Онкогеномика
Онкогеномика — это раздел геномики , который характеризует раком связанные с гены, . Основное внимание уделяется геномным, эпигеномным изменениям и изменениям транскриптов при раке.
Рак — генетическое заболевание, вызванное накоплением мутаций ДНК и эпигенетических изменений, приводящих к неограниченной пролиферации клеток и образованию новообразований . Цель онкогеномики — идентифицировать новые онкогены или гены-супрессоры опухолей , которые могут дать новое представление о диагностике рака, прогнозировании клинических исходов рака и новых мишенях для лечения рака. Успех таргетных методов лечения рака, таких как Гливек , Герцептин и Авастин, породил надежду на то, что онкогеномика позволит выявить новые цели лечения рака. [1]

Помимо понимания основных генетических механизмов, которые инициируют или способствуют прогрессированию рака, онкогеномика нацелена на персонализированное лечение рака. Рак развивается из-за мутаций ДНК и эпигенетических изменений, которые накапливаются случайным образом. Выявление и нацеливание на мутации у отдельного пациента может привести к повышению эффективности лечения.
Завершение проекта «Геном человека» облегчило развитие онкогеномики и расширило возможности исследователей по поиску онкогенов. Технологии секвенирования и методы глобального профилирования метилирования применялись для изучения онкогеномики.
История
[ редактировать ]Эра геномики началась в 1990-х годах, когда были созданы последовательности ДНК многих организмов. В 21 веке завершение проекта «Геном человека» позволило изучить функциональную геномику и изучить геномы опухолей. Рак находится в центре внимания.
Эпоха эпигеномики началась сравнительно недавно, примерно в 2000 году. [2] [3] Одним из основных источников эпигенетических изменений является изменение метилирования CpG-островков в промоторной области генов (см. Метилирование ДНК при раке ). Ряд недавно разработанных методов позволяют оценить статус метилирования ДНК при раке по сравнению с нормальными тканями. [4] Некоторые методы оценивают метилирование CpG, расположенных в разных классах локусов, включая CpG-островки, берега и полки, а также промоторы, тела генов и межгенные области. [5] Рак также является основным объектом эпигенетических исследований.
Доступ к секвенированию всего генома рака важен для исследований рака (или генома рака), потому что:
- Мутации являются непосредственной причиной рака и определяют фенотип опухоли .
- Доступ к образцам раковых и нормальных тканей одного и того же пациента, а также тот факт, что большинство раковых мутаций представляют собой соматические события, позволяют идентифицировать мутации, специфичные для рака.
- Мутации рака кумулятивны и иногда связаны со стадией заболевания. метастазы и лекарственную устойчивость. Различают [6]
Доступ к профилированию метилирования важен для исследований рака, потому что:
- Эпи-драйверы, наряду с Мут-драйверами, могут выступать в качестве непосредственных причин рака. [7]
- Эпимутации рака носят кумулятивный характер и иногда связаны со стадией заболевания. [8]
Полногеномное секвенирование
[ редактировать ]Первый геном рака был секвенирован в 2008 году. [6] В этом исследовании секвенировали типичный геном острого миелолейкоза (ОМЛ) и его нормальный геном-аналог, полученный от одного и того же пациента. Сравнение выявило десять мутировавших генов. Уже считалось, что два из них способствуют прогрессированию опухоли: внутренняя тандемная дупликация FLT3 рецептора гена тирозинкиназы , который активирует передачу сигналов киназы и связана с плохим прогнозом, и вставка четырех оснований в экзоне 12 гена NPM1 (NPMc). Эти мутации обнаруживаются в 25–30% опухолей ОМЛ и, как полагают, способствуют прогрессированию заболевания, а не вызывают его напрямую.
Остальные 8 были новыми мутациями, и все они представляли собой изменения с одним основанием: четыре были в семьях, которые тесно связаны с патогенезом рака ( PTPRT , CDH24, PCLKC и SLC15A1 ). Остальные четыре ранее не имели связи с патогенезом рака. У них действительно были потенциальные функции в метаболических путях , которые предполагали механизмы, с помощью которых они могли способствовать развитию рака (KNDC1, GPR124 , EB12, GRINC1B).
Эти гены участвуют в путях, которые, как известно, способствуют патогенезу рака, но до этого исследования большинство из них не были кандидатами на таргетную генную терапию. Этот анализ подтвердил подход секвенирования всего ракового генома при выявлении соматических мутаций и важность параллельного секвенирования геномов нормальных и опухолевых клеток. [9]
В 2011 году геном исключительного пациента с раком мочевого пузыря, опухоль которого была устранена с помощью препарата эверолимус, был секвенирован, что выявило мутации в двух генах: TSC1 и NF2 . Мутации нарушили регуляцию mTOR , белка, ингибируемого эверолимусом, что позволило ему воспроизводиться без ограничений. В результате в 2015 году в Национальном институте рака была создана Инициатива исключительных ответчиков. Инициатива позволяет таким исключительным пациентам (которые положительно реагировали в течение как минимум шести месяцев на лекарство от рака, которое обычно не помогает) секвенировать их геномы для выявления соответствующих мутаций. После выявления этих мутаций другие пациенты могут быть проверены на наличие этих мутаций, а затем им назначен препарат. В 2016 году с этой целью в 2015 году начались общенациональные испытания лекарства от рака, в которых приняли участие до двадцати четырехсот центров. Пациентам с соответствующими мутациями подбирают один из более чем сорока препаратов. [10]
В 2014 году Центр молекулярной онкологии представил тест MSK-IMPACT — инструмент скрининга, который ищет мутации в 341 гене, связанном с раком. К 2015 году было обследовано более пяти тысяч пациентов. Пациенты с соответствующими мутациями имеют право участвовать в клинических исследованиях, обеспечивающих таргетную терапию. [10]
Технологии
[ редактировать ]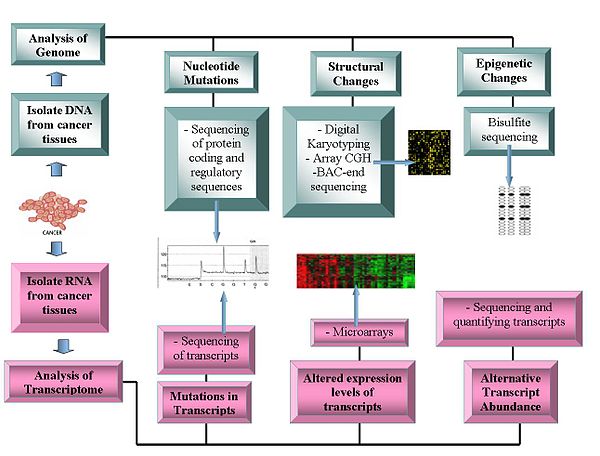
Геномные технологии включают в себя:
Секвенирование генома
[ редактировать ]- Секвенирование ДНК . Секвенаторы на основе пиросеквенирования предлагают относительно недорогой метод получения данных о последовательностях. [1] [11] [12]
- Сравнительная гибридизация генома с помощью массива : этот метод измеряет разницу в количестве копий ДНК между нормальным и раковым геномами. Он использует интенсивность флуоресценции флуоресцентно-меченных образцов, которые гибридизуются с известными зондами на микрочипе. [13] [14]
- Репрезентативный олигонуклеотидных микрочипов анализ : обнаруживает изменение числа копий с использованием амплифицированных рестрикционно расщепленных геномных фрагментов, которые гибридизуются с человеческими олигонуклеотидами, достигая разрешения от 30 до 35 кбит/с. [15]
- Цифровое кариотипирование : обнаруживает изменение количества копий с использованием геномных меток, полученных с помощью переваривания ферментами рестрикции . Эти теги затем связываются в дитаги, конкатенируются, клонируются, секвенируются и снова сопоставляются с эталонным геномом для оценки плотности тегов. [16] [17]
- Секвенирование концов бактериальной искусственной хромосомы (BAC) ( профилирование концевых последовательностей ) : идентифицирует точки разрыва хромосом путем создания библиотеки BAC из ракового генома и секвенирования их концов. Клоны BAC, содержащие хромосомные аберрации, имеют концевые последовательности, которые не картируются с аналогичной областью эталонного генома, что указывает на хромосомную точку разрыва. [18]
Транскриптомы
[ редактировать ]- Микрочипы : оценка количества транскриптов . Полезно для классификации, прогнозирования, открывает возможности дифференцированного лечения и помогает идентифицировать мутации в кодирующих областях белков. [19] [20] Относительное обилие альтернативных транскриптов стало важной особенностью исследований рака. Конкретные альтернативные формы транскриптов коррелируют с конкретными типами рака. [21]
- РНК-Seq
Биоинформатика и функциональный анализ онкогенов
[ редактировать ]Технологии биоинформатики позволяют проводить статистический анализ геномных данных. Функциональные характеристики онкогенов еще предстоит установить. Потенциальные функции включают их трансформационные способности, связанные с образованием опухолей, и специфическую роль на каждой стадии развития рака.
