Бразилодон
| Бразилодон Временный диапазон: поздний триасы
~ | |
|---|---|
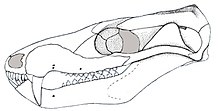
| |
| Реконструированный череп с боковым видом | |
| Научная классификация | |
| Домен: | Эукариота |
| Королевство: | Животное |
| Филум: | Chordata |
| Клада : | Synapsida |
| Клада : | Терапсида |
| Клада : | Cynodontia |
| Клада : | Маммалиаморфа |
| Семья: | † Brasilodontidae Bonaparte et al. , 2005 |
| Род: | † Бразилодон Bonaparte et al. , 2003 |
| Разновидность: | † B. Четырееху
|
| Биномиальное название | |
| † Brasilodon Quadrangularis Bonaparte et al. , 2003
| |
| Синонимы | |
| |
Brasilodon («зуб из Бразилии») является вымершим родом небольших, похожих на млекопитающих цинодонтов , которые жили в том, что сейчас является Бразилии в норья эпоху поздней триасовой эпохи, около 225,42 миллиона лет назад. Хотя полных скелетов не обнаружено, длина Бразилодона была оценена в 12 сантиметрах (4,7 дюйма). Его зубные зубные рты показывают, что это, скорее всего, насекомоядным . Род монотипический , содержащий только виды B. Quadrangularis . Бразилодон принадлежит к семейству Brasilodontidae , чьи члены были одними из самых близких родственников млекопитающих, единственными cynodonts, живыми сегодня. рода Brasilodontid, Brasilitherium и Minicynodon , теперь считаются младшими синонимами Бразилодона Два других .
Открытие и именование
[ редактировать ]
Первые три образца, упомянутые в Brasilodon Quadrangularis, были обнаружены на участке Linha Sáo Luiz, карьере возле города факсинал до Soturno в штате Рио -Гранд -ду -Сул . Скалы, где были найдены Бразилодон , принадлежат верхней части последовательности канделярийского супер -неэффективности Санта -Мария, соответствующей традиционной формированию Катурриты , которая была датирована ранним норским веком позднего триаса . [ 1 ] [ 2 ] [ 3 ] Голотип постканин (UFRGS-PV-0611-T) состоит из хорошо сохранившегося черепа, сохраняющего как левый, так и правый верхний , но отсутствует нижняя челюсть. Указанный образец UFRGS-PV-0716-T состоит из левой передней части черепа, сохраняя 6 постканинов. Образец UFRGS-PV-0628-T состоит из частичного черепа, включая нижнюю челюсть, причем большинство верхних и нижних зубных протезов сохранились. [ 1 ] С тех пор были обнаружены многие другие образцы Бразилодона , как в факсинал, так и в Канделарии , так и в содержании как черепного, так и посткраниального материала. [ 1 ] [ 2 ]
Род Brasilodon был назван в статье 2003 года Хосе Ф. Бонапарте , Агустином Дж. Мартинелли, Сезаром Л. Шульцем и Рожерио Рубертом. Общее название Брасилодон получено из страны Бразилии, где оно было найдено, и из греческого слова Одон , что означает «зуб». Предполагаемое значение - «зуб из Бразилии». Специфический эпитет Quadrangularis относится к прямоугольной форме верхних постканиновых зубов . [ 1 ]
В той же статье 2003 года был назван виды Brasilitherium riograndensis на основе шести образцов. Образцы, приписываемые Brasilithritherium были обнаружены в тех же местах, что и Brasilodon , и были отличены от этого таксона в значительной степени по их меньшим размерам, различным пропорциям черепа, , а не в Brasilodon Полем [ 1 ] [ 4 ] Бумага 2005 года построила семейство Brasilodontidae для двух родов. [ 4 ] В 2010 году был назван третий вид Brasilodontid, Minicynodon Maieri , был назван Bonaparte et al. , основываясь на одном хорошо сохранившемся черепе от факсинал до Soturno. Этот вид отличался от Бразилодона благодаря твердому прикреплению костей крыши черепа и от Бразилитерейума из -за отсутствия Cusp B в нижних постканинах. Minicynodon также был интерпретирован как обладание двойным челюстным суставом, в отличие от двух других видов. [ 5 ] Более поздние исследования поставили под сомнение обоснованность Бразилитерьера и Миницинодона , вместо этого предлагая, что они, наряду с Бразилодоном , просто представляют индивидуальные различия в одном виде. В этом случае Брасилодон является действительным таксоном, тогда как Бразилтерии и Миницинодон являются недействительными юношескими синонимами . [ 2 ] [ 6 ]
Описание
[ редактировать ]Brasilodon был небольшим, полученным Cynodont, с предполагаемой общей длиной около 12 сантиметров (4,7 дюйма). [ 7 ] На нем было много особенно похожих на млекопитающих, в том числе хорошо развитый вторичный небо , симметричное развитие зубов и более полученную анатомию ушей, чем в более ранних цинодонтах. [ 4 ]
Череп
[ редактировать ]

Различные образцы Бразилодона имели широко изменяющуюся длину черепа, в диапазоне от 20 до 55 миллиметров (от 0,79 до 2,17 дюймов), в основном из -за различий в возрасте. [ 7 ] Череп имел низкую удлиненную форму, когда виден со стороны. Верхняя часть мозгового мозга была довольно широкой, с коротким и низким темальным гребнем . [ 1 ] Бразилодону не хватало префронтальных и посторбитальных костей, которые были обнаружены у более базальных вероятногонаков, таких как Прозостродон . не было посторбитального бара За глазной розеткой . Зигоматическая арка (щечная кость) была довольно низкой и стройной. [ 2 ] [ 4 ]
