Слитый белок
Слитые белки или химерные (ки-мир-ик) белки (буквально состоящие из частей из разных источников) — это белки, созданные путем соединения двух или более генов , которые первоначально кодировали отдельные белки. Трансляция этого слитого гена приводит к образованию одного или нескольких полипептидов с функциональными свойствами, полученными из каждого из исходных белков. Рекомбинантные слитые белки создаются искусственно с помощью технологии рекомбинантной ДНК для использования в биологических исследованиях или терапии . Химерами или химерами обычно обозначают гибридные белки, состоящие из полипептидов, имеющих разные функции или физико-химические характеристики. Химерные мутантные белки возникают в природе, когда сложная мутация , такая как хромосомная транслокация , тандемная дупликация или ретротранспозиция, создает новую кодирующую последовательность, содержащую части кодирующих последовательностей двух разных генов. Встречающиеся в природе гибридные белки обычно обнаруживаются в раковых клетках, где они могут функционировать как онкопротеины . Слитый белок bcr-abl является хорошо известным примером онкогенного слитого белка и считается основным онкогенным фактором хронического миелогенного лейкоза .
Функции
[ редактировать ]Некоторые слитые белки объединяют целые пептиды и, следовательно, содержат все функциональные домены исходных белков. Однако другие слитые белки, особенно те, которые встречаются в природе, объединяют только части кодирующих последовательностей и поэтому не сохраняют исходные функции родительских генов, которые их сформировали.
Многие слияния целых генов полностью функциональны и могут заменять исходные пептиды. Однако некоторые испытывают взаимодействие между двумя белками, которое может изменить их функции. Помимо этих эффектов, некоторые слияния генов могут вызывать регуляторные изменения , которые изменяют время и место действия этих генов. При частичном слиянии генов перетасовка различных активных сайтов и связывающих доменов может привести к появлению новых белков с новыми функциями.
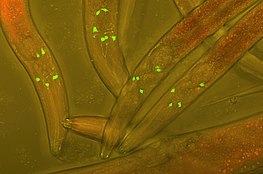
Метки флуоресцентного белка
[ редактировать ]Слияние флуоресцентных меток с белками в клетке-хозяине — широко популярный метод, используемый в экспериментальных клеточных и биологических исследованиях для отслеживания взаимодействий белков в реальном времени. Первая флуоресцентная метка, зеленый флуоресцентный белок (GFP), была выделена из Aequorea victoria и до сих пор часто используется в современных исследованиях. Более поздние разработки включают фотоконвертируемые флуоресцентные белки (PCFP), которые были впервые выделены из Anthozoa . Наиболее часто используемой PCFP является флуоресцентная метка Kaede , но разработка зелено-красной метки Kikume (KikGR) в 2005 году предлагает более яркий сигнал и более эффективное фотопреобразование. Преимуществом использования флуоресцентных меток PCFP является возможность отслеживать взаимодействие перекрывающихся биохимических путей в режиме реального времени. Метка изменит цвет с зеленого на красный, как только белок достигнет интересующей точки на пути, а белок другого цвета можно будет отслеживать на протяжении всего пути. Этот метод особенно полезен при изучении Пути рециркуляции рецептора, связанного с G-белком (GPCR). Судьба переработанных рецепторов G-белка может быть отправлена либо на плазматическую мембрану для переработки, что отмечено зеленой флуоресцентной меткой, либо может быть отправлено в лизосому для деградации, что отмечено красной флуоресцентной меткой. [ 1 ]
Химерные белковые препараты
[ редактировать ]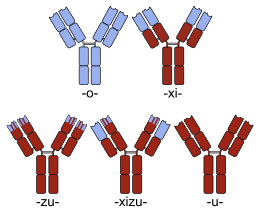
Целью создания гибридных белков при разработке лекарств является придание свойств каждого из «родительских» белков полученному химерному белку. несколько химерных белковых препаратов В настоящее время для медицинского применения доступно .
Многие химерные белковые препараты представляют собой моноклональные антитела , специфичность которых к целевой молекуле была разработана с использованием мышей и, следовательно, изначально представляли собой «мышиные» антитела. Будучи белками, не принадлежащими человеку, мышиные антитела имеют тенденцию вызывать иммунную реакцию при введении людям. Процесс химеризации включает замену сегментов молекулы антитела, которые отличают его от человеческого антитела. Например, можно ввести константные домены человека , тем самым устраняя большую часть потенциально иммуногенных частей лекарственного средства без изменения его специфичности в отношении намеченной терапевтической мишени. Номенклатура антител указывает на этот тип модификации путем вставки -xi- в непатентованное название (например, abci -xi -mab ). Если части вариабельных доменов также заменить человеческими частями, гуманизированные получают антитела. Хотя концептуально этот тип не отличается от химер, он обозначается с помощью -zu-, например, в dacli- zu -mab . см . в списке моноклональных антител Дополнительные примеры .
