Мутационные сигнатуры
Мутационные сигнатуры представляют собой характерные комбинации типов мутаций, возникающие в результате определенных процессов мутагенеза , таких как нарушение репликации ДНК , воздействие экзогенных и эндогенных генотоксинов , дефектные пути репарации ДНК и ферментативное редактирование ДНК. [ 1 ]
Этот термин используется для двух разных понятий, часто смешиваемых: сигнатуры мутагена и сигнатуры опухолей. Его первоначальное использование, мутагенная сигнатура, относится к набору мутаций, произведенных в лаборатории известным мутагеном и не вызванных другими мутагенами, что уникально для мутагена, так же как человеческая сигнатура уникальна для подписавшего. Уникальность позволяет вывести мутаген из мутаций клетки. [ 2 ] Позже эта фраза стала обозначать структуру мутаций, характерную для типа опухоли, хотя обычно она не уникальна ни для типа опухоли, ни для мутагена. [ 3 ] [ 4 ] Если мутационная сигнатура опухоли совпадает с уникальной мутационной сигнатурой мутагена, это позволяет сделать вывод о воздействии канцерогена или процессе мутагенеза, который произошел в далеком прошлом пациента. [ 2 ] Все более уточненные сигнатуры опухолей начинают ассоциироваться с сигнатурами мутагенов. [ 5 ]
Расшифровка мутационных признаков рака дает представление о биологических механизмах, участвующих в канцерогенезе и нормальном соматическом мутагенезе . [ 6 ] Мутационные сигнатуры показали свою применимость в лечении и профилактике рака. Достижения в области онкогеномики позволили разработать и использовать молекулярно- таргетную терапию , но исторически такая терапия была сосредоточена на ингибировании онкогенных факторов (например, EGFR мутации с целью увеличения функции и лечения ингибиторами EGFR при колоректальном раке). [ 7 ] ). Совсем недавно профилирование мутационных сигнатур оказалось успешным в управлении лечением онкологических заболеваний и использовании таргетных методов лечения (например, иммунотерапия при дефектах репарации несоответствий при различных типах рака, [ 8 ] платина и ингибитор PARP для использования синтетической летальности при гомологичной рекомбинации с дефицитом раке молочной железы ). [ 9 ]
Общие понятия
[ редактировать ]
Механизмы – обзор
[ редактировать ]Механизмы биологического мутагенеза, лежащие в основе мутационных сигнатур (например, COSMIC Signatures от 1 до 30), включают, помимо прочего: [ а ] [ 4 ]
- репликации ДНК Неверность
- Корректировка ДНК — это процесс, при котором ДНК-полимераза удаляет неправильно включенный нуклеотид посредством экзонуклеазы ферментативной реакции . Неспособность ДНК-полимеразы исправить эти ошибки репликации приводит к прогрессивному накоплению мутаций посредством последовательного митоза клеток .
- Генотоксины
- Эндогенные клеточные (например, спонтанное 5-метилцитозина дезаминирование приводит к переходу C>T (генетика) ) мутации (см. повреждение ДНК (естественное происхождение) ).
- Экзогенные/ канцерогены
- Ультрафиолетовое излучение: УФ-излучение вызывает прямое повреждение ДНК и является известным фактором риска рака кожи (например, меланомы ).
- Алкилирующие противоопухолевые агенты . Эта группа химиотерапевтических агентов добавляет алкильную группу к ДНК , что вызывает сшивание ДНК и препятствует репликации и восстановлению ДНК . Раковые клетки страдают больше всего из-за высокой скорости митоза .
- Табак : Табак содержит несколько канцерогенов , вредных для ДНК, в том числе полициклические ароматические углеводороды , акролеин , нитрозамины , цианиды и другие (см. Влияние табака на здоровье ).
- Дефицит репарации ДНК
- Дефицит гомологичной рекомбинации (HRD): двухцепочечный разрыв ДНК требует механизма гомологичной рекомбинации для точного восстановления точек разрыва.
- Дефицит восстановления несоответствия ДНК (MMR): механизм восстановления несоответствия распознает и исправляет ошибочную вставку, удаление или неправильное включение пары оснований.
