Chechik G, Cagney G, Mukherjee P, Silva AC, Shales M, et al. (2009). «Функциональная организация сети фосфорилирования S. cerevisiae» . Клетка . 136 (5): 952–963. doi : 10.1016/j.cell.2008.12.039 . PMC 2856666 . PMID 19269370 .
- ^ Schoeberl, B; Эйхлер-Джонссон, C; Жиль, Эд; Мюллер, G (апрель 2002 г.). «Вычислительное моделирование динамики каскада киназы карты, активируемой поверхностными и интернализованными рецепторами EGF». Nature Biotechnology . 20 (4): 370–5. doi : 10.1038/nbt0402-370 . PMID 11923843 . S2CID 9851026 .
- ^ Олдридж, BB; Берк, JM; Lauffenburger, DA; Sorger, PK (ноябрь 2006 г.). «Физико -химическое моделирование путей сигнализации клеток». Природная клеточная биология . 8 (11): 1195–203. doi : 10.1038/ncb1497 . PMID 17060902 . S2CID 14586526 .
- ^ Чжу, F; Guan, Y (11 июня 2014 г.). «Прогнозирование динамического отклика передачи сигналов при невидимых возмущениях» . Биоинформатика . 30 (19): 2772–8. doi : 10.1093/bioinformatics/btu382 . PMC 4173019 . PMID 24919880 .
- ^ Савика, Анна; Seiser, Christian (2014-08-01). «Ощущение ядра гистона фосфорилирование - вопрос идеального времени» . Biochimica et Biophysica Acta (BBA) - механизмы регуляции генов . 1839 (8): 711–718. doi : 10.1016/j.bbagrm.2014.04.013 . PMC 4103482 . PMID 24747175 .
- ^ Россетто, Дорин; Аввакумов, Никита; Кот, Жак (2012-10-01). «Гистоновое фосфорилирование» . Эпигенетика . 7 (10): 1098–1108. doi : 10.4161/epi.21975 . ISSN 1559-2294 . PMC 3469451 . PMID 22948226 .
- ^ Чжан, ты; Гриффин, Карен; Мондал, Нилима; Парвин, Джеффри Д. (2004-05-21). «Фосфорилирование гистона H2A ингибирует транскрипцию на шаблонах хроматина» . Журнал биологической химии . 279 (21): 21866–21872. doi : 10.1074/jbc.m400099200 . ISSN 0021-9258 . PMID 15010469 .
- ^ Гришам, Реджинальд Х. Гаррет, Чарльз М. (2013). Биохимия (5 -е изд.). Белмонт, Калифорния: Брукс/Коул, Cengage Learning. ISBN 978-1133106296 .
{{cite book}}: Cs1 maint: несколько имен: список авторов ( ссылка ) - ^ Гонсалес-Санчес М.Б., Ланукара Ф., Хелм М., Эйерс К.Е. (2013). «Попытка переписать историю: проблемы с анализом гистидина-фосфорилированных пептидов». Biochem Soc Trans . 41 (4): 1089–1095. doi : 10.1042/bst20130072 . PMID 23863184 .
- ^ Томасон П; Кей Р (сентябрь 2000 г.). «Эукариотическая передача сигнала через гистидин-аспартатную фосфорела» (PDF) . J. Cell Sci . 113 (18): 3141–50. doi : 10.1242/jcs.113.18.3141 . PMID 10954413 .
- ^ Путтик, Дженнифер; Бейкер, Эдвард Н.; Delbaere, Louis TJ (2008). «Гистидиновое фосфорилирование в биологических системах». Biochimica et Biophysica Acta (BBA) - белки и протеомика . 1784 (1): 100–105. doi : 10.1016/j.bbapap.2007.07.008 . ISSN 1570-9639 . PMID 17728195 .
- ^ Леммон, Марк А .; Schlessinger, Joseph (июнь 2010 г.). «Сигнализация клеток рецепторными тирозинкиназами» . Клетка . 141 (7): 1117–34. doi : 10.1016/j.cell.2010.06.011 . PMC 2914105 . PMID 20602996 .
- ^ CHO HS, Leahy DJ; Лихи (2002). «Структура внеклеточной области HER3 выявляет междоменную привязку» . Наука . 297 (5585): 1330–1333. Bibcode : 2002sci ... 297.1330c . doi : 10.1126/science.1074611 . PMID 12154198 . S2CID 23069349 .
- ^ Морган, Дэвид О. (2007). Клеточный цикл: принципы контроля. Лондон: New Science Press, 1 -е изд.
- ^ Гаррет, Реджинальд Х.; Гришам, Чарльз М. (2013). Биохимия . Мэри Финч, Cengage Learning. С. 489–491.
