Обыкновенный канюк
| Обыкновенный канюк | |
|---|---|

| |
| Степной канюк B. vulpinus | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Птицы |
| Заказ: | похожий на ястреба |
| Семья: | Accipitridae |
| Род: | Бутео |
| Разновидность: | Б. Бьют
|
| Биномиальное имя | |
| Бутео Бутео | |
| Подвиды | |
|
7–10, см. текст | |
| |
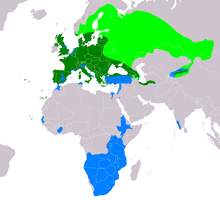
| Ареал B.buteo Ареал размножения мигрантов Резидентный диапазон Ареал зимовки переселенческих популяций
| |
| Синонимы | |
|
Фалько Бутео Линней, 1758 г. | |

Канюк обыкновенный ( Buteobuteo ) [ 2 ] средних и крупных размеров — хищная птица , имеющая большой ареал. Это представитель рода Buteo. [ 2 ] семейства Accipitridae из [ 2 ] Этот вид обитает на большей части территории Европы и расширяет свой ареал размножения на большей части Палеарктики вплоть до северо-западного Китая ( Тянь-Шань ), далекой западной Сибири и северо-западной Монголии . [ 2 ] [ 1 ] [ 3 ] На большей части своего ареала он обитает круглый год. Однако канюки из более холодных частей Северного полушария , а также те, которые размножаются в восточной части своего ареала, обычно мигрируют на юг на северную зиму, многие из них добираются до Южной Африки . [ 2 ] [ 4 ]
Обыкновенный канюк — оппортунистический хищник, который может ловить самую разнообразную добычу, но питается в основном мелкими млекопитающими , особенно грызунами , такими как полевки . Обычно он охотится с насеста. [ 5 ] Как и большинство хищных птиц ястреба, он строит гнездо, обычно на деревьях этого вида, и является преданным родителем относительно небольшого выводка птенцов. [ 3 ] Обыкновенный канюк, по-видимому, является наиболее распространенным дневным хищником в Европе, поскольку, по оценкам, его общая популяция в мире исчисляется миллионами. [ 3 ] [ 6 ]
Таксономия
[ редактировать ]Первое официальное описание обыкновенного канюка было сделано шведским натуралистом Карлом Линнеем в 1758 году в десятом издании его Systema Naturae под биномиальным названием Falcobuteo . [ 7 ] Род Buteo был введен французским натуралистом Бернаром Жерменом де Ласепедом в 1799 году путем таутонимии с видовым названием этого вида. [ 8 ] [ 9 ] Слово бутео означает в переводе с латыни канюк. [ 10 ] Его не следует путать с стервятником-индейкой , которого в американском английском иногда называют канюком.
Подсемейство Buteoninae и наиболее разнообразно произошло из Северной и Южной Америки , иногда с более широким распространением, которое привело к появлению обыкновенных канюков и других евразийских и африканских канюков. Обыкновенный канюк — представитель рода Buteo , группы хищников среднего размера с крепким телом и широкими крыльями. Виды Buteo из Евразии и Африки обычно называют «канюками», а виды, обитающие в Америке, называют ястребами. Согласно современной классификации, род включает около 28 видов, что является вторым по разнообразию из всех существующих родов ястребиных после только Accipiter . [ 11 ] Анализ ДНК показывает, что обыкновенный канюк довольно тесно связан с краснохвостым ястребом ( Buteo jamaicensis ) Северной Америки , который занимает аналогичную экологическую нишу с канюком на этом континенте. Оба вида могут принадлежать к одному видовому комплексу . [ 11 ] [ 12 ] Три канюка в Африке, вероятно, тесно связаны с обыкновенным канюком на основании генетического материала: горный канюк ( Buteo oreophilus ), лесные канюки ( Buteo trizonatus ) и мадагаскарский канюк ( Buteo brachypterus ), до такой степени, что возникает вопрос, являются ли они достаточно различны, чтобы их можно было отнести к полноценным видам. Однако самобытность этих африканских канюков в целом получила поддержку. [ 12 ] [ 13 ] [ 14 ] Генетические исследования также показали, что современные канюки Евразии и Африки представляют собой относительно молодую группу, показывая, что они разошлись примерно 300 000 лет назад. [ 15 ] Тем не менее, окаменелости возрастом более 5 миллионов лет (поздний миоцен ) показали, что виды Buteo присутствовали в Европе гораздо раньше, чем это можно было бы предположить, хотя нельзя с уверенностью утверждать, что они были связаны с современными канюками. [ 16 ]
Разделение на подвиды и виды
[ редактировать ]
В прошлом было описано около 16 подвидов, и до 11 часто считаются действительными, хотя некоторые авторитеты допускают всего семь. [ 3 ] Подвиды обыкновенного канюка делятся на две группы.
Западная группа бутео живущих на короткие расстояния состоит в основном из постоянных жителей или мигрантов, , и включает в себя:
- Б. б. бутео : Ареалы в Европе от Атлантических островов, Британских островов и Пиренейского полуострова (включая остров Мадейра , население которого когда-то считалось отдельной расой, Б. б. harterti ) более или менее непрерывно по всей Европе до Финляндии , Румынии и Малой Азии . . [ 17 ] Эта весьма индивидуально изменчивая раса описана ниже. Это относительно крупный и громоздкий вид канюков. У самцов хорда крыла колеблется от 350 до 418 мм (от 13,8 до 16,5 дюйма), а длина хвоста - от 194 до 223 мм (от 7,6 до 8,8 дюйма). Для сравнения, более крупная самка имеет хорду крыла размером от 374 до 432 мм (от 14,7 до 17,0 дюйма) и длину хвоста от 193 до 236 мм (от 7,6 до 9,3 дюйма). У обоих полов предплюсны составляет от 69 до 83 мм (от 2,7 до 3,3 дюйма). длина [ 3 ] [ 5 ] Как показывает средняя масса тела, размеры номинальной расы обыкновенного канюка, по-видимому, соответствуют правилу Бергмана , увеличиваясь к северу и уменьшаясь ближе к экватору . [ 18 ] Сообщается, что на юге Норвегии средний вес самцов составлял 740 г (1,63 фунта), а женщин - 1100 г (2,4 фунта). [ 19 ] Британские канюки имели средний размер: 214 самцов в среднем весили 781 г (1,722 фунта) и 261 самка в среднем весили 969 г (2,136 фунта). [ 18 ] Птицы на юге Испании были меньше: в среднем 662 г (1,459 фунта) у 22 самцов и 800 г (1,8 фунта) у 30 самок. [ 20 ] Крэмп и Симмонс (1980) указали, что средняя масса тела канюков-номинантов в Европе в целом составляет 828 г (1,825 фунта) у самцов и 1052 г (2,319 фунта) у самок. [ 5 ]

- Б. б. Ротшильди : Эта предполагаемая раса обитает на Азорских островах. Обычно считается действительным подвидом. [ 3 ] [ 5 ] [ 21 ] Эта раса отличается от типичного промежуточного представителя номината тем, что имеет более темный и холодный коричневый цвет как сверху, так и снизу, ближе к более темным особям номината. В среднем он меньше, чем большинство канюков-номинантов. Хорда крыла самцов колеблется от 343 до 365 мм (от 13,5 до 14,4 дюйма), а у самок - от 362 до 393 мм (от 14,3 до 15,5 дюйма). [ 3 ] [ 21 ]
- Б. б. insularum : Эта раса обитает на Канарских островах . Не все авторитеты считают эту расу достаточно отдельной, но другие выступают за сохранение ее как полноценного подвида. [ 3 ] [ 21 ] Обычно он имеет более насыщенный коричневый цвет сверху и более густые прожилки снизу по сравнению с номинальными птицами. По размеру он похож на B. b. rothschildi и в среднем немного меньше номинальной расы. Сообщается, что у самцов хорда крыла составляет от 352 до 390 мм (от 13,9 до 15,4 дюйма), а у самок - от 370 до 394 мм (от 14,6 до 15,5 дюйма). [ 3 ]
- Б. б. arrigonii : Эта раса обитает на островах Корсика и Сардиния . Обычно считается действительным подвидом. [ 3 ] [ 21 ] Верхняя сторона этих канюков имеет промежуточный коричневый цвет с очень густыми полосами внизу, часто покрывающими живот, тогда как у большинства канюков-номинантов имеется беловатая область в середине живота. Как и большинство других островных рас, эта относительно невелика. У самцов хорда крыла составляет от 343 до 382 мм (от 13,5 до 15,0 дюйма), а у самок - от 353 до 390 мм (от 13,9 до 15,4 дюйма). [ 3 ] [ 22 ]

В группу восточных вульпинов входят:

- Б. б. vulpinus : Степной канюк гнездится на западе вплоть до восточной Швеции , на южных двух третях Финляндии , в восточной Эстонии , на большей части территории Белоруссии и Украины , на востоке до северного Кавказа , северного Казахстана , Кыргызстана , на большей части территории России до Алтая и на юге. Центральная Сибирь , Тянь-Шань в Китае и западная Монголия . Б. б. vulpinus — дальний мигрант . Зимует в основном на большей части восточной и южной Африки . Реже и зачастую весьма прерывисто степные канюки зимуют на южных полуостровах Европы, Аравии и юго-западной Индии , а также в некоторых частях юго-восточного Казахстана , Узбекистана и Киргизии . На открытой местности, излюбленной местами зимовок, степных канюков часто можно увидеть сидящими на придорожных телефонных столбах. [ 1 ] [ 3 ] [ 17 ] [ 23 ] Когда-то его считали отдельным видом из-за различий в размере, форме, окраске и поведении (особенно в отношении миграционного поведения), но генетически он не отличается от номинальных канюков. [ 5 ] [ 24 ] Более того, степной канюк активно скрещивается с номинальной расой, в результате чего типичные характеристики двух рас смешиваются. Зона интеграции простирается от Скандинавии через европейский континент до Черного моря , включая любую часть пересекающихся ареалов в Швеции , Финляндии , Эстонии , Латвии , Литве , западной Украине и восточной Румынии . Иногда плодовитые гибриды этих двух рас ошибочно относят к таким расам, как B. b. промежуточный или Б. б. циммерманные . серо-коричневый тип бледных морф vulpinus . Межсортные канюки наиболее распространены там, где преобладает [ 3 ] [ 25 ] Степные канюки обычно заметно меньше, с относительно более длинными для своего размера крыльями и хвостом и поэтому часто кажутся более быстрыми и проворными в полете, чем канюки-номинанты, чьи взмахи крыльев могут выглядеть медленнее и неуклюже. Обычно их длина составляет от 45 до 50 см (от 18 до 20 дюймов), в то время как размах крыльев самцов составляет в среднем 113 см (44 дюйма), а у самок - 122,7 см (48 дюймов). составляет Хорда крыла от 335 до 377 мм (от 13,2 до 14,8 дюйма) у самцов и от 358 до 397 мм (от 14,1 до 15,6 дюйма) у самок. Длина хвоста составляет от 170 до 200 мм (от 6,7 до 7,9 дюйма) у самцов и от 175 до 209 мм (от 6,9 до 8,2 дюйма) у самок. [ 3 ] [ 17 ] Сообщается, что вес птиц из России может варьироваться от 560 до 675 г (от 1,235 до 1,488 фунтов) у самцов и от 710 до 1180 г (от 1,57 до 2,60 фунтов) у самок. [ 26 ] Вес перелетных птиц оказывается ниже, чем у степных канюков в другое время года. Два исследования перелетных канюков во время их огромного весеннего перелета в Эйлате , Израиль, показали, что 420 птиц весили в среднем 579 г (1,276 фунта), а 882 птицы - в среднем 578 г (1,274 фунта). [ 27 ] [ 28 ] Для сравнения, вес зимующих степных канюков был выше: в среднем 725 г (1,598 фунта) у 35 птиц в бывшем Трансваале ( Южная Африка ) и 739 г (1,629 фунта) у 160 птиц в Капской провинции . Вес птиц из Замбии был схожим. [ 27 ] [ 29 ] [ 30 ]
- Б. б. menetriesi : Эта раса встречается в южном Крыму, через Кавказ , в северный Иран и, возможно, в Турцию . [ 3 ] [ 31 ] Эта раса традиционно считается постоянной расой, но некоторые источники считают ее мигрантом в восточную и южную Африку. [ 3 ] [ 32 ] По сравнению с перекрывающимся подвидом степного канюка, он крупнее (примерно промежуточный между номинальной расой и vulpinus ) и имеет более тусклый общий цвет, снизу песчаный, а не рыжий, и не имеет ярко-рыжего цвета на хвосте. Хорда крыла составляет от 351 до 397 мм (от 13,8 до 15,6 дюйма) у самцов и от 372 до 413 мм (от 14,6 до 16,3 дюйма) у самок. [ 3 ] [ 24 ]
Когда-то считалось, что расы обыкновенного канюка распространены в Азии так же далеко, как гнездящаяся птица, вплоть до Гималаев , и на востоке, вплоть до северо-восточного Китая , от России до Охотского моря и всех островов Курильских островов и Япония , несмотря на то, что и гималайские, и восточные птицы демонстрируют естественный разрыв в распространении от следующего ближайшего гнездящегося обыкновенного канюка. Однако тестирование ДНК показало, что канюки этих популяций, вероятно, принадлежат к разным видам. Большинство специалистов в настоящее время признают этих канюков полноценными видами: восточный канюк ( Buteo japonicus ; имеет три собственных подвида) и гималайский канюк ( Buteo refectus ). [ 24 ] [ 33 ] [ 34 ] Канюки, обитающие на островах Кабо-Верде у побережья Западной Африки, когда-то называвшиеся подвидом B. b. Bannermani и остров Сокотра на севере Аравийского полуострова , когда-то называемый редко признаваемым подвидом B. b. socotrae , в настоящее время считается не принадлежащим к обыкновенному канюку. Анализ ДНК показал, что эти островные канюки на самом деле более тесно связаны с длинноногим канюком ( Buteo rufinus ), чем с обыкновенным канюком. Впоследствии некоторые исследователи выступали за полный видовой статус популяции Кабо-Верде, но местонахождение этих канюков обычно считается неясным. [ 3 ] [ 21 ] [ 15 ] [ 35 ]
Описание
[ редактировать ]
Обыкновенный канюк — это хищник среднего и крупного размера, оперение которого сильно различается. У большинства канюков отчетливо круглая голова, несколько тонкий клюв, относительно длинные крылья, которые либо достигают кончика хвоста, либо немного не доходят до него, когда они сидят, довольно короткий хвост и несколько короткие и в основном голые лапки. В целом они могут казаться довольно компактными, но могут также казаться большими по сравнению с другими более распространенными хищными птицами, такими как пустельги и перепелятники . [ 3 ] [ 36 ] Обыкновенный канюк имеет длину от 40 до 58 см (от 16 до 23 дюймов) и размах крыльев 109–140 см (43–55 дюймов). [ 3 ] Самки в среднем примерно на 2–7% крупнее самцов по линейному закону и весят примерно на 15% больше. Масса тела может значительно варьироваться. Только у канюков из Великобритании вес самцов может варьироваться от 427 до 1183 г (от 0,941 до 2,608 фунта), а у самок - от 486 до 1370 г (от 1,071 до 3,020 фунта). [ 3 ] [ 37 ] [ 38 ]

В Европе большинство типичных канюков темно-коричневые сверху и на верхней стороне головы и спины, но могут стать более бледными и теплыми коричневыми с потертым оперением. всегда коричневые Маховые перья сидящих европейских канюков у номинального подвида ( B. b. Buteo ) . Обычно хвост обычно имеет серо-коричневый и темно-коричневый цвет с узкой полосой, бледным кончиком и широкой темной субтерминальной полосой, но у самых светлых птиц хвост может иметь различную белую и уменьшенную субтерминальную полосу или даже казаться почти полностью белым. У европейских канюков окраска нижней стороны может быть различной, но чаще всего это белое горло с коричневыми прожилками и несколько более темная грудь. Часто присутствует бледная буква U на груди; за ней следует бледная линия, идущая вниз по животу, которая разделяет темные области на груди и боках. Эти бледные области, как правило, имеют весьма изменчивые отметины, которые имеют тенденцию образовывать неправильные полосы. Молодые канюки очень похожи на взрослых особей номинальной расы, их лучше всего отличить по более бледному глазу, более узкой субтерминальной полосе на хвосте и отметкам на нижней стороне, которые выглядят как полосы, а не полосы. Кроме того, у молодых особей могут быть бахромки на верхних кроющих крыльях от кремовых до рыжих, но они также могут отсутствовать. Если смотреть снизу в полете, канюки в Европе обычно имеют темную заднюю кромку крыльев. Если смотреть сверху, одной из лучших примет является широкая темная субтерминальная полоса на хвосте. Маховые перья типичных европейских канюков в основном сероватые, с вышеупомянутой темной подкладкой крыльев спереди и контрастной более светлой полосой вдоль срединных кроющих. В полете у более светлых особей обычно появляются темные пятна на запястье, которые могут выглядеть как черноватые дуги или запятые, но они могут быть нечеткими у более темных особей или могут казаться светло-коричневатыми или блеклыми у более бледных особей. Молодых канюков-номинантов в полете лучше всего отличить от взрослых по отсутствию отчетливой субтерминальной полосы (вместо этого с довольно равномерной перемычкой по всей длине) и снизу по менее острой и коричневатой, а не черноватой задней кромке крыла. У молодых канюков видны более светлые полосы под крылом и телом, а не полоса, как у взрослых особей. [ 3 ] [ 36 ] [ 39 ] [ 40 ] Помимо типичного коричневатого канюка среднего размера, птицы в Европе могут варьироваться от почти однородного черно-коричневого цвета сверху до преимущественно белого цвета. Очень темные особи могут варьироваться от шоколадно-коричневого до черноватого, почти без бледности, но с изменчивой блеклой буквой U на груди и со слабыми более светлыми коричневыми полосами на горле или без них. Крайне бледные птицы в основном беловатые, с переменными широко расположенными полосами или светло-коричневыми наконечниками стрел в середине груди и на боках, а также могут иметь или не иметь темные центры перьев на голове, кроющих крыльях, а иногда и на всей мантии, кроме части мантии. Особи могут демонстрировать почти бесконечные вариации цветов и оттенков между этими крайностями, и по этой причине обыкновенный канюк считается одним из дневных хищников с самым разнообразным оперением. [ 3 ] [ 41 ] Одно исследование показало, что эта вариация на самом деле может быть результатом уменьшения генетического разнообразия одного локуса . [ 42 ]

Помимо номинальной формы ( B. b. Buteo ), которая занимает большую часть европейского ареала обыкновенного канюка, существует второй основной, широко распространенный подвид, известный как степной канюк ( B. b. vulpinus ). Раса степного канюка демонстрирует три основные цветовые морфы, каждая из которых может преобладать в определенном регионе гнездового ареала. Она более отчетливо полиморфна , чем просто индивидуально очень изменчива, как номинальная раса. Это может быть связано с тем, что, в отличие от канюка-номината, степной канюк является далеко мигрирующим. Полиморфизм связан с миграционным поведением. [ 3 ] [ 43 ] Самый распространенный вид степного канюка - рыжий морф, который дал этому подвиду научное название ( vulpes в переводе с латыни означает «лиса»). Эта морфа включает большинство птиц, замеченных при пролете к востоку от Средиземного моря . Рыжие морфные канюки имеют более светлый серо-коричневый цвет, чем у большинства номинальных B. b. бутео . По сравнению с номинальной расой, рыжий vulpinus имеет рисунок, не отличающийся от него, но, как правило, гораздо более рыжий на голове, бахроме на кроющих крыльях и, особенно, на хвосте и нижней стороне. Голова обычно серо-коричневая с рыжим оттенком, а хвост рыжий, от почти незаметного до тонкой темной полосы с субтерминальной полосой. Нижняя сторона может быть от равномерно бледной до темно-рыжей, с густой или легкой полосой с рыжей или с темной полосой, обычно с более темными особями, имеющими букву U, как у номинала, но с рыжим оттенком. Бледная форма степного канюка наиболее распространена на западе ареала своего подвида, преимущественно наблюдается зимой и во время миграции на различных сухопутных мостах Средиземноморья. Как и у рыжей морфы, у бледной морфы vulpinus сверху серо-коричневый, но хвост обычно отмечен тонкими темными полосами и субтерминальной полосой, рыжей только возле кончика. Нижняя сторона бледной морфы серовато-белая с темно-серо-коричневыми или несколько полосатыми от головы до груди и полосатыми животом и грудью, иногда с более темными боками, которые могут быть несколько рыжими. Темная морфа vulpinus , как правило, встречается на востоке и юго-востоке ареала подвида, и их численность легко превосходит рыжую морфу, при этом они в основном используют схожие точки миграции. Особи темной морфы варьируются от серо-коричневого до гораздо более темного черновато-коричневого цвета, имеют хвост темно-серого или несколько смешанного серо-рыжего цвета, с отчетливыми темными полосами и широкой черной субтерминальной полосой. У темной морфы vulpinus голова и нижняя сторона в основном однородные темные, от темно-коричневого до черновато-коричневого и почти чисто черного. Молодые особи рыжеватой морфы часто имеют заметно более бледный основной цвет (вплоть до кремово-серого), чем взрослые особи, с отчетливой перемычкой внизу, которая фактически увеличивается у молоди бледноморфного типа. Отличить молодь бледной и рыжей морфы друг от друга можно лишь в крайних случаях. Молодые особи темной морфы больше похожи на взрослых особей темной морфы. vulpinus , но часто имеют небольшие беловатые полосы внизу и, как и все другие расы, имеют более светлые глаза и более равномерную перемычку на хвосте, чем у взрослых особей. Степные канюки, как правило, кажутся меньшими и более проворными в полете, чем те, чьи взмахи крыльев могут выглядеть медленнее и неуклюже. В полете у рыжей morph vulpinus все тело и нижняя часть крыла варьируются от однородных до рыжих с рисунком (если рисунок присутствует, он варьируется, но может быть на груди и часто бедрах, иногда на боках, со светлой полосой поперек срединных кроющих), тогда как нижняя часть крыла хвост обычно более бледно-рыжий, чем верх. Беловатые маховые перья более заметны, чем у номинальных, и более заметно контрастируют с яркой темно-коричневой полосой вдоль задних краев. Отметины бледного вульпинуса , видимые в полете, похожи на рыжие (например, более светлые отметины на крыльях), но более сероватые как на крыльях, так и на теле. У темной морфы vulpinus широкие черные задние края и цвет тела заставляют беловатые участки внутреннего крыла еще больше выделяться с часто более ярким и черным пятном на запястье, чем у других морф. Как и в номинации, несовершеннолетний vulpinus (рыжий/светлый), как правило, имеет гораздо менее отчетливые задние края, общую полосатость по телу и вдоль срединных подкрыльевых кроющих. Темная морфа vulpinus больше напоминает взрослую особь в полете, чем другие морфы. [ 3 ] [ 39 ] [ 40 ] [ 25 ]