- Ферментативное редактирование ДНК
- Ферменты цитидиндезаминазы: это семейство ферментов является частью врожденной иммунной системы и участвует в контроле ретровирусов и элементов транспозонов (включая эндогенные ретровирусы ). Эти ферменты ( цитидиндезаминаза /CDA, цитидиндезаминаза, индуцированная активацией , и APOBEC семейство белков цитидина ) активно вызывают дезаминирование и, следовательно, вызывают перехода C>T (генетические) . мутации
- репликации ДНК Неверность
Геномные данные
[ редактировать ]Для анализа сигнатур мутаций рака требуются геномные данные секвенирования генома рака с парным нормальным секвенированием ДНК , чтобы создать каталог опухолевых мутаций (типы и количество мутаций) конкретной опухоли. Различные типы мутаций (например, однонуклеотидные варианты, инделы, структурные варианты) можно использовать по отдельности или в комбинации для моделирования мутационных признаков рака.
Виды мутаций: замены оснований
[ редактировать ]Существует шесть классов замены оснований: C>A, C>G, C>T, T>A, T>C, T>G. Замену G>T считают эквивалентной замене C>A, поскольку невозможно различить, на какой цепи ДНК (прямой или обратной) первоначально произошла замена. Таким образом, замены C>A и G>T считаются частью класса «C>A». По той же причине мутации G>C, G>A, A>T, A>G и A>C считаются частью «C>G», «C>T», «T>A», « Классы T>C» и «T>G» соответственно.
Получение информации от соседних 5'- и 3'-оснований (также называемых фланкирующими парами оснований или тринуклеотидным контекстом) приводит к 96 возможным типам мутаций (например, A[C>A]A, A[C>A]T и т. д.). Каталог мутаций опухоли создается путем отнесения каждого однонуклеотидного варианта (SNV) (синонимы: замена пары оснований или точечная мутация замены ) к одному из 96 типов мутаций и подсчета общего количества замен для каждого из этих 96 типов мутаций. (см. рисунок).
Каталог мутаций опухолей
[ редактировать ]
После получения каталога мутаций (например, подсчета для каждого из 96 типов мутаций) опухоли существует два подхода к расшифровке вклада различных мутационных сигнатур в геномный ландшафт опухоли:
- Каталог мутаций опухоли сравнивается с эталонным каталогом мутаций или справочным набором данных сигнатур мутаций, например, с 21 сигнатурой мутационных процессов при раке человека. [ 4 ] из базы данных «Каталога соматических мутаций при раке» ( COSMIC ). [ 1 ]
- Моделирование мутационных сигнатур de novo может быть выполнено с использованием статистических методов, таких как факторизация неотрицательной матрицы, для выявления потенциальных новых мутационных процессов. [ 10 ]
Выявление вклада различных мутационных признаков в канцерогенез дает понимание биологии опухолей и может открыть возможности для таргетной терапии .
Виды мутаций: инделы
[ редактировать ]Сигнатура 3, наблюдаемая в опухоли с дефицитом гомологичной рекомбинации (HR), связана с повышенным содержанием крупных инделей (до 50 нуклеотидов) с перекрывающейся микрогомологией в точках разрыва. [ 4 ] В таких опухолях двухцепочечные разрывы ДНК восстанавливаются с помощью неточных механизмов репарации негомологичного соединения концов (NHEJ) или микрогомологического соединения концов (MMEJ) вместо высокоточной репарации HR.
Сигнатура 6, наблюдаемая в опухолях с микросателлитной нестабильностью , также характеризуется обогащением инделей длиной 1 п.н. в областях нуклеотидных повторов.
Виды мутаций: структурные варианты
[ редактировать ]Дефицит гомологичной рекомбинации приводит к паттерну замещения сигнатуры 3, а также к увеличению нагрузки структурных вариантов. В отсутствие рекомбинации гомологичной негомологичное соединение концов приводит к большим структурным вариантам, таким как хромосомные транслокации , хромосомные инверсии и варианты числа копий .
