Каталитический интрон группы I
| Каталитический интрон группы I | |
|---|---|
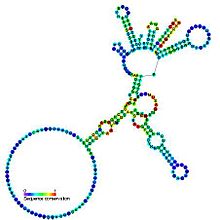 Предсказанная вторичная структура и сохранение последовательности каталитического интрона группы I | |
| Идентификаторы | |
| Символ | Интрон_gpI |
| Рфам | RF00028 |
| Другие данные | |
| РНК Тип | Интрон |
| Домен(ы) | Эукариоты ; Бактерии ; Вирусы |
| ИДТИ | ИДТИ: 0000372 |
| ТАК | ТАК: 0000587 |
| PDB Структуры | ПДБе |
Интроны группы I представляют собой крупные самосплайсинговые рибозимы . Они катализируют собственное удаление мРНК , тРНК и предшественников рРНК у широкого круга организмов. [ 1 ] [ 2 ] [ 3 ] Основная вторичная структура состоит из девяти парных регионов (P1-P9). [ 4 ] Они складываются по существу в два домена - домен P4-P6 (образованный в результате укладки спиралей P5, P4, P6 и P6a) и домен P3-P9 (образованный из спиралей P8, P3, P7 и P9). [ 2 ] Разметка вторичной структуры для этого семейства представляет только это консервативное ядро. группы I Интроны часто имеют длинные открытые рамки считывания, вставленные в петлевые области .
Катализ
[ редактировать ]Сплайсинг интронов группы I осуществляется посредством двух последовательных реакций переэтерификации . [ 3 ] Сначала экзогенный гуанозин или гуанозиновый нуклеотид ( exoG ) стыкуется с активным сайтом G-связывания, расположенным в P7, а затем его 3'-OH выравнивается, чтобы атаковать фосфодиэфирную связь в «выше» (ближе к 5'-концу) сплайсинга. сайт расположен в P1, в результате чего на верхнем экзоне появляется свободная 3'-ОН-группа , а экзоG присоединяется к 5'-концу интрона. Затем концевая G (омега G) интрона заменяет exoG и занимает сайт связывания G, подготавливая вторую реакцию переноса эфира: 3'-ОН-группа верхнего экзона в P1 выравнивается, чтобы атаковать нижестоящий сплайсинг. сайт в P10, что приводит к лигированию соседних верхних и нижних экзонов и высвобождению каталитического интрона.

механизм с двумя ионами металлов, наблюдаемый в протеин- полимеразах и фосфатазах, Было предложено использовать интронами группы I и группы II для обработки реакций переноса фосфорила. [ 5 ] что было однозначно доказано структурой интрона I группы Azoarcus с высоким разрешением в 2006 году. [ 6 ]


Интрон сворачивается
[ редактировать ]С начала 1990-х годов ученые начали изучать, как интрон группы I приобретает свою нативную структуру in vitro некоторые механизмы сворачивания РНК. , и к настоящему времени оценены [ 10 ] Принято считать, что третичная структура складывается после образования вторичной структуры. Во время сворачивания молекулы РНК быстро заселяются в различные промежуточные соединения сворачивания, промежуточные соединения, содержащие нативные взаимодействия, далее сворачиваются в нативную структуру посредством быстрого пути сворачивания, тогда как те, которые содержат ненативные взаимодействия, захватываются метастабильными или стабильными ненативными конформациями, и процесс преобразования в нативную структуру происходит очень медленно. Очевидно, что интроны I группы, различающиеся набором периферических элементов, обладают разным потенциалом вступления в путь быстрого сворачивания. Между тем, совместная сборка третичной структуры важна для сворачивания нативной структуры. Тем не менее, сворачивание интронов группы I in vitro сталкивается как с термодинамическими , так и с кинетическими проблемами. Было показано, что некоторые РНК-связывающие белки и шапероны способствуют сворачиванию интронов группы I in vitro и у бактерий путем стабилизации нативных промежуточных продуктов и дестабилизации ненативных структур соответственно.
Распространение, филогения и подвижность
[ редактировать ]Интроны I группы распространены у бактерий, низших эукариот и высших растения. Однако их встречаемость у бактерий, по-видимому, более спорадична, чем у низших видов. эукариоты, и они стали широко распространены у высших растений. Гены I группы прерывания интронов существенно различаются: они прерывают рРНК , мРНК и тРНК. гены в бактериальных геномах, а также в митохондриях и хлоропластах геномы низших эукариот, а вторгаются только в гены рРНК в ядерном геноме низшие эукариоты. У высших растений эти интроны, по-видимому, ограничены несколькими Гены тРНК и мРНК хлоропластов и митохондрий.
