Аналог нуклеиновой кислоты

Аналоги нуклеиновой кислоты — это соединения, которые аналогичны (структурно сходны) встречающимся в природе РНК и ДНК , используемым в медицине и исследованиях в области молекулярной биологии. Нуклеиновые кислоты представляют собой цепочки нуклеотидов, которые состоят из трех частей: фосфатного остова, пентозного сахара, рибозы или дезоксирибозы и одного из четырех нуклеиновых оснований . Аналог может иметь любое из этих изменений. [1] Обычно аналоговые нуклеиновые основания придают, среди прочего, различные свойства спаривания оснований и стэкинга оснований. Примеры включают универсальные основания, которые могут соединяться со всеми четырьмя каноническими основаниями, и аналоги фосфатно-сахарного остова, такие как PNA , которые влияют на свойства цепи (PNA может даже образовывать тройную спираль ). [2] Аналоги нуклеиновых кислот также называются ксенонуклеиновыми кислотами и представляют собой один из основных столпов ксенобиологии — создания новых для природы форм жизни на основе альтернативной биохимии.
Искусственные нуклеиновые кислоты включают пептид-нуклеиновые кислоты (ПНК), морфолино- и заблокированные нуклеиновые кислоты (LNA), а также гликолевые нуклеиновые кислоты (GNA), треозо-нуклеиновые кислоты (TNA) и гексит-нуклеиновые кислоты (HNA). Каждый из них отличается от встречающейся в природе ДНК или РНК изменениями в основной цепи молекулы. Однако полиэлектролитная теория гена предполагает, что для функционирования генетической молекулы требуется заряженный остов.
В мае 2014 года исследователи объявили, что они успешно внедрили два новых искусственных нуклеотида в бактериальную ДНК и, включив отдельные искусственные нуклеотиды в культуральную среду, смогли передать бактерии 24 раза; они не создали мРНК или белки, способные использовать искусственные нуклеотиды. Искусственные нуклеотиды имели два слитых ароматических кольца.
Лекарство
[ редактировать ]Некоторые аналоги нуклеозидов используются в качестве противовирусных или противораковых средств. Вирусная . полимераза включает эти соединения с неканоническими основаниями Эти соединения активируются в клетках путем преобразования в нуклеотиды. Их вводят в виде нуклеозидов, поскольку заряженные нуклеотиды не могут легко проникать через клеточные мембраны. [ нужна ссылка ]
Молекулярная биология
[ редактировать ]
Аналоги нуклеиновых кислот используются в молекулярной биологии для нескольких целей:
- Исследование возможных сценариев возникновения жизни: тестируя разные аналоги, исследователи пытаются ответить на вопрос, было ли использование ДНК и РНК в жизни выбрано с течением времени из-за ее преимуществ, или же они были выбраны случайно; [3]
- В качестве инструмента для обнаружения определенных последовательностей: XNA можно использовать для маркировки и идентификации широкого спектра компонентов ДНК и РНК с высокой специфичностью и точностью; [4]
- Показано, что как фермент, действующий на субстраты ДНК, РНК и XNA, XNA обладает способностью расщеплять и лигировать ДНК, РНК и другие молекулы XNA, аналогично действию РНК- рибозимов ; [3]
- Как средство, обладающее устойчивостью к гидролизу РНК ;
- Исследование механизмов действия фермента; и
- Исследование структурных особенностей нуклеиновых кислот.
Магистральные аналоги
[ редактировать ]Устойчивые к гидролизу РНК-аналоги
[ редактировать ]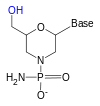
рибозы 2'-гидроксильная группа реагирует с фосфатно-связанной 3'-гидроксильной группой, что делает РНК слишком нестабильной, чтобы ее можно было надежно использовать или синтезировать. Чтобы преодолеть это, можно использовать аналог рибозы. Наиболее распространенными аналогами РНК являются 2'-О-метилзамещенная РНК, замкнутая нуклеиновая кислота (LNA) или мостиковая нуклеиновая кислота (BNA), морфолино , [5] [6] и пептид-нуклеиновая кислота ( ПНК ). Хотя эти олигонуклеотиды имеют другой сахар основной цепи — или, в случае ПНА, аминокислотный остаток вместо рибозофосфата — они все равно связываются с РНК или ДНК в соответствии с спариванием Уотсона и Крика, оставаясь при этом невосприимчивыми к активности нуклеазы. Их нельзя синтезировать ферментативно, и их можно получить только синтетически с использованием фосфорамидитной стратегии или, в случае ПНА, других методов синтеза пептидов . [ нужна ссылка ]
Другие известные аналоги, используемые в качестве инструментов.
[ редактировать ]Дидезоксинуклеотиды используются в секвенировании . Эти нуклеозидтрифосфаты содержат неканонический сахар дидезоксирибозу, в которой отсутствует 3'-гидроксильная группа, обычно присутствующая в ДНК, и поэтому она не может связываться со следующим основанием. Отсутствие 3'-гидроксильной группы прекращает цепную реакцию, поскольку ДНК-полимеразы принимают ее за обычный дезоксирибонуклеотид. Другой аналог с концевой цепью, в котором отсутствует 3'-гидроксил и имитирует аденозин, называется кордицепин . Кордицепин — противораковый препарат, нацеленный на репликацию РНК . Другим аналогом секвенирования является аналог нуклеиновых оснований, 7-деаза-GTP, который используется для секвенирования областей, богатых CG, вместо этого 7-деаза-АТФ называется туберцидином , антибиотиком. [ нужна ссылка ]
Предшественники мира РНК
[ редактировать ]Было высказано предположение, что миру РНК мог предшествовать «РНК-подобный мир», в котором существовали другие нуклеиновые кислоты с другим остовом, такие как GNA , PNA и TNA , однако доказательства этой гипотезы были названы «незначительными». . [7]
Базовые аналоги
[ редактировать ]Структура нуклеиновых оснований и номенклатура
[ редактировать ]
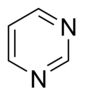
По своей структуре природные основания можно разделить на два класса:
- Пиримидины представляют собой шестичленные гетероциклы с атомами азота в положениях 1 и 3.
