Термопротеота
| Термопротеота | |
|---|---|
| |
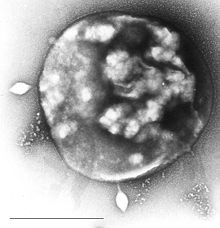
| Археи Sulfolobus заражены специфическим вирусом STSV-1 . | |
| Научная классификация | |
| Домен: | Архея |
| Королевство: | Протеоархеота |
| Супертип: | ТАК группа |
| Тип: | Термопротеота Гаррити и Холт 2021 [ 1 ] |
| Сорт | |
| |
| Синонимы | |
| |
Thermoproteota — это прокариоты , которые были классифицированы как тип домена Archaea . [ 2 ] [ 3 ] [ 4 ] Первоначально считалось, что Thermoproteota являются серозависимыми экстремофилами , но недавние исследования выявили характерную для окружающей среды рРНК Thermoproteota, указывающую на то, что эти организмы могут быть наиболее многочисленными архей в морской среде. [ 5 ] Первоначально их отделили от других архей на основе последовательностей рРНК; другие физиологические особенности, такие как отсутствие гистонов , подтверждают это разделение, хотя было обнаружено, что у некоторых кренархей гистоны имеются. [ 6 ] До 2005 года все культивируемые Thermoproteota были термофильными или гипертермофильными организмами, некоторые из которых обладали способностью расти при температуре до 113 °C. [ 7 ] Эти микроорганизмы окрашиваются грамотрицательно и морфологически разнообразны, имеют палочковидные, кокковые , нитчатые клетки и клетки необычной формы. [ 8 ] Недавние данные показывают, что некоторые представители Thermoproteota являются метаногенами.
Первоначально термопротеоты были классифицированы как часть Regnum Eocyta в 1984 году. [ 9 ] но эта классификация была отвергнута. Термин «эоцит» теперь применяется либо к TACK (ранее Crenarchaeota), либо к Thermoproteota.
Сульфобус
[ редактировать ]Одним из наиболее характерных представителей Crenarchaeota является Sulfolobus solfataricus . Этот организм был первоначально выделен из геотермально нагретых серных источников в Италии и растет при 80 °C и pH 2–4. [ 10 ] С момента его первоначальной характеристики Вольфрамом Циллигом , пионером в области исследований термофилов и архей, похожие виды одного и того же рода были обнаружены по всему миру. В отличие от подавляющего большинства культивируемых термофилов, Sulfolobus растет аэробно и хемоорганотрофно (получая энергию из органических источников, таких как сахара). Эти факторы обеспечивают гораздо более легкий рост в лабораторных условиях, чем анаэробные организмы , и привели к тому, что Sulfolobus стал модельным организмом для изучения гипертермофилов и большой группы разнообразных вирусов, которые реплицируются внутри них.
| на основе 16S рРНК ДП _06_2022 [ 11 ] [ 12 ] [ 13 ] | 53 маркерных белка на основе GTDB 08-RS214 [ 14 ] [ 15 ] [ 16 ] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
|
Рекомбинационная репарация повреждений ДНК
[ редактировать ]Облучение S. solfataricus клеток ультрафиолетовым светом сильно индуцирует образование пилей IV типа , которые затем могут способствовать агрегации клеток. [ 17 ] Аггация клеток, индуцированная ультрафиолетовым светом, была показана Ajon et al. [ 18 ] опосредовать высокочастотный межклеточный обмен маркерами хромосом . Культуры, индуцированные ультрафиолетовым светом, имели скорость рекомбинации, превышающую скорость рекомбинации неиндуцированных культур на целых три порядка. Клетки S. solfataricus способны объединяться только с другими представителями своего вида. [ 18 ] Фролс и др. [ 17 ] [ 19 ] и Аджон и др. [ 18 ] считали, что процесс переноса ДНК, индуцируемый ультрафиолетовым светом, с последующей гомологичной рекомбинационной репарацией поврежденной ДНК , является важным механизмом обеспечения целостности хромосом.
Этот процесс переноса ДНК можно рассматривать как примитивную форму сексуального взаимодействия .
