Dromaeosauridae
| Dromaeosaurids Временный диапазон:
| |
|---|---|

| |
| Коллекция Dromaeosaurid Fossil Skeletons. По часовой стрелке из верхнего левого: Deinonychus antirrhopus (сильно построенный эудромаозавр), Buitereraptor gonzalezorum (длинный неэенлагин), велоцираптор монголиенсис (маленький велоцираптор), микрораптор Gui (а-кггеночный микрораптор), halszraptor), гуи (айгеновый микрорапурный halszkaraptorine), Zhenyuanlong Suni (давно крылатый Dromaeosaurid) | |
| Scientific classification | |
| Domain: | Eukaryota |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Clade: | Dinosauria |
| Clade: | Saurischia |
| Clade: | Theropoda |
| Clade: | Paraves |
| Family: | †Dromaeosauridae Matthew & Brown, 1922 |
| Type species | |
| †Dromaeosaurus albertensis Matthew & Brown, 1922
| |
| Subgroups[5] | |
| |
| Synonyms | |
| |
Dromaeosauridae ( / ˌ d r ɒ m i . Как правило, они были малыми до пернатых хищников , которые процветали в меловой периоде . Название Dromaeosauridae означает «бегущие ящерицы», от греческого Δρομαῖος ( dromaîos ), что означает «бег на полной скорости», «Swift» и σαῦρος ( saûros ), что означает «ящерица». При неформальном использовании их часто называют хищниками [ 6 ] (после Velociraptor ), термин, популяризированный Парком юрского периода ; Несколько родов включают термин «Raptor» непосредственно в их имени, и популярная культура пришла к тому, чтобы подчеркнуть их похожий на птицу внешний вид и предполагаемое птичное поведение.
Окаменелости Dromaeosaurid были найдены по всему миру в Северной Америке , Европе , Африке , Азии и Южной Америке , причем некоторые окаменелости подтверждают возможность того, что они населяли Австралию . также [ 7 ] Самые ранние окаменелости известны из раннего мела (145-140 миллионов лет назад), и они выжили до конца меловой ( маастрихтская стадия, 66 млн. Лет), существовав до мелинового палеогенового вымирания . Присутствие дромазавридов уже в среднем юрском периоде было предложено в результате открытия изолированных ископаемых зубов, хотя в этом периоде не было обнаружено, что ископаемые тела дромазавридов не было обнаружено. [8][9]
Description
[edit]Technical diagnosis
[edit]Dromaeosaurids are diagnosed by the following features: short T-shaped frontals that form the rostral boundary of the supratemporal fenestra; a caudolateral overhanging shelf of the squamosal; a lateral process of the quadrate that contacts the quadratojugal; raised, stalked, parapophyses on the dorsal vertebrae, a modified pedal digit II; chevrons and prezygapophysis of the caudal vertebrae elongate and spanning several vertebrae; the presence of a subglenoid fossa on the coracoid.[10]
Size and general build
[edit]

Dromaeosaurids were small to medium-sized dinosaurs, ranging from 1.5–2.07 metres (4.9–6.8 ft) in length (in the case of Velociraptor) to approaching or over 6 m (20 ft) (in Utahraptor, Dakotaraptor and Achillobator).[11][12] Large size appears to have evolved at least twice among dromaeosaurids; once among the dromaeosaurines Utahraptor and Achillobator, and again among the unenlagiines (Austroraptor, which measured 5–6 m (16–20 ft) long). A possible third lineage of giant dromaeosaurids is represented by isolated teeth found on the Isle of Wight, England. The teeth belong to an animal the size of the dromaeosaurine Utahraptor, but they appear to belong to velociraptorines, judging by the shape of the teeth.[13][14]
The distinctive dromaeosaurid body plan helped to rekindle theories that dinosaurs may have been active, fast, and closely related to birds. Robert Bakker's illustration for John Ostrom's 1969 monograph,[15] showing the dromaeosaurid Deinonychus in a fast run, is among the most influential paleontological reconstructions in history.[16] The dromaeosaurid body plan includes a relatively large skull, serrated teeth, narrow snout (an exception being the derived dromaeosaurines), and forward-facing eyes which indicate some degree of binocular vision.[17]

Dromaeosaurids, like most other theropods, had a moderately long S-curved neck, and their trunk was relatively short and deep. Like other maniraptorans, they had long arms that could be folded against the body in some species, and relatively large hands with three long fingers (the middle finger being the longest and the first finger being the shortest) ending in large claws.[10] The dromaeosaurid hip structure featured a characteristically large pubic boot projecting beneath the base of the tail. Dromaeosaurid feet bore a large, recurved claw on the second toe. Their tails were slender, with long, low, vertebrae lacking transverse process and neural spines after the 14th caudal vertebra.[10] Ossified uncinate processes of ribs have been identified in several dromaeosaurids.[18][19][20]
Foot
[edit]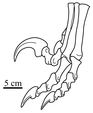
Like other theropods, dromaeosaurids were bipedal; that is, they walked on their hind legs. However, whereas most theropods walked with three toes contacting the ground, fossilized footprint tracks confirm that many early paravian groups, including the dromaeosaurids, held the second toe off the ground in a hyperextended position, with only the third and fourth toes bearing the weight of the animal. This is called functional didactyly.[21] The enlarged second toe bore an unusually large, curved, falciform (sickle-shaped, alt. drepanoid) claw (held off the ground or 'retracted' when walking), which is thought to have been used in capturing prey and climbing trees (see "Claw function" below). This claw was especially blade-like in the large-bodied predatory eudromaeosaurs.[22] One possible dromaeosaurid species, Balaur bondoc, also possessed a first toe which was highly modified in parallel with the second. Both the first and second toes on each foot of B. bondoc were also held retracted and bore enlarged, sickle-shaped claws.[23]
Tail
[edit]
Dromaeosaurids had long tails. Most of the tail vertebrae bore bony, rod-like extensions (called prezygapophyses), as well as bony tendons in some species. In his study of Deinonychus, Ostrom proposed that these features stiffened the tail so that it could only flex at the base, and the whole tail would then move as a single, rigid, lever.[15] However, one well-preserved specimen of Velociraptor mongoliensis (IGM 100/986) has an articulated tail skeleton that is curved horizontally in a long S-shape. This suggests that, in life, the tail could bend from side to side with a substantial degree of flexibility.[24] It has been proposed that this tail was used as a stabilizer or counterweight while running or in the air;[24] in Microraptor, an elongate diamond-shaped fan of feathers is preserved on the end of the tail. This may have been used as an aerodynamic stabilizer and rudder during gliding or powered flight (see "Flight and gliding" below).[25]
Feathers
[edit]

There is a large body of evidence showing that dromaeosaurids were covered in feathers. Some dromaeosaurid fossils preserve long, pennaceous feathers on the hands and arms (remiges) and tail (rectrices), as well as shorter, down-like feathers covering the body.[26][27] Other fossils, which do not preserve actual impressions of feathers, still preserve the associated bumps on the forearm bones where long wing feathers would have attached in life.[28] Overall, this feather pattern looks very much like Archaeopteryx.[26]
The first known dromaeosaurid with definitive evidence of feathers was Sinornithosaurus, reported from China by Xu et al. in 1999.[27] Many other dromaeosaurid fossils have been found with feathers covering their bodies, some with fully developed feathered wings. Microraptor even shows evidence of a second pair of wings on the hind legs.[26] While direct feather impressions are only possible in fine-grained sediments, some fossils found in coarser rocks show evidence of feathers by the presence of quill knobs, the attachment points for wing feathers possessed by some birds. The dromaeosaurids Rahonavis and Velociraptor have both been found with quill knobs, showing that these forms had feathers despite no impressions having been found. In light of this, it is most likely that even the larger ground-dwelling dromaeosaurids bore feathers, since even flightless birds today retain most of their plumage, and relatively large dromaeosaurids, like Velociraptor, are known to have retained pennaceous feathers.[28][29] Though some scientists had suggested that the larger dromaeosaurids lost some or all of their insulatory covering, the discovery of feathers in Velociraptor specimens has been cited as evidence that all members of the family retained feathers.[28][30]
More recently, the discovery of Zhenyuanlong established the presence of a full feathered coat in relatively large dromaeosaurids. Additionally, the animal displays proportionally large, aerodynamic wing feathers, as well as a tail-spanning fan, both of which are unexpected traits that may offer an understanding of the integument of large dromaeosaurids.[31] Dakotaraptor is an even larger dromaeosaurid species with evidence of feathers, albeit indirect in the form of quill knobs,[32] though the taxon is considered as chimeara by other researchers as even the dinosaurian elements with supposed traits diagnostic for dromaeosaurs also referrable to caenagnathids and ornithomimosaurians.[33][34]