- Пурины являются бициклическими и состоят из пиримидина, присоединенного к имидазольному кольцу.
Искусственные нуклеотиды ( неестественные пары оснований (UBP), названные d5SICS UBP и dNaM UBP ) были вставлены в бактериальную ДНК, но эти гены не создавали матрицу мРНК и не индуцировали синтез белка. Искусственные нуклеотиды содержали два слитых ароматических кольца, которые образовывали комплекс (d5SICS–dNaM), имитирующий природную пару оснований (dG–dC). [8] [9] [10]
Мутагены
[ редактировать ]Одним из наиболее распространенных аналогов оснований является 5-бромурацил (5BU), аномальное основание, обнаруженное в мутагенном нуклеотидном аналоге BrdU. Когда нуклеотид, содержащий 5-бромурацил, включается в ДНК, он, скорее всего, образует пару с аденином; однако он может спонтанно перейти в другой изомер , который соединяется с другим нуклеиновым основанием , гуанином . Если это произойдет во время репликации ДНК, гуанин будет вставлен в качестве аналога противоположного основания, а при следующей репликации ДНК этот гуанин соединится с цитозином. Это приводит к изменению одной пары оснований ДНК, а именно к переходной мутации . [ нужна ссылка ]
Кроме того, азотистая кислота (HNO2) является мощным мутагеном, действующим на реплицирующуюся и нереплицирующуюся ДНК. Он может вызывать дезаминирование аминогрупп аденина, гуанина и цитозина. Аденин дезаминируется до гипоксантина , который образует пары оснований с цитозином вместо тимина. Цитозин дезаминируется до урацила, основания которого соединяются с аденином вместо гуанина. Дезаминирование гуанина не обладает мутагенным действием. Мутации, вызванные азотистой кислотой, также вызывают мутации обратно к дикому типу. [ нужна ссылка ]
Флуорофоры
[ редактировать ]
Обычно флуорофоры (такие как родамин или флуоресцеин ) связаны с кольцом, связанным с сахаром (в пара) через гибкое плечо, предположительно выступающее из основной канавки спирали. Из-за низкой процессивности нуклеотидов, связанных с объемистыми аддуктами, такими как флорофоры, с помощью [Taq-полимеразы], последовательность обычно копируется с использованием нуклеотида с плечом, а затем соединяется с реактивным флуорофором (непрямое мечение):
- Реакция на амины: аминоаллильные нуклеотиды содержат первичную аминогруппу на линкере, который реагирует с аминореактивным красителем, таким как цианин или красители Alexa Fluor , которые содержат реакционноспособную уходящую группу, такую как сукцинимидиловый эфир (NHS). Аминогруппы, спаривающиеся с основаниями, не затрагиваются.
- Тиол-реактивный: тиолсодержащие нуклеотиды реагируют с флуорофором, связанным с реакционноспособной уходящей группой, такой как малеимид.
- Биотин -связанные нуклеотиды основаны на том же принципе непрямого мечения (как и флуоресцентный стрептавидин) и используются в ДНК-чипах Affymetrix .
Флуорофоры находят множество применений в медицине и биохимии.
Флуоресцентные базовые аналоги
[ редактировать ]Наиболее часто используемый и коммерчески доступный аналог флуоресцентного основания, 2-аминопурин (2-AP), имеет высокий квантовый выход флуоресценции в свободном растворе (0,68), который значительно снижается (приблизительно в 100 раз, но сильно зависит от последовательности оснований) при включены в нуклеиновые кислоты. [11] Чувствительность 2-AP к излучению непосредственного окружения аналогична другим многообещающим и полезным аналогам флуоресцентных оснований, таким как 3-MI, 6-MI, 6-MAP, [12] пирроло-dC (также имеется в продаже), [13] модифицированные и улучшенные производные пирроло-дЦ, [14] фуран-модифицированные основания [15] и многие другие (см. недавние обзоры). [16] [17] [18] [19] [20] Эта чувствительность к микроокружению использовалась в исследованиях, например, структуры и динамики ДНК и РНК, динамики и кинетики взаимодействия ДНК-белок и переноса электронов внутри ДНК. [ нужна ссылка ]
Недавно разработанная и очень интересная группа аналогов флуоресцентных оснований, квантовый выход флуоресценции которых почти нечувствителен к их непосредственному окружению, представляет собой семейство трициклических цитозинов. 1,3-Диаза-2-оксофенотиазин tC имеет квантовый выход флуоресценции примерно 0,2 как в одноцепочечной, так и в двухцепочечной форме независимо от окружающих оснований. [21] [22] Также оксо-гомолог tC называется tC. ТО (оба коммерчески доступны), 1,3-диаза-2-оксофеноксазин имеет квантовый выход 0,2 в двухцепочечных системах. [23] Однако он несколько чувствителен к окружающим основаниям в однонитях (квантовый выход 0,14–0,41). Высокие и стабильные квантовые выходы этих аналогов оснований делают их очень яркими, а в сочетании с их хорошими свойствами аналогов оснований (оставляя структуру и стабильность ДНК практически нетронутыми) они особенно полезны при измерениях анизотропии флуоресценции и FRET, в областях, где другие аналоги с флуоресцентной основой менее точны. аналог основания акцептора FRET tC nitro . Кроме того, в том же семействе аналогов цитозина был разработан [24] Совместно с ТК ТО в качестве донора FRET это первая когда-либо разработанная пара FRET-аналога нуклеиновых кислот. Семейство tC, например, использовалось в исследованиях, связанных с механизмами связывания ДНК-полимеразы и полимеризации ДНК.