Морские виды
[ редактировать ]Начиная с 1992 года были опубликованы данные о последовательностях генов, принадлежащих Thermoproteota, в морской среде. [ 20 ] , [ 21 ] С тех пор анализ обильных липидов мембран Thermoproteota, взятых из открытого океана, использовался для определения концентрации этих «низкотемпературных Crenarchaea» (см. TEX-86 ). На основании этих измерений их характерных липидов считается, что Thermoproteota очень многочисленны и являются одним из основных участников фиксации углерода . [ нужна ссылка ] Последовательности ДНК Thermoproteota также были обнаружены в почве и пресноводной среде, что позволяет предположить, что этот тип распространен повсеместно в большинстве сред. [ 22 ]
В 2005 году были опубликованы данные о первых культивированных «низкотемпературных кренархеях». Названный Nitrosopumilus maritimus , это окисляющий аммиак организм, выделенный из морского аквариума и выращенный при 28 °C. [ 23 ]
Возможные связи с эукариотами
[ редактировать ]Исследование двухдоменной системы классификации открыло возможности связи между кренархей и эукариотами . [ 24 ]
Анализ ДНК 2008 года (а позже, 2017 года) показал, что эукариоты, возможно, произошли от термопротеотоподобных организмов. К другим кандидатам в предки эукариот относятся близкородственные асгарды . Это может свидетельствовать о том, что эукариотические организмы, возможно, произошли от прокариотов.
Эти результаты аналогичны гипотезе эоцитов 1984 года, предложенной Джеймсом А. Лейком . [ 9 ] Классификация по Лейку гласит, что и кренархеи, и асгарды относятся к царству Eocyta. Хотя ученые отбросили эту идею, основная концепция остается. Термин «эоциты» теперь относится либо к группе TACK , либо к самому типу Thermoproteota.
Тем не менее, эта тема широко обсуждается, и исследования все еще продолжаются.
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Орен А., генеральный менеджер Гаррити (2021 г.). «Действительная публикация названий сорока двух типов прокариот» . Int J Syst Evol Microbiol . 71 (10): 5056. doi : 10.1099/ijsem.0.005056 . ПМИД 34694987 . S2CID 239887308 .
- ^ См. веб-страницу NCBI о Crenarchaeota.
- ^ К.Майкл Хоган. 2010. Археи . ред. Э.Моноссон и К.Кливленд, Энциклопедия Земли. Национальный совет по науке и окружающей среде, Вашингтон, округ Колумбия.
- ^ Данные извлечены из «Ресурсы таксономии NCBI» . Национальный центр биотехнологической информации . Проверено 19 марта 2007 г.
- ^ Мэдиган М; Мартинко Дж., ред. (2005). Брок Биология микроорганизмов (11-е изд.). Прентис Холл. ISBN 978-0-13-144329-7 .
- ^ Кубонова Л., Сэндман К., Халлам С.Дж., Делонг Э.Ф., Рив Дж.Н. (2005). «Гистоны в Кренархее» . Журнал бактериологии . 187 (15): 5482–5485. дои : 10.1128/JB.187.15.5482-5485.2005 . ПМК 1196040 . ПМИД 16030242 .
- ^ Блохл Э., Рэйчел Р., Бургграф С., Хафенбрадль Д., Яннаш Х.В., Стеттер КО (1997). « Pyrolobus fumarii , gen. и sp. nov., представляет собой новую группу архей, расширяющую верхний температурный предел жизни до 113 °C». Экстремофилы . 1 (1): 14–21. дои : 10.1007/s007920050010 . ПМИД 9680332 . S2CID 29789667 .
- ^ Гэррити ГМ, Бун Д.Р., ред. (2001). Руководство Берджи по систематической бактериологии, том 1: Археи, глубоко ветвящиеся и фототрофные бактерии (2-е изд.). Спрингер. ISBN 978-0-387-98771-2 .
- ^ Перейти обратно: а б Лейк Дж.А., Хендерсон Э., Оукс М., Кларк М.В. (июнь 1984 г.). «Эоциты: новая структура рибосом указывает на царство, имеющее близкое родство с эукариотами» . Труды Национальной академии наук Соединенных Штатов Америки . 81 (12): 3786–3790. Бибкод : 1984PNAS...81.3786L . дои : 10.1073/pnas.81.12.3786 . ПМК 345305 . ПМИД 6587394 .
- ^ Зиллиг В., Стеттер КО, Вундерл С., Шульц В., Присс Х., Шольц I (1980). «Группа Sulfolobus-«Калдариеллара»: Таксономия на основе структуры ДНК-зависимых РНК-полимераз». Арх. Микробиол . 125 (3): 259–269. дои : 10.1007/BF00446886 . S2CID 5805400 .
- ^ «ЛТП» . Проверено 10 мая 2023 г.
- ^ «Дерево LTP_all в формате Ньюика» . Проверено 10 мая 2023 г.
- ^ «Примечания к выпуску LTP_06_2022» (PDF) . Проверено 10 мая 2023 г.
- ^ «Выпуск GTDB 08-RS214» . База данных геномной таксономии . Проверено 10 мая 2023 г.