Classification
[edit]Relationship with birds
[edit]
Dromaeosaurids share many features with early birds (clade Avialae or Aves). The precise nature of their relationship to birds has undergone a great deal of study, and hypotheses about that relationship have changed as large amounts of new evidence became available. As late as 2001, Mark Norell and colleagues analyzed a large survey of coelurosaur fossils and produced the tentative result that dromaeosaurids were most closely related to birds, with troodontids as a more distant outgroup. They even suggested that Dromaeosauridae could be paraphyletic relative to Avialae.[35] In 2002, Hwang and colleagues utilized the work of Norell et al., including new characters and better fossil evidence, to determine that birds (avialans) were better thought of as cousins to the dromaeosaurids and troodontids.[11] The consensus of paleontologists is that there is not yet enough evidence to determine whether any dromaeosaurids could fly or glide, or whether they evolved from ancestors that could.[36]
Alternative theories and flightlessness
[edit]Dromaeosaurids are so bird-like that they have led some researchers to argue that they would be better classified as birds. First, since they had feathers, dromaeosaurids (along with many other coelurosaurian theropod dinosaurs) are "birds" under traditional definitions of the word "bird", or "Aves", that are based on the possession of feathers. However, other scientists, such as Lawrence Witmer, have argued that calling a theropod like Caudipteryx a bird because it has feathers may stretch the word past any useful meaning.[37]

At least two schools of researchers have proposed that dromaeosaurids may actually be descended from flying ancestors. Hypotheses involving a flying ancestor for dromaeosaurids are sometimes called "Birds Came First" (BCF). George Olshevsky is usually credited as the first author of BCF.[38] In his own work, Gregory S. Paul pointed out numerous features of the dromaeosaurid skeleton that he interpreted as evidence that the entire group had evolved from flying, dinosaurian ancestors, perhaps an animal like Archaeopteryx. In that case, the larger dromaeosaurids were secondarily flightless, like the modern ostrich.[29] In 1988, Paul suggested that dromaeosaurids may actually be more closely related to modern birds than to Archaeopteryx. By 2002, however, Paul placed dromaeosaurids and Archaeopteryx as the closest relatives to one another.[39]
In 2002, Hwang et al. found that Microraptor was the most primitive dromaeosaurid.[11] Xu and colleagues in 2003 cited the basal position of Microraptor, along with feather and wing features, as evidence that the ancestral dromaeosaurid could glide. In that case the larger dromaeosaurids would be secondarily terrestrial—having lost the ability to glide later in their evolutionary history.[26]
Also in 2002, Steven Czerkas described Cryptovolans, though it is a probable junior synonym of Microraptor. He reconstructed the fossil inaccurately with only two wings and thus argued that dromaeosaurids were powered fliers, rather than passive gliders. He later issued a revised reconstruction in agreement with that of Microraptor[40]
Other researchers, like Larry Martin, have proposed that dromaeosaurids, along with all maniraptorans, were not dinosaurs at all. Martin asserted for decades that birds were unrelated to maniraptorans, but in 2004 he changed his position, agreeing that the two were close relatives. However, Martin believed that maniraptorans were secondarily flightless birds, and that birds did not evolve from dinosaurs, but rather from non-dinosaurian archosaurs.[41]

In 2005, Mayr and Peters described the anatomy of a very well preserved specimen of Archaeopteryx, and determined that its anatomy was more like non-avian theropods than previously understood. Specifically, they found that Archaeopteryx had a primitive palatine, unreversed hallux, and hyper-extendable second toe. Their phylogenetic analysis produced the controversial result that Confuciusornis was closer to Microraptor than to Archaeopteryx, making the Avialae a paraphyletic taxon. They also suggested that the ancestral paravian was able to fly or glide, and that the dromaeosaurids and troodontids were secondarily flightless (or had lost the ability to glide).[43][44] Corfe and Butler criticized this work on methodological grounds.[45]
A challenge to all of these alternative scenarios came when Turner and colleagues in 2007 described a new dromaeosaurid, Mahakala, which they found to be the most basal and most primitive member of the Dromaeosauridae, more primitive than Microraptor. Mahakala had short arms and no ability to glide. Turner et al. also inferred that flight evolved only in the Avialae, and these two points suggested that the ancestral dromaeosaurid could not glide or fly. Based on this cladistic analysis, Mahakala suggests that the ancestral condition for dromaeosaurids is non-volant.[46] However, in 2012, an expanded and revised study incorporating the most recent dromaeosaurid finds recovered the Archaeopteryx-like Xiaotingia as the most primitive member of the clade Dromaeosauridae, which appears to suggest the earliest members of the clade may have been capable of flight.[47]
Taxonomy
[edit]The authorship of the family Dromaeosauridae is credited to William Diller Matthew and Barnum Brown, who erected it as a subfamily (Dromaeosaurinae) of the family Deinodontidae in 1922, containing only the new genus Dromaeosaurus.[48]
The subfamilies of Dromaeosauridae frequently shift in content based on new analysis, but typically consist of the following groups. A number of dromaeosaurids have not been assigned to any particular subfamily, often because they are too poorly preserved to be placed confidently in phylogenetic analysis (see section Phylogeny below) or are indeterminate, being assigned to different groups depending on the methodology employed in different papers. The most basal known subfamily of dromaeosaurids is Halszkaraptorinae, a group of bizarre creatures with long fingers and necks, a large number of small teeth, and possible semiaquatic habits.[49] Another enigmatic group, Unenlagiinae, is the most poorly supported subfamily of dromaeosaurids and it is possible that some or all of its members belong outside of Dromaeosauridae.[50][51] The larger, ground-dwelling members like Buitreraptor and Unenlagia show strong flight adaptations, although they were probably too large to 'take off'. One possible member of this group, Rahonavis, is very small, with well-developed wings that show evidence of quill knobs (the attachment points for flight feathers) and it is very likely that it could fly. The next most primitive clade of dromaeosaurids is the Microraptoria. This group includes many of the smallest dromaeosaurids, which show adaptations for living in trees. All known dromaeosaurid skin impressions hail from this group and all show an extensive covering of feathers and well-developed wings. Like the unenlagiines, some species may have been capable of active flight. The most advanced subgroup of dromaeosaurids, Eudromaeosauria, includes stocky and short-legged genera which were likely ambush hunters. This group includes Velociraptorinae, Dromaeosaurinae, and in some studies a third group: Saurornitholestinae. The subfamily Velociraptorinae has traditionally included Velociraptor, Deinonychus, and Saurornitholestes, and while the discovery of Tsaagan lent support to this grouping, the inclusion of Deinonychus, Saurornitholestes, and a few other genera is still uncertain. The Dromaeosaurinae is usually found to consist of medium to giant-sized species, with generally box-shaped skulls (the other subfamilies generally have narrower snouts).[5]





The following classification of the various genera of dromaeosaurids follows the table provided in Holtz, 2011 unless otherwise noted.[5]
- Family Dromaeosauridae
- Nuthetes
- Pamparaptor
- Variraptor
- Pyroraptor
- Zhenyuanlong
- Daurlong
- Subfamily Halszkaraptorinae[49]
- Subfamily Unenlagiinae
- Subfamily Microraptorinae
- Node Eudromaeosauria[22]
- Deinonychus
- Dineobellator
- Vectiraptor
- Subfamily Saurornitholestinae
- Subfamily Velociraptorinae
- Subfamily Dromaeosaurinae
Phylogeny
[edit]Dromaeosauridae впервые определили как клада Полом Серено в 1998 году, как самая инклюзивная естественная группа, содержащая Dromaeosaurus , но не Troodon , Ornithomimus или прохожих . Различные «подсемейства» также были повторно определены в виде клад, обычно определяемых как все виды, ближе к тезке групп, чем к Dromaeosaurus , или любые одноименные имены других подкладов (например, Маковицки определил кладу Unenlagiinae как все Dromaeosaurids к Unenlagia. чем к велоцираптору ). Микрораптория является единственной подкладкой Dromaeosaurid, не преобразованной из подсемейства. Сентер и коллеги прямо придумали название без подсемейства суффиксов -inae , чтобы избежать предполагаемых проблем с возведением традиционного таксона семейной группы, если группа будет обнаружена за пределами Dromaeosauridae. [53] Sereno offered a revised definition of the sub-group containing Microraptor to ensure that it would fall within Dromaeosauridae, and erected the subfamily Microraptorinae, attributing it to Senter et al., though this usage has only appeared on his online TaxonSearch database and has not been formally published.[54] The extensive cladistic analysis conducted by Turner et al. (2012) further supported the monophyly of Dromaeosauridae.[55]