Похожие виды
[ редактировать ]
Обыкновенного канюка часто путают с другими хищниками, особенно в полете или на расстоянии. Неопытные и чрезмерно восторженные наблюдатели даже ошибочно принимали более темных птиц за гораздо более крупного беркута и иных пропорций ( Aquila chrysaetos ), а также темных птиц за западного болотного луня ( Circus aeruginosus ), который также летает двугранно, но, очевидно, относительно намного длиннее и стройнее. крылатые и хвостатые и с совершенно разными методами полета. Также канюков можно спутать с орлами-сапогами темной или светлой морфы ( Hieraeetus pennatus ), которые схожи по размеру, но орел летает ровно, крылья с параллельными краями, которые обычно кажутся шире, имеют более длинный, более квадратный хвост, без запястного пятна. у бледных птиц и все темные маховые, за исключением беловатого клина на внутренних маховых перьях у темных морф. Бледных особей иногда также путают с бледноморфными короткопалыми орлами ( Circaetus Gallicus ), которые намного крупнее, со значительно большей головой, более длинными крыльями (которые обычно держатся равномерно в полете, а не двугранными) и более светлым нижним крылом без какого-либо запястного пятна. или темная подкладка крыла. [ 3 ] [ 36 ] [ 39 ] Более серьезные проблемы с идентификацией возникают у других видов Buteo и у летающих медоедов , которые выглядят совсем по-другому, если их увидеть сидящими с близкого расстояния. Считается, что европейский осох ( Pernis apivorus ) имитирует более сильных хищников, в частности, молодые особи могут имитировать оперение более мощного обыкновенного канюка. [ 44 ] Хотя в Европе осоки менее индивидуально изменчивы, нижняя часть тела более полиморфна, чем даже у обыкновенного канюка. Наиболее распространенная форма взрослого европейского осоха имеет густую рыжую полосу на нижней стороне, что сильно отличается от обыкновенного канюка, однако коричневатая молодь гораздо больше напоминает промежуточного обыкновенного канюка. Осоёд машет крыльями заметно медленнее и равномернее, чем обыкновенный канюк. Крылья также поднимаются выше при каждом взмахе вверх, создавая более правильный и механический эффект, кроме того, при полете их крылья слегка выгнуты, но не в форме буквы V. У осоха голова кажется меньше, тело тоньше, хвост длиннее. и крылья более узкие и с более параллельными краями. Расу степных канюков особенно часто принимают за молодых европейских осоев до такой степени, что первые наблюдатели миграции хищников в Израиле считали отдаленных особей неотличимыми. Однако по сравнению со степным канюком у осоха заметно более темные второстепенные струйки на нижнем крыле с меньшим количеством и более широкими полосами и более обширными черными кончиками крыльев (целыми пальцами), контрастирующими с менее бледной рукой. [ 3 ] [ 36 ] [ 40 ] ), встречающийся в том же ареале, что и степной канюк, в некоторых частях южной Сибири , а также (с зимующими степями) на юго-западе Индии , Восточный осоед ( Pernis ptilorhynchus крупнее как европейского осося, так и обыкновенного канюка. Восточный вид по строению тела больше похож на обыкновенного канюка: у него относительно более широкие крылья, более короткий хвост и более широкая голова (хотя голова все еще относительно небольшая) по сравнению с европейским осохом, но во всем оперении отсутствуют запястные пятна. [ 3 ]
На большей части Европы канюк является единственным видом канюка. Тем не менее, субарктический гнездящийся канюк ( Buteo lagopus ) зимой занимает большую часть северной части континента в тех же местах, что и обыкновенный канюк. Тем не менее, шершавый канюк обычно крупнее, у него заметно более длинные крылья, оперенные ноги, а также хвост с белым основанием и широкой субтерминальной полосой. У шершавых канюков более медленные взмахи крыльев и они парят гораздо чаще, чем у обыкновенных канюков. Пятна на запястье на нижнем крыле также более жирные и черные у всех более светлых форм ястреба с грубыми ногами. У многих бледноморфных шершавых канюков есть яркая черноватая полоса на животе на фоне контрастирующих более светлых перьев - особенность, которая редко появляется у отдельных обыкновенных канюков. Обычно у большинства бледных форм мохоногих канюков лицо также кажется несколько беловатым, что справедливо только для чрезвычайно бледных обыкновенных канюков. Грубоногие канюки темной морфы обычно заметно темнее (вплоть до почти черноватого цвета), чем даже очень темные особи обыкновенных канюков в Европе, и по-прежнему имеют отчетливый хвост с белым основанием и широкую субтерминальную полосу, как у других шерстоногих канюков. [ 3 ] [ 36 ] [ 40 ] В Восточной Европе и на большей части азиатского ареала обыкновенного канюка длинноногий канюк ( Buteo rufinus рядом с обычным видом может жить ). Как и у степных канюков, у длинноногого канюка есть три основные цветовые морфы, более или менее схожие по оттенку. Как у степного канюка, так и у длинноногого канюка основная окраска в целом довольно рыжая. В большей степени, чем степные канюки, длинноногие канюки, как правило, имеют заметно более светлые голову и шею по сравнению с другими перьями и, что более отчетливо, обычно хвост без перемычек. Кроме того, длинноногий канюк обычно представляет собой довольно крупную птицу, которую часто считают внешне напоминающей орла (хотя он действительно кажется изящным и мелкоклювым даже по сравнению с более мелкими настоящими орлами), эффект усиливается за счет его более длинных лапок, несколько длиннее шея и относительно удлиненные крылья. Стиль полета последнего вида более глубокий, медленный и более орлиный, с гораздо более частым зависанием, с более выступающей головой и немного более высоким V, удерживаемым при парении. Чем меньше Североафриканская и аравийская раса канюка ( B.r.cirtensis ) по размерам и почти всем цветовым характеристикам более сходна со степным канюком, вплоть до сильно пестрого оперения молоди, в ряде случаев таких птиц можно отличить только по окраске. пропорции и схемы полета остаются неизменными. [ 3 ] [ 40 ] Гибридизация с последней расой ( B.r.cirtensis ) и канюками-номинантами наблюдалась в Гибралтарском проливе , несколько таких птиц потенциально были зарегистрированы в южном Средиземноморье из-за взаимно сужающихся ареалов, которые, возможно, размыты из-за климата. изменять. [ 45 ]
Зимующие степные канюки могут жить рядом с горными канюками и особенно с лесными канюками во время зимовки в Африке. Молодь степного и лесного канюка более или менее неразличима и различается только пропорциями и стилем полета, причем последний вид мельче, компактнее, имеет меньший клюв, более короткие ноги и более короткие и тонкие крылья, чем у степного канюка. Однако размер не является диагностическим, если только они не расположены рядом, поскольку в этом отношении два канюка перекрывают друг друга. Наиболее достоверными являются пропорции крыльев видов и их полетные действия. У лесного канюка более гибкие взмахи крыльев, чередующиеся с планированием, к тому же он парит на более плоских крыльях и, по-видимому, никогда не парит. Взрослые лесные канюки по сравнению с типичным взрослым степным канюком (рыжая морфа) также похожи, но лесной тип обычно имеет более белую нижнюю сторону, иногда в основном просто-белую, обычно с тяжелыми пятнами или каплевидными отметинами на брюшке, с полосой на бедрах, более узкая каплевидная форма на груди и более пятнистая на передних краях нижнего крыла, обычно без отметин на белой букве U на груди (которая в остальном похожа, но обычно шире, чем у вульпинус ). Для сравнения, горный канюк, который по размеру больше похож на степного канюка и немного крупнее лесного канюка, обычно имеет более тускло-коричневый цвет сверху, чем степной канюк, и более беловатый снизу с характерными тяжелыми коричневыми пятнами от груди до живота. бока и подкладки крыльев, тогда как молодые канюки снизу охристые, с более мелкими и полосатыми отметинами. Степной канюк по сравнению с другим африканским видом, канюком-красношейкой ( Buteo auguralis ), имеющим красный хвост, похожий на хвост vulpinus , отличается во всех других аспектах оперения, несмотря на схожий размер. У последнего канюка полосатая рыжая голова, белая нижняя часть с контрастирующей яркой темной грудью во взрослом оперении, а в молодом оперении он имеет тяжелые темные пятна на груди и боках со светлой подкладкой крыльев. Шакалы и канюки-авгуры ( Buteo rufofuscus & augur ), также рыжие на хвосте, крупнее и крупнее степных канюков и имеют несколько отличительных характеристик оперения, в первую очередь оба имеют свои собственные яркие, контрастирующие узоры черно-коричневого, рыжего и кремового цвета. . [ 3 ] [ 36 ] [ 46 ]
Распространение и среда обитания
[ редактировать ]
Обыкновенный канюк встречается на нескольких островах восточной части Атлантического океана, включая Канарские острова и Азорские острова , а также почти по всей Европе. Сегодня он встречается в Ирландии и почти во всех частях Шотландии , Уэльса и Англии . Примечательно, что в материковой Европе нет существенных пробелов без размножения обыкновенных канюков от Португалии и Испании до Греции , Эстонии , Беларуси и Украины , хотя на большей части восточной половины последних трех стран они присутствуют в основном только в период размножения. Они также присутствуют на всех крупных средиземноморских островах, таких как Корсика , Сардиния , Сицилия и Крит . Дальше на север Скандинавии они встречаются в основном на юго-востоке Норвегии (хотя также в некоторых точках на юго-западе Норвегии недалеко от побережья и в одной части к северу от Тронхейма ), чуть выше южной половины Швеции и граничат с Ботническим заливом до Финляндии , где они живут как размножающийся вид почти на двух третях суши.
Обыкновенный канюк достигает своих северных пределов гнездования на Дальнем Востоке Финляндии и за границей с европейской частью России , продолжая гнездиться до самых узких проливов Белого моря и почти до Кольского полуострова . В этих северных районах обыкновенный канюк обычно обитает только летом, но круглый год обитает в значительной части южной Швеции и некоторых частях южной Норвегии. [ 1 ] [ 3 ] [ 6 ] За пределами Европы он обитает в северной Турции (в основном недалеко от Черного моря ), в других случаях встречается главным образом в качестве транзитной миграции или зимнего гостя в остальной части Турции, Грузии , спорадически, но не редко, в Азербайджане и Армении , северном Иране (в основном обнимая Каспийское море ) до северного Туркменистана . [ 1 ] Дальше на север, хотя он отсутствует по обе стороны северного Каспийского моря, обыкновенный канюк встречается на большей части западной России (хотя исключительно в качестве гнездящегося места), включая весь Центральный федеральный округ и Приволжский федеральный округ , все, кроме самых северных частей Северо -Западный и Уральский федеральные округа и почти южная половина Сибирского федерального округа , самое дальнее восточное распространение как селекционер. Он также встречается в северном Казахстане , Кыргызстане , на крайнем северо-западе Китая ( Тянь-Шань ) и северо-западной Монголии . [ 1 ] [ 47 ]
Негнездящиеся популяции встречаются либо в качестве мигрантов, либо в виде зимующих птиц на юго-западе Индии , в Израиле , Ливане , Сирии , Египте (северо-восток), северном Тунисе (и на крайнем северо-западе Алжира ), северном Марокко , вблизи побережий Гамбии , Сенегала и дальних стран. юго-запад Мавритании и Кот-д’Ивуара (и граничит с Буркина-Фасо ). В восточной и центральной Африке встречается зимой на юго-востоке Судана , Эритрее , примерно на двух третях Эфиопии , [ 48 ] большая часть Кении (хотя, очевидно, отсутствует на северо-востоке и северо-западе), Уганда , южная и восточная Демократическая Республика Конго и более или менее вся южная Африка от Анголы до Танзании и по остальной части континента (за исключением очевидного разрыв вдоль побережья от юго-западной Анголы до северо-западной Южной Африки ). [ 1 ] [ 49 ] [ 50 ]
среда обитания
[ редактировать ]Обыкновенный канюк обычно обитает на границе лесных массивов и открытых территорий ; Чаще всего этот вид обитает на опушках леса , в небольших лесах или лесополосах с прилегающими лугами , пахотными землями или другими сельскохозяйственными угодьями . Открытая вересковая пустошь оправдана , если там есть деревья для охоты и гнездования окуней. Леса, в которых они обитают, могут быть хвойными , широколиственными и смешанными лесами умеренного пояса , а также лиственными лесами умеренного пояса , иногда с предпочтением местных доминирующих деревьев. Он отсутствует в безлесной тундре , а также в Субарктике , где вид почти полностью уступает место зимняку. [ 3 ] [ 17 ] Обыкновенный канюк в безлесной степи встречается спорадически или редко , но может время от времени мигрировать через нее (несмотря на название, подвид степного канюка гнездится преимущественно в лесистых опушках степи). [ 51 ] Этот вид в некоторой степени встречается как в горной , так и в равнинной местности . Несмотря на то, что канюки адаптируются к водно-болотным угодьям и прибрежным районам и иногда встречаются на них, их часто считают более горным видом, и, по-видимому, их не привлекают регулярно водоемы или они категорически избегают их в немигрирующие времена. [ 3 ] [ 17 ] [ 52 ] Канюки в густолесистых районах восточной Польши в основном использовали большие, зрелые насаждения деревьев , которые были более влажными , богатыми и густыми, чем преобладающие в прилегающих районах, но отдавали предпочтение тем, которые находились на расстоянии от 30 до 90 м (от 98 до 295 футов) от отверстий. [ 53 ] В основном оседлые канюки живут в низинах и предгорьях , но они могут жить на лесистых хребтах и возвышенностях, а также на скалистых побережьях , иногда гнездясь на уступах скал, а не на деревьях. Канюки могут жить от уровня моря до высоты 2000 м (6600 футов), размножаясь в основном ниже 1000 м (3300 футов), но они могут зимовать на высоте 2500 м (8200 футов) и легко мигрируют до 4500 м (14 800 футов). [ 3 ] В горных итальянских Апеннинах гнезда канюков находились на средней высоте 1399 м (4590 футов) и находились, по отношению к окружающей местности, дальше от освоенных человеком территорий (например, дорог) и ближе к днищу долин в труднопроходимых местах с неравномерным топографированием. особенно те, которые смотрели на северо-восток. [ 54 ] Обыкновенные канюки довольно хорошо адаптируются к сельскохозяйственным угодьям , но могут продемонстрировать региональное снижение очевидной реакции на сельское хозяйство. Было показано, что изменения в более экстенсивных методах ведения сельского хозяйства привели к сокращению популяций канюков на западе Франции , где сокращение « живых изгородей , лесных участков и лугов » привело к сокращению численности канюков, а также в Хэмпшире , Англия на свободном выгуле , где более обширный выпас скота и лошадей привел к снижению численности канюков. канюков, вероятно, во многом из-за кажущегося сокращения популяций мелких млекопитающих . [ 55 ] [ 56 ] Напротив, канюки в центральной Польше адаптировались к вырубке сосен и сокращению добычи грызунов, меняя места гнездования и добычу на какое-то время без сильных изменений в их местной численности. [ 57 ] Обширная урбанизация, по-видимому, отрицательно влияет на канюков, поскольку этот вид, как правило, менее адаптируется к городским районам, чем его собратья из Нового Света, краснохвостый ястреб . Хотя пригородные районы могут время от времени увеличивать потенциальную популяцию добычи в определенном месте, смертность отдельных канюков, нарушение гнездования и деградация среды обитания на местах гнездования. в таких районах значительно возрастает [ 58 ] [ 59 ] [ 60 ] Обыкновенные канюки довольно хорошо адаптируются к сельской местности , а также к пригородным зонам с парками и большими садами , а также к таким местам, если они находятся рядом с фермами. [ 61 ] [ 62 ] [ 63 ]
Поведение
[ редактировать ]
Обыкновенный канюк является типичным бутео по большей части своего поведения . Чаще всего его можно увидеть либо парящим на разной высоте, либо сидящим на видном месте на верхушках деревьев, голых ветвях, телеграфных столбах, столбах забора, камнях или уступах, либо, попеременно, глубоко внутри крон деревьев. Канюки также будут стоять и добывать корм на земле. В постоянных популяциях он может проводить более половины дня в неактивном положении. Более того, его описывают как «медленную и не очень смелую» хищную птицу. [ 17 ] [ 64 ] Это одаренный парящий человек, который может подниматься в воздух в течение длительного времени, но может показаться трудоемким и тяжелым в горизонтальном полете, в большей степени канюков, чем степных канюков. [ 3 ] [ 17 ] В частности, во время миграции, как было зафиксировано в случае перемещения степных канюков над Израилем , канюки легко меняют направление, расположение хвоста и крыльев, а также высоту полета, чтобы приспособиться к окружающей среде и ветровым условиям. [ 65 ] В Израиле канюки-мигранты редко взлетают на такую высоту (максимум 1000–2000 м (3300–6600 футов) над землей) из-за отсутствия горных хребтов, которые в других районах обычно служат пролетными путями; однако попутный ветер значителен и позволяет птицам преодолевать скорость в среднем 9,8 метра в секунду (22 мили в час). [ 66 ]
Миграция
[ редактировать ]
Обыкновенный канюк удачно описывается как частичный мигрант. [ 67 ] Осенние и весенние перемещения канюков подвержены значительным вариациям, вплоть до индивидуального уровня, в зависимости от пищевых ресурсов региона, конкуренции (как со стороны других канюков, так и других хищников), степени беспокойства человека и погодных условий. Передвижения на короткие расстояния являются нормой для подростков и некоторых взрослых осенью и зимой, но в Центральной Европе и на Британских островах больше взрослых особей остаются на своих круглогодичных местах жительства, чем нет. [ 3 ] [ 68 ] Даже в случае расселения молоди канюков первого года жизни они могут не зайти слишком далеко. В Англии 96% первокурсников зимой переехали на расстояние менее 100 км (62 миль) от места своего рождения. [ 69 ] Юго-западная Польша была зарегистрирована как довольно важное место зимовки среднеевропейских канюков ранней весной, которые, по-видимому, пришли с несколько севера, а зимой средняя плотность составляла локально высокие 2,12 особи на квадратный километр. [ 68 ] Доступность среды обитания и добычи, по-видимому, была основными факторами выбора среды обитания осенью для европейских канюков. В северной Германии было зарегистрировано, что канюки осенью отдают предпочтение участкам, достаточно удаленным от места гнездования, с большим количеством нор полевок и более широко рассредоточенными насестами. [ 70 ] [ 71 ] В Болгарии средняя плотность зимовки составляла 0,34 особи на квадратный километр, и канюки отдавали предпочтение сельскохозяйственным территориям, а не лесным. [ 72 ] Подобные предпочтения среды обитания были зарегистрированы на северо-востоке Румынии , где плотность канюка составляла 0,334–0,539 особей на квадратный километр. [ 73 ] Номинативные канюки Скандинавии несколько более мигрируют, чем большинство популяций Центральной Европы. Однако птицы из Швеции демонстрируют некоторые различия в миграционном поведении. Максимум 41 000 особей было зарегистрировано в одном из основных мест миграции на юге Швеции в Фальстербо . [ 74 ] На юге Швеции зимние перемещения и миграции изучались путем наблюдения за окраской канюка. Белые особи были значительно чаще распространены на юге Швеции, а не дальше на север их шведского ареала. Южная популяция мигрирует раньше, чем средне-темные канюки, как у взрослых, так и у молодых особей. В южной популяции мигрирует большая доля молоди, чем взрослых. Особенно взрослые особи южной популяции являются резидентами в большей степени, чем заводчики более северных особей. [ 75 ]