Помимо химерных и гуманизированных антител, существуют и другие фармацевтические цели создания химерных конструкций. Этанерцепт , например, представляет собой блокатор TNFα, созданный посредством комбинации рецептора фактора некроза опухоли (TNFR) с иммуноглобулина G 1 Fc-сегментом . TNFR обеспечивает специфичность в отношении мишени лекарственного средства, а Fc-сегмент антитела, как полагают, повышает стабильность и доставляемость лекарственного средства. [ 2 ] Дополнительные химерные белки, используемые для терапевтических целей, включают:
- Афлиберцепт : человеческий рекомбинантный белок, который помогает в лечении оксалиплатин-резистентного метастатического колоректального рака , неоваскулярной макулярной дегенерации и макулярного отека . [ 2 ]
- Рилонацепт : уменьшает воспаление, предотвращая активацию рецепторов IL-1 для лечения криопирин-ассоциированных периодических синдромов (CAPS). [ 2 ]
- Алефацепт : регулирует реакцию Т-клеток путем избирательного воздействия на эффекторные Т-клетки памяти для лечения вульгарного псориаза . [ 2 ]
- Ромиплостим : пептидное антитело, которое лечит иммунную тромбоцитопению . [ 2 ]
- Абатацепт / Белатацепт : препятствует совместной стимуляции Т-клеток при лечении аутоиммунных заболеваний, таких как ревматоидный артрит , псориатический артрит и псориаз . [ 2 ]
- Денилейкин-дифтитокс : лечит лимфому кожи . [ 2 ]
Рекомбинантная технология
[ редактировать ]
Рекомбинантный слитый белок представляет собой белок, созданный посредством генной инженерии слитого гена. Обычно это включает удаление стоп- кодона из последовательности кДНК, кодирующей первый белок, а затем добавление последовательности кДНК второго белка в рамку считывания посредством лигирования или ПЦР с перекрывающимся удлинением . будет экспрессироваться клеткой Эта последовательность ДНК затем в виде одного белка. Белок можно сконструировать так, чтобы он включал полную последовательность обоих исходных белков или только часть любого из них.
Если эти два объекта являются белками, часто также добавляются линкерные (или «спейсерные») пептиды, которые повышают вероятность того, что белки сворачиваются независимо и ведут себя ожидаемым образом. Особенно в том случае, когда линкеры обеспечивают очистку белка , линкеры в слитых белках или пептидах иногда конструируются с сайтами расщепления для протеаз или химических агентов, которые обеспечивают высвобождение двух отдельных белков. Этот метод часто используется для идентификации и очистки белков путем слияния белка GST , пептида FLAG или гекса-гис-пептида (6xHis-метка), которые можно выделить с помощью аффинной хроматографии с никелевыми или кобальтовыми смолами. Ди- или мультимерные химерные белки могут быть получены посредством генной инженерии путем слияния с исходными белками пептидных доменов, которые индуцируют ди- или мультимеризацию искусственных белков (например, стрептавидиновые или лейциновые молнии ). Слитые белки также можно производить с прикрепленными к ним токсинами или антителами для изучения развития заболеваний. Промотор гидрогеназы, P SH был изучен с целью конструирования PSH слияния промотора и gfp с использованием зеленого флуоресцентного белка ( gfp) репортерного гена . [ 3 ]
Рекомбинантная функциональность
[ редактировать ]Новые рекомбинантные технологии позволили улучшить дизайн гибридных белков для использования в таких разнообразных областях, как биодетекция, бумажная и пищевая промышленность, а также биофармацевтика. Недавние улучшения включали слияние отдельных пептидов или фрагментов белков с областями существующих белков, такими как N- и C-концы , и, как известно, усиливают следующие свойства: [ 4 ]
- Каталитическая эффективность : слияние определенных пептидов обеспечивает большую каталитическую эффективность за счет изменения третичной и четвертичной структуры целевого белка. [ 4 ]
- Растворимость . Распространенной проблемой при разработке слитых белков является проблема нерастворимости вновь синтезированных слитых белков в рекомбинантном хозяине, что приводит к чрезмерной агрегации целевого белка в клетке. Могут быть добавлены молекулярные шапероны , которые способны способствовать сворачиванию белка, тем самым лучше разделяя гидрофобные и гидрофильные взаимодействия в растворенном веществе и повышая растворимость белка. [ 4 ]
