Спора


В биологии спора представляет собой единицу сексуального (в грибах) или бесполого репродукции , которое может быть адаптировано для рассеивания и для выживания, часто в течение длительных периодов времени, в неблагоприятных условиях. [ 1 ] Споры составляют часть жизненных циклов многих растений , водорослей , грибов и простейших . [ 2 ] Считалось, что они появились еще в среднем периоде ордовика в середине-заключении в качестве адаптации ранних наземных растений. [ 3 ]
Бактериальные споры не являются частью сексуального цикла, но являются устойчивыми структурами, используемыми для выживания в неблагоприятных условиях. [ 4 ] Мексозоианские споры выпускают амебоидные инфекционные микробы («амебулы») в своих хозяев для паразитарной инфекции, но также воспроизводят в хозяевах путем сочетания двух ядер в плазмодие, которое развивается из амебулы. [ 5 ]
У растений споры обычно являются гаплоидными и одноклеточными вырабатываются мейозом в спорангии диплоидного и спорофита . В некоторых редких случаях диплоидная спора также производится в некоторых водорослях или грибах. [ 6 ] В благоприятных условиях спора может превратиться в новый организм с использованием митотического разделения, производя многоклеточный гаметофит , который в конечном итоге продолжает производить гаметы. Два гамета сливаются, чтобы сформировать зиготу , которая превращается в новый спорофит. Этот цикл известен как чередование поколений .
Споры семянных растений производятся внутри, а мегапоры (образуются в яйцеклетке), а микроспоры участвуют в формировании более сложных структур, которые образуют рассеянные единицы, семена и зерна пыльцы .
Определение
[ редактировать ]Термин спора происходит от древнегреческого слова σπορά spora , означающего « семя , посев», связанное с σπόρος споро , «посев» и σπείρειν Спейреин , «Сеять».
На общем языке разница между «спорой» и « гаметой » заключается в том, что спор прорастает и превратится в спорезацию , в то время как гамете необходимо объединиться с другой гаметой, чтобы сформировать зиготу, прежде чем развиваться.
Основное различие между спорами и семенами в качестве единиц диспергирования состоит в том, что споры являются одноклеточными, первая ячейка гаметофита, в то время как семена содержат внутри них развивающее эмбрион (многоклеточный спорофит следующего поколения), создаваемого слиянием мужского гамита Пыльцевая трубка с женской гаметой, образованной мегагаметофитом в яйце. Споры прорастают, чтобы привести к гаплоидным гаметофитам, в то время как семена прорастают, чтобы привести к диплоидным спорофитам.
Классификация организмов, производящих спор
[ редактировать ]
Растения
[ редактировать ]Споры сосудистых растений всегда гаплоидны . Сосудистые растения являются либо гомоспористыми (или изоспорированными) , либо гетеропринятыми . Растения, которые являются гомоспортными, производят споры того же размера и типа.
Гетероспористые растения, такие как растения для семян , шипики , киллеры и папоротники порядка Salviniales, производят споры двух разных размеров: более крупная спор (мегаспор), функционирующая как «женская» спор, а меньшая (микроспора) функционирует как »как« мужской". Такие растения, как правило, вызывают два вида споров из отдельных спорангии, либо мегапорангий , который производит мегапоры или микроспорангий , который производит микроспоры. В цветущих растениях эти спорангии встречаются в коце и пыльниках соответственно.
Грибы
[ редактировать ]Грибы обычно производят споры во время сексуального и бесполого воспроизведения. Споры обычно являются гаплоидными и превращаются в зрелых гаплоидных индивидуумов через митотическое разделение клеток ( Urediniospores и телиоспоры среди ржавчиков являются дикариотическими). Дикариотические клетки являются результатом слияния двух гаплоидных гаметовых клеток. Среди спорогенных дикариотических клеток встречается кариогамия (слияние двух гаплоидных ядер) для получения диплоидной клетки. Диплоидные клетки подвергаются мейозу для получения гаплоидных споров. [ Цитация необходима ]
Классификация споров
[ редактировать ]Споры могут быть классифицированы несколькими способами, такими как их структура, производящая спор, функции, происхождение в течение жизненного цикла и мобильность.