Нижняя челюсть состояла в основном из зубной кости , которая носила зубы. Кончик нижней челюсти был согнут вверх. Симфиз были , сустав между двумя половинками зубного, был неисправен, с шероховатой поверхностью, где связки бы прикреплены . Меккельская канавка была расположена рядом с нижним краем зубного зубца. Постдентарные кости, набор костей в нижней челюсти, расположенной за зубной зоной, были сильно уменьшены по сравнению с состоянием в более примитивных цинодонтах. Как и в большинстве не млекопитающих цинодонтов, челюстный сустав в Бразилодоне включал в себя квадратную кость (кость черепа, гомологичный инков современных млекопитающих) и суставную кость (одна из постдентарных костей, гомологично по отношению к мальлеу ). [ 4 ] Возможно, также мог быть контакт между зубной и сквомольной костью , причем суставной процесс зубного зубца имел утолщенный конец, по -видимому, условие -предшественника для полностью развитого ххахтиля зубного мозга более производных млекопитающих . Квадратная кость имела хорошо развитый стапедиальный процесс, аналогичный базальному млекопитающему Morganucodon . [ 4 ]
Передняя часть крыши устья состояла из хорошо развитого вторичного неба , образованного челюстными и недобывающими костями , которые простирались примерно до последнего постканина. [ 6 ] У вторичного неба была канавка, в которой были бы вписывались нижние постканины, когда рот был закрыт. За вторичным вкусом был основной небо , образованное Вомером и Птеригоидными Костями . Была пара пробелов между птеригоидами, называемыми межптеригоидными ваваилами . Хорошо разработанные межптеригоидные валюты известны в базальных цинодонах, таких как Procynosuchus , но валюты, как правило, уменьшаются или отсутствуют в более продвинутых группах, поэтому их присутствие в Бразилодоне, вероятно, является производным условием. Птеригоиды имели поперечный контакт с базиптеригоидным процессом базисфеноидной кости . [ 4 ] Основнойран был широко , когда его видели снизу. Проотические . и опистотические кости были объединены в петросальную кость , которая имела четко промонториум определенную [ 2 ] Было отдельное отверстие (отверстия) для верхнечелюстных и нижнечелюстных нервов. Рядом с Crista Interfenestralis была большая депрессия, называемая «стапедиальным углублением», которая носила несколько отверстий. Затылочные мыщечки были расположены дальше, чем хрупкий гребень . [ 4 ]
Клюниальная эндокаст , внутренняя поверхность мозгового мозга, является важным источником информации о мозге доисторических животных, поскольку сами мозги очень редко сохраняются в окаменелостях. Исследование 2013 года, проведенное Rodrigues et al. описал эндокаст образца UFRGS-PV-1043-T, который первоначально был назначен роду Brasilitherium . Эндокаст имел длину 17,67 миллиметра (0,696 дюйма), что составляет 46,5% от общей длины этого черепа. Максимальная ширина эндокаста составила 7,17 миллиметра (0,282 дюйма). Форма эндокаста указывает на то, что животное обладает хорошо развитыми обонятельными луковицами , частями мозга, связанных с обонянием. С другой стороны, полушария головного мозга были меньше, чем у базальных млекопитающих, таких как Morganucodon . Неясно, ли Бразилодон обладал неокортекс , часть мозга, обнаруженная у современных млекопитающих. Коэффициент энцефализации (относительный размер мозга) был интерпретирован как, вероятно, выше, чем в более базальных цинодонах, таких как Traversodontid Massetognathus , но ниже, чем у базальных млекопитающих и современных млекопитающих. Как и другие не млекопитающие, у Бразилодона , вероятно, была лиссценсфальная (гладкая) поверхность мозга. [ 8 ]
Зубной зуб
[ редактировать ]
Как и большинство Cynodonts, Бразилодон обладал гетеродонтным зубным ртом, разделенным на резцы , клыки и постканины. Образцы, первоначально назначенные в Бразилодон, не сохраняют ни одного из верхних резцов, но некоторые образцы, приписываемые Brasilitherium , приходили 4 пары резцов в верхней челюсти. [ 9 ] Было 3 пары более низких резцов; Первый нижний резцерв был сведен (указывает вперед). [ 2 ] У взрослых образцов клыки были большими и сплющенными из стороны в сторону, но не имели зубцов . [ 4 ] У маленьких людей, приписываемых Brasilithritherium , было две пары функциональных собак, которые были лишь немного больше, чем резцы. [ 9 ]