Вся популяция степного канюка сильно мигрирующая, преодолевая за время миграции значительные расстояния. Ни на одной части ареала степные канюки не используют одни и те же места летования и зимовки. Степные канюки во время миграции немного стадны и путешествуют стаями разного размера. Эта раса мигрирует с сентября по октябрь, часто из Малой Азии до мыса Африки примерно за месяц, но не пересекает воду, следуя вокруг залива Винам озера Виктория, а не пересекая залив шириной в несколько километров. Точно так же они пройдут по обе стороны Черного моря . Миграционное поведение степных канюков повторяет поведение ширококрылых ястребов и ястребов Свенсона ( Buteo platypterus & swainsoni во всех отношениях ), что и аналогичные мигрирующие на большие расстояния Buteos , включая трансэкваториальные перемещения, избегание больших водоемов и стадное поведение. [ 4 ] [ 17 ] Мигрирующие степные канюки поднимаются вверх с утренними термальными потоками и могут преодолевать в среднем сотни миль в день, используя доступные течения вдоль горных хребтов и другие топографические особенности. Пик весенней миграции степных канюков приходится на март – апрель, но последние вульпины прибывают в места гнездования к концу апреля или началу мая. Расстояния, преодолеваемые мигрирующими степными канюками в одну сторону из северной Европы (т. е. из Финляндии или Швеции ) в южную Африку, составляют более 13 000 км (8 100 миль) за сезон. [ 3 ] [ 17 ] Для степных канюков из восточной и северной Европы и западной России (которые составляют большую часть всех степных канюков) пиковая численность миграций приходится на разные районы осенью, когда наибольшие зарегистрированные перемещения происходят через Малую Азию , например , через Турцию , чем весной. когда самое большое зарегистрированное перемещение происходит на юг Ближнего Востока , особенно в Израиль . Эти два миграционных движения в целом практически не различаются, пока не достигают Ближнего Востока и Восточной Африки, где наибольший объем мигрантов осенью приходится на южную часть Красного моря , вокруг Джибути и Йемена , тогда как основной объем весной приходится на самую северную часть. пролив, вокруг Египта и Израиля . Осенью численность степных канюков, зарегистрированных на пролете, достигает 32 000 (учет 1971 г.) в северо-западной Турции ( Босфор ) и в северо-восточной Турции (Черное море) до 205 000 (учет 1976 г.). Дальше по пути миграции в Джибути было зарегистрировано осеннее количество пролетов до 98 000 особей . [ 3 ] [ 76 ] [ 77 ] Было зарегистрировано от 150 000 до почти 466 000 степных канюков, мигрирующих через Израиль весной, что делает это не только самым многочисленным мигрирующим хищником здесь, но и одним из крупнейших миграций хищников в мире. [ 66 ] [ 78 ] Миграционные перемещения канюков южной Африки в основном происходят вдоль крупных горных хребтов, таких как Драконовы горы и горы Лебомбо . [ 46 ] гораздо более нерегулярно, встречаются в Трансваале чем в Капской области . Зимующие степные канюки зимой [ 79 ] Начало миграционного движения степных канюков обратно к местам размножения на юге Африки приходится преимущественно на март, достигая пика на второй неделе. [ 46 ] Степной канюк быстро линяет перья по прибытии на места зимовки и, по-видимому, разделяет линьку маховых перьев между местом размножения в Евразии и местом зимовки на юге Африки, при этом линька приостанавливается во время миграции. [ 80 ] За последние 50 лет было зарегистрировано, что канюки-номинанты обычно мигрируют на более короткие расстояния и зимуют дальше на север, возможно, в ответ на изменение климата, в результате чего их численность в местах миграции относительно меньше. Они также расширяют ареал своего размножения, возможно, сокращая или вытесняя степных канюков. [ 75 ] [ 81 ] [ 82 ]
Вокализации
[ редактировать ]Постоянные популяции канюков, как правило, издают звуки круглый год, тогда как мигранты, как правило, издают звуки только в период размножения. И канюки-номинанты, и степные канюки (и их многочисленные родственные подвиды внутри своих типов), как правило, имеют схожие голоса. Основной крик этого вида - жалобное, далеко разносящееся пи-йау или пи-и-у , используемое как в качестве контактного крика, так и более возбужденно в воздушных демонстрациях. Их крик более резкий, более звонкий, когда он используется в агрессии, имеет тенденцию быть более продолжительным и колеблющимся, когда преследует злоумышленников, более резким и более визгливым, когда он используется в качестве предупреждения при приближении к гнезду, или более коротким и более взрывным, когда он вызывает тревогу. Другие варианты их вокального исполнения включают кошачье мяуканье , неоднократно произносимое на крыле или сидя, особенно напоказ; Было зарегистрировано повторное мах , произнесенное парами, отвечающими друг другу, в гнездах также были зарегистрированы дальнейшие смешки и каркания. [ 3 ] [ 17 ] [ 83 ] Молодь обычно можно отличить по нестройному характеру криков по сравнению с криками взрослых. [ 84 ]
Диетическая биология
[ редактировать ]Обыкновенный канюк — универсальный хищник, который при возможности охотится на самую разнообразную добычу. Спектр их добычи простирается на самых разных позвоночных, включая млекопитающих , птиц (от любого возраста от яиц до взрослых птиц), рептилий , земноводных и, реже, рыб , а также различных беспозвоночных , в основном насекомых . Часто подвергаются нападениям молодые животные, в основном гнездящиеся детеныши различных позвоночных. Всего известно более 300 видов добычи обыкновенных канюков. Кроме того, размер добычи может варьироваться от крошечных жуков , гусениц и муравьев до крупных взрослых тетеревов и кроликов , которые почти вдвое превышают массу тела. [ 3 ] [ 17 ] [ 85 ] [ 86 ] [ 87 ] оценивалась в 179,6 г (6,34 унции) Средняя масса тела позвоночных жертв в Беларуси . [ 88 ] Иногда они также частично питаются падалью, обычно мертвыми млекопитающими или рыбой. [ 3 ] Однако диетические исследования показали, что они в основном охотятся на мелких млекопитающих , в основном на мелких грызунов . [ 48 ] Как и многие хищные птицы умеренного пояса различных родословных, полевки являются неотъемлемой частью рациона канюка. Предпочитание этой птицы граничить между лесом и открытыми пространствами часто делает ее идеальной средой обитания для полевок. [ 89 ] [ 90 ] Было обнаружено, что охота на относительно открытых территориях увеличивает успех охоты, тогда как более полный кустарниковый покров снижает успех. [ 91 ] Большую часть добычи добывают, сбрасывая с окуня, и обычно добывают на земле. Альтернативно, на добычу можно охотиться на низкой высоте. Этот вид не склонен охотиться, эффектно наклоняясь, а обычно плавно падает, а затем постепенно ускоряется внизу, держа крылья над спиной. Иногда канюк также добывает пищу, случайным образом планируя или паря над открытой местностью, опушками леса или полянами. Предпочтительно охотиться на окуня, но канюки довольно регулярно также охотятся с земли, когда этого требует среда обитания. Вне сезона размножения было зарегистрировано 15–30 канюков, добывающих корм на земле на одном большом поле, особенно молодые особи. Обычно самый редкий тип добывания пищи — парящий. Исследование, проведенное в Великобритании, показало, что зависание, похоже, не увеличивает успех охоты. [ 3 ] [ 17 ] [ 92 ]
Млекопитающие
[ редактировать ]
Если есть такая возможность, можно привлечь большое разнообразие грызунов , поскольку в пище обыкновенных канюков зарегистрировано около 60 видов грызунов. [ 85 ] [ 86 ] Кажется очевидным, что полевки являются наиболее важной добычей европейских канюков. Почти в каждом исследовании на континенте упоминается важность, в частности, двух наиболее многочисленных и широко распространенных европейских полевок: обыкновенной полевки весом 28,5 г (1,01 унции) ( Microtus arvalis ) и несколько более северной полевки массой 40 г (1,4 унции). ) полевка ( Microtus agrestis ). [ 93 ] [ 94 ] [ 95 ] На юге Шотландии полевки были наиболее представленными видами в погадках, на их долю приходилось 32,1% из 581 погадки. [ 96 ] [ 97 ] На юге Норвегии полевки снова стали основным кормом в годы пиковой численности полевок: на их долю приходилось 40,8% из 179 объектов добычи в 1985 году и 24,7% из 332 объектов добычи в 1994 году. В общей сложности грызуны составляют 67,6% и 58,4% продуктов питания в эти пиковые годы для полевок. Однако в годы с низкой численностью полевок вклад грызунов в рацион был незначительным. [ 94 ] Даже на западе, в Нидерландах , обыкновенные полевки были наиболее регулярной добычей, составляя 19,6% из 6624 объектов добычи в очень большом исследовании. [ 98 ] Обыкновенные полевки были основным кормом, зарегистрированным в центральной Словакии , и составляли 26,5% из 606 объектов добычи. [ 99 ] Обыкновенная полевка или другие родственные виды полевок иногда были основным кормом и на Украине (17,2% от 146 объектов добычи), а также на востоке России в Приволжском степном заповеднике (41,8% от 74 объектов добычи) и в Самаре ( 21,4% из 183 объектов добычи). [ 85 ] Другие записи из России и Украины показывают, что полевки варьируются от второстепенной добычи до 42,2% рациона. [ 100 ] В Беларуси на полевок, включая виды Microtus и рыжих полевок ( Myodes glareolus ) массой 18,4 г (0,65 унции), приходилось в среднем 34,8% биомассы в 1065 объектах добычи из разных районов исследований за 4 года. [ 88 ] Известно , что по крайней мере 12 видов рода Microtus являются объектами охоты обыкновенных канюков, и даже это, вероятно, консервативно, более того, аналогичные виды, такие как лемминги . если таковые имеются, будут выбраны [ 85 ] [ 87 ] [ 99 ] [ 100 ] [ 101 ]
Других грызунов ловят в основном оппортунистически, а не по предпочтениям. Известно, что несколько лесных мышей ( Apodemus ssp. ) добываются довольно часто, но, учитывая их предпочтение активности в более глубоких лесах, чем предпочтительные границы между полем и лесом, они редко являются чем-то большим, чем второстепенным продуктом питания. [ 93 ] [ 94 ] Исключением была Самара , где желтошейная мышь ( Apodemus flavicollis ), одна из крупнейших представителей своего рода с весом 28,4 г (1,00 унции), составляла 20,9%, уступая по значимости обыкновенной полевке. [ 85 ] Точно так же древесные белки охотно поедаются, но редко играют важную роль в пище канюков в Европе, поскольку канюки, по-видимому, предпочитают избегать добычи с деревьев и не обладают ловкостью, обычно необходимой для поимки значительного количества древесных белок. [ 17 ] [ 86 ] [ 102 ] [ 103 ] Все четыре суслика , обитающие (в основном) в Восточной Европе, также известны как добыча обыкновенного канюка, но мало количественного анализа было проведено, чтобы понять, насколько значимы такие отношения хищник-жертва. [ 104 ] [ 105 ] [ 106 ] [ 107 ] Размер добычи грызунов варьировался от евразийской промысловой мыши ( Micromys minutus ) весом 7,8 г (0,28 унции) до неместной ондатры ( Ondatra zibethicus ) весом 1100 г (2,4 фунта ). [ 108 ] Другие грызуны, добываемые либо редко, либо в районах, где пищевые привычки канюков известны лишь частично, включают летяг , сурков (вероятно, очень молодых, если их поймать живыми), бурундуков , колючих крыс , хомяков , землекопов , песчанок , песчанок и тушканчиков , а иногда и сытных. большое количество соней , хотя они ведут ночной образ жизни. [ 80 ] [ 88 ] [ 99 ] [ 100 ] [ 109 ] [ 110 ] [ 111 ] [ 112 ] [ 113 ] На удивление мало исследований было проведено в отношении рациона зимующих степных канюков на юге Африки, учитывая их многочисленный статус там. Однако было указано, что основной добычей являются грызуны, такие как четырехполосая травяная мышь ( Rabdomys pumilio ) и капский землекоп ( Georychus capensis ). [ 46 ] [ 114 ]
Помимо грызунов, две другие группы млекопитающих могут считаться важными для рациона канюков. Одним из этих основных видов добычи, вносимых в рацион обыкновенных канюков, являются лепориды или зайцеобразные , особенно европейский кролик ( Oryctolagus cuniculus ), где он встречается в большом количестве в диком или одичавшем состоянии. Во всех диетических исследованиях, проведенных в Шотландии, кролики играли очень важную роль в рационе канюка. В южной Шотландии кролики составляли 40,8% останков в гнездах и 21,6% содержимого погадок, в то время как зайцеобразные (в основном кролики, но также и некоторые молодые зайцы ) присутствовали в 99% останков в Морей , Шотландия . Богатство питательных веществ по сравнению с наиболее распространенной добычей в других местах, такой как полевки, может объяснить высокую продуктивность канюков здесь. Например, размеры кладок были в среднем вдвое больше там, где кролики были обычным явлением (Море), чем там, где они были редки ( Глен Уркхарт ). [ 96 ] [ 115 ] [ 116 ] В северной Ирландии , представляющей интерес, поскольку здесь отсутствуют какие-либо местные виды полевок , кролики снова стали основной добычей. Здесь зайцеобразные составляли 22,5% кормовых объектов по численности и 43,7% по биомассе. [ 117 ] Хотя кролики не являются местными, хотя и давно обосновались на Британских островах, в их родной части Пиренейского полуострова , кролики также играют важную роль в рационе канюков. В Мурсии , Испания , кролики были наиболее распространенным млекопитающим в рационе, составляя 16,8% из 167 объектов добычи. [ 87 ] В крупном исследовании, проведенном на северо-востоке Испании, кролики доминировали в пище канюка, составляя 66,5% из 598 объектов добычи. [ 118 ] В Нидерландах европейские кролики были вторыми по численности (19,1% из 6624 объектов добычи) после обыкновенных полевок и крупнейшим поставщиком биомассы в гнезда (36,7%). [ 98 ] За пределами этих (по крайней мере, исторически) богатых кроликами территорий зайчата обыкновенного зайца, обитающие в Европе, могут быть важной дополнительной добычей. Заяц-русак ( Lepus europaeus ) был четвертым по значимости видом добычи в центральной Польше и третьим по значимости видом добычи в Ставропольском крае , Россия . [ 100 ] [ 111 ] Канюки обычно нападают на молодняк европейских кроликов и зайцев. Вес большинства кроликов, пойманных канюком, по разным оценкам составлял от 159 до 550 г (от 5,6 до 19,4 унций), а иногда и до 700 г (1,5 фунта). [ 98 ] [ 118 ] [ 117 ] [ 115 ] [ 119 ] Аналогично, в разных регионах средний вес зайцев-русаков, добытых в Финляндии, составлял около 500 г (1,1 фунта). [ 120 ] Вес одного молодого зайца-беляка ( Lepus timidus ), пойманного в Норвегии, оценивался примерно в 1000 г (2,2 фунта). [ 86 ] Однако известно, что канюки иногда убивают взрослых кроликов. [ 119 ] Подтверждением этому могут служить остатки относительно крупных костей предплюсны кролика, длиной до 64 мм. [ 118 ] [ 121 ]
Другим важным типом добычи млекопитающих являются насекомоядные , среди которых, как известно, этот вид поедает более 20 видов, включая почти все виды землероек , кротов и ежей, обитающих в Европе. Среди этого отряда особенно часто попадаются кроты, поскольку, как и в случае с «полевками», канюки, вероятно, склонны наблюдать за кротовьими норами в полях на предмет активности и быстро нырять со своего насеста, когда одно из подземных млекопитающих выскакивает. [ 122 ] Наиболее широко встречающийся крот в северном ареале канюка - это европейский крот весом 98 г (3,5 унции) ( Talpa europaea ), и это один из наиболее важных объектов добычи этого вида, не являющихся грызунами. Этот вид присутствовал в 55% из 101 останков в Глен-Уркхарте , Шотландия , и был вторым по распространенности видом добычи (18,6%) в 606 объектах добычи в Словакии . [ 99 ] [ 115 ] В Бари , Италия , римский крот ( Talpa romana ), размером с европейский вид, был основной идентифицированной добычей среди млекопитающих, составляя 10,7% рациона. [ 123 ] Канюки могут поймать полноразмерный диапазон насекомоядных: от самого маленького млекопитающего в мире (по весу), этрусской землеройки весом 1,8 г (0,063 унции) ( Suncus etruscus ), до, возможно, самого тяжелого насекомоядного животного весом 800 г (28 унций). Европейский еж ( Erinaceus europaeus ). [ 115 ] [ 118 ] Добычей млекопитающих канюков, кроме грызунов, насекомоядных и зайцеобразных, добывают редко. Время от времени ласки , такие как ласка ( Mustela nivalis ) и горностай ( Mustela erminea ловятся останки молодых лесных куниц ( Martes martes ) и взрослых европейских хорьков ( Mustela putorius ). ), а в гнезде канюка обнаруживаются [ 94 ] [ 98 ] [ 100 ] [ 96 ] [ 124 ] [ 125 ] Многочисленные крупные млекопитающие, в том числе плотоядные животные среднего размера, такие как собаки , кошки , лисы и различные копытные , иногда поедаются канюками как падаль, главным образом в ненастные зимние месяцы. мертворожденные олени . Нередко посещаются и [ 88 ] [ 115 ] [ 107 ] [ 126 ]
Птицы
[ редактировать ]
Нападая на птиц, канюки в основном охотятся на птенцов и птенцов птиц малого и среднего размера, в основном воробьиных , но также и на различных диких птиц , но иногда также на раненых, больных или неосторожных, но здоровых взрослых особей. Несмотря на то, что обыкновенный канюк способен одолеть птиц крупнее его самого, обычно считается, что ему не хватает ловкости, необходимой для поимки многих взрослых птиц, даже диких птиц, которые, предположительно, будут более слабыми летчиками, учитывая их относительно тяжелое тело и маленькие крылья. [ 3 ] [ 17 ] Однако количество молодых и молодых птиц, на которых охотятся, по сравнению со взрослыми особями, варьируется. Например, в итальянских Альпах 72% пойманных птиц были птенцами или недавно оперившимися птенцами, 19% — птенцами и 8% — взрослыми особями. [ 127 ] Напротив, на юге Шотландии , несмотря на то, что канюки ловили относительно крупную птицу, в основном тетерева ( Lagopus lagopus scotica ), по сообщениям, 87% пойманных птиц были взрослыми особями. [ 96 ] В целом, как и у многих хищных птиц, далеких от птицеловов, птицы представляют собой самую разнообразную группу в спектре добычи канюка ввиду огромной численности и разнообразия птиц, немногие хищники не охотятся на них хотя бы изредка. В рационе канюка обнаружено около 150 видов птиц. В целом, несмотря на многие принятые меры, птицы обычно занимают в рационе второстепенное положение после млекопитающих. [ 94 ] [ 98 ] [ 100 ] В северной Шотландии в пище канюков было довольно много птиц. Наиболее часто регистрируемой птичьей добычей, а также 2-м и 3-м по частоте видами добычи (после только полевок) в Глен-Уркхарте ) весом 23,9 г (0,84 унции) были зяблик ( Fringilla coelebs ) весом 18,4 г (0,65 унции ) и луговые коньки ( Anthus pratensis , канюки уничтожили 195 птенцов этих видов против всего лишь 90 взрослых. Это отличалось от мурены , где наиболее частой добычей птиц и вторым по частоте видом добычи после кролика был обыкновенный лесной голубь ( Columba palumbus ) весом 480 г (17 унций ), а канюки съедали в четыре раза больше взрослых особей по сравнению с птенцами. [ 115 ]