- ^ "ar53_r214.sp_label" . База данных геномной таксономии . Проверено 10 мая 2023 г.
- ^ «История таксонов» . База данных геномной таксономии . Проверено 10 мая 2023 г.
- ^ Перейти обратно: а б Фролс С., Айон М., Вагнер М., Тейхманн Д., Золгадр Б., Фолеа М., Букема Э.Дж., Дриссен А.Дж., Шлепер С., Альберс С.В. Индуцируемая УФ-излучением клеточная агрегация гипертермофильных архей Sulfolobus solfataricus опосредована образованием пилей. Мол Микробиол. Ноябрь 2008 г.;70(4):938-52. doi: 10.1111/j.1365-2958.2008.06459.x. ПМИД 18990182
- ^ Перейти обратно: а б с Аджон М., Фролс С., ван Вулферен М., Стокер К., Тейхманн Д., Дриссен А.Дж., Гроган Д.В., Альберс С.В., Шлепер С. Обмен ДНК, индуцируемый УФ-излучением, у гипертермофильных архей, опосредованный пилями IV типа. Мол Микробиол. Ноябрь 2011 г.;82(4):807-17. doi: 10.1111/j.1365-2958.2011.07861.x. Epub 2011, 18 октября. PMID 21999488.
- ^ Фрёльс С., Уайт М.Ф., Шлепер К. Реакция на повреждение УФ-излучением у модельного архея Sulfolobus solfataricus. Biochem Soc Trans. 2009 февраль;37(Часть 1):36-41. дои: 10.1042/BST0370036. ПМИД 19143598
- ^ Фурман Дж.А., МакКаллум К., Дэвис А.А. (1992). «Новая основная группа архебактерий морского планктона». Природа . 356 (6365): 148–9. Бибкод : 1992Natur.356..148F . дои : 10.1038/356148a0 . ПМИД 1545865 . S2CID 4342208 .
- ^ Делонг Э.Ф. (1992). «Археи в прибрежных морских средах» . Proc Natl Acad Sci США . 89 (12): 5685–9. Бибкод : 1992PNAS...89.5685D . дои : 10.1073/pnas.89.12.5685 . ПМК 49357 . ПМИД 1608980 .
- ^ Барнс С.М., Делвич С.Ф., Палмер Дж.Д., Пейс Н.Р. (1996). «Перспективы разнообразия архей, термофилии и монофилии на основе последовательностей рРНК окружающей среды» . Proc Natl Acad Sci США . 93 (17): 9188–93. Бибкод : 1996PNAS...93.9188B . дои : 10.1073/pnas.93.17.9188 . ПМК 38617 . ПМИД 8799176 .
- ^ Кеннеке М., Бернхард А.Е., де ла Торре-младший, Уокер С.Б., Уотербери Дж.Б., Шталь Д.А. (2005). «Выделение автотрофного морского архея, окисляющего аммиак». Природа . 437 (7058): 543–6. Бибкод : 2005Natur.437..543K . дои : 10.1038/nature03911 . ПМИД 16177789 . S2CID 4340386 .
- ^ Ютин, Наталья; Макарова Кира С.; Мехедов Сергей Л.; Вольф, Юрий И.; Кунин, Евгений В. (2008). «Глубокие архейные корни эукариотов» . Молекулярная биология и эволюция . 25 (8): 1619–1630. дои : 10.1093/molbev/msn108 . ПМЦ 2464739 . ПМИД 18463089 .
Дальнейшее чтение
[ редактировать ]Научные журналы
[ редактировать ]- Кавальер-Смит, Т. (2002). «Неомуранское происхождение архебактерий, негибактериальный корень универсального дерева и бактериальная мегаклассификация» . Межд. Дж. Сист. Эвол. Микробиол . 52 (Часть 1): 7–76. дои : 10.1099/00207713-52-1-7 . ПМИД 11837318 .
- Штакебрандт, Э; Фредериксен В; Гаррити ГМ; Гримонт, Пенсильвания; Кампфер П; Девичья МС; Несме Х; Росселло-Мора Р; Качели J; Трупер Х.Г.; Вотерен Л; Уорд переменного тока; Уитмен ВБ (2002). «Отчет специального комитета по переоценке определения видов в бактериологии». Межд. Дж. Сист. Эвол. Микробиол . 52 (Часть 3): 1043–1047. дои : 10.1099/ijs.0.02360-0 . ПМИД 12054223 .
- Гуртлер, В; Мэйолл, Британская Колумбия (2001). «Геномные подходы к типированию, таксономии и эволюции бактериальных изолятов». Межд. Дж. Сист. Эвол. Микробиол . 51 (Часть 1): 3–16. дои : 10.1099/00207713-51-1-3 . ПМИД 11211268 .