В обеих челюстях было до 8 пар постканинов. [ 4 ] Постканины имели прямоугольную форму при взгляде на корону . В верхних постканинах было мало изменений в форме, в то время как нижние показали больше вариаций. [ 1 ] Верхние постканины были значительно меньше, чем нижние, условие, наблюдаемое во многих других вероятностях. [ 10 ] Постканины были «похожими на триконодонт», [ 2 ] с тремя основными пульсировками, расположенными примерно по прямой линии. На верхних зубах был большой центральный порог, названный половым путем в верхних зубах, а в нижних нижних в). [ 1 ] В нижних постканинах некоторых экземпляров был дополнительный порог (cusp d) за Cusp C; Отсутствие или присутствие этого Cusp использовалось для отличия Бразилодона от Бразилитерея , [ 1 ] Но вполне вероятно, что отсутствие Cusp D в некоторых образцах вызвано износом зубов у пожилых людей. [ 6 ] Постканины также имели различное количество аксессуаров, иногда формируя пояс . В более старых образцах между собаками и постканинами был большой разрыв ( диастема ), который был создан первым постканином, который пролиз, но не заменен. В средней нижней нижней части постканинов была языковая система для переплета между соседними зубами, при этом подготовительная подготовительная установка между Cusp B и аксессуарной почвой, но эта система, по-видимому, не была функциональной из-за расстояния между зубами. Корни постканинов не были полностью разделены, как они находятся на современных млекопитающих, но у них было заметное сужение в середине, что дало им 8-формированное поперечное сечение. [ 11 ] Питательные каналы корней тоже были разделены. [ 2 ] Постканиновые корни были закреплены в челюсти кольцом окостеневшей пародонтальной связки . [ 11 ]
Позвонки и ребра
[ редактировать ]
Позвонки Бразилодона известны . плохо Один экземпляр (UFRGS-PV-1043-T) сохраняет 4 изолированных предварительных позвонков, которые плохо сохранились. была Центра позвонка уплачена , имела плоский передний конец и вогнутый задний конец. Такая форма также наблюдается в некоторых тртилодонтидах и базальных формах млекопитающих, но не в более базальных цинодонах, которые обычно имели амфиковые позвонки, которые были вогнуты с обеих сторон. Нейронный канал был довольно большим, в то время как боковые стены были тонкими. Нейронные арки были слиты в центре в этом образце, показывая, что это был взрослый. Один из позвонков сохраняет небольшой и цилиндрический диапофиз (поперечный процесс). Зигапофизы (суставные процессы, четыре удлинения каждого позвонка, которые соединяют соседние позвонки вместе) , плохо сохраняются, но постзигапофии, по -видимому, были почти горизонтальными и расположены довольно высоко на позвонках. Форма основания нейронного позвоночника (заостренное разгибание на верхней поверхности позвонка) указывает на то, что он был удлиненным переднеропостериальным (в направлении спереди на спинку). [ 2 ]
Образец UFRGS-PV-1043-T также сохраняет 4 изолированных ребра фрагмента . Ребра были сплющены переднеропостериальным. не- Многие Эти тарелки отсутствуют у большинства вероятногонаков, включая Бразилодон . [ 2 ]
Конечности и опоры конечностей
[ редактировать ]Нижняя часть правой лопатки (лопаток) сохраняется в образце UFRGS-PV-1043-T, с двумя другими связанными фрагментами костей, вероятно, представляющими среднюю часть дорсального (верхнего) края и переднего (переднего) края. , соответственно. Лопатка представляла собой тонкую кость, с сутяным основанием, как и в других эпицинодонтах . Верхний край был несколько выпуклым, в то время как постеродорсальный (верхний задний) угол был указан. Передняя и задняя края имели фланцы, продюсирующие боком, как видно в других эпицинододододонтах, не являющихся млекопитающими. Был хорошо развитый процесс акромиона в конце переднего края. Акромион показал вперед, как во многих других цинодонтах. была глубокая инфраспонизионная ямка На бортовой стороне лопатки , но надпрессивная ямка, казалось бы, отсутствовала. Гленоидный плечевой фасфель лопатки (область, где она соединялась с костью ) была вогнутой и имела несколько яйцевидную форму. Гленоид, по-видимому, был указан вниз, в отличие от более обращенных боковых гленоидов некоторых ранних цинодонтов. А Коракоидные и прокоракоидные кости не сохранились, но на лопаточном лопаточнике были суставные поверхности для этих костей. [ 2 ]

Образец UFRGS-PV-1043-T сохраняет полную левую плечевую кость (кость верхней части руки) с длиной 15,6 миллиметра (0,61 дюйма). Вал плечевой кости был стройным. Он был несколько скручен, с угловой 15 градусов между противоположными концами кости. Скрученная плечевая кость встречается в большинстве цинодонтов за пределами Терии (клада, содержащая современные сумчатые и плаценты ), в том числе в современных монотринах . Тем не менее, скручивание плечевой кости было относительно низким в Бразилодоне по сравнению со многими другими нетерианскими цинодонтами. На проксимальной стороне (стороне, соединяющейся с плечом), голова плеча имела полусферическую форму, типичную для форм -млекопитающих, но в отличие от более базальных цинодонтов, где головка плеча, как правило, была более яйцевидной. В отличие от большинства Therians, голова плечевой кости столкнулась с постератологическим (назад и в стороне). был гребень, простирающийся от голова плечевой кости до герба На другом