После обнаружения соматических раковых мутаций в группе образцов рака можно провести биоинформатический компьютерный анализ для выявления вероятных функциональных и вероятных драйверных мутаций. Для этой идентификации обычно используются три основных подхода: картирование мутаций, оценка влияния мутации на функцию белка или регуляторного элемента и обнаружение признаков положительного отбора в группе опухолей. Однако эти подходы не обязательно являются последовательными, между элементами различных подходов существуют важные отношения приоритета. На каждом этапе используются разные инструменты. [22]
Оперомика
[ редактировать ]Оперомика стремится объединить геномику, транскриптомику и протеомику, чтобы понять молекулярные механизмы, лежащие в основе развития рака. [23]
Сравнительная онкогеномика
[ редактировать ]Сравнительная онкогеномика использует межвидовые сравнения для идентификации онкогенов. Это исследование включает изучение раковых геномов, транскриптомов и протеомов в модельных организмах, таких как мыши, идентификацию потенциальных онкогенов и обращение к образцам рака человека, чтобы выяснить, играют ли гомологи этих онкогенов важную роль в возникновении рака у человека. [24] Генетические изменения в моделях на мышах аналогичны тем, которые обнаруживаются при раке человека. Эти модели создаются с помощью методов, включая ретровирусов инсерционный мутагенез или трансплантацию раковых клеток.
Источник мутаций, вызывающих рак, мутагенез рака
[ редактировать ]Мутации обеспечивают сырье для естественного отбора в эволюции и могут быть вызваны ошибками репликации ДНК, действием экзогенных мутагенов или эндогенными повреждениями ДНК. Механизм репликации и поддержания генома может быть поврежден мутациями или изменен физиологическими условиями и различными уровнями экспрессии при раке (см. [25] ).
Как отметили Гао и др., [26] Стабильность и целостность человеческого генома поддерживаются системой реакции на повреждение ДНК (DDR). Невосстановленные повреждения ДНК являются основной причиной мутаций, которые приводят к канцерогенезу. [27] [28] Если репарация ДНК недостаточна, повреждения ДНК имеют тенденцию накапливаться. Такое избыточное повреждение ДНК может увеличить мутационные ошибки во время репликации ДНК из-за склонного к ошибкам синтеза транслезий . Чрезмерное повреждение ДНК также может усилить эпигенетические изменения из-за ошибок во время восстановления ДНК. [29] [30] Такие мутации и эпигенетические изменения могут привести к раку . Гены DDR часто подавляются при раке человека эпигенетическими механизмами. Такая репрессия может включать метилирование ДНК промоторных областей или репрессию генов DDR с помощью микроРНК. Эпигенетическая репрессия генов DDR происходит чаще, чем генная мутация при многих типах рака (см. Эпигенетика рака ). Таким образом, эпигенетическая репрессия часто играет более важную роль, чем мутация, в снижении экспрессии генов DDR. Это снижение экспрессии генов DDR, вероятно, является важной движущей силой канцерогенеза.
Контекст нуклеотидной последовательности влияет на вероятность мутации [31] [32] [33] и анализ мутационных (мутабельных) мотивов ДНК может иметь важное значение для понимания механизмов мутагенеза при раке. Такие мотивы представляют собой отпечатки пальцев взаимодействий между ДНК и мутагенами, между ДНК и ферментами репарации/репликации/модификации. Примерами мотивов являются мотив AID WRCY/RGYW (W = A или T, R = пурин и Y = пиримидин) с мутациями C на T/G/A, [33] и склонная к ошибкам ДНК pol η приписывала мутации, связанные со AID (от A до G/C/G), в мотивах WA/TW. [34]
Другой (независимый) способ анализа наблюдаемых мутационных спектров и контекста последовательностей ДНК мутаций в опухолях включает объединение всех мутаций разных типов и контекстов из образцов рака в дискретное распределение. Если доступно несколько образцов рака, их контекстно-зависимые мутации можно представить в виде неотрицательной матрицы. Эту матрицу можно дополнительно разложить на компоненты (мутационные сигнатуры), которые в идеале должны описывать отдельные мутагенные факторы. [35] Для решения этой проблемы разложения было предложено несколько вычислительных методов. Первая реализация метода неотрицательной матричной факторизации (NMF) доступна в Mutation Signature Framework Института Сэнгера в виде пакета MATLAB. [36] С другой стороны, если доступны мутации только из одного образца опухоли, пакет DeconstructSigs R [37] и сервер MutaGene [38] может обеспечить идентификацию вклада различных мутационных сигнатур для одного образца опухоли. Кроме того, сервер MutaGene предоставляет мутагенные или специфичные для рака фоновые модели и сигнатуры мутаций, которые можно применять для расчета ожидаемой мутабельности участков ДНК и белков, чтобы разделить относительный вклад мутагенеза и селекции в канцерогенез.
Синтетическая летальность
[ редактировать ]Синтетическая летальность возникает, когда сочетание нарушений экспрессии двух или более генов приводит к гибели клеток, тогда как дефицит только одного из этих генов не приводит. Дефициты могут возникнуть в результате мутаций, эпигенетических изменений или ингибиторов одного из генов.
Терапевтический потенциал синтетической летальности как эффективной стратегии борьбы с раком постоянно улучшается. В последнее время применимость синтетической летальности к таргетной терапии рака возросла благодаря недавней работе ученых, в том числе Рональда А. ДеПиньо и его коллег, в так называемой «побочной летальности». Мюллер и др. обнаружили, что гены-пассажиры, находящиеся в хромосомной близости к генам-супрессорам опухолей, одновременно удаляются при некоторых видах рака. [39] Таким образом, идентификация коллатерально удаленных избыточных генов, выполняющих важную клеточную функцию, может стать неиспользованным резервом для дальнейшего использования подхода синтетической летальности . Таким образом, побочная летальность имеет большой потенциал для выявления новых и селективных терапевтических целей в онкологии. [40] В 2012 году Мюллер и др. выявили, что гомозиготная делеция избыточно-существенного гликолитического гена ENO1 человека при глиобластоме (GBM) является следствием близости к делециям локуса-супрессора опухоли 1p36 и может иметь потенциал для синтетического летального подхода к ингибированию GBM. [39] ENO1 — один из трех гомологичных генов ( ENO2 , ENO3 ), которые кодируют фермент альфа-енолазу млекопитающих . [41] ENO2, который кодирует енолазу 2 , в основном экспрессируется в нервных тканях, что приводит к предположению, что при GBM с делецией ENO1 ENO2 может быть идеальной мишенью в качестве избыточного гомолога ENO1. [42] Мюллер обнаружил, что как генетическое, так и фармакологическое ингибирование ENO2 в клетках GBM с гомозиготной делецией ENO1 вызывает синтетическую летальность за счет селективного уничтожения клеток GBM. [39] В 2016 году Мюллер и его коллеги обнаружили антибиотик SF2312 как высокоэффективный ингибитор енолазы наномолярного диапазона , который преимущественно ингибирует пролиферацию клеток глиомы и гликолитический поток в клетках с делецией ENO1. [43] Было показано, что SF2312 более эффективен, чем ингибитор паненолазы PhAH, и обладает большей специфичностью к ингибированию ENO2, чем ENO1. [43] Последующая работа той же группы показала, что тот же подход можно применить к раку поджелудочной железы , при этом гомозиготное удаление SMAD4 приводит к побочной делеции митохондриального яблочного фермента 2 ( ME2 ), окислительной декарбоксилазы, необходимой для окислительно-восстановительного гомеостаза. [44] Дей и др. показали, что геномная делеция ME2 в клетках аденокарциномы протоков поджелудочной железы приводит к образованию большого количества эндогенных активных форм кислорода, что соответствует KRAS-индуцированному раку поджелудочной железы , и, по существу, подготавливает ME2-нулевые клетки к синтетической летальности за счет истощения избыточной NAD(P)+-зависимой изоформы ME3. Было обнаружено, что эффекты истощения ME3 опосредованы ингибированием синтеза нуклеотидов de novo в результате активации AMPK и митохондриального ROS-опосредованного апоптоза. [45] [44] Между тем, Оике и др. продемонстрировали возможность обобщения концепции путем воздействия на избыточные незаменимые гены, участвующие в процессах, отличных от метаболизма, а именно на субъединицы SMARCA4 и SMARCA2 ремоделирующем хроматин в комплексе SWI/SNF, . [46]
Некоторые онкогены необходимы для выживания всех клеток (не только раковых). Таким образом, лекарства, которые выбивают эти онкогены (и тем самым убивают раковые клетки), могут также повредить нормальные клетки, вызывая серьезные заболевания. Однако другие гены могут быть важны для раковых клеток, но не для здоровых клеток.