- ^ Нинфа, Александр; Дэвид П. Баллу, Дэвид (1998). Фундаментальные лабораторные подходы к биохимии и биотехнологии (2 -е изд.). Fitzgerald Science Press. С. 230–231.
- ^ Улла, Шахид; Лин, Шаофенг (2016). «DBPAF: интегративная база данных фосфорилирования белка у животных и грибов» . Научные отчеты . 6 : 23534. BIBCODE : 2016NATSR ... 623534U . doi : 10.1038/srep23534 . PMC 4806352 . PMID 27010073 . Получено 17 мая 2016 года .
- ^ Jump up to: а беременный Блом, Николай; Gammeloft, Steen; Брунак, Сёрен (1999-12-17). «Последовательность и структурно-основанные прогноз эукариотических мест фосфорилирования белка1». Журнал молекулярной биологии . 294 (5): 1351–1362. doi : 10.1006/jmbi.1999.3310 . PMID 10600390 .
- ^ Хуан, Кай-яо; Лу, Ченг-Зунг; Бретана, Нил Арвин; Ли, Цонг-Йи; Chang, Tzu-Hao (2013-01-01). «Viralphos: включение рекурсивно статистического метода для прогнозирования сайтов фосфорилирования на вирусных белках» . BMC Bioinformatics . 14 (16): S10. doi : 10.1186/1471-2105-14-S16-S10 . ISSN 1471-2105 . PMC 3853219 . PMID 24564381 .
- ^ Усса, NA (1 марта 2013 г.). «Фосфорилирование TRAF1 на серине 139 модулирует активность NF-κB ниже 4-1BB в Т-клетках». Биохимическая и биофизическая исследовательская коммуникация . 432 (1): 129–134. doi : 10.1016/j.bbrc.2013.01.073 . PMID 23376065 .
- ^ Надратовска-Весоловская, б (21 октября 2016 г.). «RSK2 регулирует эндоцитоз рецептора FGF 1 путем фосфорилирования на серине 789» . Онкоген . 33 (40): 4823–4836. doi : 10.1038/onc.2013.425 . PMID 24141780 .
- ^ Танджи, Куниказу (3 мая 2014 г.). «Фосфорилирование серина 349 P62 в мозге болезни Альцгеймера» . Acta Neuropathologica Communications . 2 (50): 50. doi : 10.1186/2051-5960-2-50 . PMC 4035093 . PMID 24886973 .
- ^ Хоу, Ших (14 ноября 2003 г.). «Фосфорилирование серина 337 NF-Kappab P50 является критическим для связывания ДНК» . Журнал биологической химии . 278 (46): 45994–8. doi : 10.1074/jbc.m307971200 . PMID 12947093 .
- ^ Редактор, Кендра К. Бенс (2013). Белковая тирозинфосфатаза контроль метаболизма . Нью -Йорк, Нью -Йорк: Springer New York. ISBN 978-1-4614-7855-3 .
{{cite book}}:|last1=имеет общее имя ( справка ) - ^ Коззоне, Ален Дж.; Грангеш, Кристоф; Дублет, Патриция; Duclos, Бертран (1 марта 2004 г.). «Фосфорилирование белка на тирозине у бактерий». Архив микробиологии . 181 (3): 171–181. doi : 10.1007/s00203-003-0640-6 . PMID 14745484 . S2CID 37161183 .
- ^ Eyers CE, Hardman G (21 августа 2019 г.). «Сильная анионная обменная фосфопротеомика выявляет обширное неканоническое фосфорилирование человека» . Embo Journal . 38 (21): E100847. doi : 10.15252/embj.2018100847 . PMC 6826212 . PMID 31433507 .
- ^ Ковач К.А., Стейнманн М; MAGISTRETTI PJ; Полуполон О; Cardinaux Jr (сентябрь 2003 г.). «CCAAT/Энхансер-связывающие члены семейства белков рекрутируют коактиватор CREB-связывающий белок и запускают его фосфорилирование» . Дж. Биол. Химический 278 (38): 36959–65. doi : 10.1074/jbc.m303147200 . ISSN 0021-9258 . PMID 12857754 .
- ^ Munton RP, Tweedie-Cullen R, Livingstone-Zatchej M, Weinandy F, Waidelich M, Longo D, Gehrig P, Potthast F, et al. (Февраль 2007 г.). «Качественный и количественный анализ фосфорилирования белка в наивных и стимулированных синаптосомных препаратах мыши» (PDF) . Мол Клетка. Протеомика . 6 (2): 283–93. doi : 10.1074/mcp.m600046-mcp200 . PMID 17114649 . S2CID 18221665 .
