Птанодон
| Птанодон | |
|---|---|

| |
| Сделанная копия взрослого мужчины P. longiceps , Американский музей естественной истории | |
| Научная классификация | |
| Домен: | Эукариота |
| Королевство: | Животное |
| Филум: | Chordata |
| Заказ: | † Птерозаврия |
| Подотряд: | † pterodactyloidea |
| Семья: | † pteranodontidae |
| Подсемейство: | † pteranodontinae |
| Род: | † Pteranodon Болото , 1876 |
| Тип видов | |
| † Pteranodon longiceps Болото, 1876
| |
| Другие виды | |
| |
| Синонимы | |
| |
Pteranodon ( / t ə ˈ r æ n ə d ɒ n / ; от древнегреческого : πτερόν , романизированный : pteron ' wing ' и ἀνόΔων , анодон ' беззубые ' ) [ 2 ] [ Лучший источник необходим ] является родом птерозавра , который включал одни из крупнейших известных летающих рептилий , причем P. longiceps имел размах крыльев более 6 м (20 футов). Они жили в поздний меловой геологический период Северной Америки в современном Канзасе , Небраске , Вайоминге , Южной Дакоте и Алабаме . [ 3 ] больше ископаемых образцов птеродона Было обнаружено , чем любой другой птерозавр, с около 1200 образцами, известными для науки, многие из которых хорошо сохранились с почти полными черепами и сочлененными скелетами. Это была важная часть сообщества животных в западном внутреннем морском пути . [ 4 ]
Pteranodon - самый известный птерозавр, часто представляемый в СМИ динозавров и тесно связанный с динозаврами широкой публикой. [ 5 ] В то время как не динозавры, птерозавры, такие как птеродон, образуют кладу , тесно связанную с динозаврами, поскольку оба попадают в кладку Avemetatarsalia .
Открытие и история
[ редактировать ]
Первые окаменелости
[ редактировать ]Птеранодон был первым птерозавром, найденным за пределами Европы . Его окаменелости были сначала обнаружены Отниэлем Чарльзом Маршем в 1871 году, [ 6 ] В позднем меловом дымном холме залежи мела Западного Канзаса. Эти меловые кровати были отложены на дне того, что когда -то было западным внутренним морем , большое мелкое море над тем, что сейчас является серединой североамериканского континента. Эти первые образцы, YPM 1160 и YPM 1161, состояли из частичных костей крыла, а также зуба из доисторической рыбы Xiphactinus , который, по ошибке, по ошибке принадлежал этому новому птерозавра (все известные птерозавры до этого момента). В 1871 году Марш назвал находку Pterodactylus oweni , присваивая его хорошо известному (но гораздо меньшим) европейскому роду Pterodactylus . название, которое он выбрал, уже использовалось для европейских видов птерозавров Гарри Сили . Понимая, что Марш также собрал больше костей крыла большого птерозавра в 1871 году . , »в его описании нового образца в 1872 году. Он назвал два дополнительных вида, основываясь на различиях в размере: Pterodactylus Ingens (самый большой образец до сих пор) и Pterodactylus Velox (самый маленький). [ 7 ]
Тем временем соперник Марша Эдвард Дрибер Коуп обнаружил несколько образцов крупного североамериканского птерозавра. Основываясь на этих образцах, Коуп назвал двух новых видов, Ornithochirus umbrosus и Ornithochirus harpyia , пытаясь назначить их крупному европейскому роду Ornithocheirus , хотя он написал имя (забыв «E»). [ 7 ] Название бумаги Коупа его вида было опубликовано в 1872 году, всего через пять дней после бумаги Марша. Это привело к спору, сражающемуся в опубликованной литературе, над чьи имена были приоритетными в том, что, очевидно, было одним и тем же видом. [ 7 ] В 1875 году Коуп признал, что имена Марша имели приоритет над своим, но утверждал, что Pterodactylus Umbrosus был отдельным видом (но не родом) от любого, как Марш назвал ранее. [ 8 ] Повторная оценка более поздними учеными поддержала дело Марша, опровергая утверждение Копа о том, что П. Умбрус представлял собой более крупные, отдельные виды. [ 7 ]
Беззубокий птерозавр
[ редактировать ]
В то время как первые кости крыла Pteranodon были собраны Marsh and Cope в начале 1870 -х годов, 2 мая 1876 года был найден первый череп Птерододона , вдоль реки Смоки -Хилл в округе Уоллес (сейчас округ Логан), Канзас, США, Самуэль Венделл Уиллистон , коллекционер ископаемых, работающий на болота. [ 4 ] Второй, меньший череп, скоро был также обнаружен. Эти черепа показали, что североамериканские птерозавры отличались от каких -либо европейских видов, потому что им не хватало зубов и имели костные гребни на своих черепах. Марш признал это основное различие, описывая образцы как «отличившись от всех ранее известных родов ордена Птерозаврии по всему отсутствию зубов». Марш признал, что эта характеристика гарантировала новый род, и в 1876 году он придумал название Pteranodon («крыло без зуба»). Марш реклассифицировал все ранее названные североамериканские виды от Pterodactylus до Pteranodon . Он считал, что меньший череп принадлежит Pteranodon occidentalis , основываясь на его размере. Marsh классифицировал более крупный череп, YPM 1117, в новых видах Pteranodon Longiceps , который, по его мнению, является видом среднего размера между небольшими P. occidentalis и большими P. Ining . [ 9 ] [ 7 ] Марш также назвал несколько дополнительных видов: Pteranodon Comptus и Pteranodon Nanus были названы в честь фрагментарных скелетов мелких людей, в то время как Pteranodon Gracilis был основан на кости крыла, которую он принял за тазовую кость. Вскоре он осознал свою ошибку и снова классифицировал этот образец в отдельный род, который он назвал Nyctosaurus . П. Нанус был также позже признан образцом из Нью -Йорка . [ 10 ] [ 7 ]
В 1892 году Сэмюэль Уиллистон изучил вопрос о классификации Pteranodon . Он заметил, что в 1871 году Сили упомянул о существовании частичного набора беззубых челюстей из птерозавров из Кембриджского зелени и Англии , которую он назвал Орнитостомой . Поскольку основным характеристическим болотом использовался для отделения птеронодона от других птерозавров, было отсутствие зубов, Уиллистон пришел к выводу, что «орнитостома» следует считать старшим синонимом птеродона . Однако в 1901 году Плейнингер указал, что «орнитостома» никогда не была с научной точки зрения или даже назначал видовое название до работы Уиллистона, и поэтому был номенем и не мог победить Птеродона для названия приоритета. Уиллистон принял этот вывод и вернулся к названию рода Pteranodon . [ 7 ] Тем не менее, и Уиллистон, и Плейнингер были неверны, потому что незамеченными им обоим был тот факт, что в 1891 году сам Сили наконец -то описал и должным образом назван Ornithostoma , присвоив его виду О. Седжвицки . В 2010 -х годах больше исследований идентичности Ornithostoma показали, что это, вероятно, не Pteranodon или даже близкий родственник, но на самом деле, возможно, был азархоидом , другим типом беззубого птерозавра. [ 11 ]