Лечение, основанное на принципе синтетической летальности, продлило выживаемость онкологических больных и открывает перспективы для будущих достижений в обращении канцерогенеза. Основной тип синтетической летальности связан с дефектом репарации ДНК, который часто инициирует рак и все еще присутствует в опухолевых клетках. Некоторые примеры приведены здесь.
Экспрессия BRCA1 или BRCA2 недостаточна при большинстве случаев рака молочной железы и яичников высокой степени злокачественности, обычно из-за эпигенетического метилирования его промотора или эпигенетической репрессии сверхэкспрессируемой микроРНК (см. статьи BRCA1 и BRCA2 ). BRCA1 и BRCA2 являются важными компонентами основного пути гомологичной рекомбинационной репарации двухцепочечных разрывов. Если одного или другого недостаточно, это увеличивает риск развития рака, особенно рака молочной железы или яичников. Резервный путь восстановления ДНК для некоторых повреждений, обычно восстанавливаемых с помощью BRCA1 и BRCA2, зависит от PARP1 . Таким образом, многие виды рака яичников реагируют на одобренное FDA лечение ингибитором PARP, вызывая синтетическую гибель раковых клеток, дефицитных по BRCA1 или BRCA2. Это лечение также оценивается при раке молочной железы и многих других видах рака в рамках клинических испытаний III фазы в 2016 году. [47]
Существует два пути гомологичной рекомбинационной репарации двухцепочечных разрывов. Основной путь зависит от BRCA1, PALB2 и BRCA2, тогда как альтернативный путь зависит от RAD52. [48] Доклинические исследования с участием эпигенетически уменьшенных или мутированных клеток с дефицитом BRCA (в культуре или инъекций мышам) показывают, что ингибирование RAD52 синтетически летально при дефиците BRCA. [49]
Мутации в генах, участвующих в восстановлении несоответствий ДНК (MMR), вызывают высокую частоту мутаций. [50] В опухолях такие частые последующие мутации часто приводят к образованию «чужих» иммуногенных антигенов. Клиническое исследование фазы II на людях с участием 41 пациента оценивало один синтетический летальный подход к опухолям с дефектами MMR или без них. [51] Продукт гена PD-1 обычно подавляет цитотоксические иммунные реакции. Ингибирование этого гена обеспечивает более сильный иммунный ответ. Когда онкологические больные с дефектом MMR в опухолях подвергались воздействию ингибитора PD-1, у 67–78% пациентов наблюдалась выживаемость без прогрессирования, связанная с иммунитетом. Напротив, для пациентов без дефектного MMR добавление ингибитора PD-1 привело только к 11% пациентов с выживаемостью без иммунозависимого прогрессирования. Таким образом, ингибирование PD-1 в первую очередь является синтетически летальным при дефектах MMR.
ARID1A , модификатор хроматина, необходим для негомологичного соединения концов , основного пути, который восстанавливает двухцепочечные разрывы в ДНК. [52] а также играет регулирующую роль транскрипции. [53] Мутации ARID1A являются одной из 12 наиболее распространенных канцерогенных мутаций. [54] Мутация или эпигенетическое снижение экспрессии [55] ARID1A обнаружен при 17 типах рака. [56] Доклинические исследования на клетках и мышах показывают, что синтетическая летальность при дефиците ARID1A возникает либо за счет ингибирования метилтрансферазной активности EZH2, либо за счет ингибирования метилтрансферазной активности EZH2, [57] [58] или с добавлением ингибитора киназы дазатиниба. [59]
Другой подход заключается в индивидуальном отключении каждого гена в геноме и наблюдении эффекта на нормальных и раковых клетках. [60] [61] Если нокаут несущественного в других отношениях гена мало влияет или вообще не влияет на здоровые клетки, но является летальным для раковых клеток, содержащих мутировавший онкоген, то общесистемное подавление подавленного гена может уничтожить раковые клетки, оставив при этом здоровые относительно неповрежденными. Этот метод использовался для идентификации ингибиторов PARP-1 для лечения рака, связанного с BRCA1/BRCA2. [62] [63] В этом случае совокупное присутствие ингибирования PARP-1 и мутаций, связанных с раком, в генах BRCA является летальным только для раковых клеток.
Базы данных для исследований рака
[ редактировать ]Проект «Геном рака» — это инициатива по выявлению всех соматических мутаций при раке. В рамках проекта систематически секвенируются экзоны и фланкирующие сплайсинговые соединения геномов первичных опухолей и линий раковых клеток. Программное обеспечение COSMIC отображает данные, полученные в результате этих экспериментов. По состоянию на февраль 2008 года CGP выявила 4746 генов и 2985 мутаций в 1848 опухолях.
Проект анатомии генома рака включает информацию об исследованиях геномов, транскриптомов и протеомов рака.
Progenetix — это онкогеномная справочная база данных, в которой представлены цитогенетические и молекулярно-цитогенетические данные об опухолях.
Компания Oncomine собрала данные профилей транскриптома рака.
Интегративная база данных онкогеномики IntOGen и наборы данных Gitools объединяют многомерные онкогеномные данные человека, классифицированные по типам опухолей. Первая версия IntOGen была сосредоточена на роли дерегулированной экспрессии генов и CNV при раке. [64] В более поздней версии особое внимание уделялось мутационным генам, вызывающим рак, в 28 типах опухолей. [65] [66] Все выпуски данных IntOGen доступны в базе данных IntOGen.
Международный консорциум генома рака — крупнейший проект по сбору данных о геноме рака человека. Данные доступны через веб-сайт ICGC. BioExpress® Oncology Suite содержит данные об экспрессии генов из образцов первичных, метастатических и доброкачественных опухолей, а также нормальных образцов, включая соответствующие соседние контрольные образцы. В комплект входят образцы гематологических злокачественных опухолей многих известных видов рака.
Конкретные базы данных для модельных животных включают Базу данных генов рака, меченных ретровирусами (RTCGD), в которой собраны исследования по инсерционному мутагенезу ретровирусов и транспозонов в опухолях мышей.