Кладограмма ниже следует анализу 2015 года Depalma et al. Использование обновленных данных из рабочей группы Theropod. [ 32 ]
| Dromaeosauridae | |


Другая кладограмма, построенная ниже, следует за филогенетическим анализом, проведенным в 2017 году Cau et al. Использование обновленных данных из рабочей группы Theropod в их описании Halskaraptor . [ 49 ]
| Dromaeosauridae | |



Палеобиология
[ редактировать ]Чувства
[ редактировать ]Сравнения между склеральными кольцами нескольких дромаозавридов ( микрораптор , синорнитозавр и велосираптор ) и современных птиц и рептилий указывают на то, что и велосираптор ) возможно включая , дромаозавриды некоторые ( микрораптор короткие интервалы). [ 56 ] Тем не менее, открытие переливающегося оперения в микрорапторе ставит под сомнение вывод о ночной жизни в этом роде, поскольку ни один современные птицы, обладающие радужными оперениями, не известны ночными. [ 57 ]
Исследования обонятельных луковиц дромозавридов показывают, что у них были сходные обонятельные соотношения для их размера к другим неавийским тероподам и современным птицам с острым обонянием, таким как тиранозавриды и стервятник индейки , вероятно, отражая важность обонятельного смысла в Ежедневная деятельность Dromaeosaurids, таких как поиск пищи. [ 58 ] [ 59 ]
Кормление
[ редактировать ]Было обнаружено, что кормление дромазауридов типично для теропод -колурозавров, с характерным методом питания «прокола и тяги». Исследования схем износа на зубах дромаозавридов Angelica Titices et al. Укажите, что зубы Dromaeosaurid имеют сходные схемы износа с теми, которые видели в Tyrannosauridae и Troodontidae. Тем не менее, Microwear на зубах указывал на то, что Dromaeosaurids, вероятно, предпочитали большие добычи, чем Troodontids, с которыми они часто разделяли окружающую среду. Такие диетические дифференциации, вероятно, позволили им населять одну и ту же окружающую среду. То же самое исследование также показало, что Dromaeosaurids, такие как Dromaeosaurus и SauroRnitholestes (два дромаозаврида, проанализированные в исследовании), вероятно, включали кость в свой рацион и были лучше адаптированы для борьбы с добычей в то время как та такие как беспозвоночные и каррион. [ 60 ]
Функция когтя
[ редактировать ]

В настоящее время существуют разногласия по поводу функции увеличенного «серпа» на втором ноге. Когда Джон Остром описал это для Deinonychus в 1969 году, он интерпретировал коготь как лезвие, похожее на срезанное оружие, так же, как клыки некоторых саб-зубчатых кошек , используемых с мощными ударами, чтобы врезать добычу. Адамс (1987) предположил, что Talon использовался для выпадения крупных цератопсских динозавров. [ 61 ] Интерпретация серпового когти как убийственного оружия, применяемого ко всем дромаэзавридам. Однако Manning et al. Утверждал, что вместо этого Коготь служил крючком, реконструируя кератиновую оболочку с помощью эллиптического поперечного сечения, вместо ранее предполагаемой перевернутой формы слезы. [ 62 ] В интерпретации Мэннинга второй когти на ногах будет использоваться в качестве помощи скалолазанию, когда подчиняет большую добычу, а также в качестве нанесенного нанесению ударного оружия.
Остром сравнил Deinonychus с страусом и кассовари . Он отметил, что виды птиц могут нанести серьезные травмы большим когтям на втором ноге. [ 15 ] Кассоварий имеет когти длиной до 125 миллиметров (4,9 дюйма). [ 63 ] Остром процитировал Гиллиарда (1958), сказав, что они могут разорвать руку или выбросить человека. [ 64 ] Кофрон (1999 и 2003 год) изучил 241 задокументированные атаки кассовой, и обнаружил, что одна человеческая и две собаки были убиты, но никаких доказательств того, что кассовари могут выбросить или расчленять других животных. [ 65 ] [ 66 ] Кассовы используют свои когти, чтобы защитить себя, атаковать угрожающих животных, и на агонистических проявлениях, таких как выставка для лука. [ 63 ] Seriema также имеет увеличенный когти на втором ноге и использует его , чтобы разорвать небольшие предметы добычи для глотания. [ 67 ]
Филипп Мэннинг и коллеги (2009) пытались проверить функцию серпового когти и когти с аналогичной формой на передних конечностях. Они проанализировали биомеханику того, как стрессы и штаммы будут распределяться вдоль когтей и в конечностях, используя рентгеновские визуализации для создания трехмерной контурной карты когти переднего когтя из велосираптора . Для сравнения, они проанализировали построение когти от современной хищной птицы, орлиной совы . Они обнаружили, что, основываясь на том, как стресс проводился вдоль коготь, они были идеальными для лазания. Ученые обнаружили, что заточенным кончиком когти был прокол и захватывающий инструмент, в то время как изогнутая и расширенная база когти помогла равномерно перенести напряженные нагрузки. Команда Мэннинга также сравнила кривизну Dromaeosaurid "Spid Claw" на ноге с кривизны у современных птиц и млекопитающих. Предыдущие исследования показали, что количество кривизны в когтях соответствовало тому, какой образ жизни у животного есть: животные с сильно изогнутыми когтями определенной формы, как правило, являются альпинистами, в то время как прямые когти указывают на стиль жизни на земле. Серп когти Deinonychus имеет кривизну 160 градусов, в значительной степени в пределах диапазона лазании животных. Когти, которые они изучали передние конечности, также попали в диапазон кривизны. [ 68 ]
Палеонтолог Питер МакКавицки прокомментировал исследование команды Мэннинга, заявив, что маленькие примитивные дромаозавриды (такие как микрораптор вероятно, были деревьями, но этот лазание не объясняло, почему позже гигантские дромаозавриды, такие как ахиллобатор ) , были слишком большими, чтобы подняться на деревья. Маковицки предположил, что гигантские дромаозавриды, возможно, адаптировали когти, который будет использоваться исключительно для привлечения добычи. [ 69 ]

В 2009 году Фил Сендер опубликовал исследование по пальцам ног Дромаозаврида и показал, что их диапазон движения был совместим с раскопками жестких гнезд насекомых. Сентер предположил, что небольшие дромаозавриды, такие как Рахонавис и Бютрераптор, были достаточно маленькими, чтобы быть частичными насекомыми , в то время как более крупные роды, такие как Deinonychus и Neuquenraptor, могли бы использовать эту способность ловить добычу позвоночных в гнездах насекомых. Тем не менее, Senter не проверял, была ли также способствовала сильная кривизна когти Dromaeosaurid. [ 70 ]
В 2011 году Денвер Фаулер и его коллеги предложили новый метод, с помощью которого Dromaeosaurids могли получить меньшую добычу. Эта модель, известная как модель хищничества «сдержанность добычи хищника» (RPR), предлагает, чтобы дромаэосавриды убили свою добычу, очень похожие на существующих добычи с добычей : подпрыгивая на их карьер, закрепляя ее под их телом, и и крепко схватив его большими когтями. Как и Accipitrids, Dromaeosaurid тогда начал бы питаться животным, пока он еще жив, пока не умер от кровопотери и недостаточности органа. Это предложение основано главным образом на сравнении морфологии и пропорциях ног и ног дромаозавридов с несколькими группами добычи хищных птиц с известным хищным поведением. Фаулер обнаружил, что ноги и ноги дромазавридов наиболее похожи на ноги орлов и ястребов , особенно с точки зрения увеличения второго когтя и аналогичного диапазона захватывающих движений. короткая плюсневая плюс Однако Совы . Метод хищничества RPR будет соответствовать другим аспектам анатомии Dromaeosaurid, такими как их необычная морфология зубов и рука. Руки, которые могли оказывать много силы, но, вероятно, были покрыты длинными перьями, могли использоваться в качестве стабилизаторов хлопья для баланса, в то же время на вершине борющегося добычи, наряду с жестким уравновешенным хвостом. Jaws Dromaeosaurid, которые Фаулер и его коллеги считали сравнительно слабыми, были бы полезны для употребления добычи живой, но не так полезно для быстрого и сильного отправки добычи. Эти хищные адаптации, работающие вместе, также могут иметь последствия для происхождения хлопья у паравицев . [ 71 ] [ 72 ]