конце кости . В отличие от более базальных cynodonts, у Бразилодона был четкий Большой бугорок , хотя он был довольно маленьким. У меньшего бугорка была толстая и выпуклая форма. Это было больше, чем больший бугорок, как у базальных млекопитающих и монотрим, но в отличие от мультитуберкулятов , спалакотеридов и терианов. Дельтопекторный гребень был сливался с большим бугорком и распространялся на 48% длины плечевой кости. Гребень был довольно большим и надежным, как и в других не млекопитающих цинодонтах, монотриах и исчезновениях (ограбления), таких как родинки . На другой стороне кости был еще один гребень, известный как Theres Crest . Этот гребень был найден в большинстве мезозойских цинодонтов, но пропал без вести в Винселесте и Теранах. Гребень Терес был примерно столько же, сколько дельтопекторный гребень, и простирался от меньшего бугорника до большого и яйцевидного выступления вблизи середины кости. Эта выступа, возможно, послужила якорем для основной мышцы Терес . Аналогичная структура существовала во многих других мезозойских цинодонтах, но не в многотуркуляциях, спалакотеридах или терицах. На передней поверхности плечевой кости был широкий Биципитальная канавка , граничащая с дельтопекторными и гребнями -терминами, аналогично многим другим мезозойским цинодонтам. Биципитальная канавка, вероятно, служила точкой прикрепления для мышц Coracobrachialis . Дистальная сторона плечевой кости (сторона, соединенная с ) имела ширину 43 радиусом и лонешкой предплечья % длины плечевой кости. Это было уже, чем во многих других не млекопитающих цинодонтах, а также в монотримах, но шире, чем у базальной млекопитающей Морганакодон и Терана Дидельфиса . Entepicondyle был довольно надежным и имел выступа , которое указывало на проксимальный конце кости. Эктопикондиль был более узким и менее выступающим , чем энтепикондиль. Entepicondylar Foramen представляла собой большую и удлиненную дыру. И наоборот, не было никакого эктипикондиларного отверстия ; Эта дыра была найдена в большинстве цинодонтов более базальных, чем Брасилодон , но отсутствует в TrityLodontids и большинстве млекопитающих. Ульнары мыщелки (части , и радиальные соединяющие ясенную и радиус, имели выпуклую форму, как видно в других не млекопитающих цинодонтах. Мыщелки были разделены узкой канавкой и не имели Трохлеарная (похожая на шкив) форма, наблюдаемая во многих млекопитающих короны, таких как Therians или стержни-монотримные Kryoryctes . [ 12 ] Ульнарный мыщелок был виден как спереди, так и сзади плечевой кости. Его передняя сторона была несколько поперечной сжатой. Радиальный мыщелок был больше, чем локтевой мыщелок, но не был виден на задней стороне кости. Фосса олеклона (депрессия на задней части плечевой кости) была довольно мелкой, в отличие от более глубокой ямки, наблюдаемой в многотуркулятах, винселесо и терицах. [ 2 ]

Левый радиус UFRGS-PV-1043-T составляет 14 миллиметров (0,55 дюйма). Радиус был стройным, с круглым поперечным сечением возле проксимального конца (ближе к локте). Голова радиуса (где он сочленен с плечевой костью) была приблизительно круговой, с чашкой депрессией посередине окружена выпуклым ободом. Голова была несколько наклонена к боковой стороне передне -передней (спереди). На задней (обратной) стороне головы была небольшая суставная граница, где он был бы сформулирован с радиальной выемкой локтевой кости. Не было радиальной бугристости для прикрепления мышц бицепса . Дистальный конец кости (ближе к запястью) был формирован как сплющенный прямоугольник. Дистальный конец был несколько изогнутым постеромерным, что позволило бы кости пересечь переднюю переднюю часть локтевой кости, как у TrityLodontids, Morganucodon и Opossums. Дистальная суставная поверхность (где она соединялась с запястью) также имела прямоугольную форму. [ 2 ]
Правая локтевая кость UFRGS-PV-1043-T имеет длину 16,5 миллиметра (0,65 дюйма). Как и в большинстве цинодонтов, локтевая кость была узкой средней (от стороны в сторону) и имела сигмоидную (S-подобную) форму. По бокам кости было два бороздка, проходящих от одного конца кости до другого, заключенных в утолщенные края на передней и задней стороне. Считается, что канавки послужили точками привязанности для мышц разгибателя и сгибателей. На проксимальном конце кости произошел хорошо развитый и окостененный процесс олекранона . За исключением Trucidocynodon и TrityLodontids, ни у Cynodonts более базальных, чем Brasilodon, он был окостененный олеклин, и, как полагают, он был хрящевым известно, что в этих таксонах . Олеверон Бразилодона составил чуть менее 20% от длины локтевой кости и был направлен вперед по сравнению с остальной частью кости. Олекранон был несколько толще из стороны в сторону, чем остальная часть кости. Полунарная выемка , где яле сформулирована с локтевым мыщелком плечевой кости, была большой и полукруглой. А Процесс AnCemeal был довольно маленьким, как в более базальных цинодонтах и Docodont Mammaliform Haldanodon , но в отличие от многих других форм для млекопитающих, где процесс AnCeleal образует выдающийся гребень. [ 2 ]