- Далеви, Д; Гугенгольц П; Блэколл Л.Л. (2001). «Подход с использованием множественных внешних групп к разрешению филогенетических отношений на уровне деления с использованием данных 16S рДНК» . Межд. Дж. Сист. Эвол. Микробиол . 51 (Часть 2): 385–391. дои : 10.1099/00207713-51-2-385 . ПМИД 11321083 .
- Кесвани, Дж; Уитмен ВБ (2001). «Связь сходства последовательности 16S рРНК с гибридизацией ДНК у прокариот» . Межд. Дж. Сист. Эвол. Микробиол . 51 (Часть 2): 667–678. дои : 10.1099/00207713-51-2-667 . ПМИД 11321113 .
- Янг, Дж. М. (2001). «Значения альтернативных классификаций и горизонтального переноса генов для бактериальной таксономии». Межд. Дж. Сист. Эвол. Микробиол . 51 (Часть 3): 945–953. дои : 10.1099/00207713-51-3-945 . ПМИД 11411719 .
- Кристенсен, Х; Бисгаард М; Фредериксен В; Бормочет Р.; Кунерт П; Олсен Дж. Э. (2001). «Достаточно ли характеристики одного изолята для действительной публикации нового рода или вида? Предложение об изменении рекомендации 30b Бактериологического кодекса (редакция 1990 г.)» . Межд. Дж. Сист. Эвол. Микробиол . 51 (Часть 6): 2221–2225. дои : 10.1099/00207713-51-6-2221 . ПМИД 11760965 .
- Кристенсен, Х; Анген О; Бормочет Р.; Олсен Дж.Э.; Бисгаард М (2000). «Гибридизация ДНК-ДНК, определенная в микролунках с использованием ковалентного присоединения ДНК» . Межд. Дж. Сист. Эвол. Микробиол . 50 (3): 1095–1102. дои : 10.1099/00207713-50-3-1095 . ПМИД 10843050 .
- Сюй, ХХ; Кавамура Ю; Ли Н; Чжао Л; Ли ТМ; Ли ЗЫ; Шу С; Эзаки Т (2000). «Экспресс-метод определения содержания G+C в бактериальных хромосомах путем мониторинга интенсивности флуоресценции при денатурации ДНК в капиллярной трубке» . Межд. Дж. Сист. Эвол. Микробиол . 50 (4): 1463–1469. дои : 10.1099/00207713-50-4-1463 . ПМИД 10939651 .
- Янг, Дж. М. (2000). «Предложения по избежанию продолжающейся путаницы с Бактериологическим кодексом» . Межд. Дж. Сист. Эвол. Микробиол . 50 (4): 1687–1689. дои : 10.1099/00207713-50-4-1687 . ПМИД 10939677 .
- Хансманн, С; Мартин В. (2000). «Филогения 33 рибосомальных и шести других белков, закодированных в древнем кластере генов, который консервативен во всех геномах прокариот: влияние исключения из анализа плохо выравниваемых участков» . Межд. Дж. Сист. Эвол. Микробиол . 50 (4): 1655–1663. дои : 10.1099/00207713-50-4-1655 . ПМИД 10939673 .
- Тиндалл, Би Джей (1999). «Предложение об изменении Правил обозначения типовых штаммов, депонированных под номерами коллекций культур, выделенных для патентных целей» . Межд. Дж. Сист. Бактериол . 49 (3): 1317–1319. дои : 10.1099/00207713-49-3-1317 . ПМИД 10490293 .
- Тиндалл, Би Джей (1999). «Предложение изменить Правило 18a, Правило 18f и Правило 30, чтобы ограничить обратную силу изменений, принятых ICSB» . Межд. Дж. Сист. Бактериол . 49 (3): 1321–1322. дои : 10.1099/00207713-49-3-1321 . ПМИД 10425797 .
- Тиндалл, Би Джей (1999). «Непонимание бактериологического кода» . Межд. Дж. Сист. Бактериол . 49 (3): 1313–1316. дои : 10.1099/00207713-49-3-1313 . ПМИД 10425796 .
- Тиндалл, Би Джей (1999). «Предложения по актуализации и внесению изменений в Бактериологический кодекс» . Межд. Дж. Сист. Бактериол . 49 (3): 1309–1312. дои : 10.1099/00207713-49-3-1309 . ПМИД 10425795 .
- Бургграф, С; Хубер Х; Стеттер КО (1997). «Реклассификация порядков и семейств кренархалей в соответствии с данными последовательности 16S рРНК» . Межд. Дж. Сист. Бактериол . 47 (3): 657–660. дои : 10.1099/00207713-47-3-657 . ПМИД 9226896 .