Мутационные сигнатуры
[ редактировать ]Краткое описание отдельных мутационных процессов и связанных с ними мутационных признаков рака будет включено в разделы ниже. Некоторые признаки повсеместно распространены при различных типах рака (например, признак 1), в то время как некоторые другие имеют тенденцию ассоциироваться с конкретными видами рака (например, признак 9 и лимфоидные злокачественные новообразования) . [ 4 ]
Некоторые мутационные сигнатуры характеризуются сильным смещением транскрипции, при этом замены преимущественно затрагивают одну из цепей ДНК, транскрибируемую или нетранскрибируемую (сигнатуры 5, 7, 8, 10, 12, 16). [ 4 ]
Возрастной мутагенез
[ редактировать ]Сигнатура 1 характеризуется преобладанием перехода C>T (генетика) в тринуклеотидных контекстах Np[C>T]G и коррелирует с возрастом пациента на момент постановки диагноза рака . В основе предлагаемого биологического механизма лежит спонтанное дезаминирование 5-метилцитозина . [ 4 ]
Сигнатура 5 имеет преобладание замен T>C в контексте тринуклеотида ApTpN со смещением цепи транскрипции. [ 6 ]
Дефицит гомологичной рекомбинации
[ редактировать ]Сигнатура 3 демонстрирует большое количество мутаций нескольких классов мутаций и связана с зародышевой и соматическими (биологическими) мутациями BRCA1 и BRCA2 при нескольких типах рака (например, молочной железы, поджелудочной железы, яичников, простаты). Эта сигнатура является результатом дефицита восстановления двухцепочечных разрывов ДНК (или дефицита гомологичной рекомбинации ). Сигнатура 3 связана с высокой нагрузкой инделей с микрогомологией в точках разрыва. [ 6 ]
ферменты АПОБЕК
[ редактировать ]APOBEC3 семейства Ферменты цитидиндезаминазы ферментативная активность APOBEC3A и APOBEC3B реагируют на вирусные инфекции путем редактирования вирусного генома, но также было обнаружено, что вызывает нежелательное редактирование генома хозяина и может даже участвовать в онкогенезе при раке, связанном с вирусом папилломы человека . [ 11 ]
Сигнатура 2 и сигнатура 13 обогащены заменами C>T и C>G и, как полагают, возникают в результате активности цитидиндезаминазы семейства ферментов AID/ APOBEC . [ 6 ]
Делеционный полиморфизм зародышевой линии, включающий APOBEC3A и APOBEC3B, связан с высоким бременем мутаций Signature 2 и Signature 13. [ 12 ] Этот полиморфизм считается умеренной пенетрантностью (в два раза выше фонового риска) для риска рака молочной железы. [ 13 ] Точные роли и механизмы, лежащие в основе редактирования генома, опосредованного APOBEC, еще полностью не определены, но считается, что индуцируемая активацией комплекс цитидиндезаминаза (AID)/ APOBEC участвует в иммунном ответе хозяина на вирусные инфекции и метаболизме липидов. [ 14 ]
И сигнатура 2, и сигнатура 13 характеризуются заменами цитозина на урацил вследствие цитидиндезаминаз. Сигнатура 2 имеет более высокую долю замен C[T>C]N, а сигнатура 13 — более высокую долю замен T[C>G]N. Мутагенез, опосредованный APOBEC3A и APOBEC3B, во время репликации преимущественно затрагивает отстающую цепь ДНК. [ 15 ]
Недостаток восстановления несоответствия
[ редактировать ]Четыре мутационные сигнатуры COSMIC были связаны с дефицитом репарации несоответствия ДНК и обнаружены в опухолях с микросателлитной нестабильностью : сигнатуры 6, 15, 20 и 26. [ 6 ] Потеря функции генов MLH1 , MSH2 , MSH6 или PMS2 вызывает дефектное восстановление несоответствия ДНК .
корректура ДНК
[ редактировать ]Сигнатура 10 имеет транскрипционную предвзятость и обогащена заменами C>A в контексте TpCpT, а также заменами T>G в контексте TpTpTp. [ 6 ] Сигнатура 10 связана с измененной функцией ДНК-полимеразы эпсилон , что приводит к недостаточной корректирующей активности ДНК. Как зародышевые, так и соматические мутации экзонуклеазного домена POLE (гена) связаны с сигнатурой 10. [ 16 ]
Базовый иссеченный ремонт
[ редактировать ]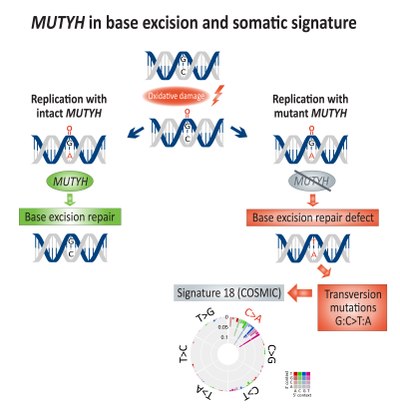
Соматическое обогащение трансверсионными мутациями (G:C>T:A) было связано с дефицитом эксцизионной репарации оснований (BER) и дефектом MUTYH , ДНК-гликозилазы , при колоректальном раке. [ 17 ] Прямое повреждение ДНК от окисления приводит к образованию 8-оксогуанина , который, если его не восстановить, приведет к включению аденина вместо цитозина во время репликации ДНК. MUTYH кодирует фермент аденингликозилазу mutY , который вырезает несовпадающий аденин из 8-оксогуанин : аденин пары оснований репарации ДНК, , тем самым обеспечивая механизмы включающие OGG1 (оксогуанингликозилазу) и NUDT1 (гидролазу 1 Nudix, также известную как MTH1 , гомолог 1 MutT) для удаления поврежденный 8-оксогуанин . [ 18 ]
Воздействие экзогенных генотоксинов
[ редактировать ]Отдельные экзогенные генотоксины/ канцерогены и их мутаген -индуцированные механизмы повреждения и восстановления ДНК связаны со специфическими молекулярными характеристиками.