Таз (тазобедренная кость) UFRGS-PV-1043 - T сохраняет полную левую вертлушку и лоток , а также неполный левый подвздошной тренас и иший . Вт в вешалку - это то, где бедренная кость задних конечностей артикулирует таз. Вт в Бразилодон был глубокой, сферической и обращенной боковой депрессией. Три кости, составляющие вертлурку (лоток, подвздошной тренажерный и иший), были слиты вместе, и не швов было видно ; Это особенность, которую Бразилодон поделился с MammaliaForms, в то время как кости были более различными в некоторых более базальных цинодонах, таких как Traversodontids . Кости сформировали три различных гребня, известных как надписные режимы вокруг вертлужной впадины. Подотры были разделены зазорами в 2–3 миллиметрах (0,079–0,118 дюйма), в отличие от современных терицев, где вокруг вертлужной впадины есть полностью очерчиваемый обод. Пробелы, скорее всего, были бы заполнены фибро -каркасовым веществом у живого животного. Лезвие подвздошного костюма сильно сохранилось, но форма основания указывает на то, что почтовая (обратная точка) часть лезвия была либо уменьшена, либо отсутствует в Бразилодон , как и в других прозостродонте . Лобковая бугриость была расположена под вертлужной впадиной, как у большинства вероятногонаков, в том числе современные терицы. В базальных эпицинододонтах монотримы и спалакотеридные акидолотесты лобковая бугристость была расположена перед вертлужкой. Лобический лоб и иший сформировал большое и яйцевидное отюраторное отверстие . [ 2 ]

Правая бедра (бедра) UFRGS-PV-1043-T составляет 15,9 миллиметра (0,63 дюйма). Вал бедренной кости был в основном прямым, но с выдающимся наклоном вперед, близким к тазобедренному суставу, как в других не млекопитающих цинодонтах; У современных млекопитающих этот изгиб менее хорошо развит. Проксимальная часть вала (ближе к бедру) была в основном квадратной формы, но она стала более сжатой спереди к спине более дистально (к колену), одновременно становилась все шире из стороны в сторону. Проксимальные и дистальные концы кости имели одинаковую ширину. На проксимальном конце кости голова бедренной кости (которая сочленена с вертлужной впадиной тазобедренного сустава) имела полусферическую форму. Голова была отделена от остальной части кости короткой шейкой бедренной кости , как у Tritylodontids и Mammaliaforms; Более примитивным цинодонтам не хватало бедренной шеи. Голова была наклонена около 60 градусов медиально (справа) по сравнению с продольной осью бедренной кости, напоминающей состояние в многотуркуляциях и спалакотеридах. В отличие от MammaliaForms, не было Фовеа Капит на головке бедренной кости. Немного дистально от головы бедра, была хорошо развитая проекция, известная как Большой Трохантер на левой стороне кости. Отчетливый больший вертел также обнаружен в TrityLodontids и MammaliaForms, но больший трохантер был слияние с головой бедренной кости в более базальных цинодонтах. На правой стороне кости был короткий гребень под названием The Messer Trochanter . Меньший Трохантер указал направо и был видим на передней стороне кости, как у многих других вероятностей. В более базальных цинодонтах это, как правило, было более непосредственным и не видно на передней стороне. Меньший вертел не распространялся на проксимальный конце кости, как у базальных млекопитающих. Там не было третьего вертела . На задней стороне бедренной кости, между большими и меньшими верточками, была глубокая и узкая депрессия, называемая межтрохантерической ямкой . На дистальном краю этой ямки был межтрохантерический гребень которые соединяли два вертола. Этот гребень также обнаружен в большинстве терицев, но пропал без вести в большинстве мезозойских групп Cynodont. Была относительно мелкая треугольная депрессия, расположенная дистально от межтрохантерического гребня; Это, вероятно, послужило точкой привязанности для мышц аддуктора бедра. На задней стороне дистального конца бедренной кости было было два мыщелка, которые были бы связаны с голенью , медиальными и боковыми мыщелками. Медиальный мыщелок был сжат из стороны в сторону. Покольная ямка имела треугольную форму и была расположена рядом с мыщелками. [ 2 ]

UFRGS-PV-1043-T сохраняет почти полную левую голень (голени). Это была стройная кость, с консервированными частями длиной 16,1 миллиметра (0,63 дюйма). Вал был в основном прямой, но с небольшой медиальной (справа) кривой в проксимальной части (к колену); Более базальные цинодонты, как правило, имели более сильную медиальную кривую голени. Вал постепенно становился все более сплющенным к дистальному концу (к лодыжке). В проксимальной части было две суставные грани, медиальные и боковые грани, которые сформулировались с мыщелками бедренной кости. Боковой аспект был шире, чем медиальный. Дистальный конец голени не сохранился. [ 2 ]
