Агната
| Агната | |
|---|---|
 | |
| Один из сохранившихся представителей семейства бесчелюстных, Petromyzontidae. | |
 | |
| Реконструкция жизни вымершего рода Agnatan Pteraspis | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Подтип: | Позвоночные животные |
| Инфрафилум: | Агната Коуп , 1889 г. |
| Группы включены | |
| |
Агната ( / ˈ æ ɡ n ə θ ə , æ ɡ ˈ n eɪ θ ə / ; [3] от древнегреческого ἀ- ( a- ) «без» и γνάθος ( gnáthos ) «челюсти») — инфратип [4] бесчелюстных рыб типа ) , Chordata , подтипа Vertebrata , состоящего как из ныне живущих ( круглоротые так и из вымерших ( конодонты , анаспиды , остракодермы ) видов . Среди современных животных круглоротые являются родственными братьями всем позвоночным животным с челюстями , известными как челюстноротые . [5]
Молекулярные данные , как по рРНК [6] и из мтДНК [7] а также эмбриологические данные, [8] убедительно подтверждает гипотезу о том, что обе группы современных бесчелюстных, миксин и миног , более тесно связаны друг с другом, чем с челюстными рыбами , образуя кладу Cyclostomi. [9]
Самые древние ископаемые бесчелюстные появились в кембрии , и до сих пор сохранились две группы: миноги и миксины , насчитывающие в общей сложности около 120 видов . Миксы считаются представителями подтипа Vertebrata , поскольку они вторично потеряли позвонки; до того, как это событие было выведено из молекулярных [6] [7] [10] и развития [11] По данным, группа Craniata была создана Линнеем (и до сих пор иногда используется как строго морфологический дескриптор) для обозначения миксин и позвоночных.
Хотя некоторые учёные до сих пор считают ныне живущих бесчелюстных сходными лишь внешне и утверждают, что многие из этих сходств, вероятно, являются общими базальными характеристиками древних позвоночных, новые таксономические исследования ясно относят миксин (Myxini или Hyperotreti) к миногам (Hyperoartii). более тесно связаны друг с другом, чем с челюстными рыбами. [6] [7] [12]
Метаболизм
[ редактировать ]Бесчелюстные — экзотермические животные , то есть они не регулируют температуру собственного тела. В холодной воде обмен веществ у агнатанов медленный, поэтому им не приходится много есть. У них нет отчетливого желудка, а есть длинная кишка, более или менее однородная на всем протяжении. Миноги питаются другими рыбами и млекопитающими. Антикоагулянтные жидкости, предотвращающие свертывание крови, вводятся хозяину, в результате чего хозяин выделяет больше крови. Миксы — падальщики, питающиеся в основном мертвыми животными. Чтобы сломать животное, они используют ряд острых зубов. Тот факт, что зубы Agnathan не могут двигаться вверх и вниз, ограничивает их возможные типы питания.
Морфология
[ редактировать ]Помимо отсутствия челюстей , для современных бесчелюстных характерно отсутствие парных плавников ; наличие хорды как у личинок, так и у взрослых особей; и семь или более парных жаберных мешков. У миног есть светочувствительный шишковидный глаз (гомологичный шишковидной железе млекопитающих . ) У всех ныне живущих и большинства вымерших Agnatha нет желудка или каких-либо придатков . Оплодотворение и развитие являются внешними. В классе Агната нет родительской опеки. Agnatha — экзотермические или холодные, с хрящевым скелетом , сердцем 2 камеры.
Покрытие тела
[ редактировать ]У современных бесчелюстных тело покрыто кожей без дермальных и эпидермальных чешуек . На коже миксины имеется большое количество слизевых желез, слизь представляет собой их защитный механизм. Иногда слизь может закупоривать жабры вражеских рыб, вызывая их гибель. Напротив, многие вымершие бесчелюстные имели обширный экзоскелет, состоящий либо из массивного тяжелого кожного панциря , либо из небольших минерализованных чешуек.