В 2019 году Питер Бишоп реконструировал скелет ног и мускулатуру Deinonychus , используя трехмерные модели мышц , сухожилий и костей . С добавлением математических моделей и уравнений епископ смоделировал условия, которые обеспечат максимальную силу на кончике серпового когти и, следовательно, наиболее вероятной функции. Среди предложенных способов использования серповидного когтя: удары по вырезанию, ударе или добыче высадки; за то, чтобы схватить на боки добычи; пронзительный помогает вес тела; атаковать жизненно важные области добычи; чтобы сдержать добычу; внутри - или межвидовая конкуренция ; и выкапывать добычу от укрытий. Результаты, полученные епископом, показали, что приседающая осанка увеличивала силы когтей, однако эти силы оставались относительно слабыми, что указывает на то, что когти не были достаточно сильными, чтобы их можно было использовать в ударах. Вместо того, чтобы использовать для сокращения, серповые когти были более вероятными при сгибающих углах ноги, таких как сдерживание добычи и нанесение удара добычи в близких кварталах. Эти результаты согласуются с Образец сражающих динозавров , который сохраняет велосираптор и протокератопы, запертые в бою, причем первые сжимались на другого с его когтями в неэтагированной осанке ноги. Несмотря на полученные результаты, епископ считал, что возможности серпового когти могли бы варьироваться в пределах таксонов, учитывая, что среди дромаозавридов у Адасаура была необычайно меньшая серповидная когтя, который сохранил характерную гиллинсмоидную предпоследний фаланги. Он не мог ни подтвердить, ни игнорировать, что педалиная цифра II может иметь потерю или сохранить ее функционально. [ 73 ]
Исследование 2020 года, проведенное Gianechini et al . Также показывает, что велоцирапторины, дромозавра и другие эвромаозавры в Лорасии сильно различались в своих локомотивах и техниках убийства от неэнлагиновых дромозавридов Гндваны. Более короткая вторая фаланга во второй цифре стопы, позволяющая увеличить силу, создаваемую этой цифрой, которая в сочетании с более короткой и более широкой плюсневой кости и заметной заметной шарнизной морфологией суставных поверхностей межевины и фалангов, возможно, возможно, возможно, возможно, возможно, возможно позволил евромаэозаврам оказывать большую захватывающую силу, чем безэнлагины, что позволяет более эффективно подчинять и убивать крупных добыча. Для сравнения, у неэнлагиновых дромозавридов были более длинные и стройные субаркктометатарса и менее хорошо перемешанные шарнирные шарнирные суставы, что, возможно, дала им большую качественную способность и допускает большую скорость. Кроме того, более длинная вторая фаланга второй цифры позволила бы быстрым движениям второго цифр их ног, чтобы охотиться на более мелкие и более неуловимых видов добычи. Эти различия в локомоторных и хищных специализациях, возможно, были ключевой особенностью, которая повлияла на эволюционные пути, которые сформировали обе группы дромаозавров в северном и южном полушарии. [ 74 ]
Групповое поведение
[ редактировать ]
Окаменелости Deinonychus были обнаружены в небольших группах возле останков травоядного Tenontosaurus , более крупного орнитишского динозавра. Это было истолковано как доказательство того, что эти дромаозавриды охотились в скоординированных упаковках, таких как некоторые современные млекопитающие . [ 76 ] Однако не все палеонтологи посчитали доказательства убедительными, и последующее исследование, опубликованное в 2007 году Роуком и Бринкманом, предполагает, что Deinonychus , возможно, фактически демонстрировал неорганизованное поведение моббинга. Современные диапсиды , в том числе птицы и крокодилы (ближайшие родственники Dromaeosaurids), демонстрируют минимальную долгосрочную кооперативную охоту (кроме Aplomado Falcon и Harris's Hawk ); Вместо этого они обычно являются одиночными охотниками, либо объединяют силы, время от времени, чтобы увеличить успех в охоте (как иногда делают крокодилы), либо притягиваются к ранее убитым тушам, где конфликт часто встречается между людьми одного и того же вида. Например, в ситуациях, когда группы драконов Комодо едят вместе, самые большие люди едят первыми и могут атаковать меньшие драконы Комодо, которые пытаются кормить; Если меньшее животное умирает, это обычно каннибализируется . Когда эта информация применяется к сайтам, содержащим предполагаемое поведение на охоте на упаковку у Dromaeosaurids, она кажется несколько соответствующей стратегии кормления, похожей на дракон, похожая на дракон. Остатки скелета Deinonychus , обнаруженные на этих участках, взяты из Subadults, с недостающими частями, которые могли быть съедены другие Deinonychus , что исследование Roach et al. Представлено в качестве доказательства против идеи, что животные сотрудничали в охоте. [ 77 ] Различные диетические предпочтения между ювенильным и взрослым Deinonychus, опубликованным в 2020 году, указывают на то, что у животного не было сложного кооперативного поведения, наблюдаемого у животных, охотящихся за упаковкой. Продолжается ли это на другие дромаозавры, в настоящее время неизвестно. [ 78 ] Третий возможный вариант состоит в том, что у Dromaeosaurids не было долгосрочного кооперативного поведения, но демонстрировали кратковременное кооперативное поведение, как видно у крокодилов, которые демонстрируют как истинное сотрудничество, так и конкуренция за добычу.
В 2001 году несколько образцов в Ютахрапторе в возрасте от полностью взрослого до крошечного ребенка длиной три фута были обнаружены на месте, рассматриваемом некоторыми, как ловушка для зыбки и хищника. Некоторые считают это доказательством семейного охотничьего поведения; Тем не менее, полный блок песчаника еще предстоит открыть, и исследователи не уверены, умерли ли животные одновременно. [ 79 ]
В 2007 году ученые описали первый известный обширный трассу Dromaeosaurid , в Шаньдун, Китай. В дополнение к подтверждению гипотезы о том, что серп -когти был удержан с земли, трасса (сделанная крупными видами, размером с ахиллобатор ), показали доказательства того, что шесть особей примерно равной размер перемещаются вместе вдоль береговой линии. Люди были расположены на расстоянии около одного метра, путешествуя в том же направлении и ходили в довольно медленном темпе. Авторы статьи, описывающие эти следы, интерпретировали дорожки как доказательство того, что некоторые виды дромазавридов жили в группах. Хотя Trackways явно не представляют охотничьего поведения, по словам авторов, идея о том, что группы Dromaeosaurids, возможно, охотились вместе. [ 21 ]
Летать и скользить
[ редактировать ]
Предплечья дромозавридов, по -видимому, хорошо адаптированы для сопротивления крутям и изгибающим напряжениям, связанным с хлопьями и скольжением, [ 80 ] и способность летать или скольжение была предложена как минимум для пяти видов Dromaeosaurid. Первый, Rahonavis ostromi (первоначально классифицированный как птичья птица, но обнаружил, что это Dromaeosaurid в последующих исследованиях [ 17 ] [ 81 ] ), возможно, был способен к питанию , о чем свидетельствует его длинные передние конечности с доказательством привязанности к перо для длинных крепких перьев полета. [ 82 ] Передатки Рахонависа были построены более мощными, чем Archeopteryx , и показывают доказательства того, что они имели сильные привязанности к связке, необходимые для хлопья. Луис Чиаппе пришел к выводу, что, учитывая эти адаптации, Рахонавис, вероятно, мог бы летать, но был бы более неуклюжим в воздухе, чем современные птицы. [ 83 ]
Другой вид Dromaeosaurid, микрораптор-графический интерфейс , возможно, был способен скользить, используя его хорошо развитые крылья как на передних, так и на задних конечностях. Исследование, проведенное в 2005 году Санкара Чаттерджи, показало, что крылья микрораптора » с доплошным уровнем функционировали как « биплан , и что он, вероятно, использовал стиль скольжения фагоида , в котором он будет запущен с окуня и вниз в U-образной кривой , затем снова поднимитесь, чтобы приземлиться на другое дерево, с хвостовыми и задними крыльями, помогающими контролировать его положение и скорость. Chatterjee также обнаружил, что Microraptor имел основные требования для поддержания полета, обеспеченного уровнем, в дополнение к скольжению. [ 25 ]
Changyuraptor Yangi является близким относительным графическим интерфейсом микрораптора , который также считается планером или флаером, основанным на наличии четырех крыльев и аналогичных пропорций конечностей. Тем не менее, это значительно большее животное, размером с дикую индейку , являющееся одним из самых больших известных мезозозой паравиан.
Другие виды Dromaeosaurid, Deinonychus antirrhopus , могут демонстрировать частичные полеты полета. Молодые из этого вида носили более длинные руки и более надежные грудные бакировки, чем взрослые, и которые были похожи на те, которые наблюдались в других развевающихся тероподах, подразумевая, что они могли быть способными к бегству, когда молодые, а затем теряли способность по мере их роста. [ 84 ]
Возможность того, что Sinornithosaurus millenii способен скользить или даже питать полет, также был воспитан несколько раз, [ 85 ] [ 86 ] Хотя дальнейших исследований не было.
Zhenyuanlong сохраняет крылышки, которые имеют аэродинамическую форму, с особенно похожими на птицами чехлы, в отличие от более длинных, более широких покрывающих покрытий таких форм, как Archeopteryx и Anchiornis , а также слитые стернальные пластины. Из -за его размера и коротких рук маловероятно, что Zhenyuanlong был способен к приводительному полету (хотя важность биомеханического моделирования в этом отношении подчеркнута [ 31 ] ), но это может указывать на относительно близкий потомки от летающих предков или даже некоторую способность к склону скольжения или наклона с помощью крыла .