Птицы были основной пищей канюков в итальянских Альпах , где они составляли 46% рациона, тогда как млекопитающие составляли 29% в 146 объектах добычи. Основными видами добычи здесь были евразийские черные дрозды массой 103 г (3,6 унции) ( Turdus merula массой 160 г (5,6 унции) ) и евразийские сойки ( Garrulus Glandarius ), хотя и у тех, и у других были пойманы в основном птенцы. [ 127 ] ведущее положение могли также занять птицы В годы с низкой численностью полевок на юге Норвегии , в частности дрозды , а именно черный дрозд, певчий дрозд массой 67,7 г (2,39 унции) ( Turdus philomelos 61 г (2,2 унции) ) и краснокрылка массой ( Turdus iliacus ). ), что в совокупности составляло 22,1% от 244 объектов добычи в 1993 году. [ 94 ] На юге Испании количество птиц в рационе было равно количеству млекопитающих, и то и другое - 38,3%, но большинство останков были классифицированы как «неопознанные птицы среднего размера», хотя наиболее часто идентифицируемыми видами из тех, которые, по-видимому, можно было определить, были евразийские сойки. и красноногие куропатки ( Alectoris rufa ). [ 87 ] Точно так же в Северной Ирландии птицы по импорту были примерно равны млекопитающим, но большинство из них были неопознанными врановыми . [ 117 ] В Северском Донце ( Украина ) птицы и млекопитающие составляют 39,3% рациона канюков. [ 100 ] Обыкновенные канюки могут охотиться почти на 80 видов воробьиных и почти на всех доступных диких птиц. Как и многие другие крупные хищники, дикие птицы привлекательны для охоты на канюков из-за их наземного образа жизни. Канюки были наиболее частыми хищниками при исследовании молоди фазанов в Англии, на их долю пришлось 4,3% из 725 смертей (против 3,2% лисиц , 0,7% сов и 0,5% других млекопитающих). [ 91 ] [ 128 ] Они также охотятся на птиц самых разных размеров, вплоть до самой маленькой птицы в Европе — золотогребня весом 5,2 г (0,18 унции) ( Regulus regulus ). [ 18 ] [ 94 ] Очень немногие отдельные птицы, на которых охотятся канюки, весят более 500 г (1,1 фунта). Тем не менее, канюками были убиты некоторые особенно крупные птицы, в том числе те, которые весили более 1000 г (2,2 фунта) или примерно самый большой средний размер канюка, в том числе взрослые особи кряквы ( Anas platyrhynchos ), тетерева ( Tetrao tetrix ), кольцевидный фазан ( Phasianus colchicus ), обыкновенный ворон ( Corvus corax ) и некоторые из более крупных чайки, если они попали в засаду на их гнезда. Самой крупной птицей, убитой канюком, и, возможно, самой крупной из известных для этого вида птиц, была взрослая самка глухаря ( Tetrao urogallus ), весившая примерно 1985 г (4,376 фунта). [ 94 ] [ 96 ] [ 98 ] [ 99 ] [ 115 ] Иногда канюки охотятся на птенцов крупных птиц, таких как цапли и журавли . [ 129 ] [ 130 ] Другая разнообразная птичья добыча включала несколько видов водоплавающих птиц , наиболее доступные голуби и голуби , кукушки , стрижи , поганки , рельсы , около 20 различных куликов , трубконосы , удоды , щурки и несколько видов дятлов . Птицы с более заметными или открытыми местами гнездования или привычками с большей вероятностью подвергаются нападению слетков или птенцов, например, водоплавающие птицы, в то время как птицы с более уединенными или недоступными гнездами, такие как голуби / голуби и дятлы, взрослые особи с большей вероятностью станут объектом охоты. [ 87 ] [ 98 ] [ 99 ] [ 107 ] [ 115 ] [ 131 ] [ 132 ] [ 133 ]
Рептилии и амфибии
[ редактировать ]
Обыкновенный канюк, возможно, является наиболее частым птичьим хищником рептилий и земноводных в Европе, за исключением тех разделов, где он симпатичен змееяду, питающемуся змееядом . [ 134 ] Всего кормовой спектр обыкновенного канюка включает около 50 герпетологических видов добычи. В исследованиях на севере и юге Испании ведущей добычей по численности были рептилии, хотя в Бискайе (северная Испания) ведущая добыча (19%) была классифицирована как «неопознанные змеи». В Мурсии самой многочисленной добычей была глазчатая ящерица ( Timon lepidus ) весом 77,2 г (2,72 унции) - 32,9%. В общей сложности на Бискайском полуострове и Мурсии на долю рептилий приходилось 30,4% и 35,9% объектов добычи соответственно. Аналогичные результаты были получены в отдельном исследовании, проведенном на северо-востоке Испании, где рептилии составляли 35,9% добычи. [ 87 ] [ 118 ] [ 135 ] В Бари , Италия , рептилии были основной добычей, составляя почти ровно половину биомассы, во главе с большой зеленой кнутовой змеей ( Hierophis viridiflavus ), составлявшей 24,2% пищевой массы. [ 123 ] В Ставропольском крае , Россия ) массой 20 г (0,71 унции) , песчаная ящерица ( Lacerta agilis была основной добычей в 23,7% из 55 объектов добычи. [ 100 ] ) массой 16 г (0,56 унции) Медленный червь ( Anguis fragilis , безногая ящерица, стал самой многочисленной добычей канюков южной Норвегии в годы низкой полевки, составляя 21,3% от 244 объектов добычи в 1993 году, а также был обычен даже в пик полевок – 1994 г. (19% из 332 объектов добычи). [ 94 ] Более или менее любая змея в Европе является потенциальной добычей, и канюк, как известно, необычайно смел в преследовании и подавлении крупных змей, таких как крысиные змеи , длиной почти до 1,5 м (4 фута 11 дюймов) и здоровых, крупные гадюки, несмотря на опасность быть пораженными такой добычей. [ 94 ] [ 136 ] [ 137 ] [ 138 ] Однако, по крайней мере, в одном случае труп самки канюка был найден отравленным над телом змеи . убитой ею [ 139 ] В некоторых частях ареала канюк приобретает привычку поедать множество лягушек и жаб. [ 3 ] Так было в Могилевской области Беларуси , ) массой 23 г (0,81 унции) где болотная лягушка ( Rana arvalis была основной добычей (28,5%) на протяжении нескольких лет, за ней следовали другие лягушки и жабы, составляющие 39,4% рациона в течение нескольких лет. годы. [ 100 ] В центральной Шотландии ) массой 46 г (1,6 унции) обыкновенная жаба ( Bufo bufo была самым многочисленным видом добычи, составляя 21,7% из 263 объектов добычи, в то время как обыкновенная лягушка ( Rana temporaria ) составляла еще 14,7% рациона. . [ 116 ] составляли около 10% рациона в центральной Польше . Лягушки также [ 111 ]
Беспозвоночные и другая добыча
[ редактировать ]Когда обыкновенные канюки питаются беспозвоночными, в Европе это в основном дождевые черви , жуки и гусеницы , и, по-видимому, на них в основном охотятся молодые канюки с менее развитыми охотничьими навыками или в районах с мягкой зимой и обильным роением или общественными насекомыми. В большинстве диетических исследований беспозвоночные в лучшем случае вносят незначительный дополнительный вклад в рацион канюка. [ 5 ] [ 99 ] в пище канюков обнаружено около десятка видов жуков Тем не менее, только в Украине . [ 85 ] Было обнаружено, что зимой на северо-востоке Испании канюки в основном переключились с добычи позвоночных, которую обычно добывают весной и летом, на рацион, в основном основанный на насекомых . Большая часть этой добычи была неопознанной, но чаще всего идентифицировались европейский богомол ( Mantis religiosa ) и европейский слепушонок ( Gryllotalpa gryllotalpa ). [ 118 ] В Украине 30,8% пищевых продуктов по численности составили насекомые . [ 85 ] Канюков часто привлекает роящаяся саранча и другие прямокрылые, особенно в зимних местах, таких как юг Африки. [ 3 ] [ 46 ] Таким образом, степной канюк может отражать аналогичного дальнего мигранта из Америки, ястреба Свенсона , который кормит своих детенышей в основном питательными позвоночными животными, но переключается на в основном насекомых, когда достигает своих отдаленных мест зимовки в Южной Америке . [ 4 ] В Эритрее были замечены 18 вернувшихся степных канюков, которые вместе питались стаями кузнечиков . [ 140 ] Что касается зимующих степных канюков в Зимбабве , один источник даже назвал их преимущественно насекомоядными, очевидно, в некоторой степени локально специализирующимися на питании термитами . [ 141 ] Содержимое желудка канюков из Малави , по-видимому, состояло в основном из кузнечиков (попеременно с ящерицами ). [ 142 ] Рыба, как правило, является самым редким классом добычи, встречающейся в пище канюка. зарегистрировано несколько случаев хищничества рыб В Нидерландах , а в других местах они, как известно, питались угрями и карпами . [ 98 ] [ 143 ] [ 144 ]
Межвидовые хищнические отношения
[ редактировать ]
Обыкновенные канюки встречаются вместе с десятками других хищных птиц в местах их размножения, проживания и зимовки. Может быть много других птиц, которые в некоторой степени частично совпадают в выборе добычи. Более того, многие хищные птицы активно используют их предпочтение границ леса и поля. Некоторые из наиболее сходных видов по питанию — это обыкновенная пустельга ( Falcotinniculus ), лунь ( Circus cyaenus ) и малый подорлик ( Clanga clanga ), не говоря уже о почти всех европейских видах сов, поскольку все, кроме двух, могут локально предпочитать грызунов. такие как полевки в их рационе. [ 145 ] [ 146 ] [ 147 ] [ 148 ] [ 149 ] было обнаружено значительное перекрытие рациона канюков и рыжих лисиц ( Vulpes vulpes ) В Польше , при этом 61,9% выбора добычи перекрываются по видам, хотя широта рациона лисицы была шире и более оппортунистична. Было обнаружено, что и берлоги лисиц, и насесты канюков находятся значительно ближе к местам обитания полевок по сравнению с окружающей средой в целом. [ 150 ] Единственный другой широко распространенный европейский бутео , шершавый канюк, часто зимует вместе с обыкновенными канюками. Он был обнаружен на юге Швеции, где среда обитания, охота и выбор добычи часто значительно пересекались. Мохоногие канюки, по-видимому, предпочитают немного более открытую среду обитания и потребляют немного меньше лесных мышей , чем обыкновенный канюк. Кроме того, шершавые ноги гораздо чаще парят и охотятся при сильном ветре. Два канюка агрессивны по отношению друг к другу и исключают друг друга из территорий зимнего кормления так же, как они исключают сородичей. В северной Германии буферность их предпочтений в среде обитания, очевидно, объясняет отсутствие влияния на размещение друг друга между двумя видами канюков. [ 151 ] [ 70 ] Несмотря на широкий диапазон совпадений, очень мало известно об экологии обыкновенного и длинноногого канюков , где они сосуществуют. Однако на основании того, что длинноногие виды предпочитают хищничество на разную добычу, такую как слепые землекопы , суслики , хомяки и песчанки , а также на основании того, что полевки обычно предпочитают обычные виды, серьезная конкуренция за пищу маловероятна. . [ 152 ] [ 153 ]
Более прямой отрицательный эффект был обнаружен при сосуществовании канюка с ястребом-тетеревятником ( Accipiter gentilis ). Несмотря на значительные различия в пищевых привычках этих двух видов, выбор среды обитания в Европе у канюков и ястребов-тетеревятников во многом аналогичен. Ястребы-тетеревятники немного крупнее канюков, являются более мощными, ловкими и, как правило, более агрессивными птицами, поэтому считаются доминирующими. В исследованиях, проведенных в Германии и Швеции, канюки оказались менее чувствительными к беспокойству, чем ястребы-тетеревятники, но, вероятно, были вытеснены доминирующими ястребами-тетеревятниками в более низкие места гнездования. Было обнаружено, что контакт канюков с чучелом ястреба-тетеревятника снижает успех размножения, тогда как контакт с чучелом канюка-тетеревятника не оказал никакого влияния на размножение ястребов-тетеревятников. [ 154 ] [ 155 ] [ 156 ] Во многих случаях в Германии и Швеции ястребы-тетеревятники вытесняли канюков из гнезд, чтобы забрать их себе. [ 157 ] [ 158 ] В Польше продуктивность канюков коррелировала с вариациями популяции добычи, особенно полевок, численность которой могла варьироваться от 10 до 80 на гектар, тогда как на ястребов-тетеревятников, по-видимому, не влияли вариации добычи; здесь встречены канюки в количестве 1,73 пары на 10 км. 2 (3,9 квадратных миль) против ястреба-тетеревятника 1,63 пары на 10 км. 2 (3,9 квадратных миль). [ 159 ] Напротив, немного более крупный аналог канюков в Северной Америке, краснохвостый ястреб (который также немного крупнее американских ястребов-тетеревятников, причем последние в среднем меньше европейских), по диете больше похож на местных ястребов-тетеревятников. Ястребы-тетеревятники не всегда доминируют над краснохвостыми, и они часто могут их вытеснить благодаря большей гибкости в питании и среде обитания. Более того, краснохвостые ястребы, по-видимому, в равной степени способны убивать ястребов-тетеревятников, как и ястребы-тетеревятники (при взаимодействии канюка и тетеревятника убийства более односторонние в пользу последних). [ 160 ] [ 161 ] [ 162 ] Другие хищные птицы, в том числе многие из них такого же или немного большего размера, чем сами канюки, могут доминировать над канюком или вытеснять его, особенно с целью захватить их гнезда. Известно , что такие виды, как черный коршун ( Milvus migrans ), орел-карлик ( Hieraeetus pennatus ) и малый подорлик, вытесняют активно гнездящихся канюков, хотя в некоторых случаях канюки могут пытаться защитить себя. Широкий спектр акципитрид, захватывающих гнезда канюков, несколько необычен. Чаще всего канюки становятся жертвами гнездового паразитизма сов и соколов , поскольку ни один из этих других видов хищных птиц не строит свои собственные гнезда, но они могут регулярно занимать уже заброшенные или альтернативные гнезда, а не те, которые канюки активно используют. . [ 127 ] [ 101 ] [ 163 ] [ 164 ] Даже птицы, традиционно не считающиеся хищными, такие как обыкновенные вороны , могут конкурировать с канюками за места гнездования. [ 165 ] В городских окрестностях юго-западной Англии было обнаружено, что сапсаны ( Falco peregrinus ) настолько настойчиво преследовали канюков, что во многих случаях приводили к травмам или смерти канюков, причем нападения имели тенденцию достигать пика в периоды размножения соколов и иметь тенденцию быть целенаправленными. на малолетних канюках. [ 166 ] Несмотря на то, что канюки часто доминируют в конфронтации в местах гнездования даже с хищниками одинакового размера, канюки, по-видимому, более смелы в прямой конкуренции за пищу с другими хищниками вне контекста размножения, и даже известно, что они вытесняют более крупных хищных птиц, таких как красные коршуны ( Milvus milvus ) и самки канюков также могут доминировать над самцами тетеревятника (которые намного меньше самок тетеревятника) при спорных добычах. [ 158 ] [ 167 ]

Обыкновенным канюкам иногда угрожает хищничество со стороны других хищных птиц. Известно, что северные ястребы-тетеревятники в некоторых случаях охотились на канюков. [ 168 ] [ 169 ] Известно, что гораздо более крупные хищники также убили несколько канюков, в том числе степных орлов ( Aquila nipalensis ) на мигрирующих степных канюках в Израиле . Дальнейшие случаи хищничества канюков были связаны с золотистым , восточным императорским ( Aquila heliaca ), орланом Бонелли ( Aquila fasciata ) и орланами-белохвостами ( Haliaeetus albicilla ) в Европе. [ 170 ] [ 171 ] [ 172 ] [ 173 ] [ 174 ] [ 175 ] Известно, что орланы-белохвосты не только охотятся на взрослых канюков, но и выращивают канюков вместе со своим потомством. Скорее всего, это случаи, когда орлы уносят молодых птенцов канюка с целью хищничества, но по неясным причинам не убивают их. Вместо этого мать-орел приходит высиживать молодого канюка. Несмотря на разницу в рационе двух видов, орланы-белохвосты на удивление успешно выращивают молодых канюков (которые заметно меньше, чем их собственные птенцы) до оперения. [ 176 ] [ 177 ] Исследования рациона орланов-белохвостов в Литве показали, что хищничество обыкновенных канюков было более частым, чем ожидалось: за 11 лет изучения летнего рациона орланов-белохвостов было обнаружено 36 останков канюка. Хотя по литовским данным птенцы канюков были в несколько раз более уязвимы для хищников, чем взрослые канюки, канюки региона тратили много времени и энергии в поздний период гнездования, пытаясь защитить свои гнезда. [ 178 ] [ 179 ] Однако самым серьезным хищником обыкновенных канюков почти наверняка является евразийский филин ( Bubo bubo ). Это очень крупная сова, средняя масса тела которой примерно в три-четыре раза превышает массу канюка. Филин, несмотря на то, что он часто охотится на мелких млекопитающих, которые во многом совпадают с теми, которых выбирают канюки, считается «суперхищником», который представляет собой серьезную угрозу почти для всех сосуществующих хищных птиц, способных уничтожать целые выводки других хищных птиц и отправляя взрослых хищников, даже таких крупных, как орлы. Из-за своей большой численности в окраинных местах обитания обыкновенные канюки часто составляют значительную часть рациона филина. Филины, как и некоторые другие крупные совы, также охотно экспроприируют гнезда канюков. [ 168 ] [ 180 ] [ 181 ] В Чехии и Люксембурге канюк был третьим и пятым по частоте добычей филинов соответственно. [ 182 ] [ 183 ] реинтродукция филин в некоторые районы Германии Было обнаружено, что оказала небольшой вредный эффект на местное заселение обыкновенными канюками. Единственным щадящим фактором является временная разница (канюк гнездится позже в году, чем филин), и на местном уровне канюки могут избегать гнездования рядом с активной семьей филин. [ 184 ] Поскольку экология зимующей популяции сравнительно мало изучена, подобная очень крупная сова, находящаяся на вершине пищевой цепи птиц, филин Верро ( Bubolacteus ), является единственным известным хищником зимующих степных канюков на юге Африки. [ 185 ] Несмотря на то, что они не являются известными хищниками канюков, известно, что другие крупные совы-полёвки вытесняют гнездящихся канюков или их избегают, например, большие серые совы ( Strix nebulosa ) и уральские совы ( Strix uralensis ). [ 169 ] [ 186 ] В отличие от крупных хищных птиц, о млекопитающих-хищниках обыкновенного канюка почти ничего не известно, несмотря на то, что млекопитающие, вероятно, истребили до нескольких птенцов и слетков. [ 17 ] [ 187 ] [ 188 ]
Сами канюки редко представляют угрозу для других хищных птиц, но иногда могут убить нескольких особей меньшего размера. Канюк - известный хищник, весящий 237 г (8,4 унции) евразийских перепелятников ( Accipiter nisus ), 184 г (6,5 унций) обыкновенной пустельги и 152 г (5,4 унции) малой пустельги ( Falco naumanni ). [ 108 ] [ 189 ] Возможно, это удивительно, но, учитывая ночной образ жизни этой добычи, группа хищных птиц, на которых канюк, как известно, охотится наиболее активно, — это совы. [ 190 ] Известная добыча совы включала 419 г (14,8 унции) сипух ( Tyto alba ), 92 г (3,2 унции) европейских сплюшек ( Otus scops ), 475 г (16,8 унции) желто-коричневых сов ( Strix aluco ), 169 г (6,0 унций) ) совы ( Athene noctua ), 138 г (4,9 унции) бореальные совы ( Aegolius funereus ), 286 г (10,1 унции) ушастые совы ( Asio otus ) и 355 г (12,5 унции) ушастые совы ( Asio flammeus ). [ 99 ] [ 118 ] [ 115 ] [ 108 ] [ 191 ] [ 192 ] Несмотря на свой относительно большой размер, коричневые совы, как известно, избегают канюков, поскольку есть несколько записей о том, как они охотились на сов. [ 98 ] [ 193 ] [ 194 ]
Разведение
[ редактировать ]
Гнездовые территории и плотность
[ редактировать ]Домашние ареалы канюков обычно составляют от 0,5 до 2 км. 2 (от 0,19 до 0,77 квадратных миль). [ 17 ] Размер территории размножения, по-видимому, обычно коррелирует с запасами пищи. [ 17 ] [ 195 ] В немецком исследовании дальность составляла от 0,8 до 1,8 км. 2 (от 0,31 до 0,69 квадратных миль) со средней площадью 1,26 км . 2 (0,49 квадратных миль). [ 70 ] Некоторые из самых низких плотностей пар обыкновенных канюков, похоже, происходят из России . Например, в Керженецком заповеднике зарегистрированная плотность составила 0,6 пары на 100 км. 2 (39 квадратных миль), а среднее расстояние до ближайших соседей составляло 3,8 км (2,4 мили). [ 196 ] Регион Сноудония на севере Уэльса удерживал пару на 9,7 км. 2 (3,7 квадратных миль) со средним расстоянием до ближайшего соседа 1,95 км (1,21 мили); в соседнем Миньене встречаемость пар составила 7,2 км. 2 (2,8 квадратных миль), со средним расстоянием 1,53 км (0,95 мили). [ 197 ] В массиве Тено на Канарских островах средняя плотность оценивалась в 23 пары на 100 км. 2 (39 квадратных миль), что соответствует численности среднего континентального населения. [ 198 ] На другом острове, на Крите, плотность пар была ниже — 5,7 пар на 100 км. 2 (39 квадратных миль); Здесь канюки, как правило, имеют неравномерное распространение: некоторые из них обитают в оливковых рощах с меньшей интенсивностью сбора урожая, но на самом деле их появление более распространено в сельскохозяйственных, чем в природных зонах. [ 199 ] В итальянских Альпах в 1993–96 годах было зафиксировано от 28 до 30 пар на 100 км. 2 (39 квадратных миль). [ 127 ] В центральной Италии средняя плотность была ниже — 19,74 пары на 100 км. 2 (39 квадратных миль). [ 200 ] Известны области с более высокой плотностью населения, чем указанные выше. В двух районах Мидлендса Англии . была заселенность 81 и 22 территориальных пар на 100 км 2 (39 квадратных миль). Высокая плотность канюков здесь была связана с высокой долей неулучшенных пастбищ и спелых лесов на оцениваемых территориях. [ 92 ] Столь же высокая плотность канюков была оценена в центральной Словакии с использованием двух разных методов: здесь плотность составляет от 96 до 129 пар на 100 км. 2 (39 квадратных миль). [ 201 ] Несмотря на утверждения, полученные в ходе исследования в центральных графствах Англии, там была самая высокая известная плотность территории для этого вида: от 32 до 51 пары на лесистой территории площадью всего 22 км . 2 (8,5 квадратных миль) в Чехии, кажется, наверняка превышает даже эту плотность. Чешское исследование предположило, что фрагментация лесов в управлении людьми землями для диких баранов и оленей приводит к исключительной концентрации добычи, такой как полевки, и отсутствию подходящей среды обитания в прилегающих регионах для исключительно высокой плотности. [ 92 ] [ 202 ]