Генные семьи
[ редактировать ]Мутационный анализ целых семейств генов показал, что гены одного и того же семейства имеют схожие функции, о чем свидетельствуют схожие кодирующие последовательности и белковые домены . Двумя такими классами являются семейство киназ , участвующих в добавлении фосфатных групп к белкам, и семейство фосфатаз , участвующих в удалении фосфатных групп из белков. [67] Эти семейства были впервые исследованы из-за их очевидной роли в передаче клеточных сигналов роста или гибели клеток. В частности, более 50% случаев колоректального рака несут мутацию в гене киназы или фосфатазы. Ген фосфатидилинозитолд-3-киназы ( PIK3CA ) кодирует липидкиназы, которые обычно содержат мутации при колоректальном раке, раке молочной железы, раке желудка, легких и различных других видах рака. [68] [69] Лекарственная терапия может ингибировать PIK3CA. Другой пример — ген BRAF , один из первых, участвующих в развитии меланомы. [70] BRAF кодирует серин / треониновую киназу, которая участвует в сигнальном пути роста RAS-RAF- MAPK . Мутации BRAF вызывают конститутивное фосфорилирование и активность в 59% меланом. До появления BRAF генетический механизм развития меланомы был неизвестен, поэтому прогноз для пациентов был плохим. [71]
Митохондриальная ДНК
[ редактировать ]Мутации митохондриальной ДНК (мтДНК) связаны с образованием опухолей. Были идентифицированы четыре типа мутаций мтДНК: [72]
Точечные мутации
[ редактировать ]Точечные мутации наблюдались в кодирующей и некодирующей области мтДНК, содержащейся в раковых клетках. У людей с раком мочевого пузыря, головы/шеи и легких точечные мутации в кодирующей области демонстрируют признаки сходства друг с другом. Это говорит о том, что когда здоровая клетка трансформируется в опухолевую (неопластическая трансформация), митохондрии кажутся гомогенными. Обильные точечные мутации, расположенные в некодирующей области D-петли раковых митохондрий, позволяют предположить, что мутации в этой области могут быть важной характеристикой некоторых видов рака. [72]
Удаление
[ редактировать ]Этот тип мутации выявляется спорадически из-за его небольшого размера (< 1 т.п.н.). Появление определенных специфических мутаций мтДНК (делеция 264 п.н. и делеция 66 п.н. в комплексе 1-субъединицы гена ND1) при множественных типах рака дает некоторые доказательства того, что небольшие делеции мтДНК могут появляться в начале онкогенеза . Это также предполагает, что количество митохондрий, содержащих эти делеции, увеличивается по мере прогрессирования опухоли. Исключением является относительно крупная делеция, которая появляется при многих видах рака (известная как «обычная делеция»), но в нормальных клетках обнаружено больше крупномасштабных делеций мтДНК, чем в опухолевых клетках. Это может быть связано с, по-видимому, адаптивным процессом опухолевых клеток по устранению любых митохондрий, содержащих эти крупномасштабные делеции («обычная делеция» составляет> 4 т.п.н.). [72]
Вставки
[ редактировать ]Две небольшие вставки мтДНК размером ~260 и ~520 п.н. могут присутствовать при раке молочной железы, раке желудка, гепатоцеллюлярной карциноме (ГЦК) и раке толстой кишки, а также в нормальных клетках. Никакой корреляции между этими инсерциями и раком не установлено. [73]
Копирование числовых мутаций
[ редактировать ]Характеристика мтДНК с помощью полимеразной цепной реакции в реальном времени показывает наличие количественного изменения количества копий мтДНК при многих видах рака. Ожидается, что увеличение числа копий произойдет из-за окислительного стресса. С другой стороны, считается, что снижение вызвано соматическими точечными мутациями в сайте начала репликации H-цепи и/или гомополимерного c-участка D310 в области D-петли, мутациями в p53 (гене-супрессоре опухоли). опосредованный путь и/или неэффективная активность фермента из-за мутаций POLG . Любое увеличение/уменьшение количества копий остается постоянным внутри опухолевых клеток. Тот факт, что количество мтДНК в опухолевых клетках постоянно, позволяет предположить, что количество мтДНК в опухолевых клетках контролируется гораздо более сложной системой, а не просто изменяется вследствие аномальной пролиферации клеток. Роль содержания мтДНК при раке человека, по-видимому, варьируется в зависимости от типа или локализации опухоли. [72]
| Тип рака | Расположение точечных мутаций | Нуклеотидное положение делеций | Увеличение количества копий мтДНК | Уменьшение количества копий мтДНК | |||
|---|---|---|---|---|---|---|---|
| D-петля | мРНК | тРНК | рРНК | ||||
| мочевой пузырь [74] | Х | Х | Х | 15,642-15,662 | |||
| Грудь [75] [76] [77] [78] | Х | Х | Х | Х | 8470-13447 и 8482-13459 | Х | |
| Голова и шея [75] [79] [80] | Х | Х | Х | Х | 8470-13447 и 8482-13459 | Х | |
| Оральный [81] | Х | Х | 8470-13447 и 8482-13459 | ||||
| Гепатоцеллюлярная карцинома (ГЦК) [82] [83] | Х | Х | Х | Х | 306-556 и 3894-3960 | Х | |
| Пищеводный [84] | Х | Х | Х | 8470-13447 и 8482-13459 | Х | ||
| Желудочный [85] [86] [87] | Х | Х | Х | 298-348 | Х | ||
| Простата [88] [89] | Х | Х | 8470-13447 и 8482-13459 | Х | |||
57,7% (500/867) содержали соматические точечные мутации, а из 1172 исследованных мутаций 37,8% (443/1127) были расположены в контрольной области D-петли, 13,1% (154/1172) были расположены в генах тРНК или рРНК и 49,1% (575/1127) были обнаружены в генах мРНК, необходимых для образования комплексов, необходимых для митохондриального дыхания.
Диагностические приложения
[ редактировать ]Некоторые противораковые препараты нацелены на мтДНК и показали положительные результаты в уничтожении опухолевых клеток. Исследования использовали митохондриальные мутации в качестве биомаркеров для терапии раковых клеток. Легче выявить мутацию в митохондриальной ДНК, чем в ядерной ДНК, поскольку митохондриальный геном намного меньше и его легче проверять на предмет конкретных мутаций. Изменения содержания мтДНК, обнаруженные в образцах крови, могут служить маркером скрининга для прогнозирования будущей восприимчивости к раку, а также для отслеживания прогрессирования злокачественных опухолей. Наряду с этими потенциально полезными характеристиками мтДНК она не находится под контролем клеточного цикла и важна для поддержания генерации АТФ и митохондриального гомеостаза. Эти характеристики делают нацеливание на мтДНК практической терапевтической стратегией. [72]
Биомаркеры рака
[ редактировать ]Некоторые биомаркеры могут быть полезны для определения стадии, прогноза и лечения рака. Они могут варьироваться от однонуклеотидных полиморфизмов (SNP), хромосомных аберраций , изменений числа копий ДНК, нестабильности микросателлитов, метилирования промоторной области или даже высоких или низких уровней белка. [90] В период с 2013 по 2019 год только 6,8% людей с онкологическими заболеваниями в двух штатах США прошли генетическое тестирование, что указывает на широкое недостаточное использование информации, которая могла бы улучшить решения о лечении и результаты лечения пациентов. [91]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б Штраусберг Р.Л., Симпсон А.Дж., Олд Л.Дж., Риггинс Г.Дж. (май 2004 г.). «Онкогеномика и разработка новых методов лечения рака». Природа . 429 (6990): 469–74. Бибкод : 2004Natur.429..469S . дои : 10.1038/nature02627 . ПМИД 15164073 . S2CID 37628107 .
- ^ Тинг А.Х., МакГарви К.М., Бэйлин С.Б. (2006). «Эпигеном рака — компоненты и функциональные корреляты» . Генс Дев . 20 (23): 3215–31. дои : 10.1101/gad.1464906 . ПМИД 17158741 .
- ^ Джонс П.А., Бэйлин С.Б. (2007). «Эпигеномика рака» . Клетка . 128 (4): 683–92. дои : 10.1016/j.cell.2007.01.029 . ПМЦ 3894624 . ПМИД 17320506 .
- ^ Ли Д., Чжан Б., Син Икс, Ван Т (2015). «Объединение MeDIP-seq и MRE-seq для исследования полногеномного метилирования CpG» . Методы . 72 : 29–40. дои : 10.1016/j.ymeth.2014.10.032 . ПМК 4300244 . ПМИД 25448294 .
- ^ Вэй Дж, Ли Г, Данг С, Чжоу Ю, Цзэн К, Лю М (2016). «Открытие и проверка гиперметилированных маркеров колоректального рака» . Дис. Маркеры . 2016 : 2192853. дои : 10.1155/2016/2192853 . ПМЦ 4963574 . ПМИД 27493446 .
- ^ Jump up to: а б Штраусберг Р.Л., Симпсон А.Дж. (январь 2010 г.). «Полногеномный анализ рака как подход к более глубокому пониманию биологии опухолей» . Бр. Дж. Рак . 102 (2): 243–8. дои : 10.1038/sj.bjc.6605497 . ПМЦ 2816661 . ПМИД 20029419 .
- ^ Фогельштейн Б., Пападопулос Н., Велкулеску В.Е., Чжоу С., Диас Л.А., Кинцлер К.В. (2013). «Пейзажи генома рака» . Наука . 339 (6127): 1546–58. Бибкод : 2013Sci...339.1546V . дои : 10.1126/science.1235122 . ПМК 3749880 . ПМИД 23539594 .
- ^ Луо Й, Вонг С.Дж., Каз А.М., Дзечатковски С., Картер К.Т., Моррис С.М., Ван Дж., Уиллис Дж.Э., Макар К.В., Ульрих СМ, Латтербо Дж.Д., Шрубсол М.Дж., Чжэн В., Марковиц С.Д., Грейди В.М. (2014). «Различия в признаках метилирования ДНК указывают на множество путей прогрессирования от аденомы до колоректального рака» . Гастроэнтерология . 147 (2): 418–29.e8. дои : 10.1053/j.gastro.2014.04.039 . ПМК 4107146 . ПМИД 24793120 .
- ^ Лей Т.Дж., Мардис Э.Р., Дин Л., Фултон Б., Маклеллан М.Д., Чен К. и др. (ноябрь 2008 г.). «Секвенирование ДНК цитогенетически нормального генома острого миелолейкоза» . Природа . 456 (7218): 66–72. Бибкод : 2008Natur.456...66L . дои : 10.1038/nature07485 . ПМК 2603574 . ПМИД 18987736 .
- ^ Jump up to: а б Пейкофф, Кира (16 октября 2015 г.). «Что чудесные выздоровления говорят нам о победе над раком» . Популярная механика . Проверено 28 апреля 2016 г.
- ^ Барделли А .; Велкулеску В.Е. (2005). «Мутационный анализ семейств генов рака человека». Текущее мнение в области генетики и развития . 15 (1): 5–12. дои : 10.1016/j.gde.2004.12.009 . ПМИД 15661527 .
- ^ Бенвенути С.; Арена С.; Барделли А. (2005). «Идентификация генов рака путем мутационного профилирования геномов опухолей». Письма ФЭБС . 579 (8): 1884–1890. дои : 10.1016/j.febslet.2005.02.015 . ПМИД 15763568 . S2CID 31708201 .
- ^ Ши ИМ; Ван Т.Л. (2005). «Применить инновационные технологии для изучения генома рака». Современное мнение в онкологии . 17 (1): 33–38. дои : 10.1097/01.cco.0000147382.97085.e4 . PMID 15608510 . S2CID 39104975 .
- ^ Грешок Дж., Нейлор Т.Л., Марголин А., Дискин С., Кливер Ш., Фуреал П.А. и др. (январь 2004 г.). «Сравнительная геномная гибридизация на основе массива с разрешением 1 Мб с использованием набора клонов BAC, оптимизированного для анализа генов рака» . Геном Рез . 14 (1): 179–87. дои : 10.1101/гр.1847304 . ПМК 314295 . ПМИД 14672980 .
- ^ Люсито Р., Хили Дж., Александр Дж., Райнер А., Эспозито Д., Чи М. и др. (октябрь 2003 г.). «Репрезентативный анализ олигонуклеотидов на микрочипах: метод высокого разрешения для обнаружения вариаций числа копий генома» . Геном Рез . 13 (10): 2291–305. дои : 10.1101/гр.1349003 . ПМК 403708 . ПМИД 12975311 .
- ^ Ху М., Яо Дж., Поляк К. (2006). «Цифровое кариотипирование, специфичное для метилирования». Нат Проток . 1 (3): 1621–36. дои : 10.1038/nprot.2006.278 . ПМИД 17406428 . S2CID 10554933 .
- ^ Кёрнер Х., Епанчинцев А., Беркинг С., Шулер-Турнер Б., Спайхер М.Р., Менссен А., Хермекинг Х. (январь 2007 г.). «Цифровое кариотипирование выявляет частую инактивацию гена дистрофина/DMD при злокачественной меланоме» . Клеточный цикл . 6 (2): 189–98. дои : 10.4161/cc.6.2.3733 . ПМИД 17314512 .
- ^ Волик С., Чжао С., Чин К., Бребнер Дж.Х., Херндон Д.Р., Тао К. и др. (июнь 2003 г.). «Профилирование конечных последовательностей: анализ аберрантных геномов на основе последовательностей» . Учеб. Натл. акад. наук. США . 100 (13): 7696–701. Бибкод : 2003PNAS..100.7696V . дои : 10.1073/pnas.1232418100 . ПМК 164650 . ПМИД 12788976 .
- ^ Ван, де Вийвер М.Дж.; и др. (2002). «Сигнатура экспрессии генов как предиктор выживаемости при раке молочной железы». Медицинский журнал Новой Англии . 347 (25): 1999–2009. doi : 10.1056/NEJMoa021967 . hdl : 1874/15577 . ПМИД 12490681 .
- ^ Ван 'т Вир Л.Дж., Дай Х., ван де Вийвер М.Дж., Хе Ю.Д., Харт А.А., Бернардс Р., Френд Ш.Х. (2003). «Профилирование экспрессии предсказывает исход рака молочной железы» . Рак молочной железы . 5 (1): 57–8. дои : 10.1186/bcr562 . ПМЦ 154139 . ПМИД 12559048 .
- ^ Сюй Ц.; Ли С. (2003). «Открытие новых форм сплайсинга и функциональный анализ специфичного для рака альтернативного сплайсинга в экспрессируемых последовательностях человека» . Исследования нуклеиновых кислот . 31 (19): 5635–5643. дои : 10.1093/нар/gkg786 . ПМК 206480 . ПМИД 14500827 .
- ^ Гонсалес-Перес А., Мустонен В., Рева Б., Ричи Г.Р., Крейкселл П., Карчин Р. и др. (август 2013 г.). «Вычислительные подходы к идентификации функциональных генетических вариантов в геномах рака» . Нат. Методы . 10 (8): 723–9. дои : 10.1038/nmeth.2562 . ПМЦ 3919555 . ПМИД 23900255 .
- ^ Ханаш С.М. (сентябрь 2000 г.). «Оперомика: молекулярный анализ тканей от ДНК до РНК и белка». Клин. хим. Лаб. Мед . 38 (9): 805–13. дои : 10.1515/CCLM.2000.116 . ПМИД 11097332 . S2CID 34163524 .
- ^ Пипер Д., Бернс А. (июнь 2006 г.). «Межвидовая онкогеномика в идентификации генов рака» . Клетка . 125 (7): 1230–3. дои : 10.1016/j.cell.2006.06.018 . ПМИД 16814709 .
- ^ Рогозин И.Б., Павлов Ю.И., Гончаренко А., Де С., Лада А.Г., Поляков Е., Панченко А.Р., Купер Д.Н. (ноябрь 2018 г.). «Мутационные сигнатуры и изменчивые мотивы в геномах рака» . Краткий. Биоинформатика . 19 (6): 1085–1101. дои : 10.1093/нагрудник/bbx049 . ПМК 6454500 . ПМИД 28498882 .
- ^ Гао Д., Герман Дж.Г. , Го М. (2016). «Клиническое значение аберрантных эпигенетических изменений генов восстановления повреждений ДНК при раке человека» . Онкотаргет . 7 (24): 37331–37346. дои : 10.18632/oncotarget.7949 . ПМК 5095080 . ПМИД 26967246 .
- ^ Кастан М.Б. (2008). «Реакция на повреждение ДНК: механизмы и роль в заболеваниях человека: лекция на премию Мемориала ГСГ Клоуза 2007 г.» (PDF) . Мол. Рак Рез . 6 (4): 517–24. дои : 10.1158/1541-7786.MCR-08-0020 . ПМИД 18403632 .
- ^ Бернштейн, К; Прасад, Арканзас; Нфонсам, В; Бернштейн, Х. (2013). «Глава 16: Повреждение ДНК, восстановление ДНК и рак». В Чен, Кларк (ред.). Новые направления исследований в области репарации ДНК . Совет директоров – Книги по запросу. п. 413. ИСБН 978-953-51-1114-6 .
- ^ О'Хаган Х.М., Мохаммад Х.П., Бэйлин С.Б. (2008). «Двухнитевые разрывы могут инициировать молчание генов и SIRT1-зависимое начало метилирования ДНК на экзогенном промоторном острове CpG» . ПЛОС Генетика . 4 (8): е1000155. дои : 10.1371/journal.pgen.1000155 . ПМЦ 2491723 . ПМИД 18704159 .
- ^ Куоццо С., Порчеллини А., Ангризано Т. и др. (июль 2007 г.). «Повреждение ДНК, репарация, направленная на гомологию, и метилирование ДНК» . ПЛОС Генетика . 3 (7): е110. дои : 10.1371/journal.pgen.0030110 . ЧВК 1913100 . ПМИД 17616978 .
- ^ Кулондр С., Миллер Дж. Х., Фарабо П. Дж., Гилберт В. (август 1978 г.). «Молекулярная основа горячих точек замещения оснований в Escherichia coli». Природа . 274 (5673): 775–80. Бибкод : 1978Natur.274..775C . дои : 10.1038/274775a0 . ПМИД 355893 . S2CID 4165194 .
- ^ Купер Д.Н., Юсуфьян Х. (февраль 1988 г.). «Динуклеотид CpG и генетические заболевания человека». Хм. Жене . 78 (2): 151–5. дои : 10.1007/bf00278187 . ПМИД 3338800 . S2CID 41948691 .
- ^ Jump up to: а б Рогозин И.Б., Колчанов Н.А. (ноябрь 1992 г.). «Соматический гипермутагенез в генах иммуноглобулинов. II. Влияние соседних последовательностей оснований на мутагенез». Биохим. Биофиз. Акта . 1171 (1): 11–8. дои : 10.1016/0167-4781(92)90134-л . ISSN 0006-3002 . ПМИД 1420357 .
- ^ Рогозин И.Б., Павлов Ю.И., Бебенек К., Мацуда Т., Кункель Т.А. (июнь 2001 г.). «Горячие точки соматических мутаций коррелируют со спектром ошибок ДНК-полимеразы эта». Нат. Иммунол . 2 (6): 530–6. дои : 10.1038/88732 . ISSN 1529-2908 . ПМИД 11376340 . S2CID 12807889 .
- ^ Ник-Зайнал С., Александров Л.Б., Ведж Д.С., Ван Лу П., Гринман К.Д., Рейн К. и др. (май 2012 г.). «Мутационные процессы, формирующие геномы 21 рака молочной железы» . Клетка . 149 (5): 979–93. дои : 10.1016/j.cell.2012.04.024 . ПМЦ 3414841 . ПМИД 22608084 .
- ^ Александров Л.Б., Ник-Зайнал С., Ведж Д.С., Кэмпбелл П.Дж., Страттон М.Р. (январь 2013 г.). «Расшифровка признаков мутационных процессов, происходящих при раке человека» . Представитель ячейки . 3 (1): 246–59. дои : 10.1016/j.celrep.2012.12.008 . ПМЦ 3588146 . ПМИД 23318258 .
- ^ Розенталь Р., МакГранахан Н., Эрреро Дж., Тейлор Б.С., Суонтон С. (февраль 2016 г.). «DeconstructSigs: определение мутационных процессов в отдельных опухолях позволяет выявить недостатки репарации ДНК и закономерности эволюции карциномы» . Геном Биол . 17:31 . дои : 10.1186/s13059-016-0893-4 . ПМЦ 4762164 . ПМИД 26899170 .
- ^ Гончаренко А., Рагер С.Л., Ли М., Санг QX, Рогозин И.Б., Панченко А.Р. (июль 2017 г.). «Изучение фоновых мутационных процессов для расшифровки генетической гетерогенности рака» . Нуклеиновые кислоты Рез . 45 (П1): W514–W522. дои : 10.1093/нар/gkx367 . ПМЦ 5793731 . ПМИД 28472504 .
- ^ Jump up to: а б с Мюллер Ф.Л., Колла С., Аквиланти Е., Манзо В.Е., Дженовезе Г., Ли Дж. и др. (август 2012 г.). «Удаления пассажиров создают терапевтические уязвимости при раке» . Природа . 488 (7411): 337–42. Бибкод : 2012Natur.488..337M . дои : 10.1038/nature11331 . ПМЦ 3712624 . ПМИД 22895339 .
- ^ Мюллер Флорида, Аквиланти Э.А., ДеПиньо Р.А. (ноябрь 2015 г.). «Побочная летальность: новая терапевтическая стратегия в онкологии» . Тенденции Рака . 1 (3): 161–173. дои : 10.1016/j.trecan.2015.10.002 . ПМК 4746004 . ПМИД 26870836 .
- ^ Пойнер Р.Р., Рид Г.Х. (август 1992 г.). «Структура бивалентного катионного комплекса с фосфоноацетогидроксаматом в активном центре енолазы». Биохимия . 31 (31): 7166–73. дои : 10.1021/bi00146a020 . ПМИД 1322695 .
- ^ Джозеф Дж., Крус-Санчес ФФ, Каррерас Дж. (июнь 1996 г.). «Активность енолазы и распределение изоферментов в областях мозга и опухолях человека». Дж. Нейрохем . 66 (6): 2484–90. дои : 10.1046/j.1471-4159.1996.66062484.x . ПМИД 8632173 . S2CID 24655147 .
- ^ Jump up to: а б Леонард П.Г., Сатани Н., Максвелл Д., Лин Ю.Х., Хаммуди Н., Пэн З. и др. (декабрь 2016 г.). «SF2312 является природным фосфонатным ингибитором енолазы» . Нат. хим. Биол . 12 (12): 1053–1058. дои : 10.1038/nchembio.2195 . ПМК 5110371 . ПМИД 27723749 .
- ^ Jump up to: а б Дей П., Баддур Дж., Мюллер Ф., Ву CC, Ван Х., Ляо В.Т. и др. (февраль 2017 г.). «Геномная делеция яблочного фермента 2 приводит к побочной летальности при раке поджелудочной железы» . Природа . 542 (7639): 119–123. Бибкод : 2017Natur.542..119D . дои : 10.1038/nature21052 . ПМК 5398413 . ПМИД 28099419 .
- ^ Лиу Г.И., Допплер Х., ДельДжорно К.Е., Чжан Л., Лейтгес М., Кроуфорд Х.К., Мерфи М.П., Сторц П. (март 2016 г.). «Мутантный KRas-индуцированный митохондриальный окислительный стресс в ацинарных клетках усиливает передачу сигналов EGFR, что приводит к образованию предраковых поражений поджелудочной железы» . Представитель ячейки . 14 (10): 2325–36. дои : 10.1016/j.celrep.2016.02.029 . ПМЦ 4794374 . ПМИД 26947075 .
- ^ Оике Т., Огивара Х., Томинага Ю., Ито К., Андо О., Цута К. и др. (сентябрь 2013 г.). «Синтетическая стратегия лечения рака, основанная на летальности, с генетической недостаточностью фактора ремоделирования хроматина BRG1» . Рак Рез . 73 (17): 5508–18. дои : 10.1158/0008-5472.CAN-12-4593 . ПМИД 23872584 .
- ^ Мурата С., Чжан С., Финч Н., Чжан К., Кампо Л., Брейер Е.К. (2016). «Предсказатели и модуляторы синтетической летальности: обновленная информация об ингибиторах PARP и персонализированной медицине» . Биомед Рес Инт . 2016 : 2346585. дои : 10.1155/2016/2346585 . ПМК 5013223 . ПМИД 27642590 .
- ^ Лок Б.Х., Карли А.С., Чанг Б., Пауэлл С.Н. (2013). «Инактивация RAD52 является синтетически летальной при дефиците BRCA1 и PALB2 в дополнение к BRCA2 в результате гомологичной рекомбинации, опосредованной RAD51» . Онкоген . 32 (30): 3552–8. дои : 10.1038/onc.2012.391 . ПМК 5730454 . ПМИД 22964643 .
- ^ Крамер-Моралес К., Неборовска-Скорска М., Шейбнер К., Пэджет М., Ирвин Д.А., Сливински Т., Хаас К., Ли Дж., Генг Х., Рой Д., Слупианек А., Рассул Ф.В., Васик М.А., Чайлдерс В., Копленд М., Мюшен М., Цивин С.И., Скорски Т. (2013). «Персонализированная синтетическая летальность, индуцированная нацеливанием на RAD52 при лейкозах, идентифицированных по генной мутации и профилю экспрессии» . Кровь . 122 (7): 1293–304. doi : 10.1182/blood-2013-05-501072 . ПМК 3744994 . ПМИД 23836560 .
- ^ Эшлеман Дж.Р., Ланг Э.З., Бауэрфинд Г.К., Парсонс Р., Фогельштейн Б., Уилсон Дж.К., Вейгл М.Л., Седвик В.Д., Марковиц С.Д. (1995). «Повышенная частота мутаций в локусе hprt сопровождает микросателлитную нестабильность при раке толстой кишки». Онкоген . 10 (1): 33–7. ПМИД 7824277 .
- ^ Ле Д.Т., Урам Дж.Н., Ван Х., Бартлетт Б.Р., Кемберлинг Х., Айринг А.Д. и др. (2015). «Блокада PD-1 в опухолях с дефицитом репарации несоответствия» . Н. англ. Дж. Мед . 372 (26): 2509–20. дои : 10.1056/NEJMoa1500596 . ПМЦ 4481136 . ПМИД 26028255 .
- ^ Ватанабе Р., Уи А., Канно С., Огивара Х., Нагасе Т., Коно Т., Ясуи А. (2014). «Факторы SWI/SNF, необходимые для клеточной устойчивости к повреждению ДНК, включают ARID1A и ARID1B и демонстрируют взаимозависимую стабильность белков» . Рак Рез . 74 (9): 2465–75. дои : 10.1158/0008-5472.CAN-13-3608 . ПМИД 24788099 .
- ^ Рааб-младший, Резник С., Магнусон Т. (2015). «Пологеномная регуляция транскрипции, опосредованная биохимически различными комплексами SWI/SNF» . ПЛОС Генет . 11 (12): e1005748. дои : 10.1371/journal.pgen.1005748 . ПМЦ 4699898 . ПМИД 26716708 .
- ^ Лоуренс М.С., Стоянов П., Мермель Ч.Х., Робинсон Дж.Т., Гарравэй Л.А., Голуб Т.Р., Мейерсон М., Габриэль С.Б., Ландер Э.С., Гетц Г. (2014). «Обнаружение и анализ насыщения генов рака в 21 типе опухолей» . Природа . 505 (7484): 495–501. Бибкод : 2014Natur.505..495L . дои : 10.1038/nature12912 . ПМК 4048962 . ПМИД 24390350 .
- ^ Чжан X, Сунь Ц, Шан М, Ню М, Лю Т, Ся Б, Лян Х, Вэй В, Сунь С, Чжан Ю, Лю XS, Сун Ц, Ян Ю, Ма Ю, Лю Ю, Ян Л, Рен Ю , Чжан Г, Пан Д (2013). «Гиперметилирование промотора гена ARID1A ответственно за низкую экспрессию его мРНК при многих инвазивных формах рака молочной железы» . ПЛОС ОДИН . 8 (1): e53931. Бибкод : 2013PLoSO...853931Z . дои : 10.1371/journal.pone.0053931 . ПМЦ 3549982 . ПМИД 23349767 .
- ^ Ву Дж.Н., Робертс CW (2013). «Мутации ARID1A при раке: еще один эпигенетический супрессор опухоли?» . Рак Дисков . 3 (1): 35–43. дои : 10.1158/2159-8290.CD-12-0361 . ПМЦ 3546152 . ПМИД 23208470 .
- ^ Битлер Б.Г., Эйрд К.М., Гарипов А., Ли Х., Аматанджело М., Коссенков А.В., Шульц Д.К., Лю К, Ши ИеМ, Конехо-Гарсия Дж.Р., Спейчер Д.В., Чжан Р. (2015). «Синтетическая летальность путем воздействия на активность метилтрансферазы EZH2 при раке с мутацией ARID1A» . Нат. Мед . 21 (3): 231–8. дои : 10.1038/нм.3799 . ПМЦ 4352133 . ПМИД 25686104 .
- ^ Ким К.Х., Ким В., Ховард Т.П., Васкес Ф., Черняк А., Ву Дж.Н., Ван В., Хасуэлл Дж.Р., Валенски Л.Д., Хан В.К., Оркин Ш., Робертс К.В. (2015). «Рак с мутациями SWI/SNF зависит от каталитической и некаталитической активности EZH2» . Нат. Мед . 21 (12): 1491–6. дои : 10.1038/нм.3968 . ПМЦ 4886303 . ПМИД 26552009 .
- ^ Миллер Р.Э., Бро Р., Байрами И., Уильямсон К.Т., МакДейд С., Кэмпбелл Дж. и др. (2016). «Синтетическое летальное воздействие на светлоклеточные опухоли яичников с мутацией ARID1A с помощью дазатиниба» (PDF) . Мол. Рак Там . 15 (7): 1472–84. дои : 10.1158/1535-7163.MCT-15-0554 . ПМИД 27364904 .
- ^ Кэлин В.Г. (2005). «Концепция синтетической летальности в контексте противораковой терапии». Обзоры природы Рак . 5 (9): 689–698. дои : 10.1038/nrc1691 . ПМИД 16110319 . S2CID 3218512 .
- ^ О'Коннор М.Дж.; Мартин НМБ; Смит GCM (2007). «Таргетная терапия рака, основанная на ингибировании восстановления разрыва цепи ДНК». Онкоген . 26 (56): 7816–7824. дои : 10.1038/sj.onc.1210879 . ПМИД 18066095 . S2CID 33955861 .
- ^ Фармер Х., Маккейб Н., Лорд СиДжей, Тутт А.Н., Джонсон Д.А., Ричардсон Т.Б., Сантароса М., Диллон К.Дж., Хиксон И., Найтс С., Мартин Н.М., Джексон С.П., Смит Г.К., Эшворт А. (апрель 2005 г.). «Нацеливание на дефект репарации ДНК в мутантных клетках BRCA как терапевтическая стратегия». Природа . 434 (7035): 917–21. Бибкод : 2005Natur.434..917F . дои : 10.1038/nature03445 . ПМИД 15829967 . S2CID 4364706 .
- ^ Брайант Х.Э.; Шульц, Никлас; Томас, Хью Д.; Паркер, Каян М.; Флауэр, Дэн; Лопес, Елена; Кайл, Сюзанна; Мейт, Марк; Кертин, Никола Дж.; Хелледей, Томас; и др. (2005). «Специфическое уничтожение опухолей с дефицитом BRCA2 ингибиторами поли(АДФ-рибозо)-полимеразы». Природа . 434 (7035): 913–917. Бибкод : 2005Natur.434..913B . дои : 10.1038/nature03443 . ПМИД 15829966 . S2CID 4391043 .
- ^ Гундем Дж., Перес-Лламас К., Жене-Санс А., Кедзерска А., Ислам А., Деу-Понс Дж., Ферни С.Дж., Лопес-Бигас Н. (февраль 2010 г.). «IntOGen: интеграция и интеллектуальный анализ многомерных онкогеномных данных». Нат. Методы . 7 (2): 92–3. дои : 10.1038/nmeth0210-92 . hdl : 10230/28107 . ПМИД 20111033 . S2CID 205417208 .
- ^ Гонсалес-Перес А, Перес-Льямас С, Деу-Понс Х, Тамбореро Д, Шредер М.П., Джене-Санс А, Сантос А, Лопес-Бигас Н (ноябрь 2013 г.). «Мутации IntOGen идентифицируют причины рака в зависимости от типа опухоли» . Нат. Методы . 10 (11): 1081–2. дои : 10.1038/nmeth.2642 . ПМК 5758042 . ПМИД 24037244 .
- ^ Рубио-Перес С., Барабанщик Д., Шредер М.П., Антолин А.А., Деу-Понс Дж., Перес-Флеймс С., Местрес Дж., Гонсалес-Перес А., Лопес-Бигас Н. (март 2015 г.). «In silico назначение противораковых препаратов группам из 28 типов опухолей открывает возможности таргетирования» . Раковая клетка 27 (3): 382–96. дои : 10.1016/j.ccell.2015.02.007 . hdl : 10230/33093 . ПМИД 25759023 .
- ^ Блюм-Дженсен П., Хантер Т. (май 2001 г.). «Передача сигналов онкогенной киназы». Природа . 411 (6835): 355–65. Бибкод : 2001Natur.411..355B . дои : 10.1038/35077225 . ПМИД 11357143 . S2CID 4428819 .
- ^ Барделли А. и др. (2003). «Мутационный анализ тирозинового кинома при колоректальном раке». Наука . 300 (5621): 949. doi : 10.1126/science.1082596 . ПМИД 12738854 . S2CID 85934154 .
- ^ Сэмюэлс Ю , Ван З, Барделли А, Силлиман Н, Птак Дж, Сабо С, Ян Х, Газдар А, Пауэлл С.М., Риггинс Г.Дж., Уилсон Дж.К., Марковиц С., Кинцлер К.В., Фогельштейн Б., Велкулеску В.Е. (апрель 2004 г.). «Высокая частота мутаций гена PIK3CA при раке человека». Наука . 304 (5670): 554. doi : 10.1126/science.1096502 . ПМИД 15016963 . S2CID 10147415 .
- ^ Дэвис Х., Бигнелл Г.Р., Кокс С., Стивенс П., Эдкинс С., Клегг С. и др. (июнь 2002 г.). «Мутации гена BRAF при раке человека» (PDF) . Природа . 417 (6892): 949–54. Бибкод : 2002Natur.417..949D . дои : 10.1038/nature00766 . ПМИД 12068308 . S2CID 3071547 .
- ^ Дэнсон С.; Лориган П. (2005). «Улучшение результатов при распространенной злокачественной меланоме – обновленная информация о системной терапии». Наркотики . 65 (6): 733–743. дои : 10.2165/00003495-200565060-00002 . ПМИД 15819587 . S2CID 46969987 .
- ^ Jump up to: а б с д и Ю, Человек (2012). Соматические мутации митохондриальной ДНК при раке человека . Достижения клинической химии. Том. 57. С. 99–138. дои : 10.1016/B978-0-12-394384-2.00004-8 . ISBN 9780123943842 . ПМИД 22870588 .
- ^ Хунг, Вайоминг; Дж. К. Лин; Л. М. Ли; и др. (2008). «Тандемное дублирование/трипликация коррелирует с вариацией растяжения полицитозина в области D-петли митохондриальной ДНК человека» . Мутагенез . 23 (2): 137–142. дои : 10.1093/mutage/gen002 . ПМИД 18252697 .
- ^ Флисс, М.С.; Усадель, Х.; Кабальеро, OL; и др. (2000). «Легкое обнаружение мутаций митохондриальной ДНК в опухолях и жидкостях организма». Наука . 287 (5460): 2017–2019. Бибкод : 2000Sci...287.2017F . дои : 10.1126/science.287.5460.2017 . ПМИД 10720328 . S2CID 24438279 .
- ^ Jump up to: а б Дэни, Массачусетс; СУ Дэни; ИП Лима; и др. (2004). «Меньшее значение ΔmtDNA4977, чем обычно, в различных типах опухолей позволяет предположить, что раковые клетки практически свободны от этой мутации». Жене. Мол. Рез . 3 (3): 395–409. ПМИД 15614730 .
- ^ Йе, К.; ХО Шу; В. Вэнь; и др. (2008). «Количественный анализ делеции 4977 п.н. митохондриальной ДНК при спорадическом раке молочной железы и доброкачественных заболеваниях молочной железы» . Рак молочной железы Рез. Обращаться . 108 (3): 427–434. дои : 10.1007/s10549-007-9613-9 . ПМЦ 3836503 . ПМИД 17541740 .
- ^ Ценг, Л.М.; ПХ Инь; CW Чи; и др. (2006). «Мутации митохондриальной ДНК и истощение митохондриальной ДНК при раке молочной железы». Гены Хромосомы Рак . 45 (7): 629–638. дои : 10.1002/gcc.20326 . ПМИД 16568452 . S2CID 21181048 .
- ^ Чжу, В.; В. Цинь; П. Брэдли; А. Вессель; К. Л. Пакетт; Э. Р. Заутер (2005). «Мутации митохондриальной ДНК в тканях рака молочной железы и в совпадающей жидкости аспирата сосков» . Канцерогенез . 26 (1): 145–152. дои : 10.1093/carcin/bgh282 . ПМИД 15375011 .
- ^ Чжоу С, Качхап С, Сунь В, Ву Г, Чуанг А, Поэта Л, Грамбине Л, Митани СК, Чаттерджи А, Кох В, Вестра ВХ, Майтра А, Глейзер С, Кардуччи М, Сидрански Д, МакФейт Т, Верма А , Калифано Дж.А. (май 2007 г.). «Частота и фенотипические последствия мутаций митохондриальной ДНК при плоскоклеточном раке головы и шеи человека» . Учеб. Натл. акад. наук. США . 104 (18): 7540–5. Бибкод : 2007PNAS..104.7540Z . дои : 10.1073/pnas.0610818104 . ПМК 1863503 . ПМИД 17456604 .
- ^ Поэтш М., Петерсманн А., Лигниц Э., Кляйст Б. (март 2004 г.). «Связь между нестабильностью митохондриальной ДНК, большими делециями митохондриальной ДНК и ядерной микросателлитной нестабильностью при плоскоклеточном раке головы и шеи». Диаг. Мол. Патол . 13 (1): 26–32. дои : 10.1097/00019606-200403000-00005 . ПМИД 15163006 . S2CID 21271445 .
- ^ Тан, диджей; Дж. Чанг; В.Л. Чен; и др. (2004). «Соматические мутации митохондриальной ДНК при раке полости рта у тех, кто жует бетель». Энн. Н-Й Акад. Наука . 1011 (1): 310–316. Бибкод : 2004NYASA1011..310T . дои : 10.1196/анналы.1293.030 . ПМИД 15126307 . S2CID 19794015 .
- ^ Ли, ХК; Ш. Ли; Дж. К. Лин; КК Ву; Округ Колумбия Да; Ю. Х. Вэй (2004). «Соматические мутации в D-петле и снижение количества копий митохондриальной ДНК при гепатоцеллюлярной карциноме человека». Мутационные исследования . 547 (1–2): 71–78. дои : 10.1016/j.mrfmmm.2003.12.011 . ПМИД 15013701 .
- ^ Инь, PH; КК Ву; Дж. К. Лин; CW Чи; Ю. Х. Вэй; ХК Ли (2010). «Соматические мутации митохондриального генома при гепатоцеллюлярной карциноме». Митохондрия . 10 (2): 174–182. дои : 10.1016/j.mito.2009.12.147 . ПМИД 20006738 .
- ^ Тан, диджей; Дж. Чанг; ЛЛ Лю; и др. (2006). «Значение соматических мутаций и изменение содержания митохондриальной ДНК при раке пищевода» . БМК Рак . 6:93 . дои : 10.1186/1471-2407-6-93 . ПМЦ 1459869 . ПМИД 16620376 .
- ^ Кассауэй К., Хаббе Н., Маллендор М.Э., Карикари