Пересмотр видов
[ редактировать ]Уиллистон также был первым ученым, который критически оценил все виды Pteranodon, классифицированные Cope и Marsh. Он согласился с большей частью классификации Марша, за некоторыми исключениями. Во -первых, он не верил, что P. Ingens и P. Umbrosus можно считать синонимами, которые даже COPE пришли к выводу. Он считал, что П. Велокс и П. Лонгийспс сомнительными; Первый был основан на не диагностических фрагментах, а второй, хотя и известный из полного черепа, вероятно, принадлежал одному из других, ранее названных видов. В 1903 году Уиллистон пересмотрел вопрос о классификации Pteranodon и пересмотрел свой более ранний вывод о том, что было семь видов до трех. Он считал, что как P. comptus, так и P. nanus являются образцами Nyctosaurus , и разделили другие на малые ( P. Velox ), среду ( P. occidentalis ) и крупные виды ( P. Inning ), основанные главным образом на форме их плечо кости. Он думал, что П. Лонгиспс , единственный, известный из черепа, может быть синонимом любого P. Velox или P. occidentalis , на основе его размера. [ 7 ]
В 1910 году Итон стал первым ученым, который опубликовал более подробное описание всего скелета Pteranodon , как было известно в то время. Он использовал свои выводы, чтобы снова пересмотреть классификацию рода, основываясь на лучшем понимании различий в анатомии Pteranodont. Итон провел эксперименты с использованием глинистых моделей костей, чтобы помочь определить влияние дробления и сглаживания на формы костей руки, которые Уиллистон использовал в своей собственной классификации. Итон обнаружил, что большинство различий в формах костей можно легко объяснить давлением окаменелости, и пришло к выводу, что ни один скелеты птеронодона не имели существенных отличий друг от друга, помимо их размера. Поэтому Итону оставалось решать свою схему классификации, основанную на различиях только в черепах, которые он назначил видам так же, как и Марш, по их размеру. В конце концов, Итон узнал только три достоверных вида: P. occidentalis , P. Inningens и P. longiceps . [ 7 ]
Открытие образцов с вертикальными гребнями, классифицированные Харксеном в 1966 году как новый вид птерододонского Стернберги , еще больше усложнило ситуацию. побуждение еще одного пересмотра рода Миллера в 1972 году. Поскольку невозможно было определить форму гребня для всех видов на основе безголовных скелетов, Миллер пришел к выводу, что все виды птерододона, за исключением двух, основанных на черепах ( P. longiceps и P. Sternbergi ) Должны считаться Номиной Дубией и заброшены. Череп Итон, который, подумал, принадлежал P. Inningens , был помещен в новый вид Pteranodon Marshi , а череп, назначенный P. occidentalis, был переименован в Pteranodon Eatoni . Миллер также узнал другой вид, основанный на черепе с гребнем, похожим на гребень P. sternbergi ; Миллер назвал это Птеранодон Уокери . Чтобы помочь внести порядок в этом путанице имен, Миллер создал для них три категории или «подрост». П. Марши и П. Лонгиспс были помещены в подрод Longicepia , хотя позже это было изменено на просто Pteranodon из -за правил приоритета. П. Стернберги и П. Уокери , вертикально-хриплую виду, получили подрод Стернбергию , которая впоследствии была изменена на Геертербергию, потому что Стернбергия уже использовалась («озабоченна»). Наконец, Миллер назвал подрод Occidentalia для P. eatoni , черепа, ранее связанный с P. occidentalis . Миллер дополнительно расширил концепцию Pteranodon , чтобы включить Nyctosaurus в качестве четвертого подрода. Миллер считал, что это эволюционное прогрессирование, с примитивным Nyctosaurus , в то время считается без гребней, что привело к ростам (с небольшим гребнем), который, в свою очередь, породил птеродон с его длинным задом наперед, наконец, приводя к геостергии С его большим, вертикальным гребнем. Тем не менее, Миллер сделал несколько ошибок в своем исследовании, касающихся того, какие образцы направили болот, и большинство ученых не принимали участие в его работе над этим предметом в их более позднем исследовании, хотя Wellnhofer (1978) последовал за списком видов Миллера. и Schoch (1984) несколько странно опубликовал еще одну пересмотр, которая по существу вернулась в первоначальную схему классификации Марша, особенно понижение P. longiceps как синоним P. Ingens . [ 7 ]
Распознавание вариаций
[ редактировать ]В начале 1990 -х годов С. Кристофер Беннетт также опубликовал несколько крупных документов, рассмотренных анатомии, таксономии и жизни Птеранодона . [ 12 ]
Фрагментарные окаменелости, назначенные в Птеранодон, также были обнаружены в Сконе , Швеция . [ 13 ]
Описание
[ редактировать ]
Виды Pteranodon очень хорошо представлены в ископаемых записях, что позволяет подробно описания их анатомии и анализа их истории жизни. Было выявлено более 1000 образцов, хотя менее половины из них достаточно полны, чтобы дать исследователям хорошую анатомическую информацию. Тем не менее, это больше материал окаменелостей, чем известен для любого другого птерозавра, и включает как мужские, так и женские образцы различных возрастных групп, так и, возможно, виды. [ 5 ]
Взрослые образцы Pteranodon от двух основных видов можно разделить на два различных класса размера. Меньший класс образцов имеет маленькие, округлые гребня для голов и очень широкие тазовые каналы, даже шире, чем у класса гораздо большего размера. Размер тазового канала, вероятно, позволил откладывать яйца, что указывает на то, что эти меньшие взрослые являются женщинами. Класс большего размера, представляющий мужчин, имеет узкие бедра и очень большие гребни, которые, вероятно, были для отображения.
Взрослый мужской птеродон были одними из крупнейших птерозавров и были самыми большими летающими животными, известными до конца 20 -го века, когда были обнаружены гигантские азархидские птерозавры. Размах крыльев среднего взрослого мужчины Птерододона составлял 5,6 м (18 футов). Взрослые женщины были намного меньше, в среднем 3,8 м (12 футов) в размахе крыльев. [ 5 ] По оценкам, большой образец Pteranodon Longiceps , USNM 50130, имеет размах крыльев 6,25–6,5 м (20,5–21,3 фута), длину тела 2,6 м (8,5 фута) и массу тела 50 кг (110 фунтов). [ 5 ] [ 14 ] [ 15 ] [ 16 ] Еще более крупные образцы имели крылья 7,25–7,6 м (23,8–24,9 фута). [ 5 ] [ 17 ] В то время как большинство образцов найдены раздавленными, существует достаточно окаменелостей, чтобы собрать подробное описание животного.