В северо-эстонском ландшафтном заповеднике Неэрути (площадь 1250 га) Марек Вахула в 1989 и 1990 годах обнаружил 9 населенных гнезд. Одно гнездо было найдено в 1982 году и, по-видимому, является самым старым известным гнездом, которое до сих пор населено. [ нужна ссылка ]

Обыкновенные канюки сохраняют свою территорию с помощью полетов. В Европе территориальное поведение обычно начинается в феврале. Тем не менее, демонстрации нередки в течение года среди местных пар, особенно со стороны самцов, и могут вызывать аналогичные проявления со стороны соседей. [ 3 ] [ 17 ] В них обыкновенные канюки обычно кружат высоко, поднимаясь по спирали вверх на слегка приподнятых крыльях. Взаимное высокое кружение парами иногда продолжается продолжительное время, особенно в период до или во время сезона размножения. При взаимном отображении пара может следовать друг за другом на расстоянии 10–50 м (33–164 фута) в горизонтальном полете. Во время взаимных демонстраций самец может преувеличенно хлопать руками или зигзагообразно кувыркаться, очевидно, в ответ на то, что самка находится слишком далеко. Иногда две или три пары могут кружить вместе, и на установленных площадках было зарегистрировано до 14 отдельных взрослых особей. [ 3 ] [ 203 ] Небесные танцы обыкновенных канюков были зарегистрированы весной и осенью, обычно самцами, но иногда и самками, почти всегда с большим призывом. Их небесные танцы напоминают американские горки, с подъемом вверх до тех пор, пока они не начинают срываться, но иногда украшенные петлями или перекатами наверху. Затем в небесном танце они пикируют на более или менее закрытых крыльях, прежде чем расправить их и снова взлететь вверх, взмахами вверх на высоту до 30 м (98 футов) с падением на высоту не менее 60 м (200 футов). Эти танцы можно повторять сериями от 10 до 20. В кульминации небесного танца волны становятся все более мелкими, часто замедляются и заканчиваются прямо на жердочке. Различные другие воздушные демонстрации включают полет по низкому контуру или петление среди деревьев, часто с глубокими взмахами и преувеличенными взмахами вверх, которые показывают рисунок подкрыльев соперникам, сидящим внизу. [ 3 ] [ 204 ] У канюков было зарегистрировано схватывание когтями и иногда перекатывание вниз со сцепленными ногами, и, как и у многих хищников, это, вероятно, является физической кульминацией агрессивной территориальной демонстрации, особенно между самцами. [ 17 ] [ 205 ] Несмотря на очень территориальный характер канюков и их преданность одному партнеру и месту размножения каждое лето, есть один случай, когда полиандрическая троица канюков гнездилась на Канарских островах . [ 206 ]
Гнезда
[ редактировать ]Обыкновенные канюки обычно строят громоздкие гнезда из палочек, веток и часто из вереска . Обычно гнезда имеют диаметр от 1 до 1,2 м (от 3 футов 3 дюйма до 3 футов 11 дюймов) и глубину 60 см (24 дюйма). При повторном использовании в течение многих лет диаметр может достигать или превышать 1,5 м (4 фута 11 дюймов), а вес гнезд может достигать более 200 кг (440 фунтов). [ 3 ] [ 17 ] Активные гнезда, как правило, выложены зеленью, чаще всего она состоит из широколистной листвы, но иногда также включает в себя камыш или водоросли местные . Высота гнезда на деревьях обычно составляет от 3 до 25 м (от 9,8 до 82,0 футов), обычно по основному стволу или основной опоре дерева. [ 3 ] [ 17 ] [ 5 ] В Германии деревья, используемые для гнездования, состояли в основном из красного бука ( Fagus sylvatica ) (в 337 случаях), тогда как еще 84 были в различных дубах . [ 207 ] Было зарегистрировано, что канюки гнездятся почти исключительно на соснах в Испании на средней высоте 14,5 м (48 футов). [ 87 ] Деревья обычно используются в качестве места гнездования, но они также могут использовать скалы или обрывы, если деревья недоступны. [ 17 ] Канюки в одном английском исследовании были на удивление неравнодушны к гнездованию на берегах с густой растительностью и из-за богатой окружающей среды обитания и популяции добычи на самом деле были более продуктивными, чем гнезда, расположенные в других местах здесь. [ 208 ] Кроме того, несколько наземных гнезд были зарегистрированы в сельскохозяйственных районах Нидерландов с высоким уровнем добычи . [ 209 ] В итальянских Альпах 81% из 108 гнезд находились на скалах. [ 127 ] Обыкновенному канюку, как правило, не хватает склонности его неарктического аналога, краснохвостого ястреба , время от времени гнездиться на искусственных сооружениях или рядом с ними (часто в густонаселенных районах), но в Испании некоторые пары зарегистрировали гнездование по периметру заброшенных зданий. [ 210 ] Пары часто имеют несколько гнезд, но некоторые пары могут использовать одно в течение нескольких лет подряд. Для обыкновенных канюков, особенно для тех, кто гнездится севернее своего ареала, типично от двух до четырех альтернативных гнезд на территории. [ 17 ] [ 211 ]
Размножение и яйца
[ редактировать ]
Сезон размножения начинается в разное время в зависимости от широты. Обычные сезоны размножения канюков могут приходиться уже на январь-апрель, но обычно на большей части территории Палеарктики сезон размножения приходится на март-июль . На северных участках ареала сезон размножения может длиться до мая – августа. Спаривание обычно происходит в гнезде или рядом с ним и длится около 15 секунд, обычно происходит несколько раз в день. [ 3 ] [ 17 ] Яйца обычно откладываются с интервалом в 2–3 дня. Размер кладки может варьироваться от 2 до 6, что является относительно большой кладкой для ястреба. Более северные и западные канюки обычно несут более крупные кладки, в среднем около 3, чем те, что восточнее и южнее. [ 3 ] [ 17 ] [ 198 ] В Испании средний размер кладки составляет от 2 до 2,3. [ 87 ] Из 4 местоположений в разных частях Европы 43% имели размер кладки 2, 41% - размер 3, кладки 1 и 4 каждая составляли около 8%. [ 212 ] Даты кладки удивительно постоянны по всей Великобритании . Однако существуют весьма существенные различия в размере кладки между британскими территориями исследований. Они не подчиняются какому-либо широтному градиенту, и вполне вероятно, что местные факторы, такие как среда обитания и доступность добычи, являются более важными определяющими размер кладки. [ 187 ] Яйца белого цвета, довольно круглой формы со спорадическими пятнами от красного до коричневого цвета, иногда слегка выступающими. У номинальной расы размер яиц составляет 49,8–63,8 мм (1,96–2,51 дюйма) в высоту и 39,1–48,2 мм (1,54–1,90 дюйма) в диаметре, в среднем 55 мм × 44 мм (2,2 дюйма × 1,7 дюйма) в диаметре. 600 яиц. У расы vulpinus высота яиц составляет 48–63 мм (1,9–2,5 дюйма) на 39,2–47,5 мм (1,54–1,87 дюйма), в среднем 54,2 мм × 42,8 мм (2,13 дюйма × 1,69 дюйма) в 303 яйцах. [ 17 ] Яйца обычно откладываются в конце марта - начале апреля на крайнем юге, где-то в апреле в большей части Европы, в мае и, возможно, даже в начале июня на крайнем севере. Если яйца потеряны хищником (в том числе человеком) или по каким-либо причинам терпят неудачу, обыкновенные канюки обычно не откладывают запасных кладок, но они регистрируются даже при трех попытках кладок одной самкой. [ 3 ] [ 17 ] [ 115 ] Самка осуществляет большую часть инкубации, но не всю, в общей сложности 33–35 дней. [ 3 ] [ 17 ] Самка остается в гнезде, высиживая детенышей на ранних стадиях, а самец приносит всю добычу. Примерно через 8–12 дней и самец, и самка принесут добычу, но самка продолжает питаться до тех пор, пока детеныши не смогут разорвать свою добычу. [ 17 ]
Развитие молодых
[ редактировать ]
После начала вылупления птенцу может потребоваться 48 часов, чтобы вылупиться. Вылупление может происходить в течение 3–7 дней, при этом масса тела новых птенцов в среднем составляет около 45 г (1,6 унции). [ 3 ] [ 17 ] [ 5 ] Нередко самый младший птенец погибает от голода, особенно в выводках из трех и более особей. [ 17 ] У птенцов первый пух заменяется более длинным и грубым пухом примерно в 7-дневном возрасте, а первые настоящие перья появляются на 12-15 день. Примерно в месячном возрасте детеныши почти полностью оперены, а не опушены, и также могут начать питаться самостоятельно. Первые попытки покинуть гнездо часто совершаются примерно через 40–50 дней, в среднем обычно 40–45 у канюков-номинантов в Европе, но быстрее в среднем на 40–42 дня у vulpinus . Окрыление обычно происходит через 43–54 дня, но в крайних случаях даже через 62 дня. Половой диморфизм очевиден у птенцов Европы, поскольку самки часто весят около 1000 г (2,2 фунта) против 780 г (1,72 фунта) у самцов. [ 17 ] [ 5 ] [ 213 ] Покинув гнездо, канюки обычно остаются поблизости, но у перелетных канюков наблюдается более четкое движение, обычно в южном направлении. Полной независимости обычно добиваются через 6–8 недель после оперения. Пернатые птицы обычно остаются в районе зимовки на следующее лето, но затем возвращаются в район, близкий к месту происхождения, а затем снова мигрируют на юг, не размножаясь. [ 3 ] [ 17 ] [ 5 ] Радиослежение показывает, что большая часть расселения, даже относительно раннего, молодых канюков осуществляется самостоятельно, а не через изгнание родителей, как это было зарегистрировано у некоторых других хищных птиц. [ 214 ] У обыкновенных канюков, вообще говоря, братья и сестры после расселения от родителей держатся довольно близко друг к другу и образуют нечто вроде социальной группы, хотя родители обычно терпят их присутствие на своей территории до тех пор, пока они не отложат очередную кладку. Однако социальная группа братьев и сестер распадается примерно в годовом возрасте. [ 213 ] [ 84 ] Молодые канюки подчиняются взрослым во время большинства встреч и склонны избегать прямых столкновений и активно защищаемых территорий, пока не достигнут соответствующего возраста (обычно не менее 2 лет). [ 5 ] То же самое произошло и с молодью степного канюка, зимующей на юге Африки, хотя в некоторых случаях молоди удавалось успешно украсть там добычу у взрослых особей. [ 215 ]

Показатели успеха размножения
[ редактировать ]На успех размножения обыкновенных канюков могут влиять многочисленные факторы. В основном среди них - популяция добычи, среда обитания, уровень беспокойства и преследования, а также внутривидовая конкуренция. [ 17 ] В Германии наиболее значимыми факторами успеха размножения считались внутри- и межвидовая конкуренция, морфа оперения, дата откладки яиц, уровень осадков и антропогенные нарушения на территории размножения (в порядке убывания). [ 195 ] В сопроводительном исследовании было обнаружено, что всего лишь 17% взрослых птиц обоих полов, обитающих на территории исследования в Германии, производят 50% потомства, поэтому успех размножения может быть ниже, чем предполагалось, и многие взрослые канюки по неизвестным причинам могут не пытаться разводить вообще. [ 216 ] Высокий успех размножения был обнаружен в Аргайле , Шотландия , вероятно, из-за обильной популяции добычи (кроликов), но также, вероятно, из-за более низкого местного уровня преследований, чем где-либо еще на Британских островах. Здесь среднее количество птенцов составляло 1,75 против 0,82–1,41 в других частях Британии. [ 187 ] было обнаружено В английском Мидлендсе , что успех размножения как по размеру кладки, так и по среднему количеству птенцов был относительно высоким, опять же благодаря высокой популяции добычи. Успех размножения был ниже по мере удаления от значительных насаждений деревьев в Мидлендсе, и большинство неудачных попыток гнездования, которые можно было определить, произошли на стадии инкубации, возможно, в связи с хищничеством яиц врановыми . [ 92 ] Было обнаружено, что конец зимы — начало весны, более важный, чем даже добыча, является, вероятно, основным фактором успеха размножения канюков из южной Норвегии . Здесь, даже в пиковые годы полевок , успешному гнездованию может существенно помешать сильный снегопад на этом решающем этапе. В Норвегии крупные кладки 3+ ожидались только в годы с минимальным снежным покровом, высокой популяцией полевок и меньшими дождями в мае–июне. [ 211 ] В итальянских Альпах среднее количество птенцов на пару составило 1,07. [ 127 ] Согласно исследованию, проведенному на юго-западе Германии , 33,4% попыток гнездования оказались неудачными , в среднем 1,06 всех попыток гнездования и 1,61 всех успешных попыток. В Германии погодные условия и популяция грызунов, по-видимому, были основными факторами успеха гнездования. [ 207 ] В Мурсии, части Испании, в отличие от Бискайи на севере, более высокий уровень межвидовой конкуренции со стороны орлов-карликов и северных ястребов-тетеревятников, по-видимому, не оказал отрицательного влияния на успех размножения из-за более обширных популяций добычи (опять же кроликов) в Мурсии, чем в Бискайе. [ 87 ]
В районе Вестфалии в Германии было обнаружено, что морфы промежуточной окраски были более продуктивными, чем более темные или светлые. [ 216 ] По причинам, которые не совсем ясны, по-видимому, было обнаружено, что меньшее количество паразитов поражают выводки канюка со средним оперением в меньшей степени, чем темные и светлые фенотипы , в частности, более высокие уровни меланина каким-то образом оказались более привлекательными для паразитических организмов, которые влияют на здоровье канюка. потомство. [ 217 ] Состав среды обитания и его связь с вмешательством человека были важными переменными для темных и светлых фенотипов, но были менее важны для промежуточных особей. Таким образом, давление отбора, вызванное различными факторами, не сильно различалось между полами, но различалось между тремя фенотипами в популяции. [ 218 ] На успех размножения в районах обитания диких европейских кроликов существенно повлиял миксоматоз кроликов и геморрагическая болезнь кроликов , которые сильно истощили популяцию диких кроликов. Было зарегистрировано снижение успешности размножения в районах, ранее богатых кроликами, с 2,6 до всего лишь 0,9 детенышей на пару. [ 165 ] [ 219 ] [ 220 ] Возраст первого размножения у нескольких канюков с радиометками показал, что только один самец размножался уже на второе лето (около года). Было обнаружено, что значительно больше канюков начинают размножаться в 3-е лето, но попытки размножения могут быть беспорядочными, учитывая наличие среды обитания, пищи и партнеров. [ 221 ] В конце 1950-х годов средняя продолжительность жизни оценивалась в 6,3 года, но это было во время сильных преследований, когда люди были причиной 50–80% смертей канюков. [ 90 ] В более современном контексте, когда уровень преследований в регионе снизился, ожидаемая продолжительность жизни может быть выше (иногда может превышать 10 лет), но все еще сильно варьируется из-за широкого спектра факторов. [ 218 ]
Статус
[ редактировать ]
Обыкновенный канюк — одна из самых многочисленных хищных птиц своего ареала. Почти наверняка это самая многочисленная дневная хищная птица во всей Европе. По скромным оценкам, общая популяция в Европе составляет не менее 700 000 пар, что более чем в два раза превышает общие оценки следующих четырех хищных птиц, которые считаются наиболее распространенными: евразийского перепелятника (более 340 000 пар), обыкновенной пустельги (более 340 000 пар). более 330 000 пар) и ястреб-тетеревятник (более 160 000 пар). [ 6 ] Фергюсон-Лис и др. По приблизительным оценкам, общая популяция обыкновенного канюка составляет почти 5 миллионов пар, но в то время в это число входили теперь отделившиеся виды восточного и гималайского канюков . [ 3 ] Эти цифры могут быть чрезмерными, но общая популяция обыкновенных канюков наверняка превышает семизначную цифру. [ 1 ] [ 3 ] Совсем недавно МСОП подсчитал, что численность обыкновенного канюка (за исключением гималайского и восточного подвидов) составляет где-то от 2,1 до 3,7 миллионов птиц, что делает этого канюка одним из самых многочисленных представителей семейства ястребиных (по оценкам евразийских перепелятников, красно-ястребов). хвостатых ястребов и ястребов-тетеревятников также может достигать более 2 миллионов). [ 1 ] В 1991 году, помимо их отсутствия в Исландии , после того, как к 1910 году они стали заводчиками, канюки повторно заселили Ирландию где-то в 1950-х годах, и к 1990-м годам их численность увеличилась до 26 пар. [ 3 ] [ 117 ] Сообщается, что дополнительное кормление помогло восстановить популяцию ирландского канюка, особенно там, где численность кроликов сократилась. [ 59 ] В большинстве других стран есть как минимум четыре особи гнездящихся пар. По состоянию на 1990-е годы в других странах, таких как Великобритания , Франция , Швейцария , Чехия , Польша , Швеция , Беларусь и Украина, количество пар было пятизначным, в то время как в Германии было около 140 000 пар, а в европейской части России, возможно, было 500 000 пар. [ 3 ] [ 6 ] К 2001 году в Великобритании гнездилось от 44 000 до 61 000 пар , причем их численность постепенно увеличивалась после прошлых преследований, изменения среды обитания и сокращения добычи, что сделало его, безусловно, самым многочисленным дневным хищником в этой стране. [ 222 ] В Вестфалии , Германия , популяция канюков увеличилась почти втрое за последние несколько десятилетий. Вестфальские канюки, возможно, получают выгоду от более теплого среднего климата, что, в свою очередь, увеличивает уязвимость полевок. Однако темпы роста были значительно выше у самцов, чем у самок, отчасти из-за повторного завоза в регион евразийских филин, охотящихся на гнезда (включая насиживающую мать), что, в свою очередь, может оказать чрезмерное давление на местную популяцию канюков. [ 223 ]
По крайней мере, 238 канюков, убитых в результате преследований, были обнаружены в Англии с 1975 по 1989 год, в основном в результате отравления. Уровень преследований ни разу за этот период времени существенно не отличался, а уровень преследований не снизился, как и не снизился по сравнению с показателями последнего исследования в 1981 году. [ 224 ] Хотя в Англии продолжаются некоторые преследования, сегодня они, вероятно, немного менее распространены. [ 222 ] Канюк оказался наиболее уязвимым хищником в результате столкновений с линиями электропередач в Испании, вероятно, потому, что это одна из самых распространенных крупных птиц, и вместе с обыкновенным вороном на его долю приходится почти треть зарегистрированных поражений электрическим током. [ 225 ] Учитывая его относительную численность, обыкновенный канюк считается идеальным биоиндикатором , поскольку он, как и другие хищники, подвергается воздействию различных пестицидов и металлов в результате загрязнения, но в значительной степени устойчив к ним на популяционном уровне. В свою очередь, это позволяет биологам интенсивно изучать (и при необходимости ловить ) канюков и окружающую среду, не влияя на их общую популяцию. Отсутствие воздействия может быть связано с приспособляемостью канюка, а также с его относительно короткой наземной пищевой цепью, которая подвергает их меньшему риску заражения и истощения популяции, чем хищники, которые в большей степени охотятся на водную добычу (например, некоторые большие орлы) или другие птицы (например, соколы ). [ 226 ] [ 227 ] [ 228 ] [ 229 ] [ 230 ] Обыкновенные канюки редко подвержены истончению скорлупы яиц под действием ДДТ , как и другие хищные птицы, но были зарегистрированы случаи истончения скорлупы яиц. [ 231 ] Другими факторами, отрицательно влияющими на хищников, которые были изучены на канюках, являются гельминты , авипоксвирус и ряд других вирусов . [ 232 ] [ 233 ] [ 234 ] [ 235 ]
Галерея
[ редактировать ]-
 В зоопарке Хамертона , Англия.