Палеопатология
[ редактировать ]В 2001 году Брюс Ротшильд и другие опубликовали исследование, в котором изучались данные о стрессовых переломах и адульсиях сухожилий у динозавров теропод и последствия для их поведения. Поскольку стрессовые переломы вызваны повторной травмой, а не сингулярными событиями, они с большей вероятностью будут вызваны регулярным поведением, чем другие виды травм. Исследователи обнаружили, что поражения, подобные тем, которые вызваны стрессовыми переломами на когти ручной работы Dromaeosaurid, одно из двух таких поражений когтей, обнаруженных в ходе исследования. Стрессовые переломы в руках имеют особое поведенческое значение по сравнению с тем, что встречаются в ногах, поскольку переломы стресса в ногах могут быть получены во время бега или во время миграции. Травмы руки, напротив, с большей вероятностью будут получены при контакте с борьбой добычей. [ 87 ]
Плавание
[ редактировать ]
По крайней мере, одна группа Dromaeosaurid, Halskaraptorinae , чьи членами являются Halskaraptorines, скорее всего, были специализированы на водных или полуаватических привычках, развивших пропорции конечности, морфологию зубов и грудную клетку , схожи с давничными птицами. [ 49 ] [ 88 ] [ 89 ]
Рыболочные привычки были предложены для неэнлагинов , включая сравнения с приписываемыми полу-квадратными спинозавридами , [ 90 ] Но любые механизмы водного движения до сих пор не обсуждались.
Размножение
[ редактировать ]
В 2006 году Греллет-Тиннер и Маковицки сообщили о яйце, связанном с образцом Deinonychus . Яйцо разделяет сходство с яйцами овирапторида , и авторы интерпретировали ассоциацию как потенциально указывающую на размышление. [ 91 ] Исследование, опубликованное в ноябре 2018 года, Norell, Yang and Wiemann et al., Указывает, что Deinonychus откладывает синие яйца, вероятно, замаски из них, а также создает открытые гнезда. Другие дромаозавриды, возможно, сделали то же самое, и теоретизируется, что они и другие манирапторанские динозавры, возможно, были происхождением для откладывания цветных яиц и создания открытых гнезд, как сегодня делают многие птицы. [ 92 ] [ 93 ] [ 94 ]
В популярной культуре
[ редактировать ]Velociraptor , Dromaeosaurid, привлек много внимания после того, как он был заметно в Спилберга фильме 1993 года . Тем не менее, размеры велоцираптора в фильме намного больше, чем самые крупные члены этого рода. Роберт Баккер вспомнил, что Спилберг был разочарован размерами велосираптора и таким образом увеличил его. [ 95 ] Грегори С. Пол , в своей книге «Хищные динозавры мира» 1988 года , также считался Deinonychus antirrhopus видом велосираптора и переигрывал виды Velociraptor Antirrhopus . [ 39 ] Это таксономическое мнение не было широко соблюдается. [ 10 ] [ 96 ] [ 97 ]
Временная шкала родов Dromaeosaurid
[ редактировать ]
Смотрите также
[ редактировать ]Ссылки
[ редактировать ]- ^ Хартман, Скотт; Мортимер, Микки; Wahl, William R.; Lomax, Dean R.; Липпинкотт, Джессика; Lovelace, David M. (2019-07-10). «Новый паравийский динозавр из позднего юрского периода Северной Америки поддерживает позднее приобретение полета птиц» . ПЕРЕЙ . 7 : E7247. doi : 10.7717/peerj.7247 . ISSN 2167-8359 . PMC 6626525 . PMID 31333906 .
- ^ FORFIRI, John D.; Страна, Маттия А.; 2 святой, Доминика Д.; Gianechini, Federico A.; Питман, Майкл; Ламанна, Мэтью С. (2024-06-14). Diuqin lechiguanae gen. et sp. nov., a new unenlagiine (Theropoda: Paraves) from the Bajo de la Carpa Formation (Neuquén Group, Upper Cretaceous) of Neuquén Province, Patagonia, Argentina"Полем BMC Экология и эволюция 24 (1): 77. Bibcode : 2024bme . doi : /s12862-024-0 10.1186 ISSN 2730-7 11177497PMC PMID 38872101 .
- ^ Аллейн, Ронан; Pearl-Suberbiola, Xabier (2003). "Динозавры Франции" Рендус учетные записи Palevol 2 (1): 27–44. Bibcode : 2003crpal . два 10.1016/S1631-0683(03)00002-2:
- ^ Agnolin, Federico L.; Motta, Matias J.; Бриссон он, Федерико; Ло Коко, Гастон; Novas, Fernando E. (2019-02-12). «Паравиская филогения и переход динозавра-птицы: ovserview» . Границы в науке Земли . 6 Doi : 10.3389/feart.2018 00252. HDL : 11336/130197 . ISSN 2296-6463 .
- ^ Jump up to: а беременный в Holtz, Thomas R. Jr. (2012) Динозавры: самая полная, современная энциклопедия для любителей динозавров всех возрастов, Приложение Winter 2011.
- ^ Acorn, J. (2007). Глубокая Альберта: ископаемые факты и раскопки динозавров . Университет Альберты Пресс. с. 13 . doi : 10.1515/9780888648518 . ISBN 978-0-88864-481-7 .
- ^ Барретт, Пол М.; Бенсон, Роджер Б.Дж.; Рич, Томас Х.; Викерс-Рич, Патриция (23 декабря 2011 г.). «Первый спинозауридный динозавр из Австралии и космополитизм меловых динозавров фауны» . Биологические письма . 7 (6): 933–936. doi : 10.1098/rsbl.2011.0466 . PMC 3210678 . PMID 21693488 .
- ^ Metcalf, SJ; Воган, RF; Бентон, MJ; Коул, Дж.; Симмс, MJ; Дартналл, Д.Л. (январь 1992 г.). «Новый батонический (средний юрский) сайт микровертозвоночных, в рамках формирования известняка Norton Norton в карьере Hornsleasow, Глостершир». Материалы Ассоциации геологов . 103 (4): 321–342. Bibcode : 1992prga..103..321m . doi : 10.1016/s0016-7878 (08) 80129-0 .
- ^ Wills, S.; Андервуд, CJ; Барретт, PM (2023). «Машинное обучение подтверждает новые записи о манирапторанских тероподах в средней юрской игре в Великобритании Фауны Микроверно -краты» . Документы по палеонтологии . 9 (2). E1487. Bibcode : 2023ppal .... 9E1487W . doi : 10.1002/spp2.1487 .
- ^ Jump up to: а беременный в дюймовый Норелл, Массачусетс; Маковицки, П.Дж. (2004). "DrimaeOssauridae" В Вешампелле, дБ; Додсон, П.; Osmóllska, H. (Eds.). Данурия (2 -е изд.). Беркли: Университет Калифорнийской прессы. стр. 196–210. ISBN 978-0-520-24209-8 .
- ^ Jump up to: а беременный в Hwang, SH; Норелл, Массачусетс; Ji, Q.; Гао, К. (2002). "Teropopoda : Dr. Dromaeridae) : 3381 : 44pp. HDL : 2246/2870 .
- ^ Перл, А.; Норелл, Массачусетс; Кларк, Дж. (1999). «Новый Maniraptoran Theropod - Achillobator Giganticus (Dromaeosauridae) - от верхнего мела Бурханта, Монголия». Вклад монгольско-американского палеонтологического проекта . 101 : 1–105.
- ^ Naish, D. Hutt и Martill, DM (2001). «Сауришские динозавры: Тероподы». В Martill, DM и Naish, D. (Eds). Динозавры острова Уайт . Палеонтологическая ассоциация, полевые направляющие к окаменелостям. 10, 242–309.
- ^ «Дино охотится на сеть редких зубов раптора» . BBC News . 26 апреля 2004 г.
- ^ Jump up to: а беременный в Ostrom, JH (1969). «Остеология Deinonychus antirrhopus , необычный теропод из нижнего мела Монтаны» (PDF) . Бюллетень музея Пибоди естественной истории . 30 : 1–165. Архивировано из оригинала (PDF) 2010-06-27 . Получено 2010-12-18 .
- ^ Пол, Грег (январь 2006 г.). «Рисование динозавров» . Геотейс .
- ^ Jump up to: а беременный Маковицки, Питер Дж.; Appestuia, Себастья; Агнолин, Федерико Л. (октябрь 2005 г.). «Эрлст -Дромауридирод из Южной Америки» Природа 437 (7061): 1007–1011. Bibcode : 2005 Natur.437.1007m . doi : 10.1038/ природа PMID 16222297 . S2CID 27078534 .
- ^ Агнолин, Федерико; Novas, Fernando E. (2013). Птиевые предки . Springerbriefs в науках о Земле. Дордрехт: Спрингер Нидерланды. doi : 10.1007/978-94-007-5637-3 . ISBN 978-94-007-5636-6 Полем S2CID 199493087 .