Естественные неканонические базы
[ редактировать ]В клетке присутствует несколько неканонических оснований: CpG-островки в ДНК (часто метилированные), все эукариотические мРНК (кэпированные метил-7-гуанозином) и несколько оснований рРНК (метилированные). Часто тРНК сильно модифицируются посттранскрипционно, чтобы улучшить их конформацию или спаривание оснований, в частности, в антикодоне или рядом с ним: инозин может образовывать пары оснований с C, U и даже с A, тогда как тиуридин (с A) более специфичен, чем урацил. (с пурином). [25] Другими распространенными модификациями оснований тРНК являются псевдоуридин (который дал название петле TΨC ), дигидроуридин (который не суммируется, поскольку он не ароматичен), кевозин, виозин и т. д. Тем не менее, все это модификации обычных оснований и не размещаются полимеразой. [25]
Спаривание оснований
[ редактировать ]Канонические основания могут иметь либо карбонильную, либо аминогруппу на атомах углерода, окружающих атом азота, наиболее удаленный от гликозидной связи, что позволяет им образовывать пары оснований (спаривание оснований Уотсона-Крика) через водородные связи (амин с кетоном, пурин с пиримидином). . Аденин и 2-аминоаденин имеют одну/две аминогруппы, тогда как тимин имеет две карбонильные группы, а цитозин и гуанин представляют собой смешанные амино-карбонильные группы (инвертированные по отношению друг к другу). [ нужна ссылка ]
| Натуральные пары оснований | |
|---|---|
 |  |
| Пара оснований GC: пуриновый карбонил/амин образует три межмолекулярные водородные связи с пиримидинамином/карбонилом. | Пара оснований АТ: пуринамин/- образует две межмолекулярные водородные связи с пиримидинкарбонилом/карбонилом. |
Точная причина того, почему существует только четыре нуклеотида, обсуждается, но есть несколько неиспользованных возможностей.Более того, аденин не является наиболее стабильным выбором для спаривания оснований: в Cyanophage S-2L диаминопурин (DAP). вместо аденина используется [26] Диаминопуриновые пары оснований идеально сочетаются с тимином, поскольку он идентичен аденину, но имеет аминную группу в положении 2, образующую 3 внутримолекулярные водородные связи, что устраняет основное различие между двумя типами пар оснований (слабая АТ и сильная CG). Эта улучшенная стабильность влияет на взаимодействия связывания белков, которые основаны на этих различиях.Другая комбинация включает в себя:
- Изогуанин и изоцитозин, у которых амин и кетон инвертированы по сравнению со стандартными гуанином и цитозином. Вероятно, они не используются, поскольку таутомеры проблематичны для спаривания оснований, но isoC и isoG можно правильно амплифицировать с помощью ПЦР даже в присутствии 4 канонических оснований. [27]
- Диаминопиримидин и ксантин, которые связываются подобно 2-аминоаденину и тимину, но имеют инвертированную структуру. Эта пара не используется, поскольку ксантин является продуктом дезаминирования.
| Неиспользованные схемы базовых пар | ||
|---|---|---|
 | 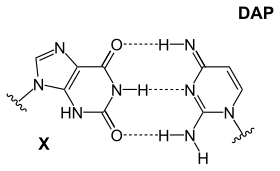 |  |
| Основание DAP-T: пуринамин/амин образует три межмолекулярные водородные связи с пиримидинкетоном/кетоном. | Основание X-DAP: пуриновый кетон/кетон образует три межмолекулярные водородные связи с пиримидинамином/амином. | Основание iG-iC: пуриновый амин/кетон образует три межмолекулярные водородные связи с пиримидиновым кетоном/амином. |
Однако правильная структура ДНК может сформироваться, даже если основания не спарены посредством водородных связей; то есть пары оснований складываются благодаря гидрофобности, как показали исследования с изостерами ДНК (аналогами с таким же количеством атомов), такими как аналог тимина 2,4-дифтортолуол (F) или аналог аденина 4-метилбензимидазол (Z). [28] Альтернативной гидрофобной парой могут быть изохинолин и пирроло[2,3-b]пиридин. [29]
Другие заслуживающие внимания пары оснований:
- Также было создано несколько флуоресцентных оснований, таких как пара оснований 2-амино-6-(2-тиенил)пурин и пиррол-2-карбальдегид. [30]
- Металл-координированные основания, такие как соединение пиридин-2,6-дикарбоксилата (тридентатный лиганд) и пиридина (монодентатный лиганд) посредством плоской квадратной координации с центральным ионом меди. [31]
- Универсальные основания могут соединяться без разбора с любым другим основанием, но, как правило, значительно снижают температуру плавления последовательности; примеры включают производные 2'-дезоксиинозина (гипоксантиндезоксинуклеотид), аналоги нитроазола и гидрофобные ароматические основания, не имеющие водородных связей (сильные эффекты суммирования). Они используются в качестве доказательства концепции и, как правило, не используются в вырожденных праймерах (которые представляют собой смесь праймеров).