Ультрафиолетовое излучение (УФ)
[ редактировать ]- Сигнатура 7 имеет преобладание замен C>T в сайтах соседних пиримидинов (соседних C или T), причем особенно диагностической подгруппой является динуклеотидная мутация CC>TT. Эта закономерность возникает потому, что основные фотопродукты ДНК, индуцированные УФ-излучением, соединяются с двумя соседними пиримидинами; фотопродуктом обычно является димер циклобутан-пиримидина (CPD). [ 19 ] Специфичность C>T, по-видимому, обусловлена ускорением дезаминирования C в миллион раз, когда он является частью CPD, в результате чего образующийся урацил действует как T. [ 20 ] [ 21 ] CPD репарируются посредством эксцизионной репарации нуклеотидов , связанной с транскрипцией , что приводит к сильному смещению замен C>T, обогащенных нетранскрибируемой цепью ДНК. [ 6 ] Области белка-супрессора опухоли, которые мутационно инактивируются при раке кожи, связанном с солнечным светом, такие же, как и при раке органов, не подвергающихся воздействию солнечного света, но мутировавший нуклеотид часто смещается на несколько оснований к месту, где может образоваться CPD. [ 22 ] Таким образом, воздействие ультрафиолетового излучения является предполагаемым основным мутагенным механизмом этой сигнатуры. УФ также демонстрирует тонкость интерпретации сигнатуры опухоли как сигнатуры мутагена: только три четверти мутаций, вызванных УФ в лаборатории, являются мутациями сигнатуры УФ, поскольку УФ также запускает клеточные окислительные процессы. [ 2 ] Поэтому, даже если все мутации в опухоли были вызваны УФ-излучением солнечного света, ожидается, что четверть мутаций не будут мутациями, связанными с УФ-сигнатурой. Для объяснения этих мутаций нет необходимости привлекать второй канцероген, но необходим второй мутационный процесс. Идентификация УФ-сигнатуры в опухоли неизвестной первичной локализации клинически важна, поскольку предполагает диагноз метастатического рака кожи и имеет важное значение для лечения. [ 23 ]
Алкилирующие агенты
[ редактировать ]- Сигнатура 11 была идентифицирована в опухолях, ранее подвергавшихся воздействию темозоламида, алкилирующего агента . [ 6 ] Эта сигнатура обогащена заменами C>T на гуаниновых основаниях вследствие транскрипционно-связанной эксцизионной репарации нуклеотидов . В этой сигнатуре присутствует сильная транскрипционная смещение цепи.