Придатки
[ редактировать ]Почти все бесчелюстные, включая всех современных бесчелюстных , не имеют парных придатков, хотя у большинства есть спинной или хвостовой плавник . Некоторые ископаемые бесчелюстные, такие как остеостраканы и питуриаспиды , действительно имели парные плавники — черта, унаследованная от их челюстных потомков . [13]
Воспроизведение
[ редактировать ]Оплодотворение у миног наружное. Способ оплодотворения миксин неизвестен. Развитие в обеих группах, вероятно, носит внешний характер. Нет никакой известной родительской заботы. О репродуктивном процессе миксины известно немного. Считается, что миксина за всю жизнь откладывает только 30 яиц. [14] Личиночной стадии, характерной для миноги, очень мало. Минога способна размножаться только один раз. миноги После внешнего оплодотворения клоаки остаются открытыми, что позволяет грибку проникнуть в их кишечник и убить их. Миноги размножаются в пресноводных руслах рек, работая парами, строя гнезда и закапывая икру примерно на дюйм ниже осадка. Вылупившиеся птенцы проходят четыре года личиночного развития, прежде чем стать взрослыми.
Эволюция
[ редактировать ]
Хотя бесчелюстные являются второстепенным элементом современной морской фауны , они занимали видное место среди ранних рыб раннего палеозоя . два типа раннекембрийских животных, по-видимому, имеющих плавники, мускулатуру позвоночных известны Из раннекембрийских сланцев Маотяньшань в Китае и жабры : Haikouichthys и Myllokunmingia . Жанвье предварительно назначил их Агнате. Третья возможная анатида из того же региона — Haikouella . О возможной бесчелюстной особи, которая не была официально описана, сообщил Симонетти из среднекембрийских сланцев Бёрджесс в Британской Колумбии . Конодонты — класс бесчелюстных, возникший в раннем кембрии. [16] оставались достаточно распространенными до их исчезновения в триасе, поэтому их зубы (единственные их части, которые обычно были окаменелыми) часто используются в качестве индексных окаменелостей от позднего кембрия до триаса. [17]
Многие ордовикские, силурийские и девонские бесчелюстные были бронированы тяжелыми костяными пластинами. Первые панцирные бесчелюстные — остракодермы , предшественники костистых рыб и, следовательно, четвероногих (включая человека ) — известны со среднего ордовика , а к позднему силуру бесчелюстные достигли высшей точки своей эволюции. Большинство остракодерм, таких как телодонты , остеостраканы и галеаспиды , были более тесно связаны с челюстноротыми, чем с выжившими бесчелюстными, известными как круглоротые. Круглоротые, очевидно, отделились от других бесчелюстных до появления дентина и кости, которые присутствуют у многих ископаемых бесчелюстных, включая конодонтов . [18] Численность Agnathans в девоне сократилась и так и не восстановилась.
Примерно 500 миллионов лет назад у позвоночных возникли два типа рекомбинаторных адаптивных иммунных систем (АИС). Челюстные позвоночные диверсифицируют свой репертуар рецепторов антигенов Т- и В-клеток на основе доменов иммуноглобулина в основном за счет реаранжировки генных сегментов V(D)J и соматической гипермутации, но ни один из фундаментальных элементов распознавания AIS у челюстных позвоночных не был обнаружен у бесчелюстных позвоночных. . Вместо этого AIS бесчелюстных позвоночных основан на вариабельных рецепторах лимфоцитов (VLR), которые генерируются посредством рекомбинаторного использования большой панели очень разнообразных последовательностей с богатыми лейцином повторами (LRR). [19] Три гена VLR (VLRA, VLRB и VLRC) были идентифицированы у миног и миксин и экспрессируются на трех различных линиях лимфоцитов. Клетки VLRA+ и клетки VLRC+ подобны Т-клеткам и развиваются в лимфоэпителиальной структуре, подобной тимусу, называемой тимоидами. Клетки VLRB+ подобны B-клеткам, развиваются в кроветворных органах и дифференцируются в плазматические клетки, секретирующие антитела VLRB. [20]
Классификация
[ редактировать ]| Подгруппа | Пример | Комментарии | |
|---|---|---|---|