UFRGS-PV-1043-T сохраняет два левых костях ласки , кальцины и астрагалу . Кальцинум (кость пятки) Бразилодона имел низкую и широкую форму, в отличие от узкого кальцины, обнаруженного в многотуркулятах, « симметродонте » Чжангеотерия и Терана. У него была выпуклая верхняя поверхность и вогнутая нижняя поверхность. был хорошо развитый клубень кальцис На заднем конце кости ; Эта особенность встречается в большинстве эуцинодонтов, но отсутствует в более базальных таксонах, таких как Тринаксодон . был перонеальный шельф На боковой (левой) стороне кальцины . С медиальной (правой) стороны была проекция, известная как Sustentaculum tali , который был бы связан с нижней стороной астрагала (кость нагнетателя). В базальных цинодонтах, таких как Тринаксодон , не было обнаружено никаких сунтикулум тали, что позволяет предположить, что он был хрящевым, если он даже существовал в этих таксонах. Левый астрагал был меньше, чем кальцеум. У него была полусферическая дорсомедиальная (верхняя правая) поверхность и плоская латеоплантарная (нижняя левая) поверхность. На переднем конце астрагала была маленькая голова. Шея, которая отделяла голову от остальной части кости, была короче, чем у современных терицев. [ 2 ]
UFRGS-PV-1043-T также сохраняет два неполных плюсневых костюмах , включающих проксимальный и дистальный конец, который был бы связан с тарзалами и с фалангами , соответственно. Проксимальный конец имел круговую депрессию, в то время как дистальный конец имел два симметричных мыщелка. Возможный фрагмент метапода (плюсневой или метакарпал ) также известен из образца UFRGS-PV-0765-T, который имел широкую проксимальную часть и более узкий вал. Оба этих образца также сохраняют некоторые фаланги (кости пальцев), а UFRGS-PV-0765-T сохраняет почти полный средний фаланги. Этот фаланги был довольно коротким, с вогнутым проксимальным концом и двумя маленькими мыщелками на дистальном конце. [ 2 ]
Классификация
[ редактировать ]Род Brasilodon принадлежит Brasilodontidae, семейству усовершенствованных вероятногогнатиков. Наряду с Бразилодоном два современных рода ( Brasilitherium и Minicynodon ) были отнесены к семье, оба из которых могут быть синонимами Бразилодона . [ 2 ] [ 6 ] Protheriodon , род из среднего триаса ( Ladinian ) из Бразилии, и Panchetocynodon , плохо известный Cynodont из раннего триаса ( Induan ) Индии, также были помещены в Brasilodontidae в статье 2013 года Хосе Бонапарта. [ 13 ] Однако в последнее время было обнаружено, что Protheriodon был ранним, вероятно, не связанным с Brasilodontids, [ 14 ] В то время как панчетоцинодон в противном случае рассматривается как таксон Incertae Sedis , чьи отношения с другими таксонами неясны из -за его неполноты. [ 9 ] [ 15 ]
Особенности черепа и зубного проведения Бразилодона указывают на то, что это был полученный цинодонт, тесно связанный с млекопитающими, единственными существующими цинодонтами. Филогенетический анализ обычно восстанавливал его в качестве члена Clade Mammaliamorpha, лежащего ближе к млекопитающим, чем TrityLodontids, но вне млекопитающих, более эксклюзивные клады, содержащие млекопитающие и их ближайшие родственники, такие как Morganucodonts , Docodonts и Haramyyidans . Кладограмма et ниже адаптирована из анализа 2019 года Wallace al. : [ 16 ]


| Progainognathia |
| ||||||||||||||||||||||||||||||||||||||||||||||||
Палеобиология
[ редактировать ]
Локомоция
[ редактировать ]Особенности посткраниума указывают на то, что Бразилодон был обобщенным животным, способным к разнообразным способам локомоции , включая копание и скалолазание. Большая бугристость вблизи дельтопекторного гребня плечевой кости подразумевает, что основная мышца Тереров (важная мышца ретрактора передней части переднего движения) была довольно большой, как видно из современных ископаемых грызунов . Однако адаптация копания Брасилодона была менее выраженной, чем у современных ископаемых млекопитающих, а также многие другие не млекопитающие цинодонты. Свидетельство о способностях сканирующего (скалолазания) включает в себя полусферические головки плечевой кости и бедренного бедра, которые позволили бы широкий спектр вращения плечевых и бедных суставов, хорошо развитого эктопикондилар присваивали высокую подвижность локте и небольшой анцехеально -процесс локтевой кости. Несколько особенностей плечевой кости, в том числе скрученный вал, указывают на то, что у Брасилодона были растянутые или полупрогровочные передние мароды. С другой стороны, особенности таза и задних конечностей, такие как плохо развитая почтовая часть лезвия подвздошной атмосферы, указывают на более прямое (более прямое (более прямое (более прямое (более прямое ( парасагиттальная ) осанка задних конечностей. [ 2 ]
Замена зуба
