рецептор ЛПНП
| ЛДЛР | ||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | LDLR , FH, FHC, LDLCQ2, рецептор липопротеинов низкой плотности, FHCL1 | |||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Опустить : 606945 ; МГИ : 96765 ; Гомологен : 55469 ; GeneCards : LDLR ; ОМА : LDLR – ортологи | |||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||






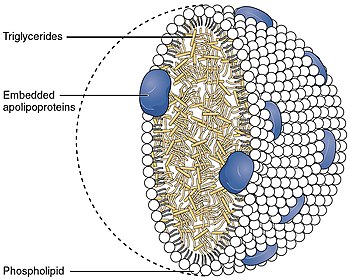
Рецептор липопротеинов низкой плотности ( ЛПНП-Р ) представляет собой мозаичный белок из 21 аминокислоты из 839 аминокислот (после удаления сигнального пептида ). [ 5 ] который опосредует эндоцитоз богатых холестерином липопротеинов низкой плотности (ЛПНП). Это рецептор клеточной поверхности, который распознает аполипопротеин B100 (ApoB100), который встроен во внешний фосфолипидный слой липопротеинов очень низкой плотности (ЛПОНП), их остатков, то есть липопротеинов промежуточной плотности (ЛПНП), и частиц ЛПНП. Рецептор также распознает аполипопротеин Е (АроЕ), который содержится в остатках хиломикронов и IDL. У человека белок рецептора ЛПНП кодируется геном на LDLR хромосоме 19 . [ 6 ] [ 7 ] [ 8 ] Он принадлежит к семейству генов рецепторов липопротеинов низкой плотности . [ 9 ] Наиболее значительно он экспрессируется в бронхов эпителиальных клетках , а также в тканях надпочечников и коры головного мозга . [ 10 ]
Майкл С. Браун и Джозеф Л. Гольдштейн были удостоены Нобелевской премии по физиологии и медицине 1985 года за определение ЛПНП-Р. [ 11 ] и его связь с метаболизмом холестерина и семейной гиперхолестеринемией . [ 12 ] Нарушение ЛПНП-Р может привести к повышению уровня ЛПНП-холестерина, а также к увеличению риска связанных с ним заболеваний. Лица с деструктивными мутациями (определяемыми как нонсенс , сайт сплайсинга или индел-сдвиг рамки считывания ) в LDLR имеют средний уровень холестерина ЛПНП 279 мг/дл по сравнению со 135 мг/дл для лиц, не имеющих ни разрушительных, ни вредных мутаций. Деструктивные мутации встречались в 13 раз чаще у людей с ранним инфарктом миокарда или ишемической болезнью сердца, чем у людей без того или иного заболевания. [ 13 ]
Структура
[ редактировать ]Ген
[ редактировать ]Ген LDLR расположен на хромосоме 19 в участке 19p13.2 и разделен на 18 экзонов . [ 8 ] Экзон 1 содержит сигнальную последовательность, которая локализует рецептор в эндоплазматическом ретикулуме для транспорта на поверхность клетки. Помимо этого, экзоны 2–6 кодируют область связывания лиганда; 7-14 кодируют домен эпидермального фактора роста (EGF); 15 кодирует богатую олигосахаридами область; 16 (и некоторые из 17) кодируют область, охватывающую мембрану; и 18 (вместе с остальными 17) кодируют цитозольный домен.
Этот ген производит 6 изоформ посредством альтернативного сплайсинга. [ 14 ]
Белок
[ редактировать ]Этот белок принадлежит к семейству LDLR и состоит из ряда функционально различных доменов , включая 3 EGF-подобных домена, 7 доменов LDL-R класса A и 6 повторов LDL-R класса B. [ 14 ]
N -концевой домен рецептора ЛПНП, отвечающий за связывание лиганда, состоит из семи повторов последовательности (идентичных примерно на 50%). Каждый повтор, называемый повтором класса А или ЛПНП-А , содержит примерно 40 аминокислот, включая 6 цистеина остатков , которые образуют дисульфидные связи внутри повтора. Кроме того, каждый повтор имеет высококонсервативные кислотные остатки, которые он использует для координации одного иона кальция в октаэдрической решетке. И дисульфидные связи, и координация кальция необходимы для структурной целостности домена во время повторных поездок рецептора в высококислотную внутреннюю часть эндосомы . Точный механизм взаимодействия между повторами класса А и лигандом (ЛПНП) неизвестен, но считается, что повторы действуют как «захватчики», удерживающие ЛПНП. Для связывания ApoB требуются повторы 2–7, тогда как для связывания ApoE требуется только повтор 5 (считающийся предковым повтором).