Табак
[ редактировать ]- Как сигнатура 4 ( табакокурение , рак легких ), так и сигнатура 29 ( жевание табака десен и щеки полости рта , плоскоклеточная карцинома ) демонстрируют смещение цепи транскрипции и обогащение заменами C>A, но их соответствующий состав и закономерности (доля каждого типа мутаций) ) немного отличаются. [ 6 ]
- Предполагаемый механизм, лежащий в основе сигнатуры 4, заключается в удалении аддуктов ДНК ( табака бензо(а)пирена , ковалентно связанного с гуанином связанного с транскрипцией ) с помощью механизма эксцизионной репарации нуклеотидов, (NER). [ 24 ]
Гипермутация гена иммуноглобулина
[ редактировать ]Сигнатура 9 была идентифицирована при хроническом лимфоцитарном лейкозе и злокачественной B-клеточной лимфоме и обогащена признаками событий трансверсии T>G . Считается, что это результат склонной к ошибкам полимеразы. η ( ген POLH )-ассоциированный мутагенез . [ 4 ]
В последнее время полимераза Сигнатура синтеза, подверженная ошибкам , была связана с негематологическими раковыми заболеваниями (например, раком кожи ), и предполагалось, что она способствует мутагенезу мотива YCG и может частично объяснить увеличение количества замен динуклеотидов TC. [ 25 ]
История
[ редактировать ]В 1990-е годы Кертис Харрис из Национального института рака США и Берт Фогельштейн из Онкологического центра Джонса Хопкинса в Балтиморе проанализировали данные, показывающие, что разные типы рака имеют свой собственный уникальный набор мутаций в р53 , которые, вероятно, были вызваны разными агенты, [ 3 ] [ 26 ] например, химические вещества в табачном дыме или ультрафиолетовый свет солнца. [ 19 ] [ 27 ] С появлением секвенирования следующего поколения увидел Майкл Стрэттон потенциал этой технологии, способной революционизировать наше понимание генетических изменений внутри отдельных опухолей, запустив огромные банки машин для секвенирования ДНК Института Уэллком Сэнгер, которые смогут читать каждую букву. ДНК в опухоли. [ 28 ] К 2009 году Стрэттон и его команда получили первые полные последовательности генома рака. Это были подробные карты, показывающие все генетические изменения и мутации, которые произошли в двух отдельных видах рака — меланоме кожи и опухоли легких. [ 29 ] [ 30 ] Геномы меланомы и рака легких стали убедительным доказательством того, что отпечатки пальцев конкретных виновников можно увидеть в раковых заболеваниях с одной основной причиной. Эти опухоли все еще содержали множество мутаций, которые нельзя было объяснить ультрафиолетом или курением табака. Детективная работа значительно усложнилась в случае рака сложного, множественного или даже совершенно неизвестного происхождения. В качестве аналогии представьте себе судебно-медицинского эксперта, смахивающего пыль с места убийства. Судмедэксперту может повезти, и он обнаружит на оконном стекле или дверной ручке набор идеальных отпечатков, которые соответствуют известному убийце. Однако у них гораздо больше шансов обнаружить мешанину отпечатков пальцев, принадлежащих целому ряду людей — от жертвы и потенциальных подозреваемых до невиновных и следователей полиции — и все они лежат друг на друге на самых разных поверхностях. [ 28 ] Это очень похоже на геномы рака, где множественные мутационные закономерности обычно накладываются друг на друга, что делает данные непонятными. К счастью, аспирант Стрэттона Людмил Александров придумал способ математического решения проблемы. Александров продемонстрировал, что мутационные закономерности отдельных мутагенов, обнаруженных в опухоли, можно отличить друг от друга с помощью математического подхода, называемого слепым разделением источников . Недавно распутанные закономерности мутаций были названы мутационными сигнатурами. [ 28 ] В 2013 году Александров и Стрэттон опубликовали первую вычислительную систему для расшифровки мутационных сигнатур из геномики рака . данных [ 31 ] Впоследствии они применили эту схему к более чем семи тысячам раковых геномов, создав первую полную карту мутационных признаков рака человека. [ 32 ] В настоящее время выявлено более ста мутационных признаков рака человека. [ 33 ] В апреле 2022 года было описано 58 новых мутационных сигнатур. [ 34 ] [ 35 ] [ 36 ]
См. также
[ редактировать ]Список заметок
[ редактировать ]- ^ Поскольку репликация, поддержание и восстановление ДНК не являются линейным процессом, некоторые признаки вызваны перекрывающимися механизмами мутагенеза.
Ссылки
[ редактировать ]- ^ Перейти обратно: а б Форбс С.А., Беар Д., Бутселакис Х., Бэмфорд С., Биндал Н., Тейт Дж. и др. (январь 2017 г.). «КОСМИЧЕСКАЯ: генетика соматического рака в высоком разрешении» . Исследования нуклеиновых кислот . 45 (Д1): Д777–Д783. дои : 10.1093/nar/gkw1121 . ПМК 5210583 . ПМИД 27899578 .
- ^ Перейти обратно: а б с Дерзкий Д.Э. (2015). «Мутации УФ-сигнатуры» . Фотохимия и фотобиология . 91 (1): 15–26. дои : 10.1111/php.12377 . ПМЦ 4294947 . ПМИД 25354245 .
- ^ Перейти обратно: а б Холлштейн М., Сидрански Д., Фогельштейн Б., Харрис К.С. (июль 1991 г.). «Мутации р53 при раке человека» . Наука . 253 (5015): 49–53. Бибкод : 1991Sci...253...49H . дои : 10.1126/science.1905840 . ПМИД 1905840 . S2CID 38527914 .