- Термостабильность : отдельные пептиды или фрагменты белка обычно добавляются для уменьшения гибкости N- или C-конца целевого белка, что усиливает термостабильность и стабилизирует диапазон pH . [ 4 ]
- Ферментативная активность: синтез, включающий введение водородных связей , может использоваться для увеличения общей активности фермента. [ 4 ]
- Уровни экспрессии: добавление многочисленных слитых фрагментов, таких как мальтоза-связывающий белок (MBP) или небольшая убиквитиноподобная молекула ( SUMO ), служит для усиления экспрессии фермента и секреции целевого белка. [ 4 ]
- Иммобилизация: PHA-синтаза, фермент, который позволяет иммобилизовать интересующие белки, является важным тегом слияния в промышленных исследованиях. [ 4 ]
- Качество кристаллов. Качество кристаллов можно улучшить путем добавления ковалентных связей между белками, что помогает в методах определения структуры. [ 4 ]
Дизайн рекомбинантного белка
[ редактировать ]Самые ранние применения дизайна рекомбинантных белков могут быть зафиксированы в использовании одиночных пептидных меток для очистки белков с помощью аффинной хроматографии . С тех пор было разработано множество методов создания слитых белков для самых разных применений, от флуоресцентных белковых меток до лекарств из рекомбинантных слитых белков. Три широко используемых метода проектирования включают тандемное слияние, вставку домена и посттрансляционную конъюгацию. [ 5 ]
Тандемный синтез
[ редактировать ]Интересующие белки просто соединяются конец в конец посредством слияния N- или C-концов между белками. Это обеспечивает гибкую структуру моста, обеспечивающую достаточное пространство между партнерами по слиянию для обеспечения правильного складывания . Однако N- или C-концы пептида часто являются решающими компонентами в получении желаемой структуры сворачивания рекомбинантного белка, что делает простое сквозное соединение доменов неэффективным в этом случае. По этой причине часто необходим белковый линкер для поддержания функциональности интересующих белковых доменов. [ 5 ]
Вставка домена
[ редактировать ]Этот метод включает слияние последовательных белковых доменов путем кодирования желаемых структур в одну полипептидную цепь, но иногда может потребоваться вставка домена в другой домен. Этот метод обычно считается более сложным для выполнения, чем тандемное слияние, из-за трудностей с поиском подходящего сайта лигирования в интересующем гене. [ 5 ]
Посттрансляционное спряжение
[ редактировать ]Этот метод объединяет белковые домены после рибосомальной трансляции интересующих белков, в отличие от генетического слияния перед трансляцией, используемого в других рекомбинантных технологиях. [ 5 ]
Белковые линкеры
[ редактировать ]
Белковые линкеры помогают проектировать слитые белки, обеспечивая соответствующее расстояние между доменами, поддерживая правильное сворачивание белка в случае, когда взаимодействия N- или C-концев имеют решающее значение для сворачивания. Обычно белковые линкеры обеспечивают важные взаимодействия доменов, повышают стабильность и уменьшают стерические препятствия, что делает их предпочтительными для использования в дизайне слитых белков, даже когда N- и C-концы могут быть слиты. Три основных типа линкеров — гибкие, жесткие и расщепляемые in vivo. [ 5 ] [ 6 ]
- Гибкие линкеры могут состоять из множества небольших остатков глицина , что придает им способность скручиваться в динамичную, адаптируемую форму. [ 6 ]
- Жесткие линкеры могут быть образованы из крупных циклических остатков пролина , что может быть полезно, когда необходимо поддерживать высокоспецифическое расстояние между доменами. [ 6 ]
- Расщепляемые in vivo линкеры уникальны тем, что они предназначены для высвобождения одного или нескольких слитых доменов при определенных условиях реакции, таких как определенный градиент pH , или при контакте с другой биомолекулой в клетке. [ 6 ]
Естественное явление
[ редактировать ]Встречающиеся в природе слитые гены чаще всего создаются, когда хромосомная транслокация заменяет концевые экзоны одного гена неповрежденными экзонами второго гена. В результате создается единственный ген, который можно транскрибировать , сплайсировать и транслировать для получения функционального слитого белка. Многие важные раку , способствующие онкогены , представляют собой слитые гены, полученные таким путем.