Рядом с лигандсвязывающим доменом находится домен гомологии предшественника EGF (домен EGFP). Это показывает примерно 30% гомологию с геном-предшественником EGF. Есть три повтора «фактора роста»; A, B и C. A и B тесно связаны, тогда как C разделен областью повтора YWTD , которая принимает конформацию бета-пропеллера ( домен LDL-R класса B ). Считается, что эта область отвечает за рН-зависимый конформационный сдвиг, который вызывает высвобождение связанных ЛПНП в эндосоме .
Третий домен белка богат О-связанными олигосахаридами , но, по-видимому, малофункционален. Эксперименты по нокауту подтвердили, что без этого домена не происходит значительной потери активности. Было высказано предположение, что этот домен мог изначально действовать как спейсер, выталкивая рецептор за пределы внеклеточного матрикса .
Единственный трансмембранный домен из 22 (в основном) неполярных остатков пересекает плазматическую мембрану в виде одной альфа-спирали .
Цитозольный покрытых С-концевой домен содержит ~50 аминокислот, включая сигнальную последовательность, важную для локализации рецепторов в клатрином ямках и для запуска рецептор-опосредованного эндоцитоза после связывания. Части цитозольной последовательности были обнаружены у других липопротеиновых рецепторов, а также у более отдаленных родственников рецепторов. [ 15 ] [ 16 ] [ 17 ]
Мутации
[ редактировать ]Известно, что мутации потери функции в гене, кодирующем рецептор ЛПНП, вызывают семейную гиперхолестеринемию.
Существует 5 широких классов мутаций рецептора ЛПНП:
- Мутации класса 1 влияют на синтез рецептора в эндоплазматическом ретикулуме (ЭР).
- Мутации класса 2 препятствуют правильному транспорту в тело Гольджи, необходимому для модификации рецептора.
- например, усечение рецепторного белка по номеру остатка 660 приводит к отсутствию доменов 3, 4 и 5 домена-предшественника EGF. Это предотвращает перемещение рецептора из ЭР в Гольджи и приводит к деградации рецепторного белка.
- Мутации класса 3 прекращают связывание ЛПНП с рецептором.
- например, удаляется повтор 6 лигандсвязывающего домена (N-конец, внеклеточная жидкость).
- Мутации класса 4 ингибируют интернализацию комплекса рецептор-лиганд.
- например, мутант «JD» возникает в результате единственной точечной мутации в домене NPVY (С-концевой, цитозольный; остаток C преобразован в Y, номер остатка 807). Этот домен рекрутирует клатрин и другие белки, ответственные за эндоцитоз ЛПНП, поэтому эта мутация ингибирует интернализацию ЛПНП.
- Мутации класса 5 приводят к появлению рецепторов, которые не могут должным образом перерабатываться. Это приводит к относительно мягкому фенотипу , поскольку рецепторы все еще присутствуют на поверхности клетки (но все они должны быть синтезированы заново). [ 18 ]
Мутации усиления функции снижают уровни ЛПНП и являются целью исследований по разработке генной терапии для лечения рефрактерной гиперхолестеринемии. [ 19 ]
Функция
[ редактировать ]Рецептор ЛПНП опосредует эндоцитоз ЛПНП, богатых холестерином, и, таким образом, поддерживает уровень ЛПНП в плазме. [ 20 ] Это происходит во всех ядросодержащих клетках, но главным образом в печени , которая удаляет из кровообращения около 70% ЛПНП. Рецепторы ЛПНП группируются в ямках, покрытых клатрином , и покрытые оболочки ямки отщипываются от поверхности, образуя покрытые эндоцитарные пузырьки, которые переносят ЛПНП в клетку. [ 21 ] После интернализации рецепторы диссоциируют от своих лигандов, когда они подвергаются воздействию более низкого pH в эндосомах . После диссоциации рецептор сворачивается, приобретая закрытую конформацию, и возвращается на поверхность клетки. [ 22 ] Быстрая переработка рецепторов ЛПНП обеспечивает эффективный механизм доставки холестерина в клетки. [ 23 ] [ 24 ] Также сообщалось, что в результате ассоциации с липопротеинами в крови такие вирусы, как вирус гепатита С , Flaviviridae вирусы и вирус бычьей диареи, могут проникать в клетки опосредованно через LDLR-опосредованный эндоцитоз. [ 25 ] LDLR был идентифицирован как основной путь проникновения вируса везикулярного стоматита у мышей и людей. [ 26 ] Кроме того, модуляция LDLR связана с ранней лимфатической дисфункцией, связанной с атеросклерозом. [ 27 ] Синтез рецепторов в клетке регулируется уровнем свободного внутриклеточного холестерина; если его в избытке для нужд клетки, то транскрипция гена рецептора будет ингибироваться. [ 28 ] Рецепторы ЛПНП транслируются рибосомами эндоплазматического ретикулума и модифицируются аппаратом Гольджи перед тем, как попасть в везикулах на поверхность клетки.
