Спора


В биологии спора — это единица полового (у грибов) или бесполого размножения , которая может быть приспособлена для распространения и выживания, часто в течение длительных периодов времени, в неблагоприятных условиях. [ 1 ] Споры являются частью жизненного цикла многих растений , водорослей , грибов и простейших . [ 2 ] Считалось, что они появились еще в середине позднего ордовика как адаптация ранних наземных растений. [ 3 ]
Споры бактерий не участвуют в половом цикле, а представляют собой устойчивые структуры, используемые для выживания в неблагоприятных условиях. [ 4 ] Споры миксозоев выделяют амебоидные инфекционные микробы («амебулы») в своих хозяев для паразитарной инфекции, но также размножаются внутри хозяев посредством спаривания двух ядер внутри плазмодия, который развивается из амебулы. [ 5 ]
У растений споры обычно гаплоидные , одноклеточные и образуются мейоза в спорангии диплоидного путем спорофита . В некоторых редких случаях диплоидные споры также образуются в некоторых водорослях или грибах. [ 6 ] При благоприятных условиях спора может развиться в новый организм путем митотического деления, образуя многоклеточный гаметофит , который в конечном итоге дает гаметы. Две гаметы сливаются, образуя зиготу , из которой развивается новый спорофит. Этот цикл известен как чередование поколений .
Споры семенных растений образуются внутри организма, а мегаспоры (образующиеся внутри семязачатков) и микроспоры участвуют в формировании более сложных структур, образующих дисперсные единицы - семена и пыльцевые зерна .
Определение
[ редактировать ]Термин «спора» происходит от древнегреческого слова σπορά spora , означающего « семя , посев», родственного σπόρος sporos , «посев», и σπείρειν speirein , «сеять».
В просторечии разница между «спорой» и « гаметой » заключается в том, что спора прорастает и развивается в спорлинг , в то время как гамете необходимо объединиться с другой гаметой, чтобы сформировать зиготу, прежде чем развиваться дальше.
Основное различие между спорами и семенами как единицами расселения состоит в том, что споры одноклеточные, первая клетка гаметофита, тогда как семена содержат внутри себя развивающийся зародыш (многоклеточный спорофит следующего поколения), образующийся в результате слияния мужской гаметы пыльцевая трубка с женской гаметой, образованной мегагаметофитом внутри семязачатка. Споры прорастают, давая гаплоидные гаметофиты, а семена прорастают, давая диплоидные спорофиты.
Классификация спорообразующих организмов
[ редактировать ]
Растения
[ редактировать ]Споры сосудистых растений всегда гаплоидны . Сосудистые растения бывают гомоспоровыми (или изоспоровыми) или разноспоровыми . Гомоспоровые растения производят споры одинакового размера и типа.
Разноспоровые растения, такие как семенные растения , колосовидные мхи , иголки и папоротники отряда Salviniales, производят споры двух разных размеров: более крупная спора (мегаспора), фактически функционирующая как «женская» спора, и меньшая (микроспора), функционирующая как «женская» спора. мужской". Такие растения обычно дают начало двум видам спор из отдельных спорангиев: либо мегаспорангия , производящего мегаспоры, либо микроспорангия , производящего микроспоры. У цветковых растений эти спорангии встречаются соответственно в плодолистике и пыльниках.
Грибы
[ редактировать ]Грибы обычно производят споры во время полового и бесполого размножения. Споры обычно гаплоидны и превращаются в зрелые гаплоидные особи путем митотического деления клеток ( урединиоспоры и телиоспоры ржавчины являются дикариотическими). Дикариотические клетки возникают в результате слияния двух гаплоидных гаметных клеток. Среди спорогенных дикариотических клеток происходит кариогамия (слияние двух гаплоидных ядер) с образованием диплоидной клетки. Диплоидные клетки подвергаются мейозу с образованием гаплоидных спор. [ нужна ссылка ]
Классификация спор
[ редактировать ]Споры можно классифицировать по нескольким признакам, например, по их спорообразующей структуре, функции, происхождению в течение жизненного цикла и подвижности.