В зоопарке Хамертона , Англия. -
 В полете, Девон, Англия. насчитывается около 40 000 гнездящихся пар . В Соединенном Королевстве
В полете, Девон, Англия. насчитывается около 40 000 гнездящихся пар . В Соединенном Королевстве -
 Яйцо, Коллекция музея Висбадена
Яйцо, Коллекция музея Висбадена -
 Окольцовывают молодого канюка
Окольцовывают молодого канюка -
 Неоплодотворённое яйцо канюка ( Buteobuteo ).
Неоплодотворённое яйцо канюка ( Buteobuteo ).





Ссылки
[ редактировать ]Цитаты
[ редактировать ]- ^ Перейти обратно: а б с д и ж г час я BirdLife International (2020). « Бутео бутео » . Красный список исчезающих видов МСОП . 2020 : e.T61695117A181770099. doi : 10.2305/IUCN.UK.2020-3.RLTS.T61695117A181770099.en . Проверено 13 ноября 2021 г.
- ^ Перейти обратно: а б с д и Уоллс, Шон; Кенвард, Роберт (23 января 2020 г.). Обыкновенный канюк . Издательство Блумсбери. ISBN 978-1-4729-7002-2 . Проверено 22 сентября 2023 г.
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и объявление но из в ах есть также и аль являюсь а к ап ак с как в В из хорошо топор является тот нет бб до нашей эры др. быть парень бг чб с Фергюсон-Лис, Дж., и Кристи, Д.А. (2001). Хищники мира . Хоутон Миффлин Харкорт.
- ^ Перейти обратно: а б с Бильдштейн, К.Л., и Заллес, Дж.И. (2005). Дальняя миграция ястребов, бутео и соколов Старого Света и Нового Света . Птицы двух миров: экология и эволюция миграций. Издательство Университета Джонса Хопкинса, Балтимор, Мэриленд, США, 154–167.
- ^ Перейти обратно: а б с д и ж г час я дж к Крамп С. и Брукс диджей (1992). Справочник птиц Европы, Ближнего Востока и Северной Африки. Птицы западной Палеарктики, т. 2 дюйма. Издательство Оксфордского университета, Оксфорд.
- ^ Перейти обратно: а б с д BirdLife, ФВБ (2005). Птицы в Европе: оценки численности, тенденции и статус охраны . Британские птицы, 98, 269–271.
- ^ Линней, К. (1758). Система природы по трем царствам природы, по классам, отрядам, родам, видам, с признаками, различиями, синонимами, местами. Томус I. Издание десятое, исправленное (на латыни). Том. 1. Гольмии (Стокгольм): Лаврентий Сальвий. с. 90.
Ф. восковые и желтые ноги, коричневое тело, коричневое брюшко.
- ^ Ласепед, Бернар Жермен де (1799). «Таблица подклассов, отделов, подразделений, отрядов и родов птиц» . Вступительные и заключительные речи курса естествознания (на французском языке). Париж: Плассан. п. 4. Нумерация страниц начинается с единицы для каждого из трех разделов.
- ^ Майр, Эрнст ; Коттрелл, Дж. Уильям, ред. (1979). Контрольный список птиц мира. Том 1 . Том. 1 (2-е изд.). Кембридж, Массачусетс: Музей сравнительной зоологии. п. 361.
- ^ Джоблинг, Джеймс А. (2010). Словарь научных названий птиц Хелма . Лондон: Кристофер Хелм. п. 81 . ISBN 978-1-4081-2501-4 .
- ^ Перейти обратно: а б Рисинг М.Дж., Крукенхаузер Л., Гамауф А. и Харинг Э. (2003). «Молекулярная филогения рода Buteo (Aves: Accipitridae) на основе последовательностей митохондриальных маркеров». Молекулярная филогенетика и эволюция . 27 (2): 328–42. Бибкод : 2003МОЛПЭ..27..328Р . дои : 10.1016/S1055-7903(02)00450-5 . ПМИД 12695095 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Перейти обратно: а б Кларк, WS (2007). «Таксономический статус лесного канюка Buteo oreophilus trizonatus » (PDF) . Страус . 78 (1): 101–104. Бибкод : 2007Остри..78..101С . дои : 10.2989/СТРУС.2007.78.1.16.60 . S2CID 6270968 . Архивировано из оригинала (PDF) 31 августа 2021 года . Проверено 26 февраля 2019 г.
- ^ Харинг Э., Рисинг М.Дж., Пинскер В. и Гамауф А. (1999). Эволюция псевдоконтролируемой области в митохондриальном геноме палеарктических канюков (род Buteo) . Журнал зоологической систематики и эволюционных исследований, 37 (4), 185–194.
- ^ Зигфрид, WR (1969). Родство небольших африканских и палеарктических бутео . Страус, 40(С1), 41–46.
- ^ Перейти обратно: а б Клуэ М. и Винк М. (2000). Канюки Кабо-Верде-Бутео (бутео) баннермани и Сокотра-Бутео (бутео) виды: первые результаты генетического анализа на основе нуклеотидных последовательностей гена цитохрома b . Алауда, 68 (1), 55–58.
- ^ Боев З. и Ковачев Д. (1998). Бутео спассови сп. N. – поздний мионец канюк (Accipitridae, Aves) из Юго-Западной Болгарии . Geologica Balkanica, 29 (1–2), 125–129.
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и объявление но из в ах Браун, Лесли и Амадон, Дин (1986) Орлы, ястребы и соколы мира . Уэллфлит Пресс. ISBN 978-1555214722 .
- ^ Перейти обратно: а б с Справочник CRC по массе тела птиц, 2-е издание Джона Б. Даннинга-младшего (редактор). ЦРК Пресс (2008), ISBN 978-1-4200-6444-5 .
- ^ Селас, В. (1997). Выбор места гнезда четырьмя симпатрическими лесными хищниками на юге Норвегии . Журнал исследований хищников, 31, 16–25.
- ^ Зуберогойтия, И., Мартинес, Х.А., Сабала, Дж., Мартинес, Х.Э., Кастильо, И., Аскона, А., и Идальго, С. (2005). Определение пола, старение и линька канюков Бутео-бутео в южной части Европы . Звон и миграция, 22 (3), 153–158.
- ^ Перейти обратно: а б с д и Джеймс, АХ (1984). Географическая изменчивость канюка Buteo Buteo (Linnaeus, 1758): острова Средней Атлантики и западного Средиземноморья (Aves: Accipitridae) . Бофорт, 34 (4), 101–116.
- ^ Тибо, Йена-Клод; Патримонио, Оливье; Торре, Хосе (июль 1992 г.). «Обладает ли дневное сообщество хищников Корсики (Западное Средиземноморье) островными характеристиками?». Журнал биогеографии . 19 (4): 363–373. Бибкод : 1992JBiog..19..363T . дои : 10.2307/2845564 . eISSN 1365-2699 . ISSN 0305-0270 . JSTOR 2845564 .
- ^ «Степной канюк – Buteo Buteo vulpinus (Gloger, 1833)» . Авибаза
- ^ Перейти обратно: а б с Крукенхаузер Л., Харинг Э., Пинскер В., Рисинг М.Дж., Винклер Х., Винк М. и Гамауф А. (2004). Генетическая и морфологическая дифференциация канюков Старого Света (род Buteo, Accipitridae) . Зоологика Скрипта, 33(3), 197–211.
- ^ Перейти обратно: а б Вус, К.Х., Хенс, Пенсильвания, и Ван Марл, Дж.Г. (1948). Отличительные черты степного канюка . ВЧ и Г. Уизерби.
- ^ Дементьев Г.П., Гладков Н.А. (1951). Птицы Советского Союза, т. 1 . Советская Наука, Москва.
- ^ Перейти обратно: а б Горни Э. и Йом-Тов Ю. (1994). Жир, состояние гидратации и линька степного канюка Buteobuteo vulpinus во время весенней миграции . Ибис, 136 (2), 185–192.
- ^ Йосеф Р., Тряновски П. и Бильдштейн К.Л. (2002). Весенняя миграция взрослых и неполовозрелых канюков (Buteobuteo) через Эйлат, Израиль: время и размер тела . Журнал исследований хищников, 36 (2), 115–120.
- ^ Бенсон, CW, и Ирвин, MPS (1967). Вклад в орнитологию Замбии (№ 1) . опубликовано.
- ^ Брокхейсен, Г.Дж., и Зигфрид, В.Р. (1969). Возраст и линька степного канюка на юге Африки . Страус, 40(С1), 223–237.
- ^ «Обыкновенный канюк (Восточная степь) – Buteobuteo menetriesi Bogdanov, 1879» . Авибаза .
- ^ Макворт-Пред, CW, и Грант, CHB (1952). Птицы восточной и северо-восточной Африки (Том 1) . Лонгманс, Грин.
- ^ Харинг Э., Рисинг М.Дж., Пинскер В. и Гамауф А. (1999). Эволюция псевдоконтролируемой области в митохондриальном геноме палеарктических канюков (род Buteo) . Журнал зоологической систематики и эволюционных исследований, 37 (4), 185–194.
- ^ Воус, К. Х. и Биджлевелд, MFIJ (1964). Заметка о гималайских канюках, Бутео бутео (Птицы ). Бофорт, 11 (134), 37–43.
- ^ Аспиналл, С. (2001). Население Бутео на Сокотре . Фалько, 8.
- ^ Перейти обратно: а б с д и ж Форсман, Д. (1999). Хищники Европы и Ближнего Востока: справочник полевой идентификации . Лондон: T&AD Poyser.
- ^ Даннинг, Джон Б. младший, изд. (1992). Справочник CRC по массе тела птиц . ЦРК Пресс. ISBN 978-0-8493-4258-5 .
- ^ дель Ойо, Дж.; Эллиотт, А.; Саргатал, Дж., ред. (1999). Справочник птиц мира . Том. 2. Барселона: Lynx Edicions. ISBN 978-84-87334-15-3 .
- ^ Перейти обратно: а б с Кларк, WS (1999). Полевой справочник по хищникам Европы, Ближнего Востока и Северной Африки . Издательство Оксфордского университета, США.
- ^ Перейти обратно: а б с д и Портер, РФ (1981). Идентификация полетов европейских хищников . А&С Черный.
- ^ Диттрих, В. (1985). Вариации оперения обыкновенного канюка (Buteobuteo) в северной Баварии . Журнал орнитологии, 126 (1), 93–97.
- ^ Шрайбер А., Стуббе А. и Стуббе М. (2001). Обыкновенный канюк (Buteobuteo): хищник с гиперполиморфными морфами оперения, но с низкой аллозимной гетерозиготностью . Журнал für Ornithologie, 142 (1), 34–48.
- ^ Ровер, С., и Полсон, Д.Р. (1987). Гипотеза избегания изображения и цветовой полиморфизм у ястребов Бутео . Скандинавский Орнис, 285–290.
- ^ Дафф, Д. (2006). Развилось ли оперение молоди осоха, чтобы имитировать оперение обыкновенного канюка? Британские птицы, 99, 118–128.
- ^ Элорриага, Дж., и Муньос, АР (2013). Гибридизация обыкновенного канюка и североафриканского канюка Buteo rufinus cirtensis в Гибралтарском проливе: прелюдия или препятствие колонизации? Страус, 84(1), 41–45.
- ^ Перейти обратно: а б с д и Стейн, П. (1983). Хищные птицы юга Африки: их идентификация и история жизни . Крум Хелм, Бекенхэм (Великобритания). 1983.
- ^ Анвин, М. (2011). Атлас птиц: разнообразие, поведение и охрана. Издательство Принстонского университета.
- ^ Перейти обратно: а б Мехерету Йонас; Лейрс, Х (2019). Участки хищных птиц для биологической борьбы с грызунами-вредителями сельского хозяйства. В: Ниссен Дж., Джейкоб М., Франкл А. (ред.). Гео-треккинг в тропических горах Эфиопии – район Догуа Тембиен . Спрингер Природа. ISBN 978-3-030-04954-6 .
- ^ Харрисон, Дж. А., и Черри, М. (1997). Атлас птиц юга Африки (Том 1) . Йоханнесбург: BirdLife, Южная Африка.
- ^ Браун, Л., Урбан, Э.К., Ньюман, К., Вудкок, М., и Хейман, П. (1982). Птицы Африки (т. 1, стр. 521) . Лондон: Академическая пресса.
- ^ Витер, С.Г. (2019). Лесохозяйственная деятельность и ее влияние на популяции хищных птиц в южной лесостепной и степной зонах (на примере Харьковской области Украины) . Сохранение хищников, 38.
- ^ Камараускайте А., Скуя С. и Трейнис Р. (2019). Места гнездования обыкновенного канюка Buteo Buteo и малого подорлика Clanga pomarina перекрываются для планирования сохранения на участках Natura 2000 . Исследование птиц, 66(2), 224–233.
- ^ Йджжеевский В., Йджеевска Б. и Келлер М. (1988). Выбор места для гнезда канюком Buteo Buteo L. в обширных лесах восточной Польши . Биологическая консервация, 43(2), 145–158.
- ^ Пентериани, В. и Фавр, Б. (1997). Выбор места обитания канюка обыкновенного (Buteobuteo) в горной местности (Аппенины Абруццо, Италия) . Дж. Раптор Рез., 31 (3), 208–212.
- ^ Бюте А., Мишель Н., Рантье Ю., Комор В., Юбер-Мой Л., Набусет Ж. и Делеттр Ю. (2010). Реакция канюка (Buteobuteo) и пустельги (Falcotinnunculus) на изменения в землепользовании в сельскохозяйственных ландшафтах Западной Франции . Сельское хозяйство, экосистемы и окружающая среда, 138 (3–4), 152–159.
- ^ Таббс, CR, и Таббс, JM (1985). Канюки Buteo Buteo и землепользование в Нью-Форест, Хэмпшир, Англия . Биологическая охрана, 31 (1), 41–65.
- ^ Грыз, Дж., и Краузе-Грыз, Д. (2019). Популяция обыкновенного канюка Buteobuteo в изменяющейся среде, Центральная Польша, на примере . Разнообразие, 11(3), 35.
- ^ Паломино, Д., и Карраскал, Л.М. (2007). Среда обитания сообщества хищников в мозаичном ландшафте Центральной Испании в условиях городской застройки . Ландшафт и городское планирование, 83 (4), 268–274.
- ^ Перейти обратно: а б Руни Э., Рид Н. и Монтгомери Висконсин (2015). Дополнительный корм увеличивает продуктивность канюка Buteobuteo, но только в условиях среды обитания низкого качества . Ибис, 157 (1), 181–185.
- ^ Стенкат Дж., Краутвальд-Юнгханнс М.Э. и Шмидт В. (2013). Причины заболеваемости и смертности свободноживущих птиц в городских условиях Германии . Экоздоровье, 10 (4), 352–365.
- ^ Пануччо М., Фоски Ф., Тодини А., Бальди А., Доминичис Н., Де Филиппис П. и Палмери А. (2019). Лучше остановиться в центре или в сельской местности? Хищные птицы зимуют на городских и сельских охраняемых территориях Рима (Центральная Италия) . Миграция раптора.