| Круглоротые | Миксини |  | Миксини (мисины) — это морские животные в форме угря , производящие слизь (иногда называемые слизевыми угрями ). Это единственные известные живые животные, у которых есть череп , но нет позвоночного столба . Эта группа пережила самую обширную потерю генов среди всех позвоночных: пропало 1386 семейств генов. [21] , как и Микины миноги, лишены челюстей и являются живыми ископаемыми ; миксины являются базальными по отношению к позвоночным, и современные миксины остаются похожими на миксин, существовавших 300 миллионов лет назад. [22] Классификация миксин вызывает споры. Вопрос в том, является ли миксина сама по себе выродившимся типом позвоночных рыб (наиболее близким родственником миног) или может представлять собой стадию, предшествующую эволюции позвоночного столба (как ланцетники ). В исходной схеме миксины и миноги группируются вместе как круглоротые (или исторически Agnatha), как старейшая из сохранившихся клад позвоночных животных, наряду с челюстноротыми (ныне повсеместно встречающимися челюстными позвоночными). Альтернативная схема предполагала, что челюстные позвоночные более тесно связаны с миногами, чем с миксинами (т.е. что позвоночные включают миног, но исключают миксин), и вводит категорию craniata для группы позвоночных, близких к миксинам. Недавние данные ДНК подтвердили первоначальную схему. [9] |
| Гипероартия |  | Hyperoartia — спорная группа позвоночных, в которую входят современные миноги и их ископаемые родственники. Примерами гипероартиан из ранней летописи окаменелостей являются Endeiolepis и Euphanerops , рыбоподобные животные с гипоцеркальными хвостами , которые жили в позднем девонском периоде. Некоторые палеонтологи до сих пор относят эти формы к « остракодермам » (бесчелюстным панцирным «рыбам») класса Anaspida , но это все чаще считается искусственным расположением, основанным на наследственных чертах . Отнесение этой группы к бесчелюстным позвоночным является предметом споров. Хотя сегодня известно достаточно разнообразия окаменелостей, чтобы сделать тесную связь между «остракодермами» маловероятной, это запутало вопрос о ближайших родственниках Hyperoartia. Традиционно группу относили к надклассу Cyclostomata вместе с миксинами (мисинами). Совсем недавно было высказано предположение, что миксины являются более базальными среди хордовых, несущих череп , тогда как гипероартии сохраняются у позвоночных. . Но даже если это, возможно, и верно, миноги представляют собой одно из старейших отклонений линии позвоночных, и вопрос о том, лучше ли они связаны с некоторыми «остракодермами» цефаласпидоморфов или не ближе к ним, чем, например, к другим «остракодермам» цефаласпидоморфов? Pteraspidomorphi . или даже давно вымершие конодонты все еще предстоит определить Даже само существование Hyperoartia оспаривается, при этом некоторые анализы отдают предпочтение трактовке «базальной Hyperoartia» как монофилетической линии Jamoytiiformes , которая на самом деле может быть очень близка к предкам челюстных позвоночных . | |
| Миллокунмингиида | † Миллокунмингииды (вымерший) |  | Myllokunmingiids кембрийского были примитивным отрядом бесчелюстных, которые были эндемиками возраста лагерштетте сланцев Маотяньшаньских в Китае . Эти существа представляют собой самую раннюю известную группу краниатов (хордовых животных с черепом из твердых костей или хрящей ). В настоящее время в группу входят 3 известных рода: Haikouichthys , Myllokunmingia и Zhongjianichthys . [23] [24] |
| Конодонта | † Конодонт |  | Конодонты — это угри, похожие на бесчелюстных, которые жили от кембрия до начала юрского периода. Они были очень разнообразны с точки зрения образа жизни: некоторые виды были фильтраторами , а другие - макрохищниками. На протяжении более столетия эти животные были известны только благодаря своим микроскопическим фосфатным структурам зубов, называемым « элементами конодонта ». Лишь в середине 1980-х годов в Шотландии и Висконсине были найдены окаменелости тел конодонтов , демонстрирующие истинный внешний вид этих животных. Их зубы являются отличными ископаемыми, поскольку многие виды жили и вымерли за относительно короткий период времени. Эти рыбы достигли своего пика разнообразия в середине ордовика, но сильно пострадали от ордовикско-силурийского вымирания . Затем они достигли еще одного всплеска разнообразия в середине позднего девона, а затем снова снизились в каменноугольном периоде . они были относительно редки В перми , но резко увеличились в численности в раннем триасе . Несмотря на это, они вымерли в нижнем юрском периоде, и некоторые из последних выживших популяций обитали в Японии. Возможно, они прожили там дольше из-за относительной удаленности местности. Первоначально считалось, что они были уничтожены крупное вымирание в конце триаса. Вместо этого теперь считается, что они вытеснили более новые мезозойские таксоны. [25] [26] [27] [28] [29] [30] |