Методы, используемые для оценки массы крупных мужских образцов Pteranodon (с помощью крыльев около 7 метров), были общеизвестно ненадежными, что дает широкий спектр оценок. В обзоре оценок размера птерозавра, опубликованных в 2010 году, исследователи Марк Виттон и Майк Хабиб утверждали, что самая большая оценка 93 кг слишком высока, а верхний предел от 20 до 35 кг является более реалистичной. Виттон и Хабиб рассмотрели методы, используемые исследователями, которые получили меньшие оценки массы, одинаково испорченные. Большинство из них были получены путем масштабирования современных животных, таких как летучих мышей и птиц до размера птеродона , несмотря на то, что у птерозавров есть совершенно разные пропорции тела и анатомию мягких тканей от любого живого животного. [ 18 ]
Другие отличительные характеристики, которые отличают птеродона от других птерозавров, включают узкие нейронные шипы на позвонках, тарелки, похожие на пластинчатые костчивые связки, укрепляющие позвонки над бедром, и относительно короткий хвост, в котором последние несколько позвонок сливаются в длинный стержень. [ 19 ] Вся длина хвоста была примерно на 3,5%, пока размах крыльев или до 25 сантиметров (9,8 дюйма) у самых крупных мужчин. [ 19 ]
Череп и клюв
[ редактировать ]
В отличие от более ранних птерозавров, таких как Rhamphorhynchus и Pterodactylus , у Pteranodon были беззубые клювы , аналогичные красам птиц . Клюки птеродона были сделаны из твердых костюшных полей, которые проецировались с основания челюстей. Клюки были длинными, стройными и заканчивались тонкими, острыми точками. Верхняя челюсть, которая была длиннее нижней челюсти, была изогнута вверх; В то время как это обычно объясняется только клювом вверх, один образец (UALVP 24238) имеет кривизну, соответствующую расширению клюва к кончику. В то время как кончик клюва не известен в этом образце, уровень кривизны предполагает, что он был бы чрезвычайно длинным. Уникальная форма клюва в этом экземпляре заставила Александра Келлнера назначить его четкому роду Dawndraco , в 2010 году. [ 12 ]
Наиболее отличительной характеристикой Pteranodon является его черепный гребень. Эти гребни состояли из костей черепа (фронталы), выступающих вверх и назад от черепа. Размер и форма этих гребней варьировались из -за ряда факторов, включая возраст, пол и виды. Самец Птеранодон Стернберги , более старые виды двух, описанных на сегодняшний день, имели более вертикальный гребень с широкой прогнозой, в то время как их потомки, птерододон-лонгийспс , развивались более узкий, более обратный герб. [ 4 ] Самки обоих видов были меньше и носили маленькие, округлые гребни. [ 7 ] Гребники, вероятно, были в основном отображаемыми структурами, хотя у них, возможно, были и другие функции. [ 20 ]
Палеобиология
[ редактировать ]Полет
[ редактировать ]
Форма крыла Птеранодона предполагает, что оно летало бы скорее как современный альбатрос . Это основано на том факте, что у Птерододона было высокое соотношение сторон (размах крыльев к аккорде ), аналогичный альбатросу - 9: 1 для Птеронодона , по сравнению с 8: 1 для альбатроса. Альбатросы проводят длинные промежутки времени при морской рыбалке и используют схему полета, называемую « динамическим парящим », который использует вертикальный градиент скорости ветра вблизи поверхности океана, чтобы проехать на большие расстояния без распада и без помощи термиков (которые не встречаются над открытым океаном так же, как они делают на земле). [ 21 ] В то время как большая часть полета Pteranodon зависела от паря, как и длинные морские птицы, это, вероятно, потребовалось случайное активное, быстрое разрыв хлопания и исследования нагрузки крыла Pteranodon (сила крыльев против веса тела) Укажите, что они были способны к существенному хлопковому полету, вопреки некоторым более ранним предположениям, что они были настолько большими, что могли только скользить. [ 18 ] Тем не менее, более недавнее исследование предполагает, что оно полагалось на тепловое паря, в отличие от современных морских птиц, но во многом как современные континентальные флаеры и вымершие пелагорнис . [ 22 ]
Как и другие птерозавры, Птеродон, вероятно, вылетел из стоячей, четвероногих положений. Используя свои длинные передние конечности для рычага, они бы сводились в воздух в быстром скачке. Почти вся энергия была бы генерирована передне. Поверхность крыльев произошел бы, когда животное очистило землю, а затем быстрый удар вниз, чтобы создать дополнительный подъем и завершить запуск в воздух. [ 18 ]
Наземная локомоция
[ редактировать ]
Исторически, наземная локомоция Птеронодона , особенно будь то двуножество или четвероногих , стала предметом дебатов. Сегодня большинство исследователей птерозавра согласны с тем, что птерозавры были четвероногими, во многом благодаря открытию птерозавра . [ 23 ]
Возможность водного локомоции через плавание была кратко обсуждена в нескольких статьях (Bennett 2001, 1994, и Bramwell & Whitfield 1974).
Диета
[ редактировать ]
Известно, что диета птеродона включала рыбу ; Ископаемые рыбные кости были обнаружены в области желудка одного птеродона , и птеродона между челюстями другого , образец AMNH 5098. Рыба составляла большую часть диеты Pteranodon ( хотя они также могли принять беспозвоночных). [ 5 ]
Традиционно большинство исследователей предполагают, что Птеранодон взял бы рыбу, погружая свои клювы в воду во время низкого, парящего полета. Тем не менее, это, вероятно, было основано на предположении, что животные не могут взлететь с поверхности воды. [ 5 ] Более вероятно, что Птеранодон мог взлетать с воды и погрузился бы на рыбу во время плавания, а не во время полета. Даже небольшая женская птеродон мог бы достичь глубины не менее 80 сантиметров (31 дюйм) с его длинным счетом и шеей во время плавания на поверхности, и они могли достичь еще больших глубин, погрузившись в воду с воздуха Как некоторые современные длиннокрытые морские птицы. [ 5 ] В 1994 году Беннетт отметил, что голова, шея и плечи птеродона были так же сильно построены, как и птицы, и предположили, что они могут погрузиться, откладывая свои крылья, как современная ганнет . [ 5 ]
Функция гребня
[ редактировать ]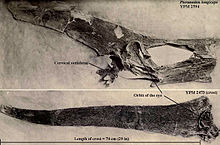
Pteranodon был замечен своим гребнем черепа, хотя функция этого гребня была предметом дебатов. Однако большинство объяснений были сосредоточены на лезвии, обратно заостренном гребне мужского P. longiceps , и игнорировали широкий спектр вариаций по возрасту и полу. Тот факт, что гребни различаются так много, определяют большинство практических функций, кроме как для использования на спаривающихся дисплеях. [ 24 ] Следовательно, дисплей был, вероятно, основной функцией гребня, и любые другие функции были вторичными. [ 20 ]
Научные интерпретации функции гребня начались в 1910 году, когда Джордж Фрэнсис Итон предложил две возможности: аэродинамический уравновешенность и точка мышечной прикрепления. Он предположил, что гребень мог бы закрепить большие длинные челюстные мышцы, но признал, что одна функция не может объяснить большой размер некоторых гребней. [ 25 ] Беннетт (1992) согласился с собственной оценкой Итона о том, что гребень был слишком большим и переменным, чтобы быть местом привязанности к мышцам. [ 20 ] Итон предположил, что вторичная функция гребня могла быть как противовес на длинный клюв, уменьшая необходимость в тяжелых мышцах шеи, чтобы контролировать ориентацию головы. [ 25 ] Испытания на аэродинамическую трубу показали, что гребень действительно функционировал в качестве эффективного противовеса в некоторой степени, но Беннетт отметил, что, опять же, гипотеза фокусируется только на длинных гребнях мужского P. longiceps , а не на более крупных гребнях P. sternbergi и очень маленьких гребни, которые существовали среди женщин. Беннет обнаружил, что гребни женщин не имели противовеса, и что гребни мужчины П. Стернберги сами по себе не будут оказывать негативное влияние на баланс головы. Фактически, для контроля баланса для контроля баланса потребовало бы большего, а не мускулатуру шеи. [ 20 ]
В 1943 году Доминик фон Крипп предположил, что гребень, возможно, служил рулем , идея, принятая несколькими более поздними исследователями. [ 20 ] [ 26 ] Один из исследователей, Росс С. Стейн, даже предположил, что гребень, возможно, поддерживал мембрану кожи, соединяющую заднюю гребень с шеей и спиной, увеличивая площадь поверхности и эффективность в качестве руля. [ 27 ] Гипотеза руля, опять же, не учитывает женщин, ни П. Стернберги , у которого был восхитительный, а не наоборот. Беннет также обнаружил, что даже в качестве руля, гребень не обеспечит почти столько направленной силы, сколько просто маневрировать крыльями. Предположение о том, что гребень был воздушным тормозом, и что животные поворачивают головы в сторону, чтобы замедлиться, страдает от аналогичной проблемы. [ 28 ] Кроме того, гипотезы руля и воздушного тормоза не объясняют, почему такие большие вариации существуют в размере гребня даже среди взрослых. [ 20 ]