Ниже приведена таблица, в которой перечислен режим классификации, названия, идентификации характеристики, примеров и изображений различных видов спор.
| Режим классификации | Имя | Идентификация характеристики | Пример споры, содержащий организм | Изображение | |
|---|---|---|---|---|---|

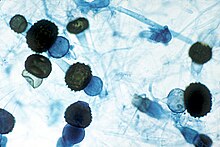
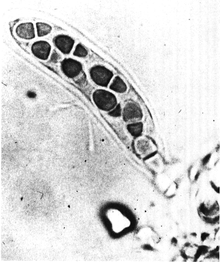


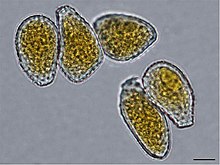
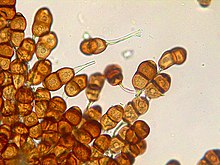












Внешняя анатомия
[ редактировать ]


При высоком увеличении споры часто имеют сложные узоры или украшения на их внешних поверхностях. Специализированная терминология была разработана для описания особенностей таких моделей. Некоторые маркировки представляют собой отверстия, где можно проникнуть в трудную внешнюю слой споры, когда происходит прорастание. Споры могут быть классифицированы на основе позиции и количества этих маркировков и отверстий. Споры Alete не показывают линий. В монолетных спорах на спор есть одна узкая линия (Laesura). [ 8 ] Указывая на предыдущий контакт двух споров, которые в конечном итоге разделялись. [ 3 ] В Trilete Spores на каждой спор показывается три узкие линии, излучающиеся от центрального полюса. [ 8 ] Это показывает, что четыре споры разделили общее происхождение и первоначально были в контакте друг с другом, образуя тетраэдр. [ 3 ] Более широкая апертура в форме канавки может быть названа Кольпусом . [ 8 ] Количество Colpi различает основные группы растений. У Eudicots есть триколпатные споры (т.е. споры с тремя Colpi). [ 9 ]
Spore tetrads and trilete spores
[ редактировать ]Тетрады споры с оболочкой принимаются в качестве самых ранних доказательств жизни растений на суше, [ 10 ] Компания от среднего ордовика (ранний Llanvirn, ~ 470 миллионов лет назад ), период, из которого еще не было восстановлено макрофоссили. [ 11 ] Индивидуальные трилетные споры, напоминающие споры современных криптогамических растений, впервые появились в отчете окаменелости в конце периода ордовика. [ 12 ]
Рассеяние
[ редактировать ]В грибах как бесполые, так и сексуальные споры или спорангиоспоры многих грибковых видов активно рассеяются путем насильственного выброса из их репродуктивных структур. Это выброс обеспечивает выход споров из репродуктивных структур, а также перемещаться по воздуху на большие расстояния. Таким образом, многие грибы обладают специализированными механическими и физиологическими механизмами, а также структурами споры, такие как гидрофобины , для выброса спор. Эти механизмы включают, например, насильственный разряд аскоспоров, обеспечиваемых структурой ASCUS и накоплением осмолитов в жидкостях ASCUS, которые приводят к взрывоопасному разряду аскоспоров в воздух. [ 13 ]
Насильственный сброс отдельных споров, называемых баллистоскорами, в себя образование небольшой капли воды ( капля Буллера ), что при контакте со спорой приводит к его выпуску снаряда с начальным ускорением более 10 000 г. включает [ 14 ] Другие грибы полагаются на альтернативные механизмы выпуска споры, такие как внешние механические силы, иллюстрируемые пуфтболами . Привлечение насекомых, таких как мухи, фрутинные структуры, в силу того, что у них живые цвета и гнилущий запах, для рассеивания грибковых споров - это еще одна стратегия, наиболее заметная используемая Стинкхорнс .
Было показано, что в общем плавном мхах ( Atrichum undulatum ) вибрация спорофита является важным механизмом высвобождения спор. [ 15 ]
сосудов, В случае сосудистых растений таких как папоротники, распределение ветра очень легких споров обеспечивает большую способность для рассеивания. Кроме того, споры менее подвержены хищничеству животных, чем семенам, потому что они почти не содержат пищевого заповедника; Однако они в большей степени подвержены грибкому и бактериальному хищничеству. Их главное преимущество состоит в том, что из всех форм потомства споры требуют наименьшей энергии и материалов для производства.