Клиническое значение
[ редактировать ]У человека ЛПНП непосредственно участвуют в развитии атеросклероза , процесса, ответственного за большинство сердечно-сосудистых заболеваний , вследствие накопления холестерина ЛПНП в крови. [ нужна ссылка ] . Гипертиреоз может быть связан со снижением уровня холестерина за счет активации рецептора ЛПНП, а гипотиреоз – наоборот. Огромное количество исследований описало значение рецепторов ЛПНП в патофизиологии атеросклероза, метаболического синдрома и стеатогепатита. [ 29 ] [ 30 ] Ранее было показано, что редкие мутации в генах ЛПНП способствуют риску инфаркта миокарда в отдельных семьях, тогда как распространенные варианты более чем в 45 локусах были связаны с риском инфаркта миокарда в популяции. По сравнению с неносителями, носители мутации LDLR имели более высокий уровень холестерина ЛПНП в плазме, тогда как носители мутации APOA5 имели более высокие уровни триглицеридов в плазме. [ 31 ] Недавние данные связали риск ИМ с мутациями кодирующей последовательности двух генов, функционально связанных с APOA5, а именно липопротеинлипазы и аполипопротеина C-III. [ 32 ] [ 33 ] В совокупности эти наблюдения позволяют предположить, что наряду с холестерином ЛПНП нарушение метаболизма липопротеинов, богатых триглицеридами, способствует риску ИМ. В целом, LDLR имеет высокую клиническую значимость в отношении липидов крови. [ 34 ] [ 35 ]
Клинический маркер
[ редактировать ]Многолокусное исследование оценки генетического риска, основанное на комбинации 27 локусов, включая ген LDLR, выявило лиц с повышенным риском как возникновения, так и рецидивов ишемической болезни сердца, а также увеличило клиническую пользу от терапии статинами . Исследование было основано на групповом исследовании на уровне сообщества (исследование диеты и рака в Мальмё) и четырех дополнительных рандомизированных контролируемых исследованиях когорт первичной профилактики (JUPITER и ASCOT) и когорт вторичной профилактики (CARE и PROVE IT-TIMI 22). [ 36 ]
Интерактивная карта маршрутов
[ редактировать ]Нажмите на гены, белки и метаболиты ниже, чтобы перейти к соответствующим статьям. [ § 1 ]

- ^ Интерактивную карту маршрутов можно редактировать на WikiPathways: «Statin_Pathway_WP430» .
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с GRCh38: Версия Ensembl 89: ENSG00000130164 – Ensembl , май 2017 г.
- ^ Перейти обратно: а б с GRCm38: выпуск Ensembl 89: ENSMUSG00000032193 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ Зюдхоф Т.К., Гольдштейн Дж.Л., Браун М.С., Рассел Д.В. (май 1985 г.). «Ген рецептора ЛПНП: мозаика экзонов, общих для разных белков» . Наука . 228 (4701): 815–22. Бибкод : 1985Sci...228..815S . дои : 10.1126/science.2988123 . ПМЦ 4450672 . ПМИД 2988123 .
- ^ Франке У., Браун М.С., Гольдштейн Дж.Л. (май 1984 г.). «Отнесение человеческого гена рецептора липопротеинов низкой плотности к хромосоме 19: синтения рецептора, лиганда и генетическое заболевание» . Труды Национальной академии наук Соединенных Штатов Америки . 81 (9): 2826–30. Бибкод : 1984PNAS...81.2826F . дои : 10.1073/pnas.81.9.2826 . ПМК 345163 . ПМИД 6326146 .
- ^ Линдгрен В., Ласки К.Л., Рассел Д.В., Франк У. (декабрь 1985 г.). «Гены человека, участвующие в метаболизме холестерина: хромосомное картирование локусов рецептора липопротеина низкой плотности и 3-гидрокси-3-метилглутарил-коэнзима А-редуктазы с помощью зондов кДНК» . Труды Национальной академии наук Соединенных Штатов Америки . 82 (24): 8567–71. Бибкод : 1985PNAS...82.8567L . дои : 10.1073/pnas.82.24.8567 . ПМК 390958 . ПМИД 3866240 .