Ниже приведена таблица, в которой перечислены способы классификации, названия, идентификационные характеристики, примеры и изображения различных видов спор.
| Режим классификации | Имя | Идентификация характеристики | Пример споросодержащего организма | Изображение | |
|---|---|---|---|---|---|

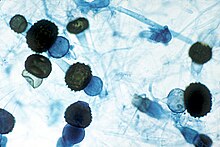
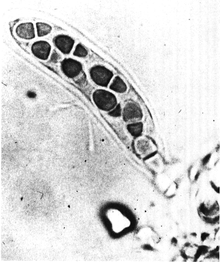


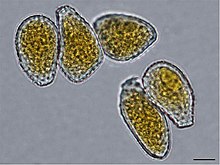
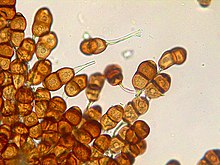












Внешняя анатомия
[ редактировать ]


При большом увеличении споры часто имеют сложные узоры или орнаменты на внешней поверхности. Для описания особенностей таких паттернов была разработана специализированная терминология. Некоторые отметки представляют собой отверстия — места, через которые можно проникнуть в прочную внешнюю оболочку споры при прорастании. Споры можно разделить на категории в зависимости от положения и количества этих отметин и отверстий. Споры Alete не имеют линий. В монолетных спорах на споре имеется одна узкая линия (лаэсура). [ 8 ] Указывает на предварительный контакт двух спор, которые в конечном итоге разделились. [ 3 ] В трилетных спорах каждая спора имеет три узкие линии, исходящие от центрального полюса. [ 8 ] Это показывает, что четыре споры имели общее происхождение и первоначально контактировали друг с другом, образуя тетраэдр. [ 3 ] Более широкое отверстие в форме желобка можно назвать кольпусом . [ 8 ] Число кольпи выделяет основные группы растений. Эвдикоты имеют трехбороздные споры (т.е. споры с тремя кольпи). [ 9 ]
Spore tetrads and trilete spores
[ редактировать ]Тетрады спор, заключенные в оболочку, считаются самым ранним свидетельством растительной жизни на суше. [ 10 ] датируемый средним ордовиком (ранний Лланвирн, ~ 470 миллионов лет назад ), периодом, из которого еще не были обнаружены макрофоссилии. [ 11 ] Отдельные трилетные споры, напоминающие споры современных криптогамных растений, впервые появились в летописи окаменелостей в конце ордовика. [ 12 ]
Рассредоточение
[ редактировать ]У грибов как бесполые, так и половые споры или спорангиоспоры многих видов грибов активно распространяются путем насильственного выброса из их репродуктивных структур. Этот выброс обеспечивает выход спор из репродуктивных структур, а также перемещение по воздуху на большие расстояния. Таким образом, многие грибы обладают специализированными механическими и физиологическими механизмами, а также структурами поверхности спор, такими как гидрофобины , для выброса спор. К этим механизмам относятся, например, насильственное выделение аскоспор, обусловленное структурой аска, и накопление осмолитов в жидкости аска, что приводит к взрывному выбросу аскоспор в воздух. [ 13 ]
Принудительный выброс одиночных спор, называемых баллистоспорами , включает образование небольшой капли воды ( капля Буллера ), которая при контакте со спорой приводит к ее выбросу снаряда с начальным ускорением более 10 000 g . [ 14 ] Другие грибы полагаются на альтернативные механизмы высвобождения спор, такие как внешние механические силы, примером которых являются дождевики . Привлечение насекомых, таких как мухи, к плодовым структурам благодаря их яркому цвету и гнилостному запаху для распространения грибковых спор - еще одна стратегия, наиболее часто используемая вонючками .
Было показано, что у мха обыкновенного гладкошапчатого ( Atrichum undulatum ) вибрация спорофита является важным механизмом высвобождения спор. [ 15 ]
В случае сосудистых растений, выделяющих споры , таких как папоротники, распространение ветром очень легких спор обеспечивает большую способность к распространению. Кроме того, споры менее подвержены хищничеству животных, чем семена, поскольку они почти не содержат запасов пищи; однако они более подвержены грибковому и бактериальному нападению. Их главное преимущество состоит в том, что из всех форм потомства спорам требуется наименьшее количество энергии и материалов для производства.