- ^ Перейти обратно: а б с д и ж г час я дж Александров Л.Б., Ник-Зайнал С., Ведж Д.С., Апарисио С.А., Бехжати С., Бьянкин А.В. и др. (август 2013 г.). «Признаки мутационных процессов при раке человека» (PDF) . Природа . 500 (7463): 415–21. Бибкод : 2013Natur.500..415. . дои : 10.1038/nature12477 . ПМК 3776390 . ПМИД 23945592 .
- ^ Кукаб Дж.Э., Зоу Х, Морганелла С., Джоэл М., Нанда А.С., Надь Э., Гомес С., Дегаспери А., Харрис Р., Джексон С.П., Арлт В.М., Филлипс Д.Х., Ник-Зайнал С. (2019). «Сборник мутационных сигнатур агентов окружающей среды» . Клетка . 177 (4): 821–36 e16. дои : 10.1016/j.cell.2019.03.001 . ПМК 6506336 . ПМИД 30982602 .
- ^ Перейти обратно: а б с д и ж г час я Александров Л.Б., Джонс П.Х., Ведж Д.С., Сэйл Дж.Е., Кэмпбелл П.Дж., Ник-Зайнал С., Страттон М.Р. (декабрь 2015 г.). «Часообразные мутационные процессы в соматических клетках человека» . Природная генетика . 47 (12): 1402–7. дои : 10.1038/ng.3441 . ПМЦ 4783858 . ПМИД 26551669 .
- ^ Соу Х., Йип В.К., Фифис Т. (март 2016 г.). «Достижения в области таргетной и иммунологической терапии колоректального рака в эпоху геномики» . Онкомишени и терапия . 9 (9): 1899–920. дои : 10.2147/OTT.S95101 . ПМЦ 4821380 . ПМИД 27099521 .
- ^ Чук М.К., Чанг Дж.Т., Теорет М.Р., Сампене Е., Хе К., Вейс С.Л., Хелмс В.С., Джин Р., Ли Х., Ю Дж., Чжао Х., Чжао Л., Пачига М., Шмиэль Д., Рават Р., Киган П., Паздур Р. (октябрь 2017 г.). «Сводка одобрения FDA: ускоренное одобрение пембролизумаба для лечения второй линии метастатической меланомы» . Клинические исследования рака . 23 (19): 5666–5670. дои : 10.1158/1078-0432.CCR-16-0663 . ПМИД 28235882 .
- ^ О'Нил, Найджел Дж.; Бейли, Мелани Л.; Хитер, Филип (26 июня 2017 г.). «Синтетическая летальность и рак». Обзоры природы Генетика . 18 (10): 613–623. дои : 10.1038/nrg.2017.47 . ПМИД 28649135 . S2CID 3422717 .
- ^ Перейти обратно: а б Чжао Э.Ю., Шен Ю., Плезанс Э., Касаян К., Лилакумари С., Джонс М. и др. (декабрь 2017 г.). «Дефицит гомологичной рекомбинации и результаты терапии на основе платины при распространенном раке молочной железы» . Клинические исследования рака . 23 (24): 7521–7530. дои : 10.1158/1078-0432.CCR-17-1941 . ПМИД 29246904 .
- ^ Уоррен С., Вестрик Дж., Дорслер К., Пён Д. (август 2017 г.). «Роль APOBEC3A и APOBEC3B в папилломавирусной инфекции и прогрессировании заболевания» . Вирусы . 9 (8): 233. дои : 10.3390/v9080233 . ПМК 5580490 . ПМИД 28825669 .
- ^ Миддлбрукс С.Д., Бандай А.Р., Мацуда К., Удким К.И., Онабаджо О.О., Пакин А. и др. (ноябрь 2016 г.). «Связь вариантов зародышевой линии в регионе APOBEC3 с риском рака и обогащением сигнатурных мутаций APOBEC в опухолях» . Природная генетика . 48 (11): 1330–1338. дои : 10.1038/ng.3670 . ПМК 6583788 . ПМИД 27643540 .
- ^ Ник-Зайнал С., Ведж Д.С., Александров Л.Б., Петляк М., Батлер А.П., Болли Н. и др. (май 2014 г.). «Связь полиморфизма числа копий зародышевой линии APOBEC3A и APOBEC3B с бременем предполагаемых APOBEC-зависимых мутаций при раке молочной железы» . Природная генетика . 46 (5): 487–91. дои : 10.1038/ng.2955 . ПМЦ 4137149 . ПМИД 24728294 .