В Spikemoss selaginella lepidophylla рассеивание частично достигается необычным типом диаспора , планой . [ 16 ]
Источник
[ редактировать ]Споры были обнаружены в микрофоссиях, среднего уровня начиная с периода ордовика . [ 3 ] Две предполагаемые начальные функции споров связаны с тем, появлялись ли они до или после наземных растений. Гипотеза с тщательно изученными состоит в том, что споры были адаптацией ранних видов наземных растений, таких как эмбриофиты , которые позволяли растениям легко рассеиваться при адаптации к их некватической среде. [ 3 ] [ 17 ] Это особенно подтверждается наблюдением толстой стенки спор в криптоспорах . Эти стены споры защищали бы потенциальное потомство от новых погодных элементов. [ 3 ] что споры были ранним предшественником наземных растений и образовались во время ошибок при мейозе водорослей Вторая более недавняя гипотеза заключается в том , , предполагаемого раннего предка земельных растений. [ 18 ]
Было ли споры до или после наземных растений, их вклад в темы в таких областях, как палеонтология и филогенетика растений , был полезен. [ 18 ] Споры, найденные в микрофоссиях, также известных как Cryptospores, хорошо сохранились из -за фиксированного материала, в котором они находятся, а также того, насколько они обильны и широко распространены в течение их соответствующих периодов времени. Эти микрофоссили особенно полезны при изучении ранних периодов Земли, поскольку макрофоссили, такие как растения, не являются общими и не сохранились. [ 3 ] Как криптоспоры, так и современные споры имеют разнообразную морфологию, которая указывает на возможные условия окружающей среды более ранних периодов земли и эволюционные отношения видов растений. [ 3 ] [ 18 ] [ 17 ]
Галерея
[ редактировать ]-
 Споры Мосс Бартрамии Итехифилла . (Микроскопический вид, 400x)
Споры Мосс Бартрамии Итехифилла . (Микроскопический вид, 400x) -
 Разобитый папоротник спорангию. (Микроскопический вид, споры не видно)
Разобитый папоротник спорангию. (Микроскопический вид, споры не видно) -
 Споры и элатеры от хвоста. ( Equisetum , микроскопический вид)
Споры и элатеры от хвоста. ( Equisetum , микроскопический вид) -
 Споры ископаемого растения ( Scylaspora ) из силурийских отложений Швеции.
Споры ископаемого растения ( Scylaspora ) из силурийских отложений Швеции. -
 Фруктовая плесень со спор и различимый клеточный рост. (2000x)
Фруктовая плесень со спор и различимый клеточный рост. (2000x) -
 плесени слизи Споры кластеры, образованные внутри спорангии ретикулярной , из сосновых лесов Восточной Украины .
плесени слизи Споры кластеры, образованные внутри спорангии ретикулярной , из сосновых лесов Восточной Украины . -
 Внутренняя поверхность перидиума плесени слизи Tubifera Dudkae со спорами.
Внутренняя поверхность перидиума плесени слизи Tubifera Dudkae со спорами.







Смотрите также
[ редактировать ]Ссылки
[ редактировать ]- ^ Setlow, Peter; Джонсон, Эрик А. (30 апреля 2014 г.), Дойл, Майкл П.; Роберт Л. (ред.), значение» , Пищевая микробиология Pres Бьюкенен , Вашингтон, округ Колумбия, США : ASM , «Споры и их 978-1-68367-058-2 , Получено 13 декабря 2023 г.
- ^ «Веб -проект Tree of Life» . Архивировано из оригинала 5 февраля 2018 года . Получено 5 февраля 2018 года .
- ^ Jump up to: а беременный в дюймовый и фон глин час Веллман, г; Грей, Дж. (29 июня 2000 г.). «Микрофоссические записи ранних наземных растений» . Философские транзакции Королевского общества Лондона. Серия B, биологические науки . 355 (1398): 717–731, обсуждение 731–732. doi : 10.1098/rstb.2000.0612 . ISSN 0962-8436 . PMC 1692785 . PMID 10905606 .
- ^ Абель-Сантос, Эрнесто (2012). Бактериальные споры: текущие исследования и применения . Норфолк: Caister Academic Press. ISBN 978-1-908230-00-3 .