| Остракодермы | † Птераспидоморфы (вымерший) |  | † Pteraspidomorphi — вымершая группа ранних бесчелюстных рыб. Окаменелости показывают обширную защиту головы. У многих были гипоцеркальные хвосты, которые создавали подъемную силу и облегчали передвижение по воде их бронированных тел, покрытых кожной костью. У них также были сосущие части рта, и некоторые виды, возможно, жили в пресной воде. Таксон содержит подгруппы Heterostraci , Astraspida , Arandaspida . |
| † Телодонти (вымерший) |  | Телодонты (сосковые зубы) — это группа мелких вымерших бесчелюстных рыб с характерной чешуей вместо больших пластин панциря. Существует много споров о том, является ли группа палеозойских рыб, известная как Thelodonti (ранее целолепиды), [31] ) представляют собой монофилетическую группу или разрозненные стволовые группы основных линий бесчелюстных и челюстных рыб . Телодонты объединены обладателями « телодонтовой чешуи ». Этот определяющий признак не обязательно является результатом общего происхождения, поскольку он мог быть развит независимо разными группами . Таким образом, обычно считается, что телодонты представляют собой полифилетическую группу. [32] хотя по этому вопросу нет твердого согласия; если они монофилетичны, нет убедительных доказательств того, каким было их предковое состояние. [33] : 206 «Телодонты» были морфологически очень похожи и, вероятно, тесно связаны с рыбами классов Heterostraci и Anaspida , отличаясь главным образом покрытием из характерных мелких колючих чешуек. Эти чешуйки легко рассеялись после смерти; их небольшой размер и устойчивость сделали их наиболее распространенными ископаемыми позвоночных своего времени. [34] [35] Рыбы жили как в пресноводной, так и в морской среде, впервые появившись в ордовике и исчезнув во время франско-фаменского вымирания в позднем девоне . Они занимали самые разнообразные экологические ниши, причем большое количество видов предпочитало рифовые экосистемы, где их гибкие тела чувствовали себя более комфортно, чем тяжелобронированные туши других бесчелюстных рыб. [36] | |
| † Анаспида (вымерший) | 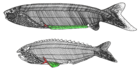 | Анаспиды (без щитка) — вымершая группа примитивных бесчелюстных позвоночных, живших в силурийский и девонский периоды. [37] Их традиционно считают предками миног. [38] Анаспиды были небольшими морскими бесчелюстными, у которых не было тяжелого костного щита и парных плавников, но они имели поразительный сильно гипоцеркальный хвост. Впервые они появились в раннем силуре и процветали до позднего девона . [39] где большинство видов, за исключением миног , вымерло из-за экологических потрясений, произошедших в то время. | |
| † Цефаласпидо- морфи (вымерший) |  | Cephalaspidomorphi — обширная группа вымерших панцирных бесчелюстных, обнаруженных в силурийских и девонских слоях Северной Америки, Европы и Китая, и названа в честь остеостраканов рода Cephalaspis . Большинство биологов считают этот таксон вымершим, но это название иногда используется при классификации миног , поскольку иногда считается, что миноги связаны с цефаласпидами. Если включить миног, это расширит известный ареал группы от раннего силура до мезозоя и до наших дней. Цефаласпидоморфы, как и большинство современных рыб, были очень хорошо бронированы. Особенно хорошо был развит головной щит, защищающий голову, жабры и передний отдел внутренностей. Корпус в большинстве случаев также был хорошо бронирован. Головной щит имел ряд борозд по всей поверхности, образующих обширный орган боковой линии . Глаза были довольно маленькими и располагались на макушке. Не было подходящей челюсти . Ротовое отверстие было окружено небольшими пластинками, делавшими губы гибкими, но лишенными возможности кусать. [40] Бесспорные подгруппы, традиционно входящие в состав Cephaloaspidomorphi, также называемые «Monorhina», включают классы Osteostraci , Galeaspida и Pituriaspida. | |
Филогения
[ редактировать ]Филогения основана на работах Микко Хаарамо и Дельсука и др. [41] [42]
| Позвоночные / |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Череп |
Хотя «агната» Conodonta действительно была безчелюстной, если бы она продолжала жить, ее потомки все равно были бы более близкими родственниками, например, людям, чем миногам, а также в то время были более близкими родственниками предка человека. По таким соображениям Agnatha не может быть объединена в последовательную группу без удаления каких-либо некруглоротых или без включения всех позвоночных, что превратит ее в младший синоним позвоночных.