[ редактировать ]Согласно одной теории, подробно описано в статье 2010 года Martinelli et al. , Брасилодон имел замену зуба полифиодонта , где зубы были заменены более одного раза на протяжении всей жизни животного, хотя неясно, сколько последовательных замен в постканине было в Бразилодоне . [ 11 ] Замена постканина прекратилась задолго до смерти человека, что позволило бы развиваться в зубах чрезвычайно сильного износа. В некоторых случаях зубы человека со временем носили менее половины высоты короны, оставшейся. [ 4 ] Размер почтовой диастемы указывает на то, что передние постканины могли быть потеряны быстрее, чем в примитивном цинодонте Тринаксодон . [ 11 ] [ 10 ]
Согласно Martinelli et al. (2010), постканины были заменены чередующимися способами. Замена постканина произошла бы с задней части на фронт, как и у некоторых других неприятных вероятностных вероятностей, и, таким образом, отличалась бы от замены ранних млекопитающих, где постканины были заменены спереди к задней части. Возможно, это было связано с неограниченным ростом черепа ранних вероятностных. [ 11 ] Изменение замены зубов, возможно, также может быть связано с диетой, с травоядными/всеядными травернодонтидами, имеющими расширенные постканины и последовательную замену зубов, [ 17 ] и плотоядные и насекомоядные, не млекопитающие, вероятно, такие как Бразилодон, имеющие чередующуюся замену зубов. [ 11 ]
Исследование 2022 года, проведенное Cabreira et al. предполагает, что Бразилодон на самом деле был Дипхёдонтом , заменяя его зубы только один раз, как и большинство современных млекопитающих. Согласно этой теории, постканины в Бразилодоне состояли из набора лиственных зубов , которые были заменены, и набора постоянных зубов, разделенных на премоляры и моляры . Исследование предполагает, что лиственные постканины в Бразилодоне последовательно разразились в направлении обратного к фронта, в то время как моляры были добавлены в противоположном направлении, спереди к задней части. [ 7 ]
Палеоэкология
[ редактировать ]
Образцы Бразилодона были найдены в двух местах. Голотип был обнаружен в местности Линха Сан -Луиз, в муниципалитете факсинал Доурно. Другие образцы были обнаружены в местности Sesmaria do Pinhal в муниципалитете Канделарии; Оба эти места находятся в бразильской части бассейна Парана . [ 18 ] Скалы, где были найдены Бразилодон , принадлежат верхней части последовательности Канделярии, которая соответствует биостратиграфической единице, известной как риограндий зона сборки . Riograndia , AZ была датирована ранним эпохом норья покойной триасовой эпохи [ 3 ] Около 225,42 миллиона лет назад. [ 7 ] Cynodonts являются одним из наиболее распространенных и таксономически разнообразных элементов в этой зоне сборки, [ 5 ] и представлены многими хорошо сохранившимися образцами. [ 19 ] В дополнение к Бразилодону , Cynodonts представлены Trithellontids Irajatherium и Riograndia , базальным млекопитающим ботукарайтером и неопределенными травернодонтидами . Другие животные включают Dicynodont Jachaleria , Procolophonid Soturnia , Lepidosauromorphs Cargninia , Clevosaurus и Lanceirosphenodon и Avemetatarsalians Faxinalipterus , Guaibasaurus , Macrocollum , Maehary , [ 3 ] Sacisaurus и Unaysaurus . Неопределенные остатки темносподинс , фитозавров и других групп также были обнаружены. [ 18 ]
Место, где были обнаружены эти окаменелости, представляет собой речную систему, характеризующуюся большим количеством мелких песчаников, образующих песчаные пласты, возникающие в результате седиментации в бассейне во время пиковых событий потока. [ 1 ] [ 19 ]
Ссылки
[ редактировать ]- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж Бонапарт, JF; Martinelli, Ag; Schultz, cl; Руберт Р. (2003). А Бразильский журнал о палеонтологии 5 : 5–27.
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л м не а п Q. ведущий с Т в Гиньярд, ML; Martinelli, Ag; Soares, MB (2019). «Посткраниальная анатомия Brasilodon Quadrangularis и приобретение признаков Mammaliaform среди некоммерческих цинодонтов» . Plos один . 14 (5): E0216672. doi : 10.1371/journal.pone.0216672 . PMC 6510408 .
- ^ Jump up to: а беременный в Келлнер, Ава; Holgado, B.; Грилло, О.; Предположим, FA; Kerber, L.; Pinheiro, FL; Soares, MB; Schultz, cl; Лопес, RT; Araújo, O.; Мюллер, RT (2022). «Переоценка Faxinalipterus Minimus , предполагаемый триасовый птерозавр из южной Бразилии с описанием нового таксона» . ПЕРЕЙ . 10 : E13276. doi : 10.7717/peerj.13276 . PMC 9074864 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k Бонапарт, JF; Martinelli, Ag; Schultz, CL (2005). «Новая информация о Brasilodon и Brasilitherium (Cynodontia, Probainognathia) из позднего триаса южной Бразилии» . Revista Brasileira de Paleontologia . 8 (1): 25–46.
- ^ Jump up to: а беременный Bonaparte, J.; Schultz, C.; Soares, M.; Мартинелли, А. (2010). «Факсина местной фауны Сотрурно, позднее триаса из Рио -Гранде -ду -Сул, Бразилия» . Бразильский журнал палеонтологии . 13 (3): 233–246. Doi : 10.4072/rbp.2010.3.07 .