Интроны группы I также обнаруживаются в генах широкого спектра бактериофагов грамположительных бактерий . [ 11 ] Однако их распространение в фаге грамотрицательных бактерий ограничивается в основном Т4 , Т-четными и Т7-подобными бактериофагами. [ 11 ] [ 12 ] [ 13 ] [ 14 ]
Теории как интронно-раннего, так и интронно-позднего нашли доказательства, объясняющие происхождение интронов группы I. Некоторые интроны группы I кодируют хоминг-эндонуклеазу (HEG), которая катализирует подвижность интронов. Предлагается, чтобы HEG переместили интрон из одного места в другое, из одного организма в другой и, таким образом, ответственны за широкое распространение эгоистических интронов I группы. Никакой биологической роли не выявлено. идентифицированы для интронов группы I, за исключением сплайсинга самих себя с предшественником чтобы предотвратить смерть хозяина, у которого они живут. Небольшое количество интронов группы I Также обнаружено, что они кодируют класс белков, называемых матуразами, которые облегчают сплайсинг интронов .
См. также
[ редактировать ]- Интрон
- База данных последовательностей и структур интронов группы I
- Место сращивания
- Ядерные интроны
- Интрон группы II
- Интрон группы III
- Твинтрон
- ЛтрА
- Циклический рибопереключатель ди-GMP-II , где есть пример рибопереключателя, действующего вместе с интроном группы I для регулирования экспрессии гена.
Ссылки
[ редактировать ]- ^ Нильсен Х, Йохансен С.Д. (2009). «Интроны группы I: движение в новых направлениях» . РНК Биол . 6 (4): 375–83. дои : 10.4161/rna.6.4.9334 . ПМИД 19667762 . Проверено 15 июля 2010 г.
- ^ Jump up to: а б Кейт Дж.Х., Гудинг А.Р., Поделл Э. и др. (сентябрь 1996 г.). «Кристаллическая структура домена рибозима группы I: принципы упаковки РНК». Наука . 273 (5282): 1678–85. Бибкод : 1996Sci...273.1678C . дои : 10.1126/science.273.5282.1678 . ПМИД 8781224 . S2CID 38185676 .
- ^ Jump up to: а б Чех ТР (1990). «Самосплайсинг интронов группы I». Анну. Преподобный Биохим . 59 : 543–68. дои : 10.1146/annurev.bi.59.070190.002551 . ПМИД 2197983 .
- ^ Вудсон С.А. (июнь 2005 г.). «Строение и сборка интронов группы I». Курс. Мнение. Структура. Биол . 15 (3): 324–30. дои : 10.1016/j.sbi.2005.05.007 . ПМИД 15922592 .
- ^ Стейтц, Т.А.; Стейц Дж. А. (1993). «Общий механизм каталитической РНК с двумя ионами металлов» . Proc Natl Acad Sci США . 90 (14): 6498–6502. Бибкод : 1993PNAS...90.6498S . дои : 10.1073/pnas.90.14.6498 . ПМК 46959 . ПМИД 8341661 .
- ^ Стейли, MR; Штробель С.А. (2006). «Сплайсинг РНК: кристаллические структуры интронов группы I раскрывают основу выбора места сплайсинга и катализа ионов металлов». Curr Opin Struct Biol . 16 (3): 319–326. дои : 10.1016/j.sbi.2006.04.005 . ПМИД 16697179 .
- ^ Голден Б.Л., Гудинг А.Р., Поделл Э.Р., Чех Т.Р. (1998). «Предварительно организованный активный центр в кристаллической структуре рибозима Tetrahymena». Наука . 282 (5387): 259–64. Бибкод : 1998Sci...282..259G . дои : 10.1126/science.282.5387.259 . ПМИД 9841391 .
- ^ Золотой Б.Л., Ким Х., Чейз Э. (2005). «Кристаллическая структура комплекса рибозим-продукт фага Twort группы I». Nat Struct Мол Биол . 12 (1): 82–9. дои : 10.1038/nsmb868 . ПМИД 15580277 . S2CID 33369317 .