У колючего мха Selaginella lepidophylla распространение частично достигается за счет необычного типа диаспоры - перекати-поле . [ 16 ]
Источник
[ редактировать ]Споры были обнаружены в микроокаменелостях, относящихся к середине-концу ордовика . [ 17 ] Две предполагаемые первоначальные функции спор связаны с тем, появились ли они до или после наземных растений. Тщательно изученная гипотеза заключается в том, что споры были адаптацией ранних видов наземных растений, таких как эмбриофиты , которые позволяли растениям легко расселяться, адаптируясь к своей неводной среде. [ 17 ] [ 18 ] Это особенно подтверждается наблюдением толстой стенки спор в криптоспорах . Эти споровые стенки защитили бы потенциальное потомство от новых погодных элементов. [ 17 ] заключается в том, что споры были ранними предшественниками наземных растений и образовались во время ошибок в мейозе водорослей Вторая, более поздняя гипотеза , , предполагаемых ранних предков наземных растений. [ 19 ]
Независимо от того, возникли ли споры до или после наземных растений, их вклад в такие области, как палеонтология и филогенетика растений , был полезен. [ 19 ] Споры, обнаруженные в микроокаменелостях, также известные как криптоспоры, хорошо сохраняются благодаря фиксированному материалу, в котором они находятся, а также тому, насколько обильными и широко распространенными они были в соответствующие периоды времени. Эти микроископаемые особенно полезны при изучении ранних периодов существования Земли, поскольку макрофоссилии, такие как растения, не распространены и не хорошо сохранились. [ 17 ] И криптоспоры, и современные споры имеют разнообразную морфологию, что указывает на возможные условия окружающей среды более ранних периодов существования Земли и эволюционные взаимоотношения видов растений. [ 17 ] [ 19 ] [ 18 ]
Галерея
[ редактировать ]-
 Споры мха Bartramia ithyphylla . (микроскопическое изображение, 400x)
Споры мха Bartramia ithyphylla . (микроскопическое изображение, 400x) -
 Расслоившиеся спорангии папоротника. (микроскопическое исследование, споры не видны)
Расслоившиеся спорангии папоротника. (микроскопическое исследование, споры не видны) -
 Споры и элятеры из хвоща. ( Хвощ , вид под микроскопом)
Споры и элятеры из хвоща. ( Хвощ , вид под микроскопом) -
 Споры ископаемых растений ( Scylaspora ) из силурийских отложений Швеции.
Споры ископаемых растений ( Scylaspora ) из силурийских отложений Швеции. -
 Плодовая плесень со спорами и заметным клеточным ростом. (2000x)
Плодовая плесень со спорами и заметным клеточным ростом. (2000x) -
 Скопления спор, образующиеся внутри спорангиев слизевика Reticularia olivacea , из сосновых лесов Восточной Украины .
Скопления спор, образующиеся внутри спорангиев слизевика Reticularia olivacea , из сосновых лесов Восточной Украины . -
 Внутренняя поверхность перидия слизевика Tubifera dudkae со спорами.
Внутренняя поверхность перидия слизевика Tubifera dudkae со спорами.







См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Сетлоу, Питер; Джонсон, Эрик А. (30 апреля 2014 г.), Дойл, Майкл П.; Бьюкенен, Роберт Л. (ред.), «Споры и их значение» , Food Microbiology , Вашингтон, округ Колумбия, США: ASM Press, стр. 45–79, doi : 10.1128/9781555818463.ch3 , ISBN 978-1-68367-058-2 , получено 13 декабря 2023 г.
- ^ «Веб-проект «Древо жизни» . Архивировано из оригинала 5 февраля 2018 года . Проверено 5 февраля 2018 г.
- ^ Перейти обратно: а б с Веллман, Швейцария; Грей, Дж. (29 июня 2000 г.). «Летопись микрофоссилий ранних наземных растений» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 355 (1398): 717–731, обсуждение 731–732. дои : 10.1098/rstb.2000.0612 . ISSN 0962-8436 . ПМЦ 1692785 . ПМИД 10905606 .
- ^ Абель-Сантос, Эрнесто (2012). Бактериальные споры: современные исследования и применение . Норфолк: академическое издательство Caister. ISBN 978-1-908230-00-3 .
- ^ Иван Фиала (10 июля 2008 г.). «Миксозоа» . Веб-проект «Древо жизни» . Архивировано из оригинала 16 февраля 2015 года . Проверено 14 января 2014 г.
Миксоспоры состоят из нескольких клеток, которые трансформируются в створки раковины, нематоцистоподобные полярные капсулы со спиральными выдавливающимися полярными нитями и амебоидные инфекционные зародыши.