- , количество возможных пар оснований удваивается xDNA Если рассматривать . xДНК содержит расширенные основания, в которые добавлено бензольное кольцо, которое может образовывать пары с каноническими основаниями, в результате чего образуются четыре дополнительные возможные пары оснований (xA-T, xT-A, xC-G, xG-C) с восемью основаниями (xA-T, xT-A, xC-G, xG-C). или 16 оснований, если используются неиспользованные расстановки). Другой формой оснований с добавлением бензола является yDNA, в которой основание расширяется за счет бензола. [32]
| Новые пары оснований с особыми свойствами | ||
|---|---|---|
 |  |  |
| Основание FZ: метилбензимидазол не образует межмолекулярных водородных связей с толуолом F/F. | Основание S-Pa: пуринтиенил/амин образует три межмолекулярные водородные связи с пирролом/карбальдегидом. | База xA-T: такое же соединение, как и у AT. |
Металлические пары оснований
[ редактировать ]При спаривании металлических оснований водородные связи Уотсона-Крика заменяются взаимодействием иона металла с нуклеозидами, выступающими в качестве лигандов. Возможные геометрии металла, которые позволили бы образовать дуплекс с двумя бидентатными нуклеозидами вокруг центрального атома металла, являются тетраэдрическими , додекаэдрическими и плоскими квадратами . Комплексообразование металлов с ДНК может происходить путем образования неканонических пар оснований из природных азотистых оснований с участием ионов металлов, а также путем обмена атомов водорода, входящих в пару оснований Уотсона-Крика, на ионы металлов. [33] Было показано, что введение ионов металлов в дуплекс ДНК обладает потенциальным магнитным потенциалом. [34] или проведение недвижимости, [35] а также повышенная стабильность. [36]
Было показано, что образование комплексов металлов происходит между природными азотистыми основаниями . Хорошо документированным примером является образование T-Hg-T, в котором участвуют два депротонированных нуклеиновых основания тимина , которые соединяются Hg. 2+ и образует связанную пару металл-основание. [37] Этот мотив не учитывает накопленную ртуть. 2+ в дуплексе из-за процесса образования внутринитевой шпильки, который предпочтительнее образования дуплекса. [38] Два тимина, расположенные напротив друг друга, не образуют пару оснований Уотсона-Крика в дуплексе; это пример, когда несоответствие пар оснований Уотсона-Крика стабилизируется за счет образования пары металл-основание. Другим примером комплексообразования металлов с природными азотистыми основаниями является образование A-Zn-T и G-Zn-C при высоком pH; Ко +2 и Ни +2 также образуют эти комплексы. Это пары оснований Уотсона-Крика, в которых двухвалентный катион координирован с азотистыми основаниями. Точная привязка обсуждается. [39]
Было разработано большое разнообразие искусственных азотистых оснований для использования в качестве пар металлических оснований. Эти модифицированные нуклеиновые основания обладают настраиваемыми электронными свойствами, размерами и сродством связывания, которые можно оптимизировать для конкретного металла. Например, было показано, что нуклеозид, модифицированный пиридин-2,6-дикарбоксилатом, прочно связывается с Cu. 2+ , тогда как другие двухвалентные ионы связаны слабо. Трехзубчатый характер способствует этой избирательности. Четвертое координационное место меди насыщено противоположно расположенным пиридиновым нуклеиновым основанием. [40] Асимметричная система спаривания металлических оснований ортогональна парам оснований Уотсона-Крика. Другим примером искусственного азотистого основания является азотистое основание гидроксипиридона, которое способно связывать Cu. 2+ внутри дуплекса ДНК. Пять последовательных пар оснований медь-гидроксипиридон были включены в двойную цепь, фланкированную только одним природным азотистым основанием на обоих концах. Данные ЭПР показали, что расстояние между медными центрами оценивается в 3,7 ± 0,1 Å, тогда как природный дуплекс ДНК B-типа лишь немного больше (3,4 Å). [41] Привлекательность укладки ионов металлов внутри дуплекса ДНК связана с надеждой получить наноскопические самоорганизующиеся металлические проволоки, хотя это еще не реализовано.
Неестественная пара оснований (UBP)
[ редактировать ]Неестественная пара оснований (UBP) — это спроектированная субъединица (или азотистое основание ) ДНК , созданная в лаборатории и не встречающаяся в природе. В 2012 году группа американских ученых во главе с Флойдом Ромесбергом, химиком-биологом из Исследовательского института Скриппса в Сан-Диего, Калифорния, опубликовала, что его команда разработала две неестественные пары оснований. названы d5SICS и dNaM . [42] С технической точки зрения, эти искусственные нуклеотиды , несущие гидрофобные нуклеиновые основания, имеют два слитых ароматических кольца , которые образуют комплекс d5SICS-dNaM или пару оснований в ДНК. [10] [43] В 2014 году та же команда сообщила, что они синтезировали плазмиду , содержащую природные пары оснований TA и CG, а наиболее эффективная лаборатория UBP Ромесберга разработала и вставила ее в клетки обычной бактерии E. coli , которая успешно воспроизвела неестественное основание. пары через несколько поколений. [44] Это первый известный пример передачи живым организмом расширенного генетического кода последующим поколениям. [10] [45] Частично это было достигнуто за счет добавления вспомогательного гена водорослей, который экспрессирует транспортер нуклеотид-трифосфата , который эффективно импортирует трифосфаты как d5SICSTP, так и dNaMTP в E. coli . бактерии [10] Затем естественные пути репликации бактерий используют их для точной репликации плазмиды, содержащей d5SICS-dNaM. [ нужна ссылка ]