Новая филогения от Miyashita et al . (2019) считается совместимым как с морфологическими, так и с молекулярными данными. [42] [43]
| Позвоночные / | Агната | |
| Череп |











См. также
[ редактировать ]- челюстноротые
- Амфирхина , альтернативное название вышеупомянутой параллельной или сестринской классификации.
- Круглоротые
Ссылки
[ редактировать ]- ^ Ян, Чуан; Ли, Сянь-Хуа; Чжу, Маоянь; Кондон, Дэниел Дж.; Чен, Цзюнюань (2018). «Геохронологические ограничения кембрийской биоты Чэнцзян, Южный Китай» (PDF) . Журнал Геологического общества . 175 (4): 659–666. Бибкод : 2018JGSoc.175..659Y . дои : 10.1144/jgs2017-103 . ISSN 0016-7649 . S2CID 135091168 .
- ^ Мерсс, Т.; Миллер, CG (2004). «Телодонты и распространение связанных с ними конодонтов из лландовери-самого нижнего лохкова валлийского пограничья». Палеонтология . 47 (5): 1211–1265. Бибкод : 2004Palgy..47.1211M . дои : 10.1111/j.0031-0239.2004.00409.x . [В. Кисслинг/М. Краузе/Э. Ито]
- ^ Краткий Оксфордский словарь английского языка
- ^ Руджеро, Майкл; Гордон, Деннис П.; Оррелл, Томас М.; Байи, Николя (апрель 2015 г.). «Классификация всех живых организмов более высокого уровня» . ПЛОС Один . 10 (4): e0119248. Бибкод : 2015PLoSO..1019248R . дои : 10.1371/journal.pone.0119248 . ПМЦ 4418965 . ПМИД 25923521 .
- ^ Хеймберг, Алиша М.; Каупер-Салари, Ричард; Семон, Мари; Донохью, Филип CJ; Петерсон, Кевин Дж. (9 ноября 2010 г.). «МикроРНК раскрывают взаимоотношения миксин, миног и челюстноротых, а также природу предков позвоночных» . Труды Национальной академии наук . 107 (45): 19379–19383. дои : 10.1073/pnas.1010350107 . ПМЦ 2984222 . ПМИД 20959416 .
- ^ Jump up to: а б с Маллат, Дж.; Салливан, Дж. (декабрь 1998 г.). «Последовательности 28S и 18S рДНК поддерживают монофилию миног и миксин». Молекулярная биология и эволюция . 15 (12): 1706–1718. doi : 10.1093/oxfordjournals.molbev.a025897 . ПМИД 9866205 .
- ^ Jump up to: а б с Делабр С., Галлут С., Барриэль В., Жанвье П., Гашелен Г. (февраль 2002 г.). «Полная митохондриальная ДНК миксины Eptatretus burgeri : сравнительный анализ последовательностей митохондриальной ДНК убедительно подтверждает монофилию круглоротых». Молекулярная филогенетика и эволюция . 22 (2): 184–92. Бибкод : 2002МОЛПЭ..22..184Д . дои : 10.1006/mpev.2001.1045 . ПМИД 11820840 .
- ^ Оиси Ю., Ота К.Г., Кураку С., Фудзимото С., Куратани С. (январь 2013 г.). «Черепно-лицевое развитие миксин и эволюция позвоночных». Природа . 493 (7431): 175–80. Бибкод : 2013Natur.493..175O . дои : 10.1038/nature11794 . hdl : 20.500.14094/D1005717 . ПМИД 23254938 . S2CID 4403344 .