- ^ Перейти обратно: а б «Рецептор липопротеина низкой плотности LDLR [Homo sapiens (человек)] - Ген - NCBI» . www.ncbi.nlm.nih.gov . Проверено 10 октября 2016 г.
- ^ Никьяер А., Уиллноу Т.Е. (июнь 2002 г.). «Семейство генов рецепторов липопротеинов низкой плотности: клеточный швейцарский армейский нож?». Тенденции в клеточной биологии . 12 (6): 273–80. дои : 10.1016/S0962-8924(02)02282-1 . ПМИД 12074887 .
- ^ «BioGPS – ваша генная портальная система» . biogps.org . Проверено 10 октября 2016 г.
- ^ «Нобелевская премия по физиологии и медицине 1985 г.» (пресс-релиз). Шведская королевская академия наук. 1985 год . Проверено 1 июля 2010 г.
- ^ Браун М.С., Гольдштейн Дж.Л. (ноябрь 1984 г.). «Как рецепторы ЛПНП влияют на холестерин и атеросклероз». Научный американец . 251 (5): 58–66. Бибкод : 1984SciAm.251c..52K . doi : 10.1038/scientificamerican0984-52 . ПМИД 6390676 .
- ^ До Р., Ститциел Н.О., Вон Х.Х., Йоргенсен А.Б., Дуга С., Анжелика Мерлини П. и др. (февраль 2015 г.). «Секвенирование экзома выявляет редкие аллели LDLR и APOA5, обуславливающие риск инфаркта миокарда» . Природа . 518 (7537): 102–6. Бибкод : 2015Natur.518..102. . дои : 10.1038/nature13917 . ПМК 4319990 . ПМИД 25487149 .
- ^ Перейти обратно: а б «LDLR - предшественник рецептора липопротеинов низкой плотности - Homo sapiens (человек) - ген и белок LDLR» . www.uniprot.org . Проверено 10 октября 2016 г.
- ^ Ямамото Т., Дэвис К.Г., Браун М.С., Шнайдер В.Дж., Кейси М.Л., Гольдштейн Дж.Л. и др. (ноябрь 1984 г.). «Человеческий рецептор ЛПНП: богатый цистеином белок с множеством последовательностей Alu в мРНК». Клетка . 39 (1): 27–38. дои : 10.1016/0092-8674(84)90188-0 . ПМИД 6091915 . S2CID 25822170 .
- ^ Браун М.С., Герц Дж., Гольдштейн Дж.Л. (август 1997 г.). «Структура ЛПНП-рецептора. Кальциевые клетки, кислотные ванны и рециклирующие рецепторы» . Природа . 388 (6643): 629–30. Бибкод : 1997Natur.388..629B . дои : 10.1038/41672 . ПМИД 9262394 . S2CID 33590160 .
- ^ Гент Дж., Браакман I (октябрь 2004 г.). «Структура и сворачивание рецептора липопротеинов низкой плотности». Клеточные и молекулярные науки о жизни . 61 (19–20): 2461–70. дои : 10.1007/s00018-004-4090-3 . ПМИД 15526154 . S2CID 21235282 .
- ^ «Рецептор липопротеинов низкой плотности» . LOVD v.1.1.0 — Лейденская открытая база данных вариаций . Архивировано из оригинала 28 января 2016 г. Проверено 17 октября 2013 г.
- ^ Шривастава Р.А. (декабрь 2023 г.). «Новые возможности в ведении и лечении рефрактерной гиперхолестеринемии с использованием CRISPR-опосредованного редактирования генома/оснований in vivo». Питание, обмен веществ и сердечно-сосудистые заболевания . 33 (12): 2317–2325. дои : 10.1016/j.numecd.2023.08.010 . ПМИД 37805309 .
- ^ Лерен Т.П. (ноябрь 2014 г.). «Сортировка рецептора ЛПНП со связанным PCSK9 для внутриклеточной деградации». Атеросклероз . 237 (1): 76–81. doi : 10.1016/j.atherosclerosis.2014.08.038 . ПМИД 25222343 .
- ^ Гольдштейн Дж.Л., Браун М.С. (апрель 2009 г.). «Рецептор ЛПНП» . Атеросклероз, тромбоз и сосудистая биология . 29 (4): 431–8. дои : 10.1161/ATVBAHA.108.179564 . ПМК 2740366 . ПМИД 19299327 .