- ^ Тринидад JC; Thalhammer A; Specht CG; Линн А.Дж; Бейкер PR; Schoepfer R; Burlingame AL (апрель 2008 г.). «Количественный анализ синаптического фосфорилирования и экспрессии белка» . Мол Клетка. Протеомика . 7 (4): 684–96. doi : 10.1074/mcp.m700170-mcp200 . PMID 18056256 .
- ^ Фрезе, Кристиан; Хуцзян Чжоу; Томас Таус; AF MAARTEN ALTELAAR; Карл Мехтлер; Альберт младший Хек; Шабаз Мохаммед (1 марта 2013 г.). «Единственная локализация фосфозирования с использованием электронного переноса/диссоциации столкновений с более высокой энергией (ETHCD)» . J Proteome Res . 12 (3): 1520–1525. doi : 10.1021/pr301130K . PMC 3588588 . PMID 23347405 .
- ^ Гербер С.А.; Rush J; Stemman O; Kirschner MW; Gygi SP (июнь 2003 г.). «Абсолютная количественная оценка белков и фосфопротеинов из клеточных лизатов тандемным MS» . Прокурор Нат. Академический Наука США . 100 (12): 6940–5. Bibcode : 2003pnas..100.6940g . doi : 10.1073/pnas.0832254100 . PMC 165809 . PMID 12771378 .
- ^ Gygi sp; Rist B; Гриффин TJ; Eng J; Aebersold R (2002). «Протеомный анализ белков с низким содержанием средств с использованием многомерной хроматографии и изотопных аффинных меток». J. Proteome Res . 1 (1): 47–54. doi : 10.1021/pr015509n . PMID 12643526 .
- ^ Olive DM (октябрь 2004 г.). «Количественные методы анализа фосфорилирования белка при развитии лекарственного средства». Эксперт Rev Proteomics . 1 (3): 327–41. doi : 10.1586/14789450.1.3.327 . PMID 15966829 . S2CID 30003827 .
- ^ Chen H, Kovar J, Sissons S, Cox K, Matter W, Chadwell F, Luan P, Vlahos CJ, et al. (Март 2005 г.). «Иммуноцитокмический анализ на основе клеток для мониторинга сигнальных путей киназы и эффективности лекарственного средства» (PDF) . Анальный. Биохимия . 338 (1): 136–42. Citeseerx 10.1.1.335.3523 . doi : 10.1016/j.ab.2004.11.015 . PMID 15707944 . Архивировано из оригинала (PDF) 2012-02-22 . Получено 2019-05-26 .
- ^ Коэн П. (2000). «Регуляция функции белка с помощью многосайтового фосфорилирования - 25 -летнего обновления». Тенденции Biochem. Наука 25 (12): 596–601. doi : 10.1016/s0968-0004 (00) 01712-6 . PMID 11116185 .
- ^ Перлман С.М., Сербер З., Феррелл Дж. (2011). «Механизм эволюции сайтов фосфорилирования» . Клетка . 147 (4): 934–946. doi : 10.1016/j.cell.2011.08.052 . PMC 3220604 . PMID 22078888 .
- ^ Холт Л.Дж., Туч Б.Б., Виллен Дж., Джонсон А.Д., Гиги С.П., Морган До (2009). «Глобальный анализ сайтов фосфорилирования субстрата CDK1 дает представление о эволюции» . Наука . 325 (5948): 1682–1686. Bibcode : 2009Sci ... 325.1682H . doi : 10.1126/science.1172867 . PMC 2813701 . PMID 19779198 .
- ^ Garcia-Garcia, Transito (2016). «Роль фосфорилирования белка в регуляции клеточного цикла и связанных с ДНК процессов в бактериях» . Границы в микробиологии . 7 : 184. doi : 10.3389/fmicb.2016.00184 . PMC 4754617 . PMID 26909079 .
- ^ Jump up to: а беременный Macek, B.; Миджакович, я.; Olsen, J.; Гнад, F; Кумар, C.; Дженсен, П. (2007). «Серин/треонин/тирозин фосфопротеом модельной бактерии Bacillus subtilis» . Мол Клетка. Протеомика . 6 (4): 697–707. doi : 10.1074/mcp.m600464-mcp200 . PMID 17218307 .
- ^ Коззоне, AJ (1988). «Фосфорилирование белка у прокариот». Annu Rev Microbiol . 42 : 97–125. doi : 10.1146/annurev.mi.42.100188.000525 . PMID 2849375 .
- ^ Вулф, Майкл С. (2012-12-19). «Роль тау в нейродегенеративных заболеваниях и его потенциал в качестве терапевтической цели» . Scientifica . 2012 : 796024. DOI : 10.6064/2012/796024 . PMC 3820460 . PMID 24278740 .