- ^ Jump up to: а б Жанвье, П. (ноябрь 2010 г.). «МикроРНК возрождают старые взгляды на дивергенцию и эволюцию бесчелюстных позвоночных» . Труды Национальной академии наук Соединенных Штатов Америки . 107 (45): 19137–19138. Бибкод : 2010PNAS..10719137J . дои : 10.1073/pnas.1014583107 . ПМК 2984170 . ПМИД 21041649 .
Хотя я был одним из первых сторонников парафилии позвоночных, меня впечатляют доказательства, предоставленные Heimberg et al . и готовы признать, что круглоротые на самом деле монофилетичны. В результате они мало что могут рассказать нам о заре эволюции позвоночных, за исключением того, что интуиция зоологов XIX века была верна, полагая, что эти странные позвоночные (в частности, миксины) сильно выродились и утратили многие признаки с течением времени. время.
- ^ Склад, ДВ; Уитт, Г.С. (август 1992 г.). «Свидетельства на основе последовательностей рибосомальной РНК 18S о том, что миноги и миксины образуют естественную группу». Наука . 257 (5071): 787–9. Бибкод : 1992Sci...257..787S . дои : 10.1126/science.1496398 . ПМИД 1496398 .
- ^ Ота К.Г., Фудзимото С., Оиси Ю., Куратани С. (июнь 2011 г.). «Идентификация позвоночноподобных элементов и возможная их дифференциация от склеротомов у миксины» . Природные коммуникации . 2 (6): 373. Бибкод : 2011NatCo...2..373O . дои : 10.1038/ncomms1355 . ПМК 3157150 . ПМИД 21712821 .
- ^ Склад, ДВ; Уитт, Г.С. (август 1992 г.). «Свидетельства на основе последовательностей рибосомальной РНК 18S о том, что миноги и миксины образуют естественную группу». Наука . 257 (5071): 787–789. Бибкод : 1992Sci...257..787S . дои : 10.1126/science.1496398 . ПМИД 1496398 .
- ^ Ромер, А.С. и Парсонс, Т.С. (1985): Тело позвоночного. (6-е изд.) Сондерс, Филадельфия.
- ^ «Хагфиш» . Aquaticcommunity.com . Проверено 30 июня 2013 г.
- ^ Бентон, MJ (2005) Палеонтология позвоночных , Блэквелл, 3-е издание, рисунок 3.25 на странице 73, ISBN 0-632-05637-1 .
- ^ Стэнли, Стивен М.; Лучай, Джон А. (2015). История системы Земли (4-е изд.). Макмиллан Образование. п. 311.
Конодонты возникли в конце раннего кембрия и диверсифицировались в ордовике. ... Подобные маленькие зубы в фаунах очень раннего кембрия ... могут представлять предков конодонтов.
- ^ Милый, туалет; Донохью, PCJ (2001). «Конодонты: прошлое, настоящее и будущее» (PDF) . Журнал палеонтологии . 75 (6): 1174–1184. Бибкод : 2001JPal...75.1174S . doi : 10.1666/0022-3360(2001)075<1174:CPPF>2.0.CO;2 . ISSN 0022-3360 . S2CID 53395896 . Архивировано (PDF) из оригинала 30 октября 2022 г.
- ^ Резюме Бейкера (декабрь 2008 г.). «Эволюция и развитие клеток нервного гребня позвоночных». Текущее мнение в области генетики и развития . 18 (6): 536–543. дои : 10.1016/j.где.2008.11.006 . ПМИД 19121930 .
- ^ Хирано, Масаюки; Дас, Сабьясачи; Го, Пэн; Купер, Макс Д. (1 января 2011 г.). «Глава 4 – Эволюция адаптивного иммунитета у позвоночных» . В Alt, Фредерик В. (ред.). Достижения иммунологии . Том. 109. Академическая пресса. стр. 125–157. дои : 10.1016/b978-0-12-387664-5.00004-2 . ISBN 9780123876645 . ПМИД 21569914 . Проверено 3 декабря 2019 г.