- ^ Руденко Г., Генри Л., Хендерсон К., Ищенко К., Браун М.С., Гольдштейн Дж.Л. и др. (декабрь 2002 г.). «Структура внеклеточного домена рецептора ЛПНП при эндосомальном pH» . Наука . 298 (5602): 2353–8. Бибкод : 2002Sci...298.2353R . дои : 10.1126/science.1078124 . ПМИД 12459547 . S2CID 17712211 .
- ^ Басу С.К., Гольдштейн Дж.Л., Андерсон Р.Г., Браун М.С. (май 1981 г.). «Моненсин прерывает рециркуляцию рецепторов липопротеинов низкой плотности в фибробластах человека». Клетка . 24 (2): 493–502. дои : 10.1016/0092-8674(81)90340-8 . ПМИД 6263497 . S2CID 29553611 .
- ^ Браун М.С., Андерсон Р.Г., Гольдштейн Дж.Л. (март 1983 г.). «Рециркуляционные рецепторы: маршрут туда и обратно мигрирующих мембранных белков». Клетка . 32 (3): 663–7. дои : 10.1016/0092-8674(83)90052-1 . ПМИД 6299572 . S2CID 34919831 .
- ^ Агнелло В., Абель Г., Эльфахал М., Найт ГБ, Чжан QX (октябрь 1999 г.). «Вирус гепатита С и другие вирусы флавивирусов проникают в клетки через рецептор липопротеинов низкой плотности» . Труды Национальной академии наук Соединенных Штатов Америки . 96 (22): 12766–71. Бибкод : 1999PNAS...9612766A . дои : 10.1073/pnas.96.22.12766 . ПМК 23090 . ПМИД 10535997 .
- ^ Финкельштейн Д., Верман А., Новик Д., Барак С., Рубинштейн М. (апрель 2013 г.). «Рецептор ЛПНП и члены его семейства служат клеточными рецепторами для вируса везикулярного стоматита» . Труды Национальной академии наук Соединенных Штатов Америки . 110 (18): 7306–11. Бибкод : 2013PNAS..110.7306F . дои : 10.1073/pnas.1214441110 . ПМЦ 3645523 . ПМИД 23589850 .
- ^ Миласан А., Даллэр Ф., Майер Дж., Мартель К. (01.01.2016). «Влияние модуляции рецепторов ЛПНП на лимфатическую функцию» . Научные отчеты . 6 : 27862. Бибкод : 2016NatSR...627862M . дои : 10.1038/srep27862 . ПМЦ 4899717 . ПМИД 27279328 .
- ^ Смит-младший, Осборн Т.Ф., Гольдштейн Дж.Л., Браун М.С. (февраль 1990 г.). «Идентификация нуклеотидов, ответственных за усиление активности регуляторного элемента стерола в гене рецептора липопротеинов низкой плотности» . Журнал биологической химии . 265 (4): 2306–10. дои : 10.1016/S0021-9258(19)39976-4 . ПМИД 2298751 . S2CID 26062629 .
- ^ Се Дж., Косеки М., Молуски М.М., Якусиджи Э., Ичи И., Вестертерп М. и др. (июль 2016 г.). «Дефицит TTC39B стабилизирует LXR, уменьшая как атеросклероз, так и стеатогепатит» . Природа . 535 (7611): 303–7. Бибкод : 2016Natur.535..303H . дои : 10.1038/nature18628 . ПМК 4947007 . ПМИД 27383786 .
- ^ Уолтер К., Мин Дж.Л., Хуан Дж., Крукс Л., Мемари Ю., Маккарти С. и др. (октябрь 2015 г.). «Проект UK10K выявляет редкие варианты здоровья и болезней» . Природа . 526 (7571): 82–90. Бибкод : 2015Natur.526...82T . дои : 10.1038/nature14962 . ПМЦ 4773891 . ПМИД 26367797 .
- ^ Роуз-Хеллекант Т.А., Шредер М.Д., Брокман Дж.Л., Жданкин О., Болстад Р., Чен К.С. и др. (август 2007 г.). «Эстрогеновый рецептор-положительный онкогенез молочной железы у трансгенных мышей TGF-альфа прогрессирует с потерей рецептора прогестерона» . Онкоген . 26 (36): 5238–46. дои : 10.1038/sj.onc.1210340 . ПМЦ 2587149 . ПМИД 17334393 .