Успешное включение третьей пары оснований является значительным прорывом на пути к значительному расширению числа аминокислот , которые могут кодироваться ДНК, с существующих 20 аминокислот до теоретически возможных 172, тем самым расширяя возможности живых организмов производить новые белки . [44] Ранее искусственные нити ДНК ничего не кодировали, но ученые предположили, что их можно спроектировать для производства новых белков, которые могут найти промышленное или фармацевтическое применение. [46] Транскрипция ДНК, содержащей неприродные пары оснований, и трансляция соответствующей мРНК были фактически достигнуты недавно. В ноябре 2017 года та же команда из Научно-исследовательского института Скриппса , которая впервые ввела два дополнительных азотистых основания в бактериальную ДНК, сообщила о создании полусинтетической бактерии E. coli , способной производить белки с использованием такой ДНК. Его ДНК содержала шесть различных азотистых оснований : четыре канонических и два искусственно добавленных, dNaM и dTPT3 (эти два образуют пару). У бактерий было два соответствующих основания РНК, включенных в два новых кодона, дополнительные тРНК, узнающие эти новые кодоны (эти тРНК также содержали два новых основания РНК внутри своих антикодонов) и дополнительные аминокислоты, позволяющие бактериям синтезировать «неестественные» белки. [47] [48]
Еще одна демонстрация UBP была проведена группой Ичиро Хирао в институте RIKEN в Японии. В 2002 году они разработали неприродную пару оснований между 2-амино-8-(2-тиенил)пурином (s) и пиридин-2-оном (y), которая действует in vitro в транскрипции и трансляции, для сайт-специфического включения нестандартные аминокислоты в белки. [49] В 2006 году они создали 7-(2-тиенил)имидазо[4,5-b]пиридин (Ds) и пиррол-2-карбальдегид (Pa) в качестве третьей пары оснований для репликации и транскрипции. [50] Впоследствии Ds и 4-[3-(6-аминогексанамидо)-1-пропинил]-2-нитропиррол (Px) были обнаружены как пара с высокой точностью при ПЦР-амплификации. [51] [52] В 2013 году они применили пару Ds-Px для генерации аптамеров ДНК путем селекции in vitro (SELEX) и продемонстрировали, что расширение генетического алфавита значительно увеличивает сродство аптамеров ДНК к белкам-мишеням. [53]
Ортогональная система
[ редактировать ]Была предложена и изучена как теоретически, так и экспериментально возможность реализации ортогональной системы внутри клеток, независимой от клеточного генетического материала, с целью создания полностью безопасной системы. [54] с возможным увеличением кодирующих потенциалов. [55] Несколько групп сосредоточили внимание на различных аспектах:
- Новые основные цепи и пары оснований, как обсуждалось выше;
- XNA- полимеразы искусственной репликации и транскрипции, обычно начинающиеся с РНК-полимеразы Т7 ; [56]
- ( 16S рибосомальные последовательности с измененными последовательностями анти-Шайна-Дальгарно, позволяющие транслировать только ортогональную мРНК с соответствующей измененной последовательностью Шайна-Дальгарно; [57] и
- Новая тРНК, кодирующая неприродные аминокислоты для расширенного генетического кода .
См. также
[ редактировать ]- Биотин
- Темный гаситель
- Дезоксирибозим
- Расширенный генетический код
- флюорофор
- Генетика
- Молекулярная биология
- Нуклеиновая кислота
- нуклеиновое основание
- нуклеозид
- Нуклеотид
- Синтез олигонуклеотидов
- Рибозим
- Синтетическая биология
- Ксенобиология
- хДНК
- ДНК Хатимодзи
- Искусственно расширенная генетическая информационная система (AEGIS)
- Ксенонуклеиновая кислота
Ссылки
[ редактировать ]- ^ Певица Э (19 июля 2015 г.). «Химики изобретают новые буквы для генетического алфавита природы» . Проводной . Проверено 20 июля 2015 г.
- ^ Петерссон Б., Нильсен Б.Б., Расмуссен Х., Ларсен И.К., Гайхеде М., Нильсен П.Е., Каструп Дж.С. (февраль 2005 г.). «Кристаллическая структура частично самокомплементарного олигомера пептидной нуклеиновой кислоты (ПНК), демонстрирующая дуплекс-триплексную сеть». Журнал Американского химического общества . 127 (5): 1424–30. дои : 10.1021/ja0458726 . ПМИД 15686374 .
- ^ Перейти обратно: а б Тейлор А.И., Пиньейро В.Б., Смола М.Дж., Моргунов А.С., Пик-Чью С., Козенс С., Уикс К.М., Хердевейн П., Холлигер П. (февраль 2015 г.). «Катализаторы из синтетических генетических полимеров» . Природа . 518 (7539): 427–30. Бибкод : 2015Natur.518..427T . дои : 10.1038/nature13982 . ПМЦ 4336857 . ПМИД 25470036 .
- ^ Ван Ц, Чен Л, Лун Ю, Тянь Х, Ву Дж (2013). «Молекулярные маяки ксенонуклеиновой кислоты для обнаружения нуклеиновой кислоты» . Тераностика . 3 (6): 395–408. дои : 10.7150/thno.5935 . ПМЦ 3677410 . ПМИД 23781286 .
- ^ Саммертон Дж., Веллер Д. (июнь 1997 г.). «Морфолиновые антисмысловые олигомеры: дизайн, получение и свойства». Разработка лекарств на основе антисмысловых и нуклеиновых кислот . 7 (3): 187–95. дои : 10.1089/oli.1.1997.7.187 . ПМИД 9212909 . S2CID 19372403 .
- ^ Саммертон Дж (декабрь 1999 г.). «Морфолино-антисмысловые олигомеры: случай структурного типа, независимого от РНКазы H». Biochimica et Biophysical Acta (BBA) – Структура и экспрессия генов . 1489 (1): 141–58. дои : 10.1016/s0167-4781(99)00150-5 . ПМИД 10807004 .
- ^ Робертсон, член парламента; Джойс, GF (01 мая 2012 г.). «Происхождение мира РНК» . Перспективы Колд-Спринг-Харбор в биологии . 4 (5): а003608. doi : 10.1101/cshperspect.a003608 . ISSN 1943-0264 . ПМК 3331698 . ПМИД 20739415 .
- ^ Поллак А (7 мая 2014 г.). «Исследователи сообщают о прорыве в создании искусственного генетического кода» . Нью-Йорк Таймс . Проверено 7 мая 2014 г.
- ^ Каллауэй Э (7 мая 2014 г.). «Первая жизнь с «чужой» ДНК». Природа . дои : 10.1038/nature.2014.15179 . S2CID 86967999 .