- ^ Ву, Фэнфан; Чен, Лиюн; Рен, Ён; Ян, Сяоцзин; Ю, Тунчжоу; Фэн, Бо; Чен, Шанву; Сюй, Аньлун (октябрь 2016 г.). «Ингибирующий рецептор VLRB у бесчелюстной миноги» . Научные отчеты . 6 (1): 33760. Бибкод : 2016NatSR...633760W . дои : 10.1038/srep33760 . ISSN 2045-2322 . ПМК 5071834 . ПМИД 27762335 .
- ^ Марлетаз, Фердинанд; Тимошевская, Наталья; Тимошевский Владимир А.; Парей, Элиза; Симаков Олег; Гаврючкина Дарья; Сузуки, Масакадзу; Кубокава, Каору; Бреннер, Сидней; Смит, Джерамиа Дж.; Рохсар, Дэниел С. (2024). «Геном миксины и эволюция позвоночных» . Природа . 627 (8005): 811–820. Бибкод : 2024Натуральный.627..811М . дои : 10.1038/s41586-024-07070-3 . ISSN 1476-4687 . ПМЦ 10972751 . ПМИД 38262590 .
- ^ Спир, Брайан Р. (1997). «Знакомство с миксинами» . Музей палеонтологии Калифорнийского университета. Калифорнийский университет, Беркли. Архивировано из оригинала 15 декабря 2017 г. Проверено 21 февраля 2013 г.
- ^ Шу, Деган (апрель 2003 г.). «Палеонтологический взгляд на происхождение позвоночных». Китайский научный бюллетень . 48 (8): 725–735. Бибкод : 2003ЧСБу..48..725С . дои : 10.1007/BF03187041 . S2CID 85163902 .
- ^ ван дер Лаан, Ричард (2016). « Названия семейной группы ископаемых рыб ». Европейский журнал таксономии (466). дои : 10.5852/ejt.2018.466 .
- ^ Мило, Уолтер К.; Купер, Барри Дж. (декабрь 2008 г.). «Знакомство Ч. Пандера с конодонтами, 1856 г.» . Эпизоды . 31 (4): 429–432. дои : 10.18814/epiiugs/2008/v31i4/009 . Проверено 3 января 2019 г.
- ^ Габботт, SE; Р. Дж. Олдридж; Дж. Н. Терон (1995). «Гигантский конодонт с сохранившейся мышечной тканью из верхнего ордовика Южной Африки». Природа . 374 (6525): 800–803. Бибкод : 1995Natur.374..800G . дои : 10.1038/374800a0 . S2CID 4342260 .
- ^ Куинтон, Пейдж С. (2016). «Влияние протоколов экстракции на изотопный состав кислорода элементов конодонтов». Химическая геология . 431 : 36–43. Бибкод : 2016ЧГео.431...36Q . doi : 10.1016/j.chemgeo.2016.03.023 .
- ^ Бергстрем, С.М.; Карнс, Дж.Б.; Этингтон, РЛ; Вотав, РБ; Вигли, П.Б. (1974). «Appalachignathus, новый многоэлементный род конодонтов из среднего ордовика Северной Америки». Журнал палеонтологии . 48 (2): 227–235. Бибкод : 2001JPal...75.1174S . doi : 10.1666/0022-3360(2001)075<1174:CPPF>2.0.CO;2 . JSTOR 1303249 . S2CID 53395896 .
- ^ Гино, Самуэль; Гудеманд, Николя (декабрь 2020 г.). «Глобальные изменения климата являются причиной основных тенденций разнообразия конодонтов, но не их окончательной гибели» (PDF) . Глобальные и планетарные изменения . 195 : 103325. Бибкод : 2020GPC...19503325G . дои : 10.1016/j.gloplacha.2020.103325 . S2CID 225005180 .
- ^ Ду, Исин; Оноуэ, Тецудзи; Томимацу, Юки; У, Цянван; Риго, Мануэль (2023). «Конодонты нижней юры из района Инуяма в Японии: последствия исчезновения конодонтов» . Границы экологии и эволюции . 11 . дои : 10.3389/fevo.2023.1135789 . hdl : 11577/3479836 . ISSN 2296-701X .