- ^ Ян Б., Ли Х, Лей Л., Чен Дж. (сентябрь 2017 г.). «APOBEC: От мутатора к редактору». Журнал генетики и геномики = И Чуань Сюэ Бао . 44 (9): 423–437. дои : 10.1016/j.jgg.2017.04.009 . ПМИД 28964683 .
- ^ Хупс Дж.И., Кортес Л.М., Мерц Т.М., Малк Э.П., Мечковски П.А., Робертс С.А. (февраль 2016 г.). «APOBEC3A и APOBEC3B преимущественно дезаминируют шаблон отстающей цепи во время репликации ДНК» . Отчеты по ячейкам . 14 (6): 1273–1282. дои : 10.1016/j.celrep.2016.01.021 . ПМЦ 4758883 . ПМИД 26832400 .
- ^ Рейнер Э., Ван Гул И.С., Паллес С., Кирси С.Э., Босс Т., Томлинсон И., Черч Д.Н. (февраль 2016 г.). «Набор ошибок: мутации домена корректуры полимеразы при раке» . Обзоры природы. Рак . 16 (2): 71–81. дои : 10.1038/nrc.2015.12 . ПМИД 26822575 . S2CID 9359891 .
- ^ Перейти обратно: а б Виль А., Брюсель А., Мечча Е. и др. (апрель 2017 г.). «Специфическая мутационная сигнатура, связанная с персистенцией ДНК 8-оксогуанина при MUTYH-дефектном колоректальном раке» . Электронная биомедицина . 20 : 39–49. дои : 10.1016/j.ebiom.2017.04.022 . ПМЦ 5478212 . ПМИД 28551381 .
- ^ Дэвид, СС ; О'Ши, В.Л.; Кунду, С (2007). «Базово-эксцизионное восстановление окислительных повреждений ДНК» . Природа . 447 (7147): 941–950. Бибкод : 2007Natur.447..941D . дои : 10.1038/nature05978 . ПМЦ 2896554 . ПМИД 17581577 .
- ^ Перейти обратно: а б Браш Д.Э., Рудольф Дж.А., Саймон Дж.А., Лин А., Маккенна Г.Дж., Баден Х.П., Гальперин А.Дж., Понтен Дж. (1991). «Роль солнечного света в раке кожи: УФ-индуцированные мутации р53 при плоскоклеточном раке» . Труды Национальной академии наук США . 88 (22): 10124–8. Бибкод : 1991PNAS...8810124B . дои : 10.1073/pnas.88.22.10124 . ПМК 52880 . ПМИД 1946433 .
- ^ Каннистраро В.Дж., Пондугула С., Сонг К., Тейлор Дж.С. (2015). «Быстрое дезаминирование фотопродуктов циклобутан-пиримидинового димера в сайтах TCG в трансляционно и вращательно расположенной нуклеосоме in vivo» . J Биол Хим . 290 (44): 26597–26609. дои : 10.1074/jbc.M115.673301 . ПМЦ 4646317 . ПМИД 26354431 .
- ^ Джин С.Г., Петтинга Д., Джонсон Дж., Ли П., Пфайфер Г.П. (2021). «Основной механизм мутаций меланомы основан на дезаминировании цитозина в пиримидиновых димерах, что определяется с помощью кругового секвенирования повреждений» . Достижения науки . 7 (31). Бибкод : 2021SciA....7.6508J . дои : 10.1126/sciadv.abi6508 . ПМЦ 8324051 . ПМИД 34330711 .
- ^ Зиглер А., Леффелл Д.Д., Кунала С., Шарма Х.В., Гайлани М., Саймон Дж.А., Гальперин А.Дж., Баден Х.П., Шапиро П.Е., Бэйл А.Е., Браш Д.Е. (1993). «Горячие точки мутаций из-за солнечного света в гене p53 немеланомного рака кожи» . Труды Национальной академии наук США . 90 (9): 4216–20. Бибкод : 1993PNAS...90.4216Z . дои : 10.1073/pnas.90.9.4216 . ПМК 46477 . ПМИД 8483937 .
- ^ Мата, Дуглас А.; Уильямс, Эрик А.; Сокол, Итан; Окснард, Джеффри Р.; Флейшманн, Зоя; Це, Джули Ю.; Декер, Бреннан (23 марта 2022 г.). «Распространенность УФ-мутационных сигнатур среди первичных опухолей кожи» . Открытая сеть JAMA . 5 (3): e223833. doi : 10.1001/jamanetworkopen.2022.3833 . ПМЦ 8943639 . ПМИД 35319765 .