Александр Келлнер предположил, что большие гребни птерозавра ленты , а также другие виды могут использоваться для теплообмена, что позволяет этим птерозаврам поглощать или сбросить тепло и регулировать температуру тела, что также будет учитывать корреляцию между размером и телом размер. Однако для этой цели нет никаких доказательств дополнительных кровеносных сосудов, и крупные мембранные крылья, заполненные кровеносными сосудами, послужили бы этой целью гораздо эффективнее. [ 20 ]
С этими гипотезами исключили, что наиболее поддерживаемая гипотеза для функции гребня, по-видимому, является сексуальным показом. Это согласуется с изменением размера, наблюдаемых в ископаемых образцах, где женщины и несовершеннолетние имеют небольшие гребни и мужчины большие, сложные, переменные гребни. [ 20 ]
Сексуальные вариации
[ редактировать ]
Образцы для взрослых Pteranodon могут быть разделены на два различных класса размера, маленькие и большие, а класс большого размера составляет примерно в полтора раза больше, чем маленький класс, а небольшой класс был в два раза больше, чем большой класс. Оба класса размера жили рядом друг с другом, и, хотя исследователи ранее предположили, что они представляют разные виды, Кристофер Беннетт показал, что различия между ними согласуются с концепцией, что они представляют самок и мужчин, и что птеродона виды были сексуально диморфными . Черепа из класса большего размера сохраняют большие, вверх и назад гребни, направленные на направление, в то время как гребни класса меньшего размера маленькие и треугольные. Некоторые более крупные черепа также показывают доказательства второго гребня, который простирался длинным и низким, к кончику клюва, который не наблюдается в небольших образцах. [ 20 ]
Пол классов различных размеров был определен не из черепов, а из костей таза. Вопреки тому, что можно ожидать, класс меньшего размера имел непропорционально большие и широкоутолечные тазовые кости. Беннетт истолковал это как указывающий на более просторный родовой канал, через который проходили яйца. Он пришел к выводу, что небольшой класс с небольшими треугольными гребнями представляют самок, а более крупные, большие хитровые образцы представляют мужчин. [ 20 ]
Обратите внимание, что общий размер и размер гребня также соответствуют возрасту. Незрелые образцы известны как у женщин, так и у мужчин, и незрелые мужчины часто имеют небольшие гребни, сходные с взрослыми женщинами. Следовательно, кажется, что крупные гребни развивались только у мужчин, когда они достигли своего большого, взрослого размера, что затрудняет установление пола незрелых образцов из частичных останков. [ 14 ]
Тот факт, что женщины, по-видимому, имеют превосходные мужчины с двумя к одному, предполагает, что, как и в случае с современными животными с сексуальным диморфизмом, связанными с размерами, такими как морские львы и другие хриптивы , птеронодон мог быть полигинными , и несколько мужчин, конкурирующих за ассоциацию с группами, состоящими большого количества женщин. Подобно современным выпечкам, Pteranodon , возможно, соревновался, чтобы создать территорию на скалистых, оффшорных корпусах, с самыми большими и крупнейшими хитрыми, мужчины, получающие наибольшую территорию и имеют больший успех спариваться с женщинами. Гребники мужского Pteranodon не использовались бы в соревнованиях, а скорее как «символы визуального доминирования», с ритуалами дисплея, занимающих место физической конкуренции с другими мужчинами. Если эта гипотеза верна, вполне вероятно, что мужской птеродон практически не играл в воспитании молодежи; Такое поведение не встречается у мужчин современных полигинозных животных, которые одновременно отчитывают много потомков. [ 20 ]
Палеоэкология
[ редактировать ]
Образцы, назначенные Птеранодону, были обнаружены как в отложениях мела Smoky Hill в формировании Ниобрары , так и в немного более молодых месторождениях Шарон -Спрингс формирования сланца Пьера . Когда Птеранодон был жив, этот район был покрыт большим внутренним морем, известным как западный интерьер . Известные окаменелостями, собранными с 1870 года, эти формирования простираются от юга до Канзаса в Соединенных Штатах до Манитобы в Канаде. Тем не менее, образцы птеродона (или любые образцы птерозавра) были обнаружены только в южной половине формирования, в Канзасе, Вайоминге и Южной Дакоте . Несмотря на то, что в современных частях формирования в Канаде были обнаружены многочисленные окаменелости, образцы птерозавров никогда не было обнаружено. Это убедительно говорит о том, что естественный географический диапазон птеродона покрывал только южную часть Нибрары, и что его среда обитания не простиралась дальше на север, чем Южная Дакота. [ 7 ]
Некоторые очень фрагментарные окаменелости, принадлежащие к птеродонтианским птерозаврам, и, возможно, сам Птеранодон , также были обнаружены на побережье Мексиканского залива и восточном побережье Соединенных Штатов . Например, некоторые фрагменты костей из Муревилля Алабамы . и формирования Мерчанвилля Делавэра формирования могли поступить из Птеронодона , хотя они слишком неполны, чтобы сделать определенную идентификацию [ 7 ] Некоторые останки из Японии также были предварительно приписаны Птеранодону , но их расстояние от ее известной западной внутренней среды обитания морских путей делает эту идентификацию маловероятным. [ 7 ]

Pteranodon Longiceps поделил бы небо с гигантским хищником Nyctosaurus . По сравнению с P. longiceps , который был очень распространенным видом, Nyctosaurus был редким, составлял только 3% окаменелостей птерозавра из формирования. Также менее распространенной была ранняя зубчатая птица , Ихтиорнис . [ 29 ]
Вполне вероятно, что, как и у других полигинозных животных (в которых мужчины конкурируют за ассоциацию с гаремами женщин), Птанодон жил в основном на оффшорных петух, где они могли уклоняться от наземных хищников и кормить далеко от берега; Большинство окаменелостей птеродона встречаются в местах, которые в то время находились в сотнях километров от береговой линии. [ 20 ]
Под поверхностью море было заполнено в основном беспозвоночными, такими как аммониты и кальмары . Жизнь позвоночных, кроме базальной рыбы, включала морские черепахи , такие как токсошели , плезиоооры Elasmosaurus и Styxosaurus , а также безлетная птица Diving ParaHesperornis . Мозасавры были наиболее распространенными морскими рептилиями, с родами, включая клидаст , мозазавр и тилозавр . [ 4 ] По крайней мере, некоторые из этих морских рептилий, как известно, питались на птеродоне . Barnum Brown , в 1904 году, сообщил о содержимое желудка Plesiosaur, содержащее кости «птеродактиль», скорее всего, от Pteranodon . [ 30 ]
Окаменелости от наземных динозавров также были обнаружены в мелах Нибрары, что позволяет предположить, что животных, которые умерли на берегу, должны были быть вымыты в море (один образец хадрозавра, по -видимому, был выбит акулой ) . [ 31 ]
Классификация
[ редактировать ]Времена и эволюция
[ редактировать ]
Окаменелости птеродона известны в первую очередь из формирования Ниобрары Центральных Соединенных Штатов. В широком смысле, Птанодон существовал более четырех миллионов лет, на стадии сантонианской стадии мелового периода. [ 7 ] Род присутствует в большинстве слоев формирования Ниобрары, за исключением двух верхних двух; В 2003 году Кеннет Карпентер обследовал распределение и датирование окаменелостей в этой форме, демонстрируя, что Птеранодон Стернберги существовал там с 88 до 85 миллионов лет назад, в то время как P. longiceps существовал от 86 до 84,5 миллионов лет назад. Возможный третий вид, который Келлнер назвал Geosternbergia Maiseyi в 2010 году, известен от члена Шарон -Спрингс -Формирования сланца в Канзасе, Вайоминге и Южной Дакоте , датируемой от 81,5 до 80,5 миллионов лет назад. [ 29 ]
В начале 1990 -х годов Беннетт отметил, что два основных морфа птерододонта, присутствующих в формации Нибрары, были точно разделены во времени с небольшим перекрытием. Из -за этого и с их общим сходством он предположил, что они, вероятно, представляют хроноспецифические в рамках единой эволюционной линии, которая длится около 4 миллионов лет. Другими словами, только один вид Pteranodon присутствовал бы в любое время, а P. sternbergi (или Geasternbergia ), по всей вероятности, был прямым видом предков P. longiceps . [ 5 ]
Действительный вид
[ редактировать ]Многие исследователи считают, что есть как минимум два вида птеродона . Однако, помимо различий между мужчинами и женщинами, описанными выше, посткраниальные скелеты птеродона показывают незначительное или отсутствие различий между видами или образцами, а тела и крылья всех птерододонтов были по существу идентичными. [ 7 ]
Два вида Pteranodon традиционно признаются как достоверные: Pteranodon longiceps , типовые виды и птеродон -штернберги . Виды различаются только по форме гребня у взрослых мужчин (описанных выше) и, возможно, под углом некоторых костей черепа. [ 7 ] Поскольку хорошо сохранившиеся окаменелости черепа Pteranodon чрезвычайно редки, исследователи используют стратиграфию (то есть, какой слой породы геологического образования, в большинстве случаев, в большинстве случаев определение видовой идентичности.