- ^ Кодд, Джонатан Р; Мэннинг, Филипп Л; Норэлл, Марк А; Перри, Стивен Ф. (2008-01-22). «Птиеподобная механика для дыхания в динозаврах манирапторов» . Труды Королевского общества B: Биологические науки . 275 (1631): 157–161. doi : 10.1098/rspb.2007.1233 . ISSN 0962-8452 . PMC 2596187 . PMID 17986432 .
- ^ Пол, Грегори С. (1988). Хищные динозавры мира: полный иллюстрированный гид . Нью-Йорк. ISBN 0-671-61946-2 Полем OCLC 18350868 .
{{cite book}}: CS1 Maint: местоположение отсутствует издатель ( ссылка ) - ^ Jump up to: а беременный Ли, Рихуи; Локли, Мартин Дж.; Маковицки, Питер Дж.; Мацукава, Масаки; Norell, Mark A.; Харрис, Джеральд Д.; Лю, Мингвей (19 октября 2007 г.). «Поведенческие и фауновые последствия ранних меловых дорожек -деинонихозавра из Китая». Naturwissenschaften . 95 (3): 185–191. Bibcode : 2008nw ..... 95..185L . doi : 10.1007/s00114-007-0310-7 . PMID 17952398 . S2CID 16380823 .
- ^ Jump up to: а беременный Longrich, NR; Керри, PJ (2009). «Микрорапторин (Dinosauria -Dromaeosauridae) из позднего мела Северной Америки» . ПНА . 106 (13): 5002–7. Bibcode : 2009pnas..106.5002L . doi : 10.1073/pnas.0811664106 . PMC 2664043 . PMID 19289829 .
- ^ Csiki, Zoltán; Времир, Матиас; Брусатт, Стивен Л.; Норэлл, Марк А. (31 августа 2010 г.). «Аберрантный динозавр теропода с аберрантным островом из покойного мела Румынии» . Труды Национальной академии наук Соединенных Штатов Америки . 107 (35): 15357–15361. BIBCODE : 2010PNAS..10715357C . doi : 10.1073/pnas.1006970107 . PMC 2932599 . PMID 20805514 .
- ^ Jump up to: а беременный Norell, Mark A.; Маковицки, Питер Дж. (1999). «Важные особенности скелета Dromaeosaurid II: Информация из недавно собранных образцов Velociraptor mongoliensis ». Американский музей новичок (3282): 1–45. HDL : 2246/3025 .
- ^ Jump up to: а беременный Чаттерджи, Санкар; Templin, R. Jack (30 января 2007 г.). «Биплановый план плана и эффективность полета пернатого графического интерфейса микрораптора динозавров» . Труды Национальной академии наук Соединенных Штатов Америки . 104 (5): 1576–1580. Bibcode : 2007pnas..104.1576c . doi : 10.1073/pnas.0609975104 . PMC 1780066 . PMID 17242354 .
- ^ Jump up to: а беременный в дюймовый , Сюй xing ; ..
- ^ Jump up to: а беременный Сюй, Син; Ван, Сяо-Лин; Wu, Xiao-Chun (сентябрь 1999 г.). «Динозавр Dromaeosaurid с нитевидным покровом из формирования Йикса в Китае». Природа . 401 (6750): 262–266. Bibcode : 1999natur.401..262X . doi : 10.1038/45769 . S2CID 4430574 .
- ^ Jump up to: а беременный в Тернер, ах; Маковицки, PJ; Норэлл, Массачусетс (21 сентября 2007 г.). «Ручки перо перо в велосирапторе динозавра» . Наука . 317 (5845): 1721. Bibcode : 2007sci ... 317.1721t . doi : 10.1126/science.1145076 . PMID 17885130 .
- ^ Jump up to: а беременный Пол, Грегори С. (2002). Динозавры воздуха: эволюция и потеря полета у динозавров и птиц. Балтимор: издательство Джона Хопкинса. 472 стр.
- ^ Prum, Richard O.; Brush, Alan H. (сентябрь 2002 г.). «Эволюционное происхождение и диверсификация перьев». Ежеквартальный обзор биологии . 77 (3): 261–295. doi : 10.1086/341993 . PMID 12365352 . S2CID 6344830 .
- ^ Jump up to: а беременный Лю, Джунчан; Брусатт, Стивен Л. (16 июля 2015 г.). «Большой, короткий, крылатый дромозаврид (Dinosauria: Theropoda) из раннего мела Китая и его последствия для эволюции перьев» . Научные отчеты . 5 (1): 11775. Bibcode : 2015natsr ... 511775L . doi : 10.1038/srep11775 . PMC 4504142 . PMID 26181289 .
- ^ Jump up to: а беременный «Первый гигантский Raptor (Theropoda: Dromaeosauridae) из формирования Hell Creek» . Палеонтологические вклад . 30 октября 2015 года. DOI : 10.17161/paleo.1808.18764 .
- ^ Jasinski, Steven E.; Салливан, Роберт М.; Додсон, Питер (2020-03-26). «Новый динозавр Dromaeosaurid (Theropoda, Dromaeosauridae) из Нью -Мексико и биоразнообразие Dromaeosaurids в конце меловых» . Научные отчеты . 10 (1): 5105. Bibcode : 2020natsr..10.5105J . doi : 10.1038/s41598-020-61480-7 . ISSN 2045-2322 . PMC 7099077 . PMID 32218481 .
- ^ Кау, А. (2024). «Унифицированная структура для макроэволюции хищных динозавров» (PDF) . Bollettino della società paleontologica italiana . 63 (1). doi : 10.4435/bspi.2024.08 . Архивировано из оригинала (PDF) 27 апреля 2024 года.
- ^ Norell, M. Clark, JM, Makovicky, PJ (2001). « Филогенетические отношения между колурозаврными тероподами. Архивируемые 2007-07-13 на машине Wayback « Новые перспективы происхождения и эволюции птиц: Материалы Международного симпозиума в честь Джона Х. Острома », Музей Йельского Пибоди: 49–67
- ^ Падиан, Кевин; Диал, Кеннет П. (2005). "Происхождение полета: могут ли« четырехкрытые динозавры летать? " Полем Природа . 438 (7066): E3, обсуждение E3–4. Bibcode : 2005natur.438e ... 3p . doi : 10.1038/nature04354 . PMID 16292258 . S2CID 4314459 .
- ^ Witmer, LM (2005) «Дебаты о птичьем происхождении; филогения, функция и окаменелости». В «Мезозойских птицах: над головами динозавров», стр. 3–30. ISBN 0-520-20094-2
- ^ Ольшевский, Джордж. (1994). «Сначала птицы? Теория, соответствующая фактам - эволюции рептилий в птиц». Omni , июнь 1994. Том 16 № 9
- ^ Jump up to: а беременный Пол, Грегори С. (1988). Хищные динозавры мира. Нью -Йорк: Саймон и Шустер. 464 стр.
- ^ Czerkas, SA, Zhang, D., Li, J. и Li, Y. (2002). «Летающие дромаозавры», в Черкасе, SJ (ред.): Пернатые динозавры и происхождение полета: журнал музея динозавров .
- ^ Мартин, Л.Д. (2004). " PDF) . Acta Geologic Synica 50 (6): 978–990.
- ^ National Geographic News- у самой ранней птицы были такие ноги, как динозавр, ископаемые шоу -Николас Бакалар, 1 декабря 2005 г., стр. 2. Получено 2006-10-18.
- ^ Mayr, G.; Пол, б; Петерс, DS (2 декабря 2005 г.). «Хорошо сохранившийся образец Archeopteryx с функциями Theropod» (PDF) . Наука . 310 (5753): 1483–1486. Bibcode : 2005sci ... 310.1483m . doi : 10.1126/science.1120331 . PMID 16322455 . S2CID 28611454 .
- ^ Mayr, G. (1 сентября 2006 г.). «Ответ на комментарий на« хорошо сохранившийся образец Archeopteryx с функциями Theropod » . Наука . 313 (5791): 1238c. Bibcode : 2006sci ... 313.1238m . doi : 10.1126/science.1130964 .
- ^ Корф, IJ; Батлер, RJ (1 сентября 2006 г.). «Прокомментируйте« хорошо сохранившийся образец Archeopteryx с функциями Theropod » . Наука . 313 (5791): 1238b. Bibcode : 2006sci ... 313.1238c . doi : 10.1126/science.1130800 . PMID 16946054 . S2CID 19859320 .
- ^ Тернер, Алан Х.; Пол, Диего; Кларк, Джулия А.; Эриксон, Грегори М.; Норэлл, Марк (2007). «Базальный Dromaeosaurid и размер эволюция, предшествующая полету птичьего вещества» . Наука . 317 (5843): 1378–1381. Bibcode : 2007sci ... 317.1378t . doi : 10.1126/science.1144066 . PMID 17823350 .