- ^ Jump up to: а беременный в дюймовый Лю, Дж.; Олсен, П. (2010). «Филогенетические отношения эуцинодонтии (Amniota: Synapsida)» . Журнал эволюции млекопитающих . 17 (3): 151–176. doi : 10.1007/s10914-010-9136-8 .
- ^ Jump up to: а беременный в дюймовый Cabreira, SF; Schultz, cl; Da Silva, LR; Лора, LHP; Pakulski, C.; Do Rêgo, RCB; Soares, MB; Смит, М.М.; Рихтер М. (2022). «Замена зуба дифёдонта Бразилодона - поздний триасовый эукинодонт, который бросает вызов времени происхождения млекопитающих». Журнал анатомии : JOA.13756. doi : 10.1111/joa.13756 .
- ^ Rodrigues, pg; Руф, я.; Schultz, CL (2014). «Изучение цифровой краниальной эндокаст не млекопитающей Cynodont brasilitherium riograndensis (более поздний триасы, Бразилия) и его отношение к эволюции мозга млекопитающих». Paläontologische Zeitschrift . 88 (3): 329–352. doi : 10.1007/s12542-013-0200-6 .
- ^ Jump up to: а беременный в Martinelli, AG (2017). «Вклад в знание вероятностных проделок (Therapside, Cynodontics, Probainognathia) Южной Америки и его влияние на происхождение форм -млекопитающих» (на португальском языке).
- ^ Jump up to: а беременный Soares, MB; Martinelli, Ag; De Oliveira, TV (2014). «Новый прозостродонтский Cynodont (Therapsida) из поздней ферменной зоны риограндий (Santa Maria Supersecence) на юге Бразилии» . Материалы Бразильской академии наук . 86 (4): 1673–1691. Doi : 10.1590/0001-3765201420140455 . HDL : 10183/119079 .
- ^ Jump up to: а беременный в дюймовый и фон Martinelli, Ag; Bonaparte, JF (2010). «Замена постканина в Бразилодоне и Бразилитерье (Cynodontia, Probainognhatia) и ее подшипник в эволюции цинотодирования» . Paleontología y Dinosaurios desde América Latina : 179–186.
- ^ Придмор, Пенсильвания; Рич, Т.Г.; Викерс-Рич, П.; Gambaryan, PP (2005). «Тахилоссид, похожий на плечевой кости с раннего мела юго-восточной Австралии». Журнал эволюции млекопитающих . 12 (3–4): 359–378. doi : 10.1007/s10914-005-6959-9 .
- ^ Bonaparte, JF (2013). «Эволюция Brasilodontidae (Cynodontia-Eucynodontia)». Историческая биология . 25 (5–6): 643–653. doi : 10.1080/08912963.2012.731154 .
- ^ Martinelli, Ag; Eltink, E.; Da-Rosa, á. КАК; Лангер, MC (2017). «Новый Cynodont из формации Санта -Мария, Южная Бразилия, улучшает поздние триасовые вероятность разнообразия». Документы по палеонтологии . 3 (3): 401–423. doi : 10.1002/spp2.1081 .
- ^ Das, DP; Гупта, А. (2012). «Новая запись Cynodont из нижней триасовой формы Панкета, Долина Дамодар». Журнал Геологического общества Индии . 79 (2): 175–180. doi : 10.1007/s12594-012-0022-2 .
- ^ Уоллес, RVS; Martínez, R.; Роу, Т. (2019). «Первая запись базального млекопитающего из раннего позднего триасового образования ишигуаласто в Аргентине» . Plos один . 14 (8): E0218791. doi : 10.1371/journal.pone.0218791 . PMC 6685608 .
- ^ Martinelli, Ag; Soares, MB; Schwanke, C. (2016). «Два новых cynodonts (Therapsida) из среднего раннего позднего триаса Бразилии и комментируют южноамериканские вероятного препарата» . Plos один . 11 (10): E0162945. doi : 10.1371/journal.pone.0162945 . PMC 5051967 .
- ^ Jump up to: а беременный Martinelli, Ag; Эскобар, JA; Francischi, H.; Cerber, L.; Мюллер, RT; Руберт, Р.; Schultz, cl; Da- Rose, á .. как (2021). «Новая запись о STahleckeriid Dicynodont (Therapsida, Dicynodontia) из позднего триаса южной Бразилии и биостратиграфических замечаний по зоне сборки риограндий». болология Старинная 33 (11): 3101–3110. doi : 10.1080/ 0
- ^ Jump up to: а беременный Де Оливейра, телевидение; Martinelli, Ag; Soares, MB (2011). «Новая информация об Иражамии Эрнандези Мартинелли, Бонапарте, Шульц и Руберт 2005 (Euynodontia, Tritheledontidae) из верхнего триаса (образование Caturrita, бассейн Парана) Бразилии». Paläontologische Zeitschrift . 85 (1): 67–82. Doi : 10.1007/s12542-010-0078-5 .