- ^ Jump up to: а беременный в дюймовый Коларава, Михала; Гарсия-Сьерра, Франциско; Бартос, Эльс; Рикни, Ян; Рипова, Даниэла (2012-05-29). «Структура и патология белка тау при болезни Альцгеймера» . Международный журнал болезни Альцгеймера . 2012 : 731526. DOI : 10.1155/2012/731526 . ISSN 2090-8024 . PMC 3368361 . PMID 22690349 .
- ^ Креспо-Биль, Наталья; Theunis, Клара; Leuven, Fred Van (2012-06-08). «Белок тау: основная причина синаптической и нейрональной дегенерации при болезни Альцгеймера» . Международный журнал болезни Альцгеймера . 2012 : 251426. DOI : 10.1155/2012/251426 . ISSN 2090-8024 . PMC 3376502 . PMID 22720188 .
- ^ Spillantini, Mg; Кроутер, Ра; Джейкс, R; Хасегава, м; Goedert, M (26 мая 1998 г.). «Альфа-синуклеин в нити, включении тел Леви от болезни Паркинсона и деменции с леви телами» . Труды Национальной академии наук Соединенных Штатов Америки . 95 (11): 6469–73. Bibcode : 1998pnas ... 95.6469S . doi : 10.1073/pnas.95.11.6469 . PMC 27806 . PMID 9600990 .
- ^ «Генетика дома ссылка: SNCA» . Национальная библиотека медицины США. 12 ноября 2013 года . Получено 14 ноября 2013 года .
- ^ Burré, J; Шарма, м; Südhof, TC (1 марта 2018 г.). «Клеточная биология и патофизиология α-синуклеина» . Перспективы Cold Spring Harbor в медицине . 8 (3): A024091. doi : 10.1101/cshperspect.a024091 . PMC 5519445 . PMID 28108534 .
- ^ Резерфорд, Нью -Джерси; Брукс, м; Гиассон, Би (8 августа 2016 г.). «Новые антитела к фосфорилированному α-синуклеиново-серину 129 и серину NFL 473 демонстрируют тесную молекулярную гомологию этих эпитопов» . Acta Neuropathologica Communications . 4 (1): 80. DOI : 10.1186/S40478-016-0357-9 . PMC 4977832 . PMID 27503460 .
- ^ Paleologou, Ke; Шмид, AW; Rospigliosi, CC; Ким, Хи; Ламберто, Гр; Фреденбург, Ра; Lansbury Pt, Jr; Фернандес, CO; Eliezer, D; Zweckstetter, m; Лашуэль, Ха (13 июня 2008 г.). «Фосфорилирование в Ser-129, но не фосфомимика S129E/D ингибирует фибрилляцию альфа-синуклеина» . Журнал биологической химии . 283 (24): 16895–905. doi : 10.1074/jbc.m800747200 . PMC 2423264 . PMID 18343814 .
- ^ «Болезнь Паркинсона | Выяснение роли фосфорилирования в модулировании альфа-синуклеинового агрегации и токсичности при болезни Паркинсона и связанных с ними расстройствах» . Болезнь Паркинсона | Фонд Майкла Дж. Фокса для исследований Паркинсона . Получено 2016-05-14 .
- ^ Ван, Ю; Ши, мин; Чунг, Кэтрин А.; Забетик, Сайрус П.; Леверенц, Джеймс Б.; Берг, Даниэла; Srulijes, Карин; Трояновский, Джон Q.; Ли, Вирджиния М.-Ю. (2012-02-15). «Фосфорилированный α-синуклеин при болезни Паркинсона» . Научная трансляционная медицина . 4 (121): 121RA20. doi : 10.1126/scitranslmed.3002566 . ISSN 1946-6234 . PMC 3302662 . PMID 22344688 .
- ^ Стюарт, Тессандра; Соси, Весна ; Айси, Ян О; Wszolek, Zbignieew k; Утти, Райан Дж; Хасегава, Казуко; Йокояма, Теруо; Забетик, Сайрус П; Леверенц, Джеймс Б. (2015-01-31). «Фосфорилированный α-синуклеин при болезни Паркинсона: корреляция зависит от заболевания нескольких ». Acta Невропатологическая связь 3 (1): 7. doi : 10.1186/ s40478-015-0185-3 ISSN 2051-5 PMC 4362824 25637461PMID
- ^ Laferrière, Флорент; Он, Синь; Цингхирино, Федерика; Дудникофф, Эвелин; Фаггиани, Эмили; Мейснер, Вассилиос Г.; Безард, Эрван; Де Гиорги, Франческа; Ичас, Франсуа (2020-10-29). «Сверхэкспрессия α-синуклеина олигодендроцитами у трансгенных мышей не повторяет фибриллярную агрегацию, наблюдаемую при многочисленной системной атрофии» . Ячейки 9 (11): 2371. doi : 10.3390/cells9112371 . ISSN 2073-4409 . PMC 7693764 . PMID 33138150 .