- ^ Jump up to: а беременный Senter, P.; Киркленд, Джи; Deblieux, DD; Madsen, S.; Тот, Н. (2012). Додсон, Питер (ред.). «Новые дромазавриды (Dinosauria: Theropoda) из нижнего мела Юты и эволюция хвоста Dromaeosaurid» . Plos один . 7 (5): E36790. Bibcode : 2012ploso ... 736790S . doi : 10.1371/journal.pone.0036790 . PMC 3352940 . PMID 22615813 .
- ^ Мэтью, WD; Браун, Б. (1922). «Семейство Deinodontidae, с уведомлением о новом роде из мела Альберты». Бюллетень Американского музея естественной истории . 46 : 367–385. HDL : 2246/1300 .
- ^ Jump up to: а беременный в дюймовый Кау, а.; Бейран, В.; Voeten, dfae; Фернандес, В.; Tafforeau, P.; Stein, K.; Barsbold, R.; Tsogtbaatar, K.; Керри, PJ; Godefroit, P. (2017). «Синхротроновое сканирование выявляет амфибийную экоморфологию в новой кладе динозавров, похожих на птицы» . Природа . 552 (7685): 395–399. Bibcode : 2017natur.552..395c . doi : 10.1038/nature24679 . PMID 29211712 . S2CID 4471941 .
- ^ Тернер, Алан Х.; Hwang, Sunny H.; Норелл, Марк А. (2007). «Небольшой производный теропод из Оёша, раннего мела, Бэйкхангор Монголия» . Американский музей новичок (3557): 1. doi : 10.1206/0003-0082 (2007) 3557 [1: asdtfs] 2.0.co; 2 . HDL : 2246/5845 . S2CID 31096081 .
- ^ Agnolin, Federico L.; Новости, Фернандо Э. (март 2011 г.). «Unenlagiid Theropods: они члены Dromaeosauridae (Theropoda, Manirato)?» Полем Материалы Бразильской академии наук . 83 (1): 117–162. Doi : 10.1590/s0001-37652011000100008 . PMID 21437379 .
- ^ Лонгрич, Николас Р.; Керри, Филипп Дж. (31 марта 2009 г.). «Микрорапторин (Dinosauria -Dromaeosauridae) из позднего мела Северной Америки» . Труды Национальной академии наук Соединенных Штатов Америки . 106 (13): 5002–5007. Bibcode : 2009pnas..106.5002L . doi : 10.1073/pnas.0811664106 . PMC 2664043 . PMID 19289829 .
- ^ Сендер, Фил; Barsbold, R.; Бритт, Брукс Б.; Бернхэм, Дэвид Б. (2004). «Систематика и эволюция Dromaeosauridae (Dinosauria, Theropoda)». Бюллетень музея естественной истории Ганма . 8 : 1–20.
- ^ Серено, PC 2005. STEM Archosauria-Taxonsearch Archived 2009-01-15 на машине Wayback , версия 1.0, 7 ноября 2005 г.
- ^ Тернер, Алан Х.; Маковицки, Питер Дж.; Норэлл, Марк А. (17 августа 2012 г.). «Обзор систематики Dromaeosaurid и Paravian Phologeny» . Бюллетень Американского музея естественной истории . 371 : 1–206. doi : 10.1206/748.1 . HDL : 2246/6352 . S2CID 83572446 .
- ^ Schmitz, L.; Мотани Р. (14 апреля 2011 г.). «Ночная территория у динозавров выводится из морфологии склерального кольца и орбиты». Наука . 332 (6030): 705–708. Bibcode : 2011sci ... 332..705s . doi : 10.1126/science.1200043 . PMID 21493820 . S2CID 33253407 .
- ^ Ли, Quanguo; Гао, К.-Q.; Meng, Q.; Кларк, JA; Shawkey, MD; D'Alba, L.; PEI, R.; Эллисон, М.; Норелл, Массачусетс; Винтер Дж. (9 марта 2012 г.). «Реконструкция микрораптора и эволюция переливающегося оперения». Наука . 335 (6073): 1215–1219. BIBCODE : 2012SCI ... 335.1215L . doi : 10.1126/science.1213780 . PMID 22403389 . S2CID 206537426 .
- ^ Зеленицкий, Дарла К; Терриен, Франсуа; Кобаяши, Йошицугу (28 октября 2008 г.). «Околочная острота в тероподах: палеобиологические и эволюционные последствия» . Труды Королевского общества B: Биологические науки . 276 (1657): 667–673. doi : 10.1098/rspb.2008.1075 . PMC 2660930 . PMID 18957367 .
- ^ Зеленицкий, Дарла К.; Терриен, Франсуа; Риджли, Райан С.; МакГи, Аманда Р.; Витмер, Лоуренс М. (13 апреля 2011 г.). «Эволюция обоняния у неавийских тероподных динозавров и птиц» . Труды Королевского общества B: Биологические науки . 278 (1725): 3625–3634. doi : 10.1098/rspb.2011.0238 . PMC 3203493 . PMID 21490022 .
- ^ Ищак, а.; Уилкинсон, Р.; Arbor, VM; Руиз-Оменья, Джи; Currie, PJ (2018). «Биомеханика прокола и удара в зубах хищных динозавров колурозавры» . Текущая биология . 28 (9): 1467–1474. BIBCODE : 2018CBIO ... 28E1467T . doi : 10.1016/j.cub.2018.03.042 . PMID 29706515 .
- ^ Адамс, Дон (1987). «Чем больше они, тем сложнее они падают: последствия ишиальной кривизны у цератопсских динозавров». В Керри, Филипп Дж.; Костер Э. (ред.). Четвертый симпозиум на мезозойских наземных экосистемах . Драмхеллер, Канада: Музей Тиррелла. С. 1–6.
- ^ Мэннинг, Филипп Л; Пейн, Дэвид; Пенникотт, Джон; Барретт, Пол М; Эннос, Роланд А (11 октября 2005 г.). "Убийцы динозавров или кольки для подъема?" Полем Биологические письма . 2 (1): 110–112. doi : 10.1098/rsbl.2005.0395 . PMC 1617199 . PMID 17148340 .
- ^ Jump up to: а беременный Дэвис, SJJF (2002) «Рэтиты и тинамоуссовая» издательство Оксфордского университета. Нью -Йорк, США [ страница необходима ]
- ^ Гиллиард, Эрнест Томас (1958). Живые птицы мира . Doubleday. OCLC 681791956 . [ страница необходима ]
- ^ Кофрон, Кристофер П. (1999). «Атаки на людей и домашних животных южного кассовари (Касуарий Касуарий Джонсон) в Квинсленде, Австралия». Журнал зоологии . 249 (4): 375–381. doi : 10.1111/j.1469-7998.1999.tb01206.x .
- ^ Кофрон, Кристофер П. (2003). «История случаев атак южного кассоварию в Квинсленде». Мемуары музея Квинсленда . 49 (1): 335–338.
- ^ Редфорд, Кент Х.; Петерс, Густав (1986). «Примечания о биологии и песне красной ноги с серимой (Кариама Кристата)». Журнал поля орнитологии . 57 (4): 261–269. JSTOR 4513154 .
- ^ Мэннинг, Филипп Л.; Маргеттс, Ли; Джонсон, Марк Р.; Withers, Philip J.; Продавцы, Уильям I.; Фалкингем, Питер Л.; Mummery, Paul M.; Барретт, Пол М.; Реймонт, Дэвид Р. (сентябрь 2009 г.). «Биомеханика когтей динозавров дромаозавридов: применение рентгеновской микротомографии, наноинентация и анализа конечных элементов» . Анатомическая запись: достижения в области интегративной анатомии и эволюционной биологии . 292 (9): 1397–1405. doi : 10.1002/ar.20986 . PMID 19711472 .
- ^ «Когти -когти велосираптора были для лазания» . Новый ученый . 203 (2725): 10. Сентябрь 2009 г.. DOI : 10.1016/S0262-4079 (09) 62381-2 . Получено 2009-09-15 .
- ^ Центр, Фил (2009). ) (PDF . Исторические гисторты 13 : 1–1
- ^ Фаулер, Денвер W.; Фридман, Элизабет А.; Scannella, John B.; Камбич, Роберт Э. (14 декабря 2011 г.). «Хищная экология Deinonychus и происхождение хлопья у птиц» . Plos один . 6 (12): E28964. BIBCODE : 2011PLOSO ... 628964F . doi : 10.1371/journal.pone.0028964 . PMC 3237572 . PMID 22194962 .
- ^ Чой, Чарльз (14 декабря 2011 г.). «Убийственные когти велосирапторов помогли им съесть добычу» . Livescience .
- ^ Епископ, PJ (2019). «Проверка функции Dromaeosaurid (Dinosauria, Theropoda)« серповые когти »посредством моделирования и оптимизации опорно -двигательного аппарата» . ПЕРЕЙ . 7 : E7577. doi : 10.7717/peerj.7577 . PMC 6717499 . PMID 31523517 .