Pteranodon Sternbergi является единственным известным видом Pteranodon с вертикальным гребнем. Нижняя челюсть П. Стернберги составляла 1,25 метра (4,1 фута) длиной. [ 32 ] Он был собран Джорджем Ф. Стернбергом в 1952 году и описан Джоном Кристианом Харксеном в 1966 году из нижней части формирования Ниобрары. Он был старше P. longiceps и считается Беннеттом прямым предком более позднего вида. [ 7 ]
Поскольку окаменелости, идентифицируемые как P. sternbergi , обнаружены исключительно в нижних слоях формирования Niobrara, и ископаемые P. longiceps исключительно в верхних слоях, в нем может быть идентифицировано окаменелости, на основе его положения в геологической колонке (хотя для многих. Ранние ископаемые находки, точные данные о его местоположении не были зарегистрированы, что делало много окаменелостей неопознаваемыми). [ 12 ]

Ниже приведена кладограмма , показывающая филогенетическое размещение этого рода в Pteranodontia от Andres and Myers (2013). [ 33 ]
| Pteranodontia |
| ||||||||||||||||||||||||
Альтернативные классификации
[ редактировать ]
Из -за тонких изменений между образцами птеронодонтида из формации Нибрары большинство исследователей назначили их всем им на единый род Pteranodon , по крайней мере, у двух видов ( P. longiceps и P. Sternbergi ), отличающимися в основном по форме гребня. Тем не менее, классификация этих двух форм варьировалась от исследователя к исследователю. В 1972 году Хэлси Уилкинсон Миллер опубликовал статью, утверждая, что различные формы птеродона были достаточно различными, чтобы быть помещенными в различные подростки. Он назвал эти Pteranodon (Occidentalia) occidentalis (для ныне раззумученных видов P. occidentalis ) и Pteranodon (Sternbergia) Sternbergi . Тем не менее, название Sternbergia было озабочено, и в 1978 году Миллер повторно назвал вида Pteranodon (Geosternbergia) Sternbergi и назвал комбинацию третьего подрода/видов для P. longiceps , как Pteranodon (Longicepia) Longiceps . Однако наиболее выдающиеся исследователи птерозавра в конце 20 -го века, включая С. Кристофер Беннетт и Питер Велнхофер не принял эти субгенные названия и продолжал размещать все виды PteranoDont в однознанный Pteranodon .
В 2010 году исследователь птерозавра Александр Келлнер пересмотрел классификацию HW Miller. Келлнер последовал за мнением Миллера, что различия между видами птеродона были достаточно великими, чтобы поместить их в разные роды. Он поместил П. Стернберги в род, названный Миллером, Гетернбергия , вместе с образцом черепа Пьера, который, ранее считал Беннетт крупным мужским P. longiceps . Келлнер утверждал, что гребень этого образца, хотя и не полностью сохраненный, был наиболее похож на Гетернбергию . Поскольку образец был миллионы лет моложе любой известной Geosternbergia , он назначил его новым видам Geosternbergia Maiseyi . Многочисленные другие образцы Pteranodont известны из того же периода формирования и времени, и Келлнер предположил, что они могут принадлежать к тому же виду, что и Г. Масея , но, поскольку им не хватает черепов, он не мог с уверенностью идентифицировать их. [ 12 ] Тем не менее, оба вида, ранее упоминавшиеся в геостербергию, были отдельно включены как виды Pteranodon ( P. Sternbergi и P. maiseyi ) на основе филогенетического анализа в 2024 году. [ 34 ]
Разработанные виды
[ редактировать ]
С 1870 -х годов был назван ряд дополнительных видов Pteranodon , хотя большинство в настоящее время считаются младшими синонимами двух или трех действительных видов. Лучше всего поддерживается типовой вид , P. longiceps , основанный на хорошо сохранившемся образе, включая первый известный череп, найденной SW Williston. У этого человека был размах крыльев 7 метров (23 фута). [ 35 ] Другие достоверные виды включают, возможно, больше P. sternbergi , с размахом крыльев, первоначально оцениваемым в 9 м (30 футов). [ 35 ] P. Oweni ( P. occidentalis ), P. Velox , P. Umbrosus , P. Harpyia и P. comptus считаются номинами Dubia Беннеттом (1994) и другими, которые ставят под сомнение их достоверность. Все, вероятно, являются синонимом более известных видов.
Поскольку ключевым отличительным характерным болотом, отмеченным для Птеронодона, было отсутствие зубов, любой беззубевый фрагмент челюсти из беззубой, где бы он ни был обнаружен в мире, имел тенденцию охватывать Птеронодон в течение конца девятнадцатого и начала двадцатого веков. Это привело к множеству видов и большой путанице. Название стало таксоном мусора динозавров , скорее как мегалозавр , чтобы пометить любые останки птерозавра, которые нельзя различить, кроме отсутствия зубов. Виды (часто сомнительные, которые, как известно, в настоящее время основаны на сексуальных вариациях или юношеских символах) были реклассифицированы несколько раз, и несколько подрост в 1970 -х годах было возведено Subgenera включает в себя Longicepia , Occidentalia и Geasternbergia ). Примечательные авторы, которые обсудили различные аспекты Птеранодона, включают Беннетт, Падиан, Анвин, Келлнер и Веллнхофер. Два вида, P. oregonensis и P. orientalis не являются птеранодонтидами и были переименованы в Bennettazhia Oregonensis и Bogolubovia orientalis соответственно.
Список видов и синонимов
[ редактировать ]Статус названий, перечисленных ниже, следуйте опросу Bennett, 1994, если не указано иное. [ 7 ]
| Имя | Автор | Год | Статус | Примечания |
|---|---|---|---|---|
| Pterodactylus oweni | Болото | 1871 | Имя без сомнения | Переименовано в Pterodactylus occidentalis Marsh 1872 на основе Оуени , озабоченной "Pterodactylus Oweni" Seeley 1864 ( nomen nudum для Ornithocheirus Oweni Seeley 1870) |
| Pterodactylus Ingens | Болото | 1872 | Переклассифицирован как птеродон | |
| Pterodactylus occidentalis | Болото | 1872 | Младший объективный синоним pterodactylus oweni | Реклассифицирован из Pterodactylus Oweni Marsh 1871 на основе P. oweni , озабоченного "Pterodactylus Oweni" Seeley 1864 ( Nomen Nudum для Ornithocheirus Oweni Seeley 1870) |
| Pterodactylus Velox | Болото | 1872 | Имя без сомнения | Реклассифицирован как Pteranodon Velox |
| Орнитохирус Тень | Справляться | 1872 | Имя без сомнения | |
| Орнитохирус Harpyia | Справляться | 1872 | Имя без сомнения | |
| Pterodactylus Umbrosus | (Справиться) справиться | ( 1872 ) 1874 | Реклассификация орнитохируса Umbrosus | |
| Pteranodon longiceps | Болото | 1876 | Действительный | Тип видов |
| Pteranodon Огромный | (Марш) Уиллистон | ( 1872 ) 1876 | Имя без сомнения | Переклассифицирован из Pterodactylus Ingens |
| Pteranodon occidentalis | Болото | ( 1872 ) 1876 | Младший объективный синоним pterodactylus oweni | Переклассифицирован из pterodactylus occidentalis |
| Pteranodon Velox | Болото | ( 1872 ) 1876 | Имя без сомнения | Рекссифицирован из Pterodactylus Velox , на основе ювенильного образца |