- ^ Го Ф, Гудинг А.Р., Чех Т.Р. (2004). «Структура рибозима Tetrahymena: тройной сэндвич основания и ион металла в активном центре» . Мол Клетка . 16 (3): 351–62. doi : 10.1016/j.molcel.2004.10.003 . ПМИД 15525509 .
- ^ Брайон П., Вестхоф Э (1997). «Иерархия и динамика сворачивания РНК». Annu Rev Biophys Biomol Struct . 26 : 113–37. doi : 10.1146/annurev.biophys.26.1.113 . ПМИД 9241415 .
- ^ Jump up to: а б Эджелл Д.Р., Белфорт М. , Шуб Д.А. (октябрь 2000 г.). «Барьеры на пути интронной распущенности у бактерий» . Дж. Бактериол . 182 (19): 5281–9. дои : 10.1128/jb.182.19.5281-5289.2000 . ПМК 110968 . ПМИД 10986228 .
- ^ Сандегрен Л., Сьёберг Б.М. (май 2004 г.). «Распределение, гомология последовательностей и хоминг интронов группы I среди Т-четных бактериофагов: свидетельства недавнего переноса старых интронов» . Ж. Биол. Хим . 279 (21): 22218–27. дои : 10.1074/jbc.M400929200 . ПМИД 15026408 .
- ^ Бонокора Р.П., Шуб Д.А. (декабрь 2004 г.). «Интрон группы I самосплайсинга в генах ДНК-полимеразы Т7-подобных бактериофагов» . Дж. Бактериол . 186 (23): 8153–5. дои : 10.1128/JB.186.23.8153-8155.2004 . ПМК 529087 . ПМИД 15547290 .
- ^ Ли К.Н., Линь Дж.В., Венг С.Ф., Ценг Ю.Х. (декабрь 2009 г.). «Геномная характеристика интронсодержащего Т7-подобного фага phiL7 Xanthomonas Campestris» . Прил. Окружающая среда. Микробиол . 75 (24): 7828–37. Бибкод : 2009ApEnM..75.7828L . дои : 10.1128/АЕМ.01214-09 . ПМК 2794104 . ПМИД 19854925 .
Дальнейшее чтение
[ редактировать ]- Чаухан, С; Калискан Г; Взяткодатель Р.М.; Перес-Салас У; Ранган П; Тирумалай Д; Вудсон С.А. (2005). «Третичные взаимодействия РНК опосредуют нативный коллапс рибозима бактериальной группы I». Дж Мол Биол . 353 (5): 1199–1209. дои : 10.1016/j.jmb.2005.09.015 . ПМИД 16214167 .
- Хауген, П; Саймон ДМ; Бхаттачарья Д. (2005). «Естественная история интронов группы I». Тенденции в генетике . 21 (2): 111–119. дои : 10.1016/j.tig.2004.12.007 . ПМИД 15661357 .
- Ранган, П; Маскида, Б; Вестхоф Е; Вудсон С.А. (2003). «Сборка основных спиралей и быстрое третичное сворачивание небольшого рибозима бактериальной группы I» . Proc Natl Acad Sci США . 100 (4): 1574–1579. Бибкод : 2003PNAS..100.1574R . дои : 10.1073/pnas.0337743100 . ПМК 149874 . ПМИД 12574513 .
- Шредер, Р; Барта А; Семрад К. (2004). «Стратегии сворачивания и сборки РНК». Nat Rev Mol Cell Biol . 5 (11): 908–919. дои : 10.1038/nrm1497 . ПМИД 15520810 . S2CID 22030359 .
- Тирумалай, Д; Ли Н; Вудсон С.А.; Климов Д (2001). «Ранние события сворачивания РНК». Annu Rev Phys Chem . 52 : 751–762. Бибкод : 2001ARPC...52..751T . doi : 10.1146/annurev.physchem.52.1.751 . ПМИД 11326079 .
- Трейбер, ДК; Уильямсон-младший (1999). «Выявление кинетических ловушек при сворачивании РНК». Curr Opin Struct Biol . 9 (3): 339–345. дои : 10.1016/S0959-440X(99)80045-1 . ПМИД 10361090 .
- Сяо, М; Лейбовиц М.Дж.; Чжан Ю (2003). «Согласованное сворачивание рибозима Candida в каталитически активную структуру после быстрого уплотнения РНК» . Нуклеиновые кислоты Рез . 31 (14): 3901–3908. дои : 10.1093/нар/gkg455 . ПМК 165970 . ПМИД 12853605 .