- ^ Перейти обратно: а б с д Малышев Д.А., Дхами К., Лавернь Т., Чен Т., Дай Н., Фостер Дж.М., Корреа И.Р., Ромесберг Ф.Е. (май 2014 г.). «Полусинтетический организм с расширенным генетическим алфавитом» . Природа . 509 (7500): 385–88. Бибкод : 2014Natur.509..385M . дои : 10.1038/nature13314 . ПМК 4058825 . ПМИД 24805238 .
- ^ Уорд, округ Колумбия, Райх Э., Страйер Л. (март 1969 г.). «Флуоресцентные исследования нуклеотидов и полинуклеотидов. I. Формицин, 2-аминопуринрибозид, 2,6-диаминопуринрибозид и их производные» . Журнал биологической химии . 244 (5): 1228–37. дои : 10.1016/S0021-9258(18)91833-8 . ПМИД 5767305 .
- ^ Хокинс М.Э. (2001). «Флуоресцентные аналоги нуклеозидов птеридина: окно во взаимодействия ДНК» . Клеточная биохимия и биофизика . 34 (2): 257–81. дои : 10.1385/cbb:34:2:257 . ПМИД 11898867 . S2CID 12134698 .
- ^ Берри Д.А., Юнг К.Ю., Уайз Д.С., Серсель А.Д., Пирсон В.Х., Маки Х., Рэндольф Дж.Б., Сомерс Р.Л. (2004). «Пирроло-дЦ и пирроло-С: флуоресцентные аналоги цитидина и 2'-дезоксицитидина для изучения олигонуклеотидов». Тетраэдр Летт . 45 (11): 2457–61. дои : 10.1016/j.tetlet.2004.01.108 .
- ^ Войцеховский Ф., Хадсон Р.Х. (сентябрь 2008 г.). «Флуоресцентные и гибридизационные свойства пептидной нуклеиновой кислоты, содержащей замещенный фенилпирролоцитозин, предназначенный для взаимодействия гуанина с дополнительной H-связью». Журнал Американского химического общества . 130 (38): 12574–75. дои : 10.1021/ja804233g . ПМИД 18761442 .
- ^ Греко, Нью-Джерси, Тор Ю (август 2005 г.). «Простые флуоресцентные аналоги пиримидина обнаруживают наличие абазовых участков ДНК». Журнал Американского химического общества . 127 (31): 10784–85. дои : 10.1021/ja052000a . ПМИД 16076156 .
- ^ Рист М.Дж., Марино Дж.П. (2002). «Флуоресцентные аналоги нуклеотидных оснований как зонды структуры, динамики и взаимодействия нуклеиновых кислот». Курс. Орг. Хим . 6 (9): 775–93. дои : 10.2174/1385272023373914 .
- ^ Уилсон Дж. Н., Кул ET (декабрь 2006 г.). «Замены оснований флуоресцентной ДНК: репортеры и сенсоры для биологических систем». Органическая и биомолекулярная химия . 4 (23): 4265–74. дои : 10.1039/b612284c . ПМИД 17102869 .
- ^ Вильгельмссон и Тор (2016). Флуоресцентные аналоги биомолекулярных строительных блоков: конструкция и применение . Нью-Джерси: Уайли. ISBN 978-1-118-17586-6 .
- ^ Вильгельмссон Л.М. (май 2010 г.). «Флуоресцентные аналоги оснований нуклеиновых кислот» . Ежеквартальные обзоры биофизики . 43 (2): 159–83. дои : 10.1017/s0033583510000090 . ПМИД 20478079 . S2CID 10783202 .
- ^ Синкелдам Р.В., Греко, Нью-Джерси, Тор Ю (май 2010 г.). «Флуоресцентные аналоги биомолекулярных строительных блоков: конструкция, свойства и применение» . Химические обзоры . 110 (5): 2579–619. дои : 10.1021/cr900301e . ПМЦ 2868948 . ПМИД 20205430 .
- ^ Вильгельмссон Л.М., Холмен А., Линкольн П., Нильсен П.Е., Норден Б. (2001). «Высокофлуоресцентный аналог оснований ДНК, который образует пары оснований Уотсона-Крика с гуанином». Дж. Ам. хим. Соц . 123 (10): 2434–35. дои : 10.1021/ja0025797 . ПМИД 11456897 .
- ^ Сандин П., Вильгельмссон Л.М., Линкольн П., Пауэрс В.Е., Браун Т., Альбинссон Б. (2005). «Флуоресцентные свойства аналога основания ДНК tC при включении в ДНК – незначительное влияние соседних оснований на квантовый выход флуоресценции» . Исследования нуклеиновых кислот . 33 (16): 5019–25. дои : 10.1093/nar/gki790 . ПМК 1201328 . ПМИД 16147985 .
- ^ Сандин П., Бёрьессон К., Ли Х., Мортенссон Дж., Браун Т., Вильгельмссон Л.М., Альбинссон Б. (январь 2008 г.). «Охарактеризация и использование беспрецедентно яркого и структурно ненарушающего флуоресцентного аналога основания ДНК» . Исследования нуклеиновых кислот . 36 (1): 157–67. дои : 10.1093/нар/gkm1006 . ПМК 2248743 . ПМИД 18003656 .
- ^ Бёрьессон К., Преус С., Эль-Сагир А.Х., Браун Т., Альбинссон Б., Вильгельмссон Л.М. (апрель 2009 г.). «Аналог оснований нуклеиновых кислот FRET-пара, облегчающая детальные структурные измерения в системах, содержащих нуклеиновые кислоты» . Журнал Американского химического общества . 131 (12): 4288–93. дои : 10.1021/ja806944w . ПМИД 19317504 .