- ^ Высочин, МО (2019). Динамика численности и типы местообитаний в местах гнездования хищных птиц (Falconiformes) Донецкой гряды по градиенту антропогенной нарушенности . Регуляторные механизмы в биосистемах, 10 (4), 464-469.
- ^ Копий, Г. (2018). Экологическое распределение и плотность населения хищников во внутренней и внешней зоне центральноевропейского города . Биологический вестник Мелитопольского государственного педагогического университета имени Богдана Хмельницкого, 8(1).
- ^ Вергара, П. (2010). Смещение дневной численности и богатства хищников в зависимости от времени суток оценивается по данным дорожных обследований . Revista Catalana d'Ornitologia, 26, 22–30.
- ^ Спаар Р. и Брудерер Б. (1997). Оптимальное летное поведение парящих мигрантов: исследование мигрирующих степных канюков Buteo Buteo vulpinus . Поведенческая экология, 8 (3), 288–297.
- ^ Перейти обратно: а б Спаар, Р. (1995). Летное поведение степных канюков (Buteobuteo vulpinus) во время весенней миграции на юге Израиля: исследование с помощью радиолокатора . Израильский журнал зоологии, 41 (3), 489–500.
- ^ Портер, РФ, и Биман, MAS (1985). Возобновление миграции хищников в Европе и на Ближнем Востоке. Исследования по сохранению хищников . Техническая публикация ICBP, 5, 237–242.
- ^ Перейти обратно: а б Вучинский, А. (2003). Численность канюка (Buteobuteo) на зимовках в Центральной Европе в зависимости от погодных условий и кормовой базы . Бутео, 13, 11–20.
- ^ Уоллс, СС, и Кенвард, Р.Э. (1998). Передвижение канюков с радиометками Бутео-бутео в молодости . Ибис, 140 (4), 561–568.
- ^ Перейти обратно: а б с Шиндлер С., Хоманн У., Пробст Р., Немешкал Х.Л. и Спитцер Г. (2012). Территориальность и использование среды обитания обыкновенного канюка (Buteobuteo) поздней осенью на севере Германии . Журнал исследований хищников, 46 (2), 149–157.
- ^ Шиндлер, С. (2002). Территориальность и использование мест обитания зимующих канюков (Buteobuteo) в земле Шлезвиг-Гольштейн, Германия (Докторская диссертация).
- ^ Николов С., Спасов С. и Камбурова Н. (2006). Плотность, численность и использование мест обитания канюка обыкновенного (Buteobuteo), зимующего в низинах Болгарии . Бутео, 15, 39–47.
- ^ Балтаг, Э.С., Покора, В., Сфика, Л., и Болбоака, Л.Е. (2013). Популяция канюка (Buteobuteo) в зимний сезон на северо-востоке Румынии: влияние плотности, выбора среды обитания и погоды . Орнис Фенника, 90(3), 186.
- ^ Челлен Н. и Роос Г. (2000). Тенденции численности шведских хищников, продемонстрированные подсчетами миграций в Фальстербо, Швеция, 1942–97 гг . Исследование птиц, 47 (2), 195–211.
- ^ Перейти обратно: а б Челлен, Н. (1999). Различные миграционные стратегии среди шведских канюков Buteobuteo выявляются по доле белых птиц . Орнис svecica, 9 (1), 11–18.
- ^ Пануччо М., Дучи А., Люсия Г. и Агостини Н. (2017). Видоспецифическое поведение мигрирующих через Турецкие проливы хищников в зависимости от погоды и географии . Ардеола, 64 (2), 305–325.
- ^ Портер Р. и Уиллис И. (1968). Осенняя миграция парящих птиц над Босфором . Ибис, 110 (4), 520–536.
- ^ Лешем, Ю. (1985). Израиль: международная ось миграции хищников . ИКБП Тех. Опубл, 5, 243–250.
- ^ Шмитт, М.Б., Баур, С. и Фон Мальтиц, Ф. (1980). Наблюдения за степным канюком в Трансваале. Страус, 51(3), 151–159.
- ^ Перейти обратно: а б Херреманс, М. (2000). «Хаотичная» линька лётных перьев степного канюка Buteo Buteo vulpinus . Исследование птиц, 47(3), 332–343.
- ^ Мартин Б., Онрубия А. и Феррер Массачусетс (2014). Влияние изменения климата на миграционное поведение обыкновенного канюка Buteobuteo . Исследования климата, 60(3), 187–197.
- ^ Бенсусан, К.Дж., Гарсия, Э.Ф., и Кортес, Дж.Э. (2007). Тенденции численности мигрирующих хищников в Гибралтаре весной . Ардеа, 95(1), 83–91.
- ^ Фрайер, Г. (1986). Заметки по биологии размножения канюка . Британские птицы, 79 (1), 18–28.
- ^ Перейти обратно: а б Уоллс, С.С., и Кенвард, Р.Э. (2001). Пространственные последствия родства и возраста канюков . Поведение животных, 61 (6), 1069–1078.
- ^ Перейти обратно: а б с д и ж г Рединов, К.А. (2012). Трофические связи канюка на территории Украины . Русские птицы: 155–168.
- ^ Перейти обратно: а б с д Селас В., Твейтен Р. и Анонсен О.М. (2007). Рацион канюка (Buteo Buteo) на юге Норвегии определен по останкам добычи и видеозаписям . Орнис Фенника, 84(3), 97.
- ^ Перейти обратно: а б с д и ж г час я Зуберогойтия И., Мартинес Х.Э., Мартинес Х.А., Сабала Х., Кальво Х.Ф., Кастильо И., Аскона А., Ирата А. и Идальго С. (2006). Влияние методов управления на выбор места гнездования, размножение и рацион обыкновенного канюка Бутео-бутео в двух разных районах Испании . Ардеола, 53 (1), 83–98.
- ^ Перейти обратно: а б с д Сидорович А.А., Ивановский В.В., Сидорович В.Е., Соловей И.А. (2016). Ландшафтные изменения в составе рациона канюка обыкновенного (Buteobuteo) в Беларуси . Словацкий журнал Raptor, 10 (1), 65–74.
- ^ Спидсё, Т.К., и Селос, В. (1988). Выбор добычи и успех размножения обыкновенного канюка Buteo Buteo в сравнении с циклами мелких грызунов на юге Норвегии . Fauna Norvegica, Серия C, 11 (2), 61–66.
- ^ Перейти обратно: а б Мебс, Т. (1964). О биологии и динамике численности канюка обыкновенного (Buteo Buteo) (Особое внимание уделено зависимости массового изменения полевой мыши Microtus arvalis) . Журнал орнитологии, 105 (3), 247–306.
- ^ Перейти обратно: а б Кенвард Р.Э., Холл Д.Г., Уоллс С.С. и Ходдер К.Х. (2001). Факторы, влияющие на хищничество канюков Buteo Buteo на выпущенных фазанов Phasianus colchicus . Журнал прикладной экологии, 38 (4), 813–822.
- ^ Перейти обратно: а б с д Сим, IMW, Кросс, А.В., Ламакрафт, DL и Пейн, DJ (2001). Корреляты плотности канюка Buteo Buteo и успеха размножения в Уэст-Мидлендсе . Исследование птиц, 48(3), 317–329.
- ^ Перейти обратно: а б Галле, С. (1988). Хищничество птиц на смешанное сообщество обыкновенных полевок (Microtus arvalis) и лесных мышей (Apodemus sylvaticus) . Экология, 75(3), 451–455.
- ^ Перейти обратно: а б с д и ж г час я дж Селас, В. (2001). Хищничество обыкновенного канюка Buteo Buteo на рептилий и птиц в связи с изменениями его основной добычи — полевок . Канадский журнал зоологии, 79 (11), 2086–2093.
- ^ Андерссон, М., и Эрлинг, С. (1977). Влияние хищников на популяции грызунов . Ойкос, 591–597.
- ^ Перейти обратно: а б с д и Грэм, И.М., Редпат, С.М. и Тиргуд, С.Дж. (1995). Рацион питания и плотность размножения обыкновенного канюка Buteobuteo в зависимости от показателей обилия добычи . Исследование птиц, 42 (2), 165–173.
- ^ Франксен, Р.М.; Уиттингем, MJ; Людвиг, Южная Каролина; Роос, С.; Бейнс, Д. (2017). «Численные и функциональные реакции обыкновенных канюков Buteo Buteo на изобилие добычи на болотах шотландских тетеревов» . Ибис . 159 (3): 541–553. дои : 10.1111/ibi.12471 .
- ^ Перейти обратно: а б с д и ж г час я Байлсма, Р.Г. (2012). Рептилии как пища для молодых канюков Buteobuteo . Снасть, 20 (2), 133–144.
- ^ Перейти обратно: а б с д и ж г час Шотнар К. и Обуч Дж. (2009). Экология питания гнездовой популяции обыкновенного канюка (Buteobuteo) в районе Верхней Нитры, Центральная Словакия . Словацкий журнал Raptor, 3, 13–20.
- ^ Перейти обратно: а б с д и ж г час я Черничко И.И., Давыгора А.В. (редакторы). (2012). Канюки Северной Евразии: распространение, состояние популяции, биология . Материалы VI Международной конференции по хищным птицам и совам Северной Евразии, Кривой Рог, 27–30 сентября 2012. – Кривой Рог: Пресс- 272 с.
- ^ Перейти обратно: а б Коста, ASC (2017). Исследование питания орла-краснокрыла (Buteobuteo [Linnaeus, 1758]) и орла-карлика (Hieraetus pennatus [Gmelin, 1788]) в пробковых дубовых лесах Альто-Алентежу (докторская диссертация).
- ^ МакКомб, AM (2004). Попытки хищничества хищных птиц на серых белок Sciurus carolinensis (Gmelin) . Журнал ирландских натуралистов, 27 (12), 483–484.
- ^ Лима, С.Л., Валоне, Т.Дж., и Карако, Т. (1985). Компромисс между добычей, эффективностью, хищничеством и риском у серой белки . Поведение животных, 33 (1), 155–165.
- ^ Рамос-Лара, Н., Копровски, Дж. Л., Криштуфек, Б., и Хоффманн, И. Е. (2014). Spermophilus citellus (Rodentia: sciuridae) . Виды млекопитающих, 46 (913), 71–87.
- ^ Матросова В.А., Шнейдерова И., Володин И.А., Володина Е.В. (2012). Видоспецифичность и общие черты вокального репертуара трех евразийских сусликов (род Spermophilus) . Acta Theriologica, 57 (1), 65–78.
- ^ Макомаска-Юхевич М. и Баран П. (2012). Мониторинг видов животных . Методическое руководство. Часть III. ГИОС, Варшава, 292–496.
- ^ Перейти обратно: а б с Паломар, CJP (2005). El ratonero común (Buteo Buteo insularum) на Фуэртевентуре, Канарские острова (Птицы, Accipitridae) . Вирея, 33, 1–7.
- ^ Перейти обратно: а б с Штайнер, Х. (1999). Канюк обыкновенный (Buteobuteo) как индикатор структуры и землепользования сельской местности: продуктивность в гетерогенном ареале, влияние пищи и погоды и сравнение с ястребом-тетеревятником (Accipiter gentilis) (Т. 62) . Биологический центр Верхней Австрии. Государственный музей.
- ^ Оберг, Б., и Хангейт, Ф.П. (ред.). (2013). Радиоэкологические процессы концентрации: материалы международного симпозиума, состоявшегося в Стокгольме , 25–29 апреля 1966 г. . Elsevier.
- ^ Шапюи, JL (2006). Тамиас сибирский . Инвентаризация чужеродных инвазивных видов.
- ^ Перейти обратно: а б с Пиновски Дж. и Рышковски Л. (1962). Универсальность канюка как хищника . Британские птицы, 55, 470–475.
- ^ Тойран, К., и Албайрак, И. (2009). Вклад в биологические характеристики Allactaga williamsi Thomas, 1897 в провинции Кырыккале (Mammalia: Rodentia) . Международный журнал естественных и технических наук, (1), 13–17.
- ^ Найтингейл, В. (1996). Соня (Glis glis; Mammalia: Rodentia: Gliridae) как добыча обыкновенного канюка (Buteo Buteo, Aves: Falconiformes: Accipitridae) . (Труды к фауне млекопитающих Саксонии, № 1). Фаунистические трактаты, 20: 320.
- ^ Гвинн, Л. (2015). Идентичность, происхождение и влияние «нового» вида канюка, размножающегося в Южной Африке (докторская диссертация, Кейптаунский университет).
- ^ Перейти обратно: а б с д и ж г час я дж Суонн Р.Л. и Этеридж Б. (1995). Сравнение успеха размножения и добычи обыкновенного канюка Бутео-бутео в двух районах северной Шотландии . Исследование птиц, 42 (1), 37–43.
- ^ Перейти обратно: а б Свон, Г. (2011). Пространственные различия в успехе размножения обыкновенного канюка Buteo Buteo в зависимости от типа среды обитания и рациона питания (докторская диссертация, факультет наук о жизни, Силвуд-Парк, Имперский колледж Лондона).
- ^ Перейти обратно: а б с д Руни Э. и Монтгомери Висконсин (2013). Разнообразие рациона обыкновенного канюка (Buteobuteo) в среде, где нет полевок . Исследование птиц, 60 (2), 147–155.
- ^ Перейти обратно: а б с д и ж г Маноза С. и Кордеро П.Дж. (1992). СЕЗОННЫЕ И ПОЛОВЫЕ ИЗМЕНЕНИЯ В ПИТАНИИ ОБЫКНОВЕННОГО КАНЮКА В СЕВЕРО-ВОСТОЧНОЙ ИСПАНИИ . Дж. Раптор Рез., 26 (4), 235–238.
- ^ Перейти обратно: а б Уоллс, Шон и Роберт Кенвард. Обыкновенный канюк. Издательство Блумсбери, 2020.
- ^ Рейф В., Торнберг Р., Юнгелл С. и Корпимяки Э. (2001). Вариации в рационе обыкновенных канюков в Финляндии подтверждают гипотезу об альтернативной добыче . Экография, 24(3), 267–274.
- ^ Тобахас, Хорхе и др. «Влияет ли численность добычи филина (Bubo bubo) на выбор размера добычи?». Орнитологический журнал 162.3 (2021): 699-708.
- ^ Скерчинский, М. (2006). Перекрытие пищевой ниши трех симпатрических хищников, размножающихся в сельскохозяйственном ландшафте Западно-Поморского региона Польши .
- ^ Перейти обратно: а б Сциллитани, Г. (1999). Рацион канюка Бутео бутео (L., 1758) в провинции Бари . Авосет, 23: 169.
- ^ Корпимяки, Э., и Норрдал, К. (1989). Птичье хищничество куньих в Европе 1: возникновение и влияние на изменение размеров тела и жизненные особенности . Ойкос, 205–215.
- ^ Франксен, Р.М., Уиттингем, М.Дж., и Бэйнс, Д. (2016). Оценка добычи птенцов обыкновенного канюка Buteobuteo: сравнение методов . Исследование птиц, 63(3), 303–310.
- ^ Сельва Н., Енджеевска Б., Енджеевски В. и Вайрак А. (2003). Сбор туш зубров в девственной Беловежской пуще (восточная Польша) . Эконаука, 10(3), 303–311.
- ^ Перейти обратно: а б с д и ж Серджио Ф., Бото А., Скандолара К. и Больяни Г. (2002). Плотность, места гнездования, рацион и продуктивность канюка (Buteobuteo) в итальянских предальпийских регионах . Журнал исследований хищников, 36 (1), 24–32.
- ^ Пэрротт, Д. (2015). Воздействие и борьба с канюками Buteo Buteo и загонами для выпуска фазанов Phasianus colchicus в Великобритании: обзор . Европейский журнал исследований дикой природы, 61 (2), 181–197.
- ^ Бунман, М. и де Врислаан, Х. (1992). [Калюк Бутео Бутео осматривает гнездо серой цапли Ardea cinerea] . Лимоза, 65: 29–30.
- ^ Джонсгард, Пенсильвания (1983). Журавли мира: Черношейный журавль (Grus nigricollis) . Журавли мира, Пол Джонсгард, Том. 5.
- ^ Монтейро, Л.Р., и Фернесс, Р.В. (1998). Видообразование посредством временной сегрегации популяций мадейрского кастро (Oceanodroma castro) на Азорских островах? Философские труды Лондонского королевского общества. Серия B: Биологические науки, 353(1371), 945–953.
- ^ Рамос, Дж. А., Монис, З., Сола, Э., и Монтейро, Л. Р. (2003). Репродуктивные меры и обеспечение птенцами буревестника Кори Calonectris diomedea Borealis на Азорских островах . Исследование птиц, 50 (1), 47–54.
- ^ Хой, Х., Хой, К., Кристофик, Дж., и Даролова, А. (2002). Репродуктивный успех снижается с увеличением размера семьи у европейской щурки . Этология, экология и эволюция, 14 (2), 99–110.
- ^ Мартин, Дж. (1990). Земноводные и рептилии как добыча птиц в Юго-Западной Европе . Смитсоновская герпетологическая информационная служба.
- ^ Бустаманте, Дж. (1985). Кормление канюка обыкновенного (Buteobuteo, L. 1758) в Северной Испании . Донана, Acta Vertebrata, 12, 51–62.
- ^ Мик, Р. (2013). Движения заливной гадюки после спячки Vipera aspis . Герпетологический вестник, 125, 22–24.
- ^ Эттлинг, Дж. А., Агасян, Л. А., Агасян, А. Л., и Паркер, П. Г. (2013). Пространственная экология армянских гадюк Montivipera raddei в измененном человеком ландшафте . Копейя, 2013(1), 64–71.
- ^ Спеллерберг, IF (1975). Уж в Великобритании . Орикс, 13 (2), 179–184.
- ^ Гатри, Дж. Э. (1932). Змеи против птиц; птицы против змей . Бюллетень Вильсона, 44 (2), 88–113.
- ^ Эш, CP, и Аткинс, JD (2009). Птицы Эфиопии и Эритреи: атлас распространения . А&С Черный.
- ^ Ирвин, MPS (1981). Птицы Зимбабве . Квест Паб.
- ^ Лонг, RC (1960). Птицы округа Порт-Геральд.(Часть I) . Страус, 31(3), 85–104.
- ^ Невель, Дж. и Уинтерс, Б. (1987). [Калюк Бутео-бутео ест угря] . Лимоза, 60:149.
- ^ Мачковски, К. и Тряновски, П. (1993). Карп (Cyprinus carpio) в пище канюка (Buteobuteo) . Пшеглад Пширодничи, 42:76.
- ^ Дэвис, TAW (1960). Погадки пустельги на зимнем ночлеге . Брит. Птицы, 53 (7), 281–284.
- ^ Редпат С.М., Кларк Р., Мэддерс М. и Тергуд С.Дж. (2015). Оценка рациона хищников: сравнение погадок, останков добычи и данных наблюдений за гнездами луней-куриц .
- ^ Лохмус, А., и Вали, Ю. (2004). Влияние качества среды обитания и размера самки на продуктивность малого подорлика Aquila pomarina в свете гипотезы альтернативной добычи . Журнал птичьей биологии, 35 (5), 455–464.
- ^ Менье, Ф.Д., Верхейден, К., и Жувентен, П. (2000). Использование обочин дорог дневными хищниками в сельскохозяйственных ландшафтах . Биологическая охрана, 92 (3), 291–298.
- ^ Бергман, Г. (1961). Питание хищных птиц и сов в Фенно-Скандии . Британские птицы, 54, 307–320.
- ^ Янковяк Л. и Тряновски П. (2013). Совместное присутствие и перекрытие пищевых ниш двух обычных хищников (рыжая лисица Vulpes vulpes и канюк Buteo Buteo) в сельскохозяйственном ландшафте . Турецкий журнал зоологии, 37 (2), 157–162.