- ^ Тернер С., Тарлинг Д.Х. (1982). «Распространение телодонта и других бесчелюстных как тест континентальных реконструкций нижнего палеозоя». Палеогеография, Палеоклиматология, Палеоэкология . 39 (3–4): 295–311. Бибкод : 1982PPP....39..295T . дои : 10.1016/0031-0182(82)90027-X .
- ^ Сарджант В.А., Холстед Л.Б. (1995). Окаменелости позвоночных и эволюция научных концепций: сочинения в честь Беверли Холстед . Гордон и Брич. ISBN 978-2-88124-996-9 .
- ^ Донохью ПК, Фори П.Л., Олдридж Р.Дж. (май 2000 г.). «Сродство конодонтов и филогения хордовых». Биологические обзоры Кембриджского философского общества . 75 (2): 191–251. дои : 10.1111/j.1469-185X.1999.tb00045.x . ПМИД 10881388 . S2CID 22803015 .
- ^ Тернер С. (1999). «Комплексы телодонтов от раннего силура до раннего девона и их возможное экологическое значение». В А. Дж. Буко, Дж. Лоусоне (ред.). Палеосообщества – Экостратиграфия проекта, Заключительный отчет (Отчет). Международная программа геологической корреляции. Том. 53. Издательство Кембриджского университета . стр. 42–78.
- ^ Ранний и средний силур. Видеть Казлев М.А., Уайт Т (6 марта 2001 г.). «Телодонти» . Палеос.com . Архивировано из оригинала 28 октября 2007 г. Проверено 30 октября 2007 г.
- ^ Феррон Х.Г., Ботелла Х. (2017). «Сквамация и экология телодонтов» . ПЛОС ОДИН . 12 (2): e0172781. Бибкод : 2017PLoSO..1272781F . дои : 10.1371/journal.pone.0172781 . ПМК 5328365 . ПМИД 28241029 .
- ^ Альберг П.Е. (2001). Основные события ранней эволюции позвоночных: палеонтология, филогения, генетика и развитие . Вашингтон, округ Колумбия: Тейлор и Фрэнсис. п. 188. ИСБН 978-0-415-23370-5 .
- ^ Паттерсон, Колин (1987). Молекулы и морфология в эволюции: конфликт или компромисс? . Кембридж, Великобритания: Издательство Кембриджского университета. п. 142. ИСБН 978-0-521-32271-3 .
- ^ Холл, Брайан Кейт; Ханкен, Джеймс (1993). Череп . Чикаго, Иллинойс: Издательство Чикагского университета. п. 131. ИСБН 978-0-226-31568-3 .
- ^ Кольбер, Майкл; Моралес, Эдвин Х. (1991). Эволюция позвоночных: история позвоночных животных во времени (4-е изд.). Нью-Йорк, штат Нью-Йорк: Вили-Лисс. ISBN 978-0-471-85074-8 .
- ^ Хаарамо, Микко (2007). « Хордовые – ланцеты, оболочники и позвоночные» . Архив филогении Микко . Проверено 30 декабря 2016 г.
- ^ Jump up to: а б Дельсук Ф, Филипп Х, Цагкогеорга Г, Симион П, Тилак МК, Турон Х, Лопес-Легентиль С, Пиетт Дж, Лемэр П, Дузери Э.Дж. (апрель 2018 г.). «Филогеномная основа и временные рамки сравнительных исследований оболочников» . БМК Биология . 16 (1): 39. дои : 10.1186/s12915-018-0499-2 . ПМК 5899321 . ПМИД 29653534 .
- ^ Мияшита, Тецуто; Коутс, Майкл И.; Фаррар, Роберт; Ларсон, Питер; Мэннинг, Филип Л.; Вогелиус, Рой А.; и др. (2019). «Мексика из мелового моря Тетис и урегулирование морфолого-молекулярного конфликта в ранней филогении позвоночных» . Труды Национальной академии наук Соединенных Штатов Америки . 116 (6): 2146–2151. Бибкод : 2019PNAS..116.2146M . дои : 10.1073/pnas.1814794116 . ПМК 6369785 . ПМИД 30670644 .

| Базы данных органов управления : Национальные |
|---|