- ^ «Диплоидные споры — обзор | Темы ScienceDirect» . www.sciencedirect.com . Архивировано из оригинала 13 декабря 2023 года . Проверено 13 декабря 2023 г.
- ^ «Биология микроспоридий» . 26 июня 2008 г. Архивировано из оригинала 26 июня 2008 г. Проверено 24 марта 2024 г.
- ^ Перейти обратно: а б с Пунт, В.; Хоэн, ПП; Блэкмор, С.; Нильссон С. и Ле Томас А. (2007). «Словарь терминологии пыльцы и спор». Обзор палеоботаники и палинологии . 143 (1): 1–81. Бибкод : 2007RPaPa.143....1P . дои : 10.1016/j.revpalbo.2006.06.008 .
- ^ Джадд, Уолтер С. и Олмстед, Ричард Г. (2004). «Обзор филогенетических взаимоотношений трехбороздных (эвдикотовых)» . Американский журнал ботаники . 91 (10): 1627–44. дои : 10.3732/ajb.91.10.1627 . ПМИД 21652313 .
- ^ Грей, Дж.; Чалонер, В.Г.; Уэстолл, Т.С. (1985). «Летопись микрофоссилий ранних наземных растений: достижения в понимании ранней террестриализации, 1970–1984» . Философские труды Королевского общества Б. 309 (1138): 167–195. Бибкод : 1985РСТБ.309..167Г . дои : 10.1098/rstb.1985.0077 . JSTOR 2396358 .
- ^ Веллман Ч., Грей Дж. (2000). «Летопись микрофоссилий ранних наземных растений» . Философские труды Королевского общества Б. 355 (1398): 717–732. дои : 10.1098/rstb.2000.0612 . ПМЦ 1692785 . ПМИД 10905606 .
- ^ Стиманс, П.; Херисс, Алабама; Мелвин, Дж.; Миллер, Массачусетс; Пэрис, Ф.; Верньерс, Дж.; Веллман, Швейцария (2009). «Происхождение и распространение древнейших сосудистых наземных растений» (PDF) . Наука . 324 (5925): 353. Бибкод : 2009Sci...324..353S . дои : 10.1126/science.1169659 . hdl : 1854/LU-697223 . ISSN 0036-8075 . ПМИД 19372423 . S2CID 206518080 . Архивировано (PDF) из оригинала 22 сентября 2017 года . Проверено 1 ноября 2017 г.
- ^ Трейл Ф. (2007). «Грибные пушки: взрывной выброс спор у Ascomycota» . Письма FEMS по микробиологии . 276 (1): 12–8. дои : 10.1111/j.1574-6968.2007.00900.x . ПМИД 17784861 .
- ^ Прингл А., Патек С.Н., Фишер М., Штольце Дж., Money NP (2005). «Захваченный запуск баллистоспоры». Микология . 97 (4): 866–71. дои : 10.3852/микология.97.4.866 . ПМИД 16457355 .
- ^ Йоханссон, Лённелл, Сундберг и Хайландер (2014) Пороги высвобождения спор мха: важность турбулентности и длины спорофитов. Журнал экологии, н/д/д.
- ^ «Ложная роза Иерихона – Selaginella lepidophylla» Ложная роза Иерихона – Selaginella lepidophylla» . Путеводитель по растениям и цветам . Февраль 2009 г. Архивировано из оригинала 15 июля 2011 г. Проверено 1 февраля 2010 г.
- ^ Перейти обратно: а б с д и Веллман, Швейцария; Грей, Дж. (29 июня 2000 г.). «Летопись микрофоссилий ранних наземных растений» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 355 (1398): 717–731, обсуждение 731–732. дои : 10.1098/rstb.2000.0612 . ISSN 0962-8436 . ПМЦ 1692785 . ПМИД 10905606 .
- ^ Перейти обратно: а б Норем, WL (1958). «Определители классификации ископаемых спор и пыльцы» . Журнал палеонтологии . 32 (4): 666–676. ISSN 0022-3360 . JSTOR 1300785 .
- ^ Перейти обратно: а б с Стротер, Пол К.; Фостер, Клинтон (13 августа 2021 г.). «Ископаемая летопись происхождения наземных растений из харофитовых водорослей» . Наука . 373 (6556): 792–796. Бибкод : 2021Sci...373..792S . дои : 10.1126/science.abj2927 . ISSN 0036-8075 . ПМИД 34385396 .