- ^ Сильвен, М. (1978). Межвидовые отношения симпатрически зимующих канюков Buteo Buteo и зимняков Buteo lagopus . Орнис Скандинавица, 197–206.
- ^ Кассинис, Н. (2009). Гнездовое распространение и численность канюка Buteo rufinus rufinus на Кипре . Авосетта, 33, 75–78.
- ^ Бакалоудис, Д.Э., Иезекииль, С., Влахос, К.Г., Бонцорлос, В.А., Папакоста, М., и Биррер, С. (2012). Оценка предвзятости в методах питания усача Buteo rufinus . Журнал засушливой среды, 77, 59–65.
- ^ Крюгер, О. (2002). Анализ занятости гнезд и воспроизводства гнезд у двух симпатрических хищников: канюка Buteo Buteo и ястреба-тетеревятника Accipiter gentilis . Экография, 25(5), 523–532.
- ^ Крюгер, О. (2002). Взаимодействие между обыкновенным канюком Buteo Buteo и ястребом-тетеревятником Accipiter gentilis: компромиссы, выявленные в ходе полевого эксперимента . Ойкос, 96(3), 441–452.
- ^ Тейлор К., Хадсон Р. и Хорн Г. (1988). Распространение и численность гнездящихся канюков в Великобритании и Северной Ирландии в 1983 году . Исследование птиц, 35 (2), 109–118.
- ^ Костшева, А. (1991). Межвидовая интерференционная конкуренция у трех европейских видов хищных птиц . Этология, экология и эволюция, 3 (2), 127–143.
- ^ Перейти обратно: а б Хаккарайнен Х., Микра С., Курки С., Торнберг Р. и Юнгелл С. (2004). Конкурентные взаимоотношения хищников в бореальных лесах . Экология, 141(3), 420–424.
- ^ Гощиньский, Дж. (1997). Плотность и продуктивность популяций канюка Buteo Buteo и ястреба-тетеревятника Accipiter gentilis в Рогове, Центральная Польша . Орнитологический закон, 32 (2), 149–155.
- ^ Гатто, Анджела Э.; Грабб, Терил Г.; Чемберс, Кэрол Л. (2006). «Питание краснохвостого ястреба совпадает с рационом северных ястребов-тетеревятников на плато Кайбаб, штат Аризона» . Журнал исследований хищников . 39 (4): 439–444.
- ^ Боал, CW (2005). «Продуктивность и смертность ястребов-тетеревятников в Миннесоте» . Дж. Раптор Рез . 39 (3): 222–228.
- ^ Люттич С., Раш Д.Х., Меслоу Э.К. и Кейт Л.Б. (1970). «Экология хищничества краснохвостого ястреба в Альберте». Экология . 51 (2): 190–203. Бибкод : 1970Ecol...51..190L . дои : 10.2307/1933655 . JSTOR 1933655 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Серджио Ф. и Бото А. (1999). Распространение гнезд, рацион питания и успех размножения черных коршунов (Milvus migrans) в итальянских предальпийских районах . Журнал исследований хищников, 33, 207–217.
- ^ Мейбург, BU (1973). Исследования менее знакомых птиц: 172. Малый подорлик . Британские птицы.
- ^ Перейти обратно: а б Мур, Северо-Запад (1957). Прошлое и настоящее положение канюка на Британских островах . Британские птицы, 50, 173–197.
- ^ Диксон, Н., и Гиббс, А. (2018). Чрезвычайная территориальная агрессия городских сапсанов по отношению к обыкновенным канюкам на юго-западе Англии . Орнис Хунгарика, 26(2), 232-242.
- ^ О'Коннор, TP (1993). Птицы и ниша падальщиков . Археофауна, (2).
- ^ Перейти обратно: а б Серхио Ф. и Хиральдо Ф. (2008). Внутригильдийное хищничество в сообществах хищных птиц: обзор птиц . Ибис, 150, 132–145.
- ^ Перейти обратно: а б Бьёрклунд Х., Сантанджели А., Бланше Ф.Г., Уиту О., Лехторанта Х., Линден Х., Валкама Дж. и Лааксонен Т. (2016). Внутригильдийное хищничество и конкуренция влияют на подчиненных хищников птиц . Экология, 181(1), 257–269.
- ^ Вайс, Н., и Джозеф, Р. (2010). Степной орел (Aquila nipalensis) охотится на канюка (Buteobuteo vulpinus) во время миграции над Эйлатом, Израиль . Журнал исследований хищников, 44 (1), 77–79.
- ^ Тодд, DM (1980). Беркут убивает канюка . Британские птицы, 73: 536–537.
- ^ Чавко Дж., Данко Ш., Обуч Дж. и Михок Дж. (2007). Питание орла-могильника (Aquila heliaca) в Словакии . Словацкий журнал Raptor, 1, 1–18.
- ^ Хорват М., Шолти Б., Фатер И., Юхас Т., Харасти Л., Ситта Т., Баллок З. и Пастори-Ковач С. (2018). Временные изменения в составе рациона восточного орла-могильника (Aquila heliaca) в Венгрии . Орнис Хунгарика, 26 (1), 1–26.
- ^ Градоц, П. (1996). [Обыкновенный канюк Buteobuteo, пойманный в полете орланом-белохвостом Haliaetus albicilla.] 62: 165–166.
- ^ Ресано Дж., Бэйл П., Реал Дж., Эрнандес А., Винсент-Мартин Н. и Равайрол А. (2012). Анализ рациона орла Бонелли Hiraaetus fasciatus (Vieillot, 1822) в период размножения 2010 года во Франции . Университет Барселоны – Оборудование биологии консервации, 1: 95–101.
- ^ Дементавичюс, Д. (2004). Обыкновенный канюк (Buteobuteo) и орлан-белохвост (Haliaeetus albicilla): гнездовой паразитизм или нетипичное пищевое поведение? Acta Zoologica Lituanica, 14 (1), 76–79.
- ^ Литерак И. и Мраз Дж. (2011). Усыновление молодых канюков в гнездах орлана-белохвоста . Орнитологический журнал Уилсона, 123 (1), 174–176.
- ^ Камараускайте А., Дементавичюс Д., Скуя С., Дагис М. и Трейнис Р. (2020). Взаимодействие орлана-белохвоста и канюка оценивается на основе анализа рациона и защитного поведения выводка . Орнис Фенника, 97(1), 26–37.
- ^ Дементавичюс Д., Румбутис С., Вирбицкас Т., Вайткувене Д., Дагис М. и Трейнис Р. (2020). Пространственные и временные изменения в гнездовом рационе орлана-белохвоста Haliaeetus albicilla, выявленные по остаткам добычи . Исследование птиц, 67 (2), 206–216.
- ^ Лоуренсо, Р., Рабаса, Дж. Э., Дельгадо, МДМ, и Пентериани, В. (2009). Внутригильдийное хищничество со стороны главного европейского хищника – филина .
- ^ Чакаров Н. и Крюгер О. (2010). Выпуск месохищника появившимся суперхищником: естественный эксперимент хищничества в трехуровневой гильдии . ПЛОС ОДИН, 5(12), е15229.
- ^ Земанова, К. (2009). Пища большой рогатой совы в Центральной Моравии .
- ^ Бэйл П. и Шаулс Р. (2011). Биология четырех пар Великого Герцога Европы Бубо-бубо в Люксембурге . Бык. Соц. Нат. Люксембург, 112, 51.
- ^ Шиндлер, С. (2002). Территориальность и использование мест обитания зимующих канюков (Buteobuteo) в земле Шлезвиг-Гольштейн, Германия (Докторская диссертация, uniwien).
- ^ Эйвери, Г., Робертсон, А.С., Палмер, Н.Г., и Принс, А.Дж. (1985). Добыча гигантских филинов в заповеднике де Хооп, Капская провинция, и некоторые наблюдения по стратегии охоты . Страус, 56(1–3), 117–122.
- ^ Бышнев, И.И. (2002). Интересен случай агрессивного взаимодействия обыкновенного канюка (Buteobuteo) и уральской совы (Strix uralensis) . Субутео, 5: 46.
- ^ Перейти обратно: а б с Остин, GE, и Хьюстон, округ Колумбия (1997). Результаты размножения канюка Бутео-бутео в Аргайле, Шотландия, и сравнение с другими районами Великобритании . Исследование птиц, 44 (2), 146–154.
- ^ Таббс, CR (1972). Анализ карт учета гнезд канюка . Исследование птиц, 19 (2), 97–104.
- ^ Сара, М., Кампобелло, Д., и Занка, Л. (2012). Влияние особенностей гнезда и колонии на репродуктивный успех пустельги (Falco naumanni) . Исследования биологии птиц, 5 (4), 209–217.
- ^ Миккола, Х. (1976). Совы убивают и убивают другие совы и хищники в Европе . Британские птицы, 69, 144–154.
- ^ Ван Ньювенхейз, Д., Жено, Дж. К., и Джонсон, Д. Х. (ред.). (2008). Маленькая сова: сохранение, экология и поведение Афины-ноктуа . Издательство Кембриджского университета, Эдинбургское здание, Шафтсбери-роуд, Кембридж CB 2 2 RU, Великобритания.
- ^ Барнард. СС (1981). Канюк охотится на ушастую сову . Британские птицы, 74: 226.
- ^ Вус, К.Х. 1988. Совы Северного полушария . MIT Press, 0262220350.
- ^ Сунде, П. (2005). Хищники контролируют смертность коричневых сов Strix aluco после оперения . Ойкос, 110(3), 461–472.
- ^ Перейти обратно: а б Крюгер, О. (2004). Важность конкуренции, пищи, среды обитания, погоды и фенотипа для размножения канюка Buteo Buteo . Исследование птиц, 51 (2), 125–132.
- ^ Новикова, Л. (2009). Численность и распространение обыкновенного канюка в Керженском государственном заповеднике, Россия . Сохранение хищников, (16).
- ^ Дэйр, П.Дж., и Барри, Дж.Т. (1990). Размер популяции, плотность и регулярность расположения гнезд канюков Бутео-бутео в двух горных регионах Северного Уэльса . Исследование птиц, 37 (1), 23–29.
- ^ Перейти обратно: а б Родригес Б., Сиверио Ф., Родригес А., Сиверио М., Эрнандес Дж. Дж. и Фигерола Дж. (2010). Плотность, выбор среды обитания и биология размножения обыкновенных канюков Buteobuteo в островной среде . Исследование птиц, 57 (1), 75–83.
- ^ Цорцакаки О., Симаякис С. и Ксирушакис С. (2012). Обилие канюков (Buteobuteo) в монокультурах оливок на острове Крит . Журнал биологических исследований, 17, 44.
- ^ Черасоли, М., и Пентериани, В. (1996). Канюки обыкновенные (Buteobuteo) в Центральной Италии . Дж. Раптор Рез., 30 (3), 130–135.
- ^ Шотнар К. и Топерсер Дж. (2009). Оценка плотности, численности и динамики численности канюка (Buteobuteo) в Западно-Карпатском регионе новым методом . Словацкий журнал Raptor, 3, 1–12.
- ^ Воржишек, П. (2000). Чрезвычайно высокая плотность популяции обыкновенного канюка (Buteobuteo) в биосферном заповеднике Палава (Чехия) и ее возможные причины . Бутео, 11, 51–56.
- ^ Форсман Д. и Солонен Т. (1984, январь). Учет гнездящихся хищных птиц на юге Финляндии: методы и результаты . В Annales Zoologici Fennici (стр. 317–320). Академия наук Финляндии, Societas Scientiarum Fennica, Societas pro Fauna et Flora Fennica и Societas Biologica Fennica Vanamo.
- ^ Негро, Джей Джей, и Гальван, И. (2018). Поведенческая экология хищников . В «Хищных птицах» (стр. 33–62). Спрингер, Чам.
- ^ Симмонс, Р.Э., и Мендельсон, Дж.М. (1993). Критический обзор кувыркающихся полетов хищников . Страус, 64(1), 13–24.
- ^ Барриентос, Р., и Лопес-Дариас, М. (2006). Случай полиандрической трио евразийских канюков (Buteobuteo) на острове Фуэртевентура, Канарские острова . Журнал исследований хищников, 40 (4), 305–306.
- ^ Перейти обратно: а б Рокенбаух, Д. (1975). Двенадцатилетние исследования экологии обыкновенного канюка (Buteobuteo) в Швабском Альбе . Журнал орнитологии, 116 (1), 39–54.
- ^ Остин, GE (1992). Распространение и эффективность размножения канюка Buteo Buteo в зависимости от среды обитания: применение с использованием дистанционного зондирования и географических информационных систем (докторская диссертация, Университет Глазго).
- ^ Джонкерс, Д.А., и Рудхарт, Дж. (2003). Наземное гнездо канюка Бутео-бутео на мосту . Снасти, 11 (3), 234–236.
- ^ Касл-Гомез, К., и Морено-Уил, Г. (2011). Запись о гнездовании обыкновенного канюка (Buteobuteo) в заброшенном здании . Журнал исследований хищников, 45 (3), 275–277.
- ^ Перейти обратно: а б Селас, В. (2001). Плотность гнездования и размер выводка обыкновенного канюка Buteobuteo в зависимости от снежного покрова весной . Ардеа, 89 (3), 471–479.
- ^ Холдсворт, М. (1971). Биология размножения канюков в Седберге в 1937–67 гг . Британские птицы, 64, 412–420.
- ^ Перейти обратно: а б Тайак, Эй.Дж., Уоллс, С.С. и Кенвард, Р.Э. (1998). Поведение обыкновенных канюков с радиометками Buteo Buteo в период зависимости после птенцов . Ибис, 140 (1), 58–63.
- ^ Уоллс, С.С., Маньоса, С., Фуллер, Р.М., Ходдер, К.Х., и Кенвард, Р.Э. (1999). Ранний разгон — предприятие или изгнание? Свидетельства канюков с радиометками . Журнал птичьей биологии, 407–415.
- ^ Херреманс, М. (2001). Возрастные различия в численности и местообитаниях степных канюков, Buteobuteo vulpinus, в Западно-Капской провинции, Южная Африка . Страусиный журнал африканской орнитологии, 72 (1–2), 105–108.
- ^ Перейти обратно: а б Крюгер О. и Линдстрем Дж. (2001). Пожизненный репродуктивный успех обыкновенного канюка Buteo Buteo: от индивидуальных вариаций к демографии популяции . Ойкос, 93(2), 260–273.
- ^ Чакаров Н., Бёрнер М. и Крюгер О. (2008). Приспособленность обыкновенных канюков на стыке противоположных взаимодействий меланина и паразита . Функциональная экология, 22 (6), 1062–1069.
- ^ Перейти обратно: а б Крюгер, О. (2002). Анализ продолжительности жизни канюка и репродуктивного успеха в течение всей жизни: относительная важность еды, конкуренции, погоды, среды обитания и индивидуальных качеств . Экология, 133(4), 474–482.
- ^ Мур, Северо-Запад (1956). Кролики, канюки и зайцы. Два исследования косвенных эффектов миксоматоза . Биологические последствия обусловлены наличием миксоматоза.
- ^ Лиз, AC, и Белл, DJ (2008). Природоохранный парадокс XXI века: европейский дикий кролик Oryctolagus cuniculus, инвазивный чужеродный и находящийся под угрозой исчезновения местный вид . Обзор млекопитающих, 38(4), 304–320.
- ^ Дэвис, PE, и Дэвис, JE (1992). Распространение и возраст первого размножения канюков в центральном Уэльсе . Британские птицы, 85 (11), 578–587.
- ^ Перейти обратно: а б Клементс, Р. (2002). Обыкновенный канюк в Великобритании . Британские птицы, 95, 377–383.
- ^ Йонкер, Р.М., Чакаров, Н., и Крюгер, О. (2014). Изменение климата и неоднородность среды обитания приводят к увеличению популяции обыкновенных канюков Buteo Buteo из-за воздействия на выживаемость . Ибис, 156 (1), 97–106.
- ^ Эллиотт, Г.Д., и Эйвери, Мичиган (1991). Обзор сообщений о преследовании канюков в 1975–1989 гг . Исследование птиц, 38 (1), 52–56.
- ^ Янсс, GF (2000). Птичья смертность от линий электропередач: морфологический подход к видовой смертности. Биологическая охрана, 95(3), 353–359.
- ^ Наккари К., Кристани М., Чимино Ф., Аркорачи Т. и Тромбетта Д. (2009). Канюки обыкновенные (Buteobuteo) — биоиндикаторы загрязнения тяжелыми металлами на Сицилии (Италия) . Международная защита окружающей среды, 35(3), 594–598.
- ^ Карнейро М., Коласу Б., Брандао Р., Феррейра К., Сантос Н., Соэйру В. и Лавин С. (2014). Биомониторинг тяжелых металлов (Cd, Hg и Pb) и металлоидов (As) с помощью португальского канюка (Buteobuteo) . Экологический мониторинг и оценка, 186(11), 7011–7021.
- ^ Ягер, Л.П., Рейньерс, Ф.В., Эсселинк, Х., и Баарс, А.Дж. (1996). Биомониторинг с использованием канюка Бутео-бутео в Нидерландах: тяжелые металлы и источники вариаций . Журнал für Ornithologie, 137 (3), 295–318.
- ^ Маноза С., Матео Р., Фрейша К. и Гитарт Р. (2003). Стойкие хлорорганические загрязнители в яйцах ястреба-тетеревятника и канюка из северо-восточной Испании: временные тенденции, связанные с изменениями в рационе . Загрязнение окружающей среды, 122(3), 351–359.
- ^ Таббс, CR (1967). Исследование популяции канюков в Нью-Форесте в 1962–66 гг . Британские птицы, 60 (10), 381–395.
- ^ Рэтклифф, Д.А. (1967). Уменьшение массы яичной скорлупы у некоторых хищных птиц . Природа, 215(5097), 208.
- ^ Санторо, М., Трипепи, М., Кинселла, Дж. М., Панебьянко, А., и Маттиуччи, С. (2010). Гельминтозная инвазия хищных птиц (Accipitriformes и Falconiformes) в Южной Италии . Ветеринарный журнал, 186 (1), 119–122.
- ^ Ширази, С., Хесараки, С., Мостафаи, Т.С. и Давуди, Дж. (2014). Первое сообщение о Centrorhynchus aluconis у канюка (Buteobuteo) на северо-западе Ирана . Закон о ветеринарии, 64 (2), 276–280.
- ^ ОЗМЕН, О., и ДОРРЕШТЕЙН, генеральный менеджер (2002). Авипокс канюка обыкновенного (Buteobuteo) . Турецкий журнал ветеринарии и зоотехники, 26 (5), 1193–1195.
- ^ Фрелих К., Прусас К., Шеттлер Э. и Хафез Х.М. (2002). Антитела к аденовирусам у свободноживущих канюков из Германии . Журнал болезней диких животных, 38 (3), 633–636.
Общие источники
[ редактировать ]- Малларни, Киллиан; Свенссон, Ларс; Зеттерстром, Дэн; Грант, Питер (1999). Путеводитель по птицам Коллинза . Лондон: ХарперКоллинз. ISBN 978-0-00-219728-1 .
- Синклер, Ян; Хоккей, Фил; Тарботон, Уорик (2002). САСОЛ Птицы Южной Африки . Струйк. ISBN 978-1-86872-721-6 .
Внешние ссылки
[ редактировать ]- Текст о видах степного канюка в Атласе птиц Южной Африки
- Птицы Мадейры: Канюк . Страница о спорном подвиде хартерти . Проверено 28 ноября 2006 г.
- Старение и определение пола (PDF; 4,2 МБ), авторы: Хавьер Бласко-Зумета и Герд-Майкл Хайнце
- Перья канюка (Buteo Buteo). Архивировано 12 июня 2013 года в Wayback Machine.
- Информационный бюллетень о видах BirdLife для Бутео-бутео
- «Бутео бутео» . Авибаза .
- «Евразийские канюковые СМИ» . Интернет-коллекция птиц .
- Фотогалерея канюка в VIREO (Университет Дрекселя)
- Аудиозаписи канюка на Xeno-canto .
| Базы данных органов управления : Национальные |
|---|