- ^ Gianechini, Federico A.; Эколи, Маркос Д.; Диас-Мартинес, Игнасио (5 февраля 2020 г.). «Дифференциальные локомоторные и хищные стратегии Гондванана и полученных лауразианских дромазавридов (Dinosauria, Theropoda, Paraves): выводы из морфометрических и сравнительных анатомических исследований» . Журнал анатомии . 236 (5): 772–797. doi : 10.1111/joa.13153 . PMC 7163733 . PMID 32023660 .
- ^ Mudroch, A.; Richter, U.; Joger, U.; Косма, Р.; Idé, o.; Мага, А. (2011). «Дидактильные треки парависких теропод (Манираптора) из« Средней юры Африки » . Plos один . 6 (2): E1462. Bibcode : 2011ploso ... 614642m . doi : 10.1371/journal.pone.0014642 . PMC 3038851 . PMID 21339816 . Рисунок 4.
- ^ Максвелл, В. Десмонд; Остром, Джон Х. (27 декабря 1995 г.). «Тафономия и палеобиологические последствия ассоциаций Tenontosaurus - deinonychus ». Журнал палеонтологии позвоночных . 15 (4): 707–712. Bibcode : 1995jvpal..15..707m . doi : 10.1080/02724634.1995.10011256 .
- ^ Роуч, Брайан Т.; Бринкман, Даниэль Л. (апрель 2007 г.). «Переоценка кооперативной охоты и общительности в Deinonychus antirrhopus и других неавийских динозаврах теропод». Бюллетень музея естественной истории Пибоди . 48 (1): 103–138. doi : 10.3374/0079-032x (2007) 48 [103: Acocph] 2.0.co; 2 . S2CID 84175628 .
- ^ Фредериксон, JA; Энгель, MH; Cifelli, RL (15 августа 2020 г.). «Онтогенетические диетические сдвиги в Deinonychus antirrhopus (Theropoda; Dromaeosauridae): понимание экологии и социального поведения динозавров с посредством стабильного изотопного анализа». Палеогеография, палеоклиматология, палеоэкология . 552 : 109780. Bibcode : 2020ppp ... 55209780f . doi : 10.1016/j.palaeo.2020.109780 . S2CID 219059665 .
- ^ Switek, Брайан (7 января 2015 г.). «Смертельная ловушка« Динозавр Юты »раскрывает Trove из гигантских хищников» . National Geographic News . Архивировано с оригинала 30 ноября 2019 года.
- ^ Хатсон, Джоэл Д.; Хатсон, Келда Н. (25 сентября 2015 г.). «Исследование подвижности кости предплечья в аллигаторе Mississippiensis (Daudin, 1802) и Struthio Camelus Linnaeus, 1758, показывает, что археоптерикс и дромаозавры разделяют адаптацию для скольжения и/или хлопания» . Geodiversitas . 37 (3): 325–344. doi : 10.5252/g2015n3a3 . S2CID 55278850 . Получено 2 декабря 2022 года .
- ^ Norell, Mark A.; Кларк, Джеймс М.; Тернер, Алан Х.; Маковицки, Питер Дж.; Барсболд, Ринчен; Роу, Тимоти (2006). «Новый Dromasomaurid Theropod от Ukhaa Tolgod (ömnögov, Mongolia) » Американский музей новичок (3545): 1. doi : 10.1206/0003-0082 (2006) 3545 [1: итфу] .0.co ; HDL : 2246/5
- ^ Форстер, Калифорния; Сэмпсон, SD; Chiappe, LM; Краузе, DW (20 марта 1998 г.). «Тероподский происхождение птиц: новые доказательства от покойного мела Мадагаскара». Наука . 279 (5358): 1915–1919. Bibcode : 1998sci ... 279.1915f . doi : 10.1126/science.279.5358.1915 . PMID 9506938 .
- ^ Chiappe, LM (2007-02-02). Прославленные динозавры: происхождение и ранняя эволюция птиц . Сидней: UNSW Press. ISBN 978-0-471-24723-4 . [ страница необходима ]
- ^ Парсонс, Уильям Л.; Парсонс, Кристен М.; Додсон, Питер (15 апреля 2015 г.). «Морфологические вариации в онтогенности Deinonychus antirrhopus (Theropoda, Dromaeosauridae)» . Plos один . 10 (4): E0121476. BIBCODE : 2015PLOSO..1021476P . doi : 10.1371/journal.pone.0121476 . PMC 4398413 . PMID 25875499 .
- ^ Чаттерджи, Санкар; Templin, RJ (2004). «Пернатые коэлурозавры из Китая: новый свет на древесное происхождение птичьего полета» . В Керри, Филипп Дж.; Koppelhus, Eva B.; Шугар, Мартин А.; Райт, Джоанна Л. (ред.). Печатные драконы: исследования перехода от динозавров к птицам . Издательство Университета Индианы. С. 251–281. ISBN 978-0-253-34373-4 .
- ^ Найш, Даррен (2012). Планета Динозавр: следующее поколение убийственных гигантов . Книги огня. п. 192 . ISBN 978-1-77085-049-1 .
- ^ Rothschild, B.; Танк, DH; Ford, TL (2001). «Темопод стрессовые переломы и авульсии сухожилия как подсказка для активности». В Танке, DH; Карпентер, К. (ред.). Мезозойская жизнь позвоночных . Издательство Университета Индианы. С. 331–336.
- ^ Кау, А. (2020). «План тела Halskaraptor Escuilliei (Dinosauria, Theropoda) не является переходной формой вдоль эволюции гиперкарнословия Dromaeosaurid» . ПЕРЕЙ . 8 : E8672. doi : 10.7717/peerj.8672 . PMC 7047864 . PMID 32140312 .
- ^ Lee, S.; Lee, Y.-N.; Керри, PJ; Sissons, R.; Парк, J.-Y.; Ким, С.-Х.; Barsbold, R.; Цогтбаатар, К. (2022). «Неавийский динозавр с упорядоченным телом демонстрирует потенциальную адаптацию для плавания» . Биология связи . 5 (1185): 1185. DOI : 10.1038/S42003-022-04119-9 . ISSN 2399-3642 . PMC 9715538 . PMID 36456823 .
- ^ Керри, Филипп Дж.; Карабоджал, Ариана Полин (декабрь 2012 г.). ) . Pol, Canale, Porfiri, Porfiri, меловые (Dosauliaidae Амегина 49 (4): 662–667. doi : 10.5710/amgh.30.8.8.2012.57 . HDL : 11336/9090 . S2CID 129058582 .
- ^ Греллет-Тиннер, Джеральд; Маковицки, Питер (1 июня 2006 г.). «Возможное яйцо Dromaeosaur deinonychus antirrhopus : филогенетические и биологические последствия». Канадский журнал наук о Земле . 43 (6): 705–719. Bibcode : 2006cajes..43..705g . doi : 10.1139/e06-033 .
- ^ Wiemann, Jasmina; Ян, Цзы-Рюй; Норелл, Марк А. (31 октября 2018 г.). «Яичный цвет динозавров имел одно эволюционное происхождение». Природа . 563 (7732): 555–558. Bibcode : 2018natur.563..555W . doi : 10.1038/s41586-018-0646-5 . PMID 30464264 . S2CID 53188171 .
- ^ «Яичный цвет динозавров имел одно эволюционное происхождение» . Ноябрь 2018 года.
- ^ «Динозавры кладут все цветные птичьи яйца в одну корзину, эволюционно говоря» . 2018-10-31.
- ^ Баккер, Роберт Т. (1995). Raptor Red . Нью -Йорк: Bantam Books. п. 4. ISBN 978-0-553-57561-3 .
- ^ Pérez-Mooreno, BP; Sanz, JL; Sudre, J.; Сиге Б. (1994). «Тероподный динозавр из нижнего мела на юге Франции». Динозавры и другие ископаемые рептилии Европы, Симпозиум Cuvier Second Georges, Монбэлиард; Палеобиология обзор, специальный том . 7 : 173–188.
- ^ Currie, PJ (1995). «Новая информация об анатомии и отношениях Dromaeosaurus albertensis (Dinosauria: Theropoda)». Журнал палеонтологии позвоночных . 15 (3): 576–591. Bibcode : 1995jvpal..15..576c . doi : 10.1080/02724634.1995.10011250 . ( Аннотация Архивировано 2007-09-27 на машине Wayback )
Внешние ссылки
[ редактировать ]- Dromaeosauridae: The Raptors! , из Калифорнийского университета Музея палеонтологии Беркли.
- Dinosaurs - Полное и бесплатное онлайн -издание книги «Динозавры», написанная WD Matthew (цитируется в этой статье с авторством семейства Dromaeosauridae), и бывшего куратора палеонтологии позвоночных в Американском музее естественной истории в Нью -Йорке; Первоначально опубликовано в 1915 году
- DROMAEOSAURIDAE , Ссылка на мир динозавров с подробным описанием и фотографиями многих DROMAEOSAURIDAE DINOSAURS