- ^ Перейти обратно: а б Родригес-Эрнандес А., Спирс Дж.Л., Гастон К.В., Лимбах П.А., Гампер Х., Хоу Ю.М., Кайзер Р., Агрис П.Ф., Перона Дж.Дж. (октябрь 2013 г.). «Структурная и механистическая основа повышения эффективности трансляции с помощью 2-тиуридина в положении колебания антикодона тРНК» . Журнал молекулярной биологии . 425 (20): 3888–906. дои : 10.1016/j.jmb.2013.05.018 . ПМК 4521407 . ПМИД 23727144 .
- ^ Кирнос М.Д., Худяков И.Ю., Александрушкина Н.И., Ванюшин Б.Ф. (ноябрь 1977 г.). «2-аминоаденин представляет собой аденин, заменяющий основание в ДНК цианофага S-2L». Природа . 270 (5635): 369–70. Бибкод : 1977Natur.270..369K . дои : 10.1038/270369a0 . ПМИД 413053 . S2CID 4177449 .
- ^ Джонсон С.К., Шерил С.Б., Маршалл Д.Д., Мозер М.Дж., Прудент Дж.Р. (2004). «Третья пара оснований для полимеразной цепной реакции: вставка isoC и isoG» . Исследования нуклеиновых кислот . 32 (6): 1937–41. дои : 10.1093/nar/gkh522 . ПМК 390373 . ПМИД 15051811 .
- ^ Танигучи Ю., Кул ET (июль 2007 г.). «Неполярные изостеры поврежденных оснований ДНК: эффективная имитация мутагенных свойств 8-оксопуринов». Журнал Американского химического общества . 129 (28): 8836–44. дои : 10.1021/ja071970q . ПМИД 17592846 .
- ^ Хван Г.Т., Ромесберг FE (ноябрь 2008 г.). «Репертуар неестественных субстратов ДНК-полимераз семейств A, B и X» . Журнал Американского химического общества . 130 (44): 14872–82. дои : 10.1021/ja803833h . ПМЦ 2675700 . ПМИД 18847263 .
- ^ Кимото М, Мицуи Т, Харада Й, Сато А, Ёкояма С, Хирао И (2007). «Флуоресцентное зондирование молекул РНК с помощью неестественной системы пар оснований» . Исследования нуклеиновых кислот . 35 (16): 5360–69. дои : 10.1093/нар/gkm508 . ПМК 2018647 . ПМИД 17693436 .
- ^ Этвелл, Шейн; Меггерс, Эрик; Спраггон, Глен; Шульц, Питер Г. (декабрь 2001 г.). «Структура медь-опосредованной пары оснований в ДНК» . Журнал Американского химического общества . 123 (49): 12364–12367. дои : 10.1021/ja011822e . ISSN 0002-7863 . ПМИД 11734038 .
- ^ Лю Х., Гао Дж., Линч С.Р., Сайто Ю.Д., Мейнард Л., Кул Э.Т. (октябрь 2003 г.). «Парная генетическая спираль из четырех оснований увеличенного размера». Наука . 302 (5646): 868–71. Бибкод : 2003Sci...302..868L . дои : 10.1126/science.1088334 . ПМИД 14593180 . S2CID 37244007 .
- ^ Веттиг С.Д., Ли Дж.С. (2003). «Термодинамическое исследование М-ДНК: новый комплекс ион металла-ДНК». Журнал неорганической биохимии . 94 (1–2): 94–99. дои : 10.1016/S0162-0134(02)00624-4 . ПМИД 12620678 .
- ^ Чжан Х.И., Кальцолари А., Ди Феличе Р. (август 2005 г.). «О магнитном выравнивании ионов металлов в двойной спирали, имитирующей ДНК». Журнал физической химии Б. 109 (32): 15345–48. дои : 10.1021/jp052202t . ПМИД 16852946 .
- ^ Айх П., Скиннер Р.Дж., Веттиг С.Д., Стир Р.П., Ли Дж.С. (август 2002 г.). «Поведение молекулярных проводов на больших расстояниях в металлическом комплексе ДНК». Журнал биомолекулярной структуры и динамики . 20 (1): 93–98. дои : 10.1080/07391102.2002.10506826 . ПМИД 12144356 . S2CID 41568646 .
- ^ Клевер Г.Х., Полборн К., Карелл Т. (2005). «Пара оснований металл-сален с высокой степенью стабилизации дуплекса ДНК». Энджью. хим. Эд . 117 (44): 7370–74. Бибкод : 2005АнгЧ.117.7370С . дои : 10.1002/anie.200501589 .
- ^ Бансел Э., Бун С., Джоли Х., Кумар Р., Норрис А.Р. (1985). «Взаимодействия ионов металлов с биомолекулами. XII. ЯМР 1H и 13C свидетельствуют о предпочтительной реакции тимидина над гуанозином в реакциях обмена и конкуренции с ртутью (II) и метилртутью (II)». Неорг. Биохим . 25 : 61–73. дои : 10.1016/0162-0134(85)83009-9 .
- ^ Оно А, Тогаши Х (август 2004 г.). «Высокоселективный сенсор ртути(II) на основе олигонуклеотидов в водных растворах». Ангеванде Хеми . 43 (33): 4300–02. дои : 10.1002/anie.200454172 . ПМИД 15368377 .
- ^ Меггерс Э., Холланд П.Л., Толман В.Б., Ромесберг Ф.Е., Шульц П.Г. (2000). «Новая пара оснований ДНК, опосредованная медью». Дж. Ам. хим. Соц . 122 (43): 10714–15. дои : 10.1021/ja0025806 .
- ^ Ли Дж.С., Латимер Л.Дж., Рид Р.С. (1993). «Совместное конформационное изменение дуплексной ДНК, индуцированное Zn2+ и ионами других двухвалентных металлов». Биохимия и клеточная биология . 71 (3–4): 162–68. дои : 10.1139/o93-026 . ПМИД 8398074 .