| Pteranodon Gracilis | Болото | 1876 | Реклассифицирован как Nyctosaurus gracilis | |
| Pteranodon Kettle | Болото | 1876 | Имя без сомнения | |
| Птерододон Дварф | Болото | 1876 | Реклассифицирован как Nyctosaurus naus | |
| Ornithocheirus shadow | (Cope) Ньютон | ( 1872 ) 1888 | Реклассифицирован как Pteranodon Umbrosus | Правописание орнитохируса Umbrosus |
| Ornithocheirus harpyia | (Cope) Ньютон | ( 1872 ) 1888 | Реклассифицирован как Pteranodon Harpyia | Правописание орнитохирусной гарпии |
| Pteranodon Umbrosus | (Коуп) Уиллистон | ( 1872 ) 1892 | Имя без сомнения | Реклассификация орнитохируса Umbrosus |
| Ornithostoma огромный | (Марш) Уиллистон | ( 1872 ) 1893 | Синонимы Pteranodon огромного | Переклассифицирован из птеранодон |
| Орнитостома оттенок | (Коуп) Уиллистон | ( 1872 ) 1897 | Синоним Pteranodon Umbrosus | Реклассифицирован из Pteranodon Umbrosus |
| Pteranodon Oregonensis | Гилмор | 1928 | Реклассифицирован как Беннеттцзия Oregonensis | |
| Птеранодон Стернберги | Харк | 1966 | Действительный | |
| Pteranodon marshi | Миллер | 1972 | Синоним Pteranodon longiceps | |
| Pteranodon Bonneri | Миллер | 1972 | Реклассифицирован как Nyctosaurus bonneri | |
| ПЕРАНОДОН Уокер | Миллер | 1972 | Синоним Pteranodon longiceps | |
| Pteranodon (Occidentalia) eatoni | (Миллер) Миллер | ( 1972 ) 1972 | Синоним птеродона Стернберги | |
| Pteranodon eatoni | (Миллер) Миллер | ( 1972 ) 1972 | Синоним птеродона Стернберги | Реклассифицирован из Pteranodon (Occidentalia) eatoni |
| Pteranodon (Longicepia) Longicps [ sic ] | (Болото) Миллер | ( 1872 ) 1972 | Синоним Pteranodon longiceps | Переклассифицирован из Pteranodon longiceps |
| Pteranodon (Longicepia) marshi | (Миллер) Миллер | ( 1972 ) 1972 | Синоним Pteranodon longiceps | Reclassified from Pteranodon marshi |
| Pteranodon (Sternbergia) Стернберги | (Харксен) Миллер | ( 1966 ) 1972 | Рекссифицирован как птеродон (Geosternbergia) sternbergi | Переклассифицирован из Птерододонского Стернберги |
| ПЕРАНОДОН (Стернбергия) Уокер | (Миллер) Миллер | ( 1972 ) 1972 | Рекссифицирован как птеродон (Geososternbergia) Walker | Переклассифицирован из Pteranodon Walkeri |
| Pteranodon (Pteranodon) marshi | (Миллер) Миллер | ( 1972 ) 1973 | Синоним Pteranodon longiceps | Reclassified from Pteranodon marshi |
| Pteranodon (Western) West | (Марш) Ольшевский | ( 1872 ) 1978 | Синоним Pteranodon occidentalis | Переклассифицирован из Pteranodon occidentalis |
| Pteranodon (Longicepia) Огромный | (Марш) Ольшевский | ( 1872 ) 1978 | Синонимы Pteranodon огромного | Переклассифицирован из птеранодон |
| Pteranodon (Pteranodon) Inning | (Марш) Ольшевский | ( 1872 ) 1978 | Синонимы Pteranodon огромного | Переклассифицирован из птеранодон |
| Pteranodon (Geososternbergia) Уокер | (Миллер) Миллер | ( 1972 ) 1978 | Синоним Pteranodon longiceps | Переклассифицирован из Pteranodon Walkeri |
| Pteranodon (Geasternbergia) Sternbergi | (Харксен) Миллер | ( 1966 ) 1978 | Синоним птеродона Стернберги | Реклассифицирован из птеродона (Стернбергия) Стернберги |
| Pteranodon Orientalis | (Bogolubov) Nesov & Yarkov | ( 1914 ) 1989 | Реклассифицирован как Bogolubovia Orientalis | Реклассифицирован из Ornithostoma orientalis |
| Geosterbergia Walker | (Миллер) Ольшевский | ( 1972 ) 1991 | Синоним птеродона Стернберги | Рекссифицирован из птеродона (Стернбергия) Уокри |
| Geosterbergia Sternberg | (Харксен) Ольшевский | ( 1966 ) 1991 | Синоним птеродона Стернберги | Рекссифицирован из Птеронодона (Геертербергия) Стернберги |
Смотрите также
[ редактировать ]Ссылки
[ редактировать ]- ^ Старки, Ребекка; Хабиб, Майкл; Лица, В. Скотт (2024-06-01). «Крыло птеронодонтидного со следами обширного поглощения беспозвоночных от Pierre Shale (участник Шарон -Спрингс, Кампанийский - Маастрихтский), Восточный Центральный Вайоминг (округ Нибрара)» . Мерашные исследования . 158 : 105837. DOI : 10.1016/j.cretres.2024.105837 . ISSN 0195-6671 .
- ^ Колберт, Эдвин Х. (Эдвин Харрис); Рыцарь, Чарльз Роберт (1951). Книга динозавров: правящие рептилии и их родственники . Нью-Йорк: МакГроу-Хилл. п. 153
- ^ Эрет, DJ; Harrell, TL Jr. (2018). «Следы кормления на птеродонской (рептилия: Птерозаврия) кость из покойного мелового (кампанианского) мела Муревилля в Алабаме, США». Палаис . 33 (9): 414–418. Bibcode : 2018palai..33..414e . doi : 10.2110/palo.2018.024 . S2CID 135332458 .
- ^ Jump up to: а беременный в дюймовый Беннетт, Южная Каролина (2000). «Вывод стратиграфического положения ископаемых позвоночных из ниобрара мела Западного Канзаса». Текущие исследования в области наук о Земле: Геологический бюллетень Канзаса , 244 (часть 1): 26 стр.
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж Беннетт, Южная Каролина (1994). «Птерозавры ниобрара мела». Ученый Земля . 11 (1): 22–25.
- ^ Виттон, Марк Пол (2010). «Птеранодон и за его пределами: история гигантских птерозавров с 1870 года» . Геологическое общество, Лондон, Специальные публикации . 343 (1): 313–323. BIBCODE : 2010GSLSP.343..313W . doi : 10.1144/sp343.19 . S2CID 128801077 - через ResearchGate.
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л м не а п Q. ведущий с Беннетт, Южная Каролина (1994). «Таксономия и систематика позднего мелового птерозавра птеронодон (Pterosauria, pterodactyloida)». Случайные документы Музея естественной истории, Университет Канзаса . 169 : 1–70.
- ^ Cope, ed (1875). «Позвонки меловых образов Запада». Отчет, Геологическая служба США территорий (Хейден) , 2 : 302 с., 57 пл.
- ^ Marsh, OC (1876a). «Уведомление о новом подпоможне Птерозаврии» . Американский журнал науки . Серия 3. 11 (65): 507–509. Bibcode : 1876 Amsjs ... 11..507M . doi : 10.2475/ajs.s3-11.66.507 . S2CID 130203580 .
- ^ Marsh, OC (1876b). «Основные персонажи американских птеродактилов» . Американский журнал науки . Серия 3. 12 (72): 479–480. Bibcode : 1876 Amsjs ... 12..479M . doi : 10.2475/ajs.s3-12.72.479 . S2CID 131057784 .
- ^ Averianov, AO (2012). « Ornithostoma sedgwicki - действительный таксон азархоидных птерозавров» . Труды зоологического института Рас . 316 (1): 40–49. doi : 10.31610/trudyzin/2012.316.1.40 . S2CID 67809186 .
- ^ Jump up to: а беременный в дюймовый Келлнер, Ава (2010). «Комментарии к Pteranodontidae (Pterosauria, Pterodactyloidea) с описанием двух новых видов» (PDF) . Anais Da Academia Brasileira de Ciências . 82 (4): 1063–1084. doi : 10.1590/s0001-37652010000400025 . PMID 21152777 .
- ^ Все в науке, 10-2012, с.72
- ^ Jump up to: а беременный Беннетт, Южная Каролина (2001). «Остеология и функциональная морфология позднего мела птерозавра птерододона . Общее описание остеологии». Palaeontographica, Abteilung a . 260 : 1–112. doi : 10.1127/pala/260/2001/1 . S2CID 90380603 .