- Палис, Т; Накамура Л.К.; Коэн FM (1997). «Открытие и классификация экологического разнообразия в мире бактерий: роль данных о последовательностях ДНК» . Межд. Дж. Сист. Бактериол . 47 (4): 1145–1156. дои : 10.1099/00207713-47-4-1145 . ПМИД 9336922 .
- Юзеби, JP (1997). «Список названий бактерий, имеющих номенклатурное положение: папка доступна в Интернете» . Межд. Дж. Сист. Бактериол . 47 (2): 590–592. дои : 10.1099/00207713-47-2-590 . ПМИД 9103655 .
- Клейтон, РА; Саттон Дж; Хинкль П.С. младший; Балка С; Филдс С (1995). «Внутривидовая изменчивость последовательностей малых субъединиц рРНК в GenBank: почему отдельные последовательности не могут адекватно представлять таксоны прокариот» . Межд. Дж. Сист. Бактериол . 45 (3): 595–599. дои : 10.1099/00207713-45-3-595 . ПМИД 8590690 .
- Мюррей, Р.Г.; Шлейфер К.Х. (1994). «Таксономические заметки: предложение по регистрации свойств предполагаемых таксонов прокариотов» . Межд. Дж. Сист. Бактериол . 44 (1): 174–176. дои : 10.1099/00207713-44-1-174 . ПМИД 8123559 .
- Винкер, С; Вёзе ЧР (1991). «Определение доменов Archaea, Bacteria и Eucarya с точки зрения характеристик малых субъединиц рибосомальной РНК». Сист. Прил. Микробиол . 14 (4): 305–310. дои : 10.1016/s0723-2020(11)80303-6 . ПМИД 11540071 .
- Вёзе, ЧР; Кандлер О; Уилис М.Л. (1990). «На пути к естественной системе организмов: предложение по доменам архей, бактерий и эукариев» . Учеб. Натл. акад. наук. США . 87 (12): 4576–4579. Бибкод : 1990PNAS...87.4576W . дои : 10.1073/pnas.87.12.4576 . ПМК 54159 . ПМИД 2112744 .
- Ахенбах-Рихтер, Л; Вёзе ЧР (1988). «Спейсерная область рибосомального гена у архебактерий». Сист. Прил. Микробиол . 10 (3): 211–214. дои : 10.1016/s0723-2020(88)80002-x . ПМИД 11542149 .
- МакГилл, Ти Джей; Юрка Дж; Собеский Дж.М.; Пикетт М.Х.; Вёзе ЧР; Фокс Дж.Э. (1986). «Характерные архебактериальные олигонуклеотиды 16S рРНК». Сист. Прил. Микробиол . 7 (2–3): 194–197. дои : 10.1016/S0723-2020(86)80005-4 . ПМИД 11542064 .
- Вёзе, ЧР; Гупта Р; Хан СМ; Зиллиг В; Ту Дж (1984). «Филогенетические взаимоотношения трех серозависимых архебактерий». Сист. Прил. Микробиол . 5 : 97–105. дои : 10.1016/S0723-2020(84)80054-5 . ПМИД 11541975 .
- Вёзе, ЧР; Олсен Дж.Дж. (1984). «Филогенетические взаимоотношения трех серозависимых архебактерий». Сист. Прил. Микробиол . 5 : 97–105. дои : 10.1016/S0723-2020(84)80054-5 . ПМИД 11541975 .
- Вёзе, ЧР; Фокс Дж.Э. (1977). «Филогенетическая структура прокариотического домена: первичные царства» . Учеб. Натл. акад. наук. США . 74 (11): 5088–5090. Бибкод : 1977PNAS...74.5088W . дои : 10.1073/pnas.74.11.5088 . ПМК 432104 . ПМИД 270744 .
Научные справочники
[ редактировать ]- Гэррити, Холт Дж.Г. (2001). «Тип AI. Crenarchaeota phy. nov.» . В Д. Р. Буне, Р. В. Кастенхольце (ред.). Руководство Берджи по систематической бактериологии, том 1: Археи и глубоко ветвящиеся и фототрофные бактерии (2-е изд.). Нью-Йорк: Springer Verlag. стр. 169 . ISBN 978-0-387-98771-2 .
Внешние ссылки
[ редактировать ]- «Кренархеота» . Виртуальная микробиология (bact.wisc.edu) . Университет Висконсина .
- «Сравнительный анализ геномов кренархей» . Интегрированная система микробных геномов . Министерство энергетики США .