- ^ Танака К., Тенгейджи А., Като Т., Тояма Н., Сионоя М. (февраль 2003 г.). «Дискретный самособирающийся металлический массив в искусственной ДНК». Наука . 299 (5610): 1212–13. Бибкод : 2003Sci...299.1212T . дои : 10.1126/science.1080587 . ПМИД 12595687 . S2CID 22413126 .
- ^ Малышев Д.А., Дхами К., Куах Х.Т., Лавернь Т., Ордуханян П., Торкамани А., Ромесберг Ф.Е. (июль 2012 г.). «Эффективная и независимая от последовательности репликация ДНК, содержащей третью пару оснований, создает функциональный шестибуквенный генетический алфавит» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (30): 12005–10. Бибкод : 2012PNAS..10912005M . дои : 10.1073/pnas.1205176109 . ПМК 3409741 . ПМИД 22773812 .
- ^ Каллауэй Э (7 мая 2014 г.). «Ученые создали первый живой организм с «искусственной» ДНК» . Новости природы . Хаффингтон Пост . Проверено 8 мая 2014 г.
- ^ Перейти обратно: а б Файкс Би Джей (8 мая 2014 г.). «Жизнь создана с использованием расширенного генетического кода» . Сан-Диего Юнион Трибьюн . Проверено 8 мая 2014 г.
- ^ Образец I (7 мая 2014 г.). «Первые формы жизни, передавшие искусственную ДНК, созданную американскими учеными» . Хранитель . Проверено 8 мая 2014 г.
- ^ Поллак А (7 мая 2014 г.). «Ученые добавляют буквы в алфавит ДНК, вселяя надежду и страх» . Нью-Йорк Таймс . Проверено 8 мая 2014 г.
- ^ Чжан Й., Птацин Дж.Л., Фишер Э.К., Эрни Х.Р., Каффаро С.Э., Сан-Хосе К., Фельдман А.В., Тернер Ч.Р., Ромесберг Ф.Е. (2017). «Полусинтетический организм, который хранит и извлекает увеличенную генетическую информацию» . Природа . 551 (7682): 644–47. Бибкод : 2017Natur.551..644Z . дои : 10.1038/nature24659 . ПМЦ 5796663 . ПМИД 29189780 .
- ^ «Неестественный» микроб может производить белки . Новости Би-би-си . 29 ноября 2017 г.
- ^ Хирао И, Оцуки Т, Фудзивара Т, Мицуи Т, Ёкогава Т, Окуни Т, Накаяма Х, Такио К, Ябуки Т, Кигава Т, Кодама К, Ёкогава Т, Нисикава К, Ёкояма С (февраль 2002 г.). пара для включения аналогов аминокислот в белки». Nature Biotechnology . 20 (2): 177–82. : 10.1038 /nbt0202-177 . PMID 11821864. . S2CID 22055476 doi
- ^ Хирао И., Кимото М., Мицуи Т., Фудзивара Т., Каваи Р., Сато А., Харада Ю., Ёкояма С. (сентябрь 2006 г.). «Неестественная гидрофобная система пар оснований: сайт-специфическое включение аналогов нуклеотидов в ДНК и РНК». Природные методы . 3 (9): 729–35. дои : 10.1038/nmeth915 . ПМИД 16929319 . S2CID 6494156 .
- ^ Кимото М., Каваи Р., Мицуи Т., Ёкояма С., Хирао И. (февраль 2009 г.). «Неестественная система пар оснований для эффективной ПЦР-амплификации и функционализации молекул ДНК» . Исследования нуклеиновых кислот . 37 (2): е14. дои : 10.1093/нар/gkn956 . ПМЦ 2632903 . ПМИД 19073696 .
- ^ Ямашигэ Р., Кимото М., Такезава Ю., Сато А., Мицуи Т., Ёкояма С., Хирао И. (март 2012 г.). «Высокоспецифичные системы неприродных пар оснований в качестве третьей пары оснований для ПЦР-амплификации» . Исследования нуклеиновых кислот . 40 (6): 2793–806. дои : 10.1093/nar/gkr1068 . ПМЦ 3315302 . ПМИД 22121213 .
- ^ Кимото М., Ямасигэ Р., Мацунага К., Ёкояма С., Хирао И. (май 2013 г.). «Поколение аптамеров ДНК с высоким сродством с использованием расширенного генетического алфавита». Природная биотехнология . 31 (5): 453–57. дои : 10.1038/nbt.2556 . ПМИД 23563318 . S2CID 23329867 .
- ^ Шмидт М. «Ксенобиология: новая форма жизни как лучший инструмент биобезопасности» Bioessays Vol 32 (4): 322–31.
- ^ Хердевейн П., Марльер П. (июнь 2009 г.). «На пути к безопасным генетически модифицированным организмам посредством химической диверсификации нуклеиновых кислот». Химия и биоразнообразие . 6 (6): 791–808. дои : 10.1002/cbdv.200900083 . ПМИД 19554563 . S2CID 8572188 .
- ^ Синкай А., Патель П.Х., Леб Л.А. (июнь 2001 г.). «Консервативный мотив А ДНК-полимеразы I Escherichia coli сильно изменчив» . Журнал биологической химии . 276 (22): 18836–42. дои : 10.1074/jbc.M011472200 . ПМИД 11278911 .
- ^ Рэкхэм О, Чин Дж.В. (август 2005 г.). «Сеть пар ортогональных рибосом х мРНК». Химическая биология природы . 1 (3): 159–66. дои : 10.1038/nchembio719 . ПМИД 16408021 . S2CID 37181098 .
Виды нуклеиновых кислот | |||||||
|---|---|---|---|---|---|---|---|
| Составляющие | |||||||
| Рибонуклеиновые кислоты (кодирование, некодирование ) |
| ||||||
| Дезоксирибонуклеиновая кислоты | |||||||
| Аналоги | |||||||
| Клонирование векторов | |||||||