- ^ Larramendi, A.; Пол, GS; HSU, S.-Y. (2021). «Обзор и переоценка специфической тяжести существующих и прошлых многоклеточных организмов, с акцентом на тетраподы» . Анатомическая запись . 304 (9): 1833–1888. doi : 10.1002/ar.24574 . PMID 33258532 .
- ^ Пол, Грегори С. (2022). Полевой гид Принстона к птерозаврам . ПРИЗНАЯ УНИВЕРСИТЕТА ПРИСЕТА. С. 170–172. doi : 10.1515/9780691232218 . ISBN 9780691232218 Полем S2CID 249332375 .
- ^ Виттон, Марк Пол (2010). «Птеранодон и за его пределами: история гигантских птерозавров с 1870 года» . Геологическое общество, Лондон, Специальные публикации . 343 (1): 313–323. BIBCODE : 2010GSLSP.343..313W . doi : 10.1144/sp343.19 . S2CID 128801077 - через ResearchGate.
- ^ Jump up to: а беременный в Виттон, депутат; Habib, MB (2010). «На размер и разнообразие полетов гигантских птерозавров использование птиц в качестве аналогов птерозавров и комментариев о бегемости птерозавра» . Plos один . 5 (11): E13982. BIBCODE : 2010PLOSO ... 513982W . doi : 10.1371/journal.pone.0013982 . PMC 2981443 . PMID 21085624 .
- ^ Jump up to: а беременный Беннетт, Южная Каролина (1987). «Новые доказательства на хвосте птерозавра птерододона ( Archosauria: Pterosauria)». Стр. 18–23 в Керри, П.Дж. и Эх Костер (ред.), Четвертый симпозиум на мезозойских наземных экосистемах, короткие бумаги. Случайные документы Музея палеонтологии Тиррелла, № 3.
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л Беннетт, Южная Каролина (1992). «Сексуальный диморфизм птеродона и других птерозавров, с комментариями о черепных гребнях». Журнал палеонтологии позвоночных . 12 (4): 422–434. doi : 10.1080/02724634.1992.10011472 .
- ^ Падиан К. (1983). «Функциональный анализ полетов и ходьбы в птерозаврах». Палеобиология . 9 (3): 218–239. doi : 10.1017/s009483730000765x . S2CID 88434056 .
- ^ Гото, Юсуке; Йода, Кен; Веймерскирх, Анри; Сато, Кацуфуми (2020). «Парящие стили вымерших гигантских птиц и птерозавров» . Biorxiv . doi : 10.1101/2020.10.31.354605 . S2CID 226263538 .
- ^ Unwin, David M. (2006). Птерозавры: из глубокого времени . Нью -Йорк: Pi Press. С. 210–222. ISBN 978-0-13-146308-0 .
- ^ Томкинс, JL; Лебас, NR; Виттон, депутат; Martill, DM; Humphries, S. (2010). «Положительная аллометрия и предыстория сексуального отбора» (PDF) . Американский натуралист . 176 (2): 141–148. doi : 10.1086/653001 . PMID 20565262 . S2CID 36207 . Архивировано из оригинала (PDF) 2017-08-09 . Получено 2018-08-06 .
- ^ Jump up to: а беременный Итон, GF (1910). «Остеология Птеранодона ». Мемуары Академии искусств и наук Коннектикута , 2 : 1–38, пл. I - XXXI.
- ^ Крипп, Д. (1943). «Жизненный образ Птерододон Иннгененс на основе полета». Nova Acta Leopoldina, NF , 12 (83): 16–32 [на немецком языке].
- ^ Stein, RS (1975). «Динамический анализ птерододонских инженеров : адаптация рептилий к полету». Журнал палеонтологии . 49 : 534–548.
- ^ Bramwell, CD и Whitfield, GR (1974). «Биомеханика Птерододона». Философские транзакции Королевское общество B , 267 .
- ^ Jump up to: а беременный Carpenter, K (2003). «Биостратиграфия позвоночных комого мела дымного холма (формация Нибрара) и член Шарон -Спрингс (Пьер Слайл)». Подходы с высоким разрешением в стратиграфической палеонтологии . Темы в геобиологии. 21 : 421–437. doi : 10.1007/978-1-4020-9053-0 . ISBN 978-1-4020-1443-7 .
- ^ Браун, Б. (1904). «Стоучи желудка и пища плезиооров» . Наука . 20 (501): 184–185. Bibcode : 1904sci .... 20..184b . doi : 10.1126/science.20.501.184 . PMID 17737868 .
- ^ Everhart, MJ; Ewell, K. (2006). «Связанные с акулами динозавры (Hadrosauridae) позвонки из мела Нибрара (верхний конец) западного Канзаса». Труды Канзасской академии науки . 109 (1–2): 27–35. doi : 10.1660/0022-8443 (2006) 109 [27: sdhcvf] 2.0.co; 2 . S2CID 86366930 .
- ^ Zimmerman, H., Preiss, B. и Sovak, J. (2001). Помимо динозавров!: Небесные Драконы, Морские монстры, мегамические и другие доисторические звери , Саймон и Шустер. ISBN 0-689-84113-2 .
- ^ Андрес, Б.; Майерс, Т.С. (2013). "Одинокая звезда птерозавров". Земля и экологическая наука Сделок Королевского общества Эдинбурга . 103 (3–4): 383–398. doi : 10.1017/s1755691013000303 . S2CID 84617119 .
- ^ Пегас, Родриго В. (2024-06-10). «Таксономическая заметка о птерозаврах ленты с участка кладбища птерозавров (группа Caiuá, раннее мелодиозное ягодило южного Бразилии): свидетельство наличия двух видов» . Историческая биология : 1–22. doi : 10.1080/08912963.2024.23555664 . ISSN 0891-2963 .
- ^ Jump up to: а беременный Wellnhofer, Peter (1996) [1991]. Иллюстрированная энциклопедия птерозавров . Нью -Йорк: Барнс и Благородные Книги. п. 139. ISBN 978-0-7607-0154-6 .
Дальнейшее чтение
[ редактировать ]- Анонимный. 1872. На двух новых орнитозаврянах из Канзаса. Американский журнал науки , серия 3, 3 (17): 374–375. (Вероятно, OC Marsh)
- Беннетт, SC 2000. Новая информация о скелетах Nyctosaurus . Журнал палеонтологии позвоночных 20 (дополнение к номеру 3): 29а. (Абстрактный)
- Беннетт, Южная Каролина (2001). «Остеология и функциональная морфология позднего мела птерозавра птерододон . Часть II. Функциональная морфология». Palaeontographica, Abteilung a . 260 : 113–153. doi : 10.1127/pala/260/2001/113 . S2CID 210463400 .
- Беннетт, Южная Каролина (2003). «Новые хриплые образцы позднего критока -птерозавра Nyctosaurus ». Палеонтологический журнал . 77 : 61–75. Doi : 10.1007/bf03004560 . S2CID 129438441 .
- Беннетт, Южная Каролина (2007). «Артикуляция и функция птероидной кости птерозавров» (PDF) . Журнал палеонтологии позвоночных . 27 (4): 881–891. doi : 10.1671/0272-4634 (2007) 27 [881: aafotp] 2.0.co; 2 . S2CID 86326537 .
- Беттс, CW 1871. Экспедиция Йельского колледжа 1870 года. Новый ежемесячный журнал Harper's, 43 (257): 663–671. (Выпуск октября 1871 года)
- Боннер, OW 1964. Остеологическое исследование Nyctosaurus и TrinacRomerum с описанием нового вида Nyctosaurus . Невозмутимый Тезис Мастерс, Университет штата Форт -Хейс, 63 страницы.
- Brower, JC (1983). «Аэродинамика Птеронодона и Нью -Йорка , двух крупных птерозавров из верхнего мела Канзаса». Журнал палеонтологии позвоночных . 3 (2): 84–124. doi : 10.1080/02724634.1983.10011963 .
- Коуп, изд 1872 года. О геологии и палеонтологии меловых слоев Канзаса. Годовой отчет Геологической службы США территорий 5: 318–349 (отчет за 1871 год).
- Коуп, изд (1872). «На двух новых орнитозаврах из Канзаса». Труды Американского философского общества . 12 (88): 420–422.
- Коуп, изд (1874). «Обзор позвонков мелового периода, найденный к западу от реки Миссисипи». Геологическая служба США Бюллетена территорий . 1 (2): 3–48.
- Итон, GF 1903. Персонажи Pteranodon . Американский журнал науки , Ser. 4, 16 (91): 82–86, пл. 6-7.
- Итон, GF 1904. Персонажи Pteranodon (вторая статья). Американский журнал науки , Ser. 4, 17 (100): 318–320, пл. 19-20.
- Мы, GF (1908). "Скрауон Птеронона " Наука XXVII : 254–2
- Everhart, MJ 1999. Раннее появление Птерододонского Стернберги из члена Смоки -Хилла (позднего мела) Ниобрара Мел в Западном Канзасе. Сделки Канзасской академии науки 18 (тезисы): 27.
- Everhart, MJ 2005. Oceans of Kansas - естественная история западного внутреннего моря. Издательство Университета Индианы, 320 стр.
- Harksen, JC (1966). « Птеронодон Стернберги , новый ископаемый птеродактиль из ниобрара мела Канзаса». Труды Южной Дакоты Академия науки . 45 : 74–77.
- Крипп, Д. Фон. (1943). «Жизненный образ Птерододон Иннгененс на основе полета». Новая Акта Леопольтина . NF 12 (83): 16–32.
- Лейн, HH 1946. Обзор ископаемых позвоночных Канзаса, Часть III, рептилии, Канзасская академия наука, сделки 49 (3): 289–332, рис. 1–7.
- Марш, OC 1871. Научная экспедиция в Скалистые горы. Американский журнал науки Ser. 3, 1 (6): 142–143.
- Marsh, OC 1871. Уведомление о некоторых новых ископаемых рептилиях из меловых и третичных образов. Американский журнал науки , серия 3, 1 (6): 447–459.
- Marsh, OC 1871. Обратите внимание на новый и гигантский вид pterodactyle. Американский журнал науки , серия 3, 1 (6): 472.
- Marsh, OC 1872. Открытие дополнительных остатков Pterosauria, с описаниями двух новых видов. Американский журнал науки , серия 3, 3 (16): 241–248.
- Marsh, OC 1881. Примечание об американских птеродактиле. Американский журнал науки , серия 3, 21 (124): 342–343.
- Марш, OC 1882. Крылья Pterodactyles. Американский журнал науки , серия 3, 23 (136): 251–256, пл. Iii.
- Marsh, OC 1884. Основные персонажи американских меловых птеродактилов. Часть I. Череп Птеранодона. Американский журнал науки , серия 3, 27 (161): 422–426, пл. 15
- Миллер, HW (1971). «Таксономия видов Птерододона из Канзаса». Труды Канзасской академии науки . 74 (1): 1–19. doi : 10.2307/3627663 . JSTOR 3627663 .
- Миллер, HW (1971). «Череп болота Pteranodon ( Longicepia ) Longiceps , связанный с крылом и частями тела». Труды Канзасской академии науки . 74 (10): 20–33. doi : 10.2307/3627664 . JSTOR 3627664 .
- Падиан, К (1983). «Функциональный анализ полетов и ходьбы в птерозаврах». Палеобиология . 9 (3): 218–239. doi : 10.1017/s009483730000765x . S2CID 88434056 .
- Рассел, DA 1988. Специальный список морских меловых позвоночных в Северной Америке, включая рыб с пресной водой, случайная бумага музея палеонтологии Тиррелла, (4): 57.
- Schultze, H.-P., L. Hunt, J. Chorn and Am Neuner, 1985. Тип и фигурные образцы ископаемых позвоночных в коллекции Музея естественной истории Университета Канзас, часть II. Ископаемые амфибии и рептилии. Разное публикации Музея естественной истории Университета Канзаса 77:66 с.
- Сили, Гарри Г. 1871. Дополнительные доказательства структуры головы у орнитозавров из Кембриджского верхнего зелени; быть дополнением к «Ornithosauria». Анналы и журнал естественной истории , серия 4, 7: 20–36, пожалуйста. 2–3. (Открытие беззубых птерозавров в Англии)
- Шор, EN 1971. окаменелости и мух; Жизнь ученого -комплекса - Сэмюэль Уэнделл Уиллистон, 1851–1918, Университет Оклахома Пресс, 285 стр.
- Sternberg, CH 1990. Жизнь ископаемого охотника, издательство Университета Индианы, 286 с. (Первоначально опубликовано в 1909 году Генри Холтом и Компанией)
- Стернберг, GF; Уокер, MV (1958). «Наблюдение за сочлененными костями конечностей недавно обнаруженного птеродона в ниобрара -меловом меловом виде Канзаса». Труды Канзасской академии науки . 61 (1): 81–85. doi : 10.2307/3626742 . JSTOR 3626742 .
- Стюарт, JD 1990. Ниобрара формация позвоночных стратиграфии. С. 19–30 в Беннетте, Южная Каролина (ред.), НИОБРАРА ЧЕЛОВОЧЕСКИЙ ЭКСПУРИОНАЛЬНЫЙ РУКОВОДСТВО, Музей естественной истории Университета Канзаса и Геологическая служба Канзаса.
- Ван, х.; Zhou, Z. (2004). «Эмбрион птерозавра с раннего мела» . Природа . 429 (6992): 621. Bibcode : 2004natur.429..621W . doi : 10.1038/429621a . PMID 15190343 . S2CID 4428545 .
- Wellnhofer, P. 1991. Иллюстрированная энциклопедия птерозавров. Полумесяц, Нью -Йорк, 192 стр.
- Уиллистон, SW (1891). «Череп и задние конечности Птерододона ». Американский натуралист . 25 (300): 1124–1126. doi : 10.1086/275456 .
- Уиллистон, SW 1892. Канзас Птеродактилс. Часть I. Канзасский университет Ежеквартально 1: 1–13, пл. я.
- Уиллистон, SW 1893. Канзас Птеродактильс. Часть II. Канзасский университет Ежеквартально 2: 79–81, с 1 рис.
- Уиллистон, SW (1895). «Примечание о нижней челюсти Орнитостомы ». Канзасский университет ежеквартально . 4 : 61.
- Уиллистон, SW 1896. На черепе Ornithostoma . Канзасский университет ежеквартально 4 (4): 195–197, с пл. я.
- Уиллистон, SW 1897. Восстановление Ornithostoma ( Pteranodon ) " Канзасский университет Ежеквартально 6: 35–51, с Pl. II.
- Уиллистон, SW (1902). «На скелете Nyctodactylus , с реставрацией» . Американский журнал анатомии . 1 (3): 297–305. doi : 10.1002/aja.1000010306 .
- Williston, SW 1902. На черепе Nyctodactylus , верхний меховой птеродактил. Журнал геологии 10: 520–531, 2 пл.
- Уиллистон, SW 1902. Крылатые рептилии. Поп Science Monthly 60: 314–322, 2 рис.
- Уиллистон, SW 1903. О остеологии Nyctosaurus ( Nyctodactylus ), с заметками об американских птерозаврах. Полевой мус. Издательство (Геологический Сер.) 2 (3): 125–163, 2 Рис., Пл. Xl-xliv.
- Уиллистон, SW 1904. Пальцы птеродактилов. Геологический журнал , серия 5, 1: 59–60.
- Уиллистон, SW (1911). «Крылоки птеродактилов, с восстановлением Nyctosaurus » . Журнал геологии . 19 (8): 696–705. Bibcode : 1911jg ..... 19..696W . doi : 10.1086/621914 .
- Уиллистон, SW (1912). «Обзор GB Eaton's« остеология птеродона ». Журнал геологии . 20 (3): 288. Bibcode : 1912jg ..... 20..288e . doi : 10.1086/621967 .