- ^ Кросби Дж., Пелосо Г.М., Ауэр П.Л., Кросслин Д.Р., Ститциел Н.О., Ланге Л.А. и др. (июль 2014 г.). «Мутации потери функции в APOC3, триглицеридах и ишемической болезни сердца» . Медицинский журнал Новой Англии . 371 (1): 22–31. дои : 10.1056/NEJMoa1307095 . ПМК 4180269 . ПМИД 24941081 .
- ^ Йоргенсен А.Б., Фрикке-Шмидт Р., Нордестгаард Б.Г., Тыбьёрг-Хансен А. (июль 2014 г.). «Мутации потери функции в APOC3 и риск ишемической болезни сосудов» . Медицинский журнал Новой Англии . 371 (1): 32–41. дои : 10.1056/NEJMoa1308027 . ПМИД 24941082 . S2CID 26995834 .
- ^ Шульдинер А.Р., Поллин Т.И. (август 2010 г.). «Геномика: вариации липидов крови» . Природа . 466 (7307): 703–4. Бибкод : 2010Natur.466..703S . дои : 10.1038/466703а . ПМИД 20686562 . S2CID 205057802 .
- ^ Теслович Т.М., Мусунуру К., Смит А.В., Эдмондсон А.С., Стилиану И.М., Косеки М. и др. (август 2010 г.). «Биологическая, клиническая и популяционная значимость 95 локусов липидов крови» . Природа . 466 (7307): 707–13. Бибкод : 2010Natur.466..707T . дои : 10.1038/nature09270 . ПМК 3039276 . ПМИД 20686565 .
- ^ Мега Дж.Л., Ститциел Н.О., Смит Дж.Г., Чесман Д.И., Колфилд М.Дж., Девлин Дж.Дж. и др. (июнь 2015 г.). «Генетический риск, события ишемической болезни сердца и клиническая польза терапии статинами: анализ исследований первичной и вторичной профилактики» . Ланцет . 385 (9984): 2264–71. дои : 10.1016/S0140-6736(14)61730-X . ПМЦ 4608367 . ПМИД 25748612 .
Дальнейшее чтение
[ редактировать ]- Браун М.С., Гольдштейн Дж.Л. (июль 1979 г.). «Рецепторно-опосредованный эндоцитоз: данные о рецепторной системе липопротеинов» . Труды Национальной академии наук Соединенных Штатов Америки . 76 (7): 3330–7. Бибкод : 1979PNAS...76.3330B . дои : 10.1073/pnas.76.7.3330 . ПМЦ 383819 . ПМИД 226968 .
- Хоббс Х.Х., Браун М.С., Гольдштейн Дж.Л. (1993). «Молекулярная генетика гена рецептора ЛПНП при семейной гиперхолестеринемии» . Человеческая мутация . 1 (6): 445–66. дои : 10.1002/humu.1380010602 . ПМИД 1301956 . S2CID 5756814 .
- Фогельман А.М., Ван Лентен Б.Дж., Уорден С., Хаберланд М.Э., Эдвардс П.А. (1989). «Макрофагальные липопротеиновые рецепторы» . Журнал клеточной науки. Добавка . 9 : 135–49. doi : 10.1242/jcs.1988.supplement_9.7 . ПМИД 2855802 .
- Барретт П.Х., Уоттс Г.Ф. (март 2002 г.). «Смена парадигмы рецепторов ЛПНП при семейной гиперхолестеринемии: новые данные недавних кинетических исследований метаболизма аполипопротеина B-100». Атеросклероз. Дополнения . 2 (3): 1–4. дои : 10.1016/S1567-5688(01)00012-5 . ПМИД 11923121 .
- Мэй П., Бок Х.Х., Герц Дж. (апрель 2003 г.). «Интеграция эндоцитоза и передачи сигнала липопротеиновыми рецепторами». СТКЭ науки . 2003 (176): ПЭ12. дои : 10.1126/stke.2003.176.pe12 . ПМИД 12671190 . S2CID 24468290 .
- Гент Дж., Браакман I (октябрь 2004 г.). «Структура и сворачивание рецептора липопротеинов низкой плотности». Клеточные и молекулярные науки о жизни . 61 (19–20): 2461–70. дои : 10.1007/s00018-004-4090-3 . ПМИД 15526154 . S2CID 21235282 .
Внешние ссылки
[ редактировать ]- Описание пути рецептора ЛПНП на веб-странице лаборатории Брауна-Гольдштейна
- Рецептор ЛПНП в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
галерея PDB |
|---|












Липиды : метаболизм липопротеиновых частиц . |
|---|