- ^ Александров Л.Б., Джу Ю.С., Хаасе К., Ван Лоо П., Мартинкорена И., Ник-Зайнал С., Тотоки Ю., Фудзимото А., Накагава Х., Сибата Т., Кэмпбелл П.Дж., Винейс П., Филлипс Д.Х., Страттон М.Р. (ноябрь 2016 г.). «Мутационные сигнатуры, связанные с курением табака при раке человека» . Наука 354 (6312): 618–622. Бибкод : 2016Sci...354..618A . дои : 10.1126/science.aag0299 . ПМК 6141049 . ПМИД 27811275 .
- ^ Рогозин И.Б., Гончаренко А., Лада А.Г., Де С., Юрченкод В., Нудельман Г., Панченко А.Р., Купер Д.Н., Павлов Ю.И. (февраль 2018 г.). «Мутационные сигнатуры ДНК-полимеразы η обнаруживаются при различных типах рака» . Клеточный цикл . 17 (3): 348–355. дои : 10.1080/15384101.2017.1404208 . ПМЦ 5914734 . ПМИД 29139326 .
- ^ Оливье М., Хуссейн С.П., Карон де Фромантель С., Эно П., Харрис CC (2004). «Спектры мутаций TP53 и нагрузка: инструмент для генерации гипотез об этиологии рака». Научные публикации МАИР (157): 247–70. ПМИД 15055300 .
- ^ Пфайфер ГП, Эно П (2003). «О происхождении G --> T-трансверсий при раке легких». Мутационные исследования . 526 (1–2): 39–43. дои : 10.1016/s0027-5107(03)00013-7 . ПМИД 12714181 .
- ^ Перейти обратно: а б с Мозаика, Кэт Арни. «Детективы ДНК, которые ищут причины рака» . CNN . Проверено 25 сентября 2018 г.
- ^ Pleasance ED, Cheetham RK, Stephens PJ, McBride DJ, Humphray SJ, Greenman CD и др. (январь 2010 г.). «Полный каталог соматических мутаций генома рака человека» . Природа . 463 (7278): 191–6. Бибкод : 2010Natur.463..191P . дои : 10.1038/nature08658 . ПМК 3145108 . ПМИД 20016485 .
- ^ Pleasance ED, Stephens PJ, O'Meara S, McBride DJ, Meynert A, Jones D и др. (январь 2010 г.). «Геном мелкоклеточного рака легких со сложными признаками воздействия табака» . Природа . 463 (7278): 184–90. Бибкод : 2010Natur.463..184P . дои : 10.1038/nature08629 . ПМЦ 2880489 . ПМИД 20016488 .
- ^ Александров Л.Б., Ник-Зайнал С., Ведж Д.С., Кэмпбелл П.Дж., Страттон М.Р. (январь 2013 г.). «Расшифровка признаков мутационных процессов, происходящих при раке человека» . Отчеты по ячейкам . 3 (1): 246–59. дои : 10.1016/j.celrep.2012.12.008 . ПМЦ 3588146 . ПМИД 23318258 .
- ^ Александров Л.Б., Ник-Зайнал С., Ведж Д.С., Апарисио С.А., Бехжати С., Бьянкин А.В. и др. (август 2013 г.). «Признаки мутационных процессов при раке человека» . Природа . 500 (7463): 415–21. Бибкод : 2013Natur.500..415. . дои : 10.1038/nature12477 . ПМК 3776390 . ПМИД 23945592 .
- ^ Александров Л., Ким Дж., Харадхвала Н.Дж., Хуанг М.Н., Нг А.В., Бут А., Ковингтон К.Р., Горденин Д.А., Бергстрем Е (15 мая 2018 г.). «Репертуар мутационных сигнатур при раке человека». bioRxiv 10.1101/322859 .
- ^ Дегаспери, Андреа; и др. (21 апреля 2021 г.). «Признаки мутационных замен при полногеномном секвенировании рака у населения Великобритании» . Наука . 376 (6591). дои : 10.1126/science.abl9283 . ПМЦ 7613262 . ПМИД 35949260 .
- ^ Ледфорд, Хайди (21 апреля 2022 г.). «Находка опухолевых геномов дает ключ к разгадке происхождения рака» . Природа . 604 (7907): 609. Бибкод : 2022Natur.604..609L . дои : 10.1038/d41586-022-01095-2 . ПМИД 35449305 . S2CID 248323597 .
- ^ «Рак: огромный анализ ДНК открывает новые подсказки» . Новости Би-би-си . 21 апреля 2022 г. Проверено 22 апреля 2022 г.