- ^ Иван Фиала (10 июля 2008 г.). "Myxozoa" . Веб -проект Tree of Life . Архивировано из оригинала 16 февраля 2015 года . Получено 14 января 2014 года .
Миксоспоры состоят из нескольких ячеек, которые преобразуются в оболочки, нематоцистаподобные полярные капсулы со спиральными экструдируемыми полярными филаментами и амебоидными инфекционными микробами.
- ^ «Диплоидная спора - обзор | темы ScienceDirect» . www.sciencedirect.com . Архивировано из оригинала 13 декабря 2023 года . Получено 13 декабря 2023 года .
- ^ «Биология микроспоридии» . 26 июня 2008 года. Архивировано с оригинала 26 июня 2008 года . Получено 24 марта 2024 года .
- ^ Jump up to: а беременный в Punt, W.; Hoen, pp; Blackmore, S.; Нильссон, С. и Ле Томас, А. (2007). «Глоссарий терминологии пыльцы и споры». Обзор палеоботании и палинологии . 143 (1): 1–81. Bibcode : 2007rpapa.143 .... 1p . doi : 10.1016/j.revpalbo.2006.06.008 .
- ^ Джадд, Уолтер С. и Олмстед, Ричард Г. (2004). «Обзор филогенетических отношений триколпата (Eudicot)» . Американский журнал ботаники . 91 (10): 1627–44. doi : 10.3732/ajb.91.10.1627 . PMID 21652313 .
- ^ Грей, Дж.; Шалонер, WG; Westoll, TS (1985). «Микрофоссические записи ранних наземных растений: достижения в понимании ранней земной земли, 1970–1984» . Философские транзакции Королевского общества б . 309 (1138): 167–195. BIBCODE : 1985RSPTB.309..167G . doi : 10.1098/rstb.1985.0077 . JSTOR 2396358 .
- ^ Wellman CH, Grey J (2000). «Микрофоссические записи ранних наземных растений» . Философские транзакции Королевского общества б . 355 (1398): 717–732. doi : 10.1098/rstb.2000.0612 . PMC 1692785 . PMID 10905606 .
- ^ Steemans, P.; Herisse, AL; Мелвин, Дж.; Миллер, Массачусетс; Париж, Ф.; Verniers, J.; Wellman, CH (2009). «Происхождение и радиация самых ранних сосудистых земельных растений» (PDF) . Наука . 324 (5925): 353. Bibcode : 2009Sci ... 324..353S . doi : 10.1126/science.1169659 . HDL : 1854/LU-697223 . ISSN 0036-8075 . PMID 19372423 . S2CID 206518080 . Архивировано (PDF) из оригинала 22 сентября 2017 года . Получено 1 ноября 2017 года .
- ^ Трейл Ф. (2007). «Грибковые пушки: взрывчатая спора в Аскомикоте» . Письма микробиологии FEMS . 276 (1): 12–8. doi : 10.1111/j.1574-6968.2007.00900.x . PMID 17784861 .
- ^ Прингл А., Патек С.Н., Фишер М., Стользе Дж., Деньги Н.П. (2005). «Захваченный запуск баллистоспоры». Микология . 97 (4): 866–71. doi : 10.3852/mycologia.97.4.866 . PMID 16457355 .
- ^ Johansson, Lönnell, Sundberg and Hylander (2014) выпускают пороговые значения для спор: важность турбулентности и длины спорофита. Журнал экологии, n/an/a.
- ^ «Ложная роза Иерихона - Selaginella lepidophyllafalse Rose of Jericho - Selaginella lepidophylla» . Руководство для растений и цветов . Февраль 2009 г. Архивировано из оригинала 15 июля 2011 года . Получено 1 февраля 2010 года .
- ^ Jump up to: а беременный Норем, WL (1958). «Ключи для классификации ископаемых споров и пыльцы» . Журнал палеонтологии . 32 (4): 666–676. ISSN 0022-3360 . JSTOR 1300785 .
- ^ Jump up to: а беременный в Стротер, Пол К.; Фостер, Клинтон (13 августа 2021 г.). «Окаменечная запись о происхождении земельных заводов из водорослей харофита» . Наука . 373 (6556): 792–796. Bibcode : 2021sci ... 373..792S . doi : 10.1126/science.abj2927 . ISSN 0036-8075 . PMID 34385396 .