Примеры включают в себя:
Антитела представляют собой слитые белки, образующиеся в результате рекомбинации V(D)J .
Существуют также редкие примеры встречающихся в природе полипептидов, которые представляют собой слияние двух четко определенных модулей, в которых каждый модуль проявляет свою характерную активность или функцию независимо от другого. Двумя основными примерами являются: двойная химера PP2C у Plasmodium falciparum (паразита малярии), в которой каждый модуль PP2C проявляет ферментативную активность протеинфосфатазы 2C, [ 7 ] и иммунофилины двойного семейства , которые встречаются у ряда одноклеточных организмов (таких как простейшие паразиты и флавобактерии ) и содержат полноразмерные циклофилина и шаперона FKBP . модули [ 8 ] [ 9 ] Эволюционное происхождение таких химер остается неясным.
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Шмидт А, Визнер Б, Шуляйн Р, Тайхманн А (2014). «Использование зелено-красных слияний Каэде и Кикуме для визуализации живых клеток рецепторов, связанных с G-белком». Экзоцитоз и эндоцитоз . Методы молекулярной биологии. Том. 1174. Нью-Йорк, штат Нью-Йорк: Humana Press. стр. 139–156. дои : 10.1007/978-1-4939-0944-5_9 . ISBN 9781493909438 . ПМИД 24947379 .
- ^ Jump up to: а б с д и ж г Бальдо Б.А. (май 2015 г.). «Химерные слитые белки, используемые для терапии: показания, механизмы и безопасность». Безопасность лекарств . 38 (5): 455–79. дои : 10.1007/s40264-015-0285-9 . ПМИД 25832756 . S2CID 23852865 .
- ^ Джагдер Б.Е., Уэлч Дж., Брейди Н., Маркиз С.П. (26 июля 2016 г.). «Создание и использование слияния растворимого промотора гидрогеназы (PSH) Cupriavidus necator H16 с gfp (зеленым флуоресцентным белком)» . ПерДж . 4 : е2269. дои : 10.7717/peerj.2269 . ПМЦ 4974937 . ПМИД 27547572 .
- ^ Jump up to: а б с д и ж г час Ян Х, Лю Л, Сюй Ф (октябрь 2016 г.). «Перспективы и проблемы слитых конструкций в биохимии и энзимологии белков». Прикладная микробиология и биотехнология . 100 (19): 8273–81. дои : 10.1007/s00253-016-7795-y . ПМИД 27541749 . S2CID 14316893 .
- ^ Jump up to: а б с д и Ю К., Лю С., Ким Б.Г., Ли Д.И. (01.01.2015). «Разработка и применение синтетических слитых белков». Достижения биотехнологии . 33 (1): 155–164. doi : 10.1016/j.biotechadv.2014.11.005 . ПМИД 25450191 .
- ^ Jump up to: а б с д Чен X, Заро JL, Shen WC (октябрь 2013 г.). «Линкеры слитых белков: свойства, дизайн и функциональность» . Обзоры расширенной доставки лекарств . 65 (10): 1357–69. дои : 10.1016/j.addr.2012.09.039 . ПМЦ 3726540 . ПМИД 23026637 .
- ^ Мамун CB, Салливан DJ, Банерджи Р., Голдберг Д.Э. (май 1998 г.). «Идентификация и характеристика необычной двойной серин/треониновой протеинфосфатазы 2C у малярийного паразита Plasmodium falciparum» . Журнал биологической химии . 273 (18): 11241–7. дои : 10.1074/jbc.273.18.11241 . ПМИД 9556615 .
- ^ Адамс Б., Мусиенко А., Кумар Р., Барик С. (июль 2005 г.). «Новый класс иммунофилинов двойного семейства» . Журнал биологической химии . 280 (26): 24308–14. дои : 10.1074/jbc.M500990200 . ПМК 2270415 . ПМИД 15845546 .
- ^ Барик С. (ноябрь 2017 г.). «О роли, экологии, филогении и строении иммунофилинов двойного семейства» . Клеточные стрессы и шапероны . 22 (6): 833–845. дои : 10.1007/s12192-017-0813-x . ПМЦ 5655371 . ПМИД 28567569 .
Внешние ссылки
[ редактировать ]- Мутанты + химерные + белки Национальной медицинской библиотеки США в медицинских предметных рубриках (MeSH)
- ChiPPI. Архивировано 10 ноября 2021 г. в Wayback Machine : Серверное белок-белковое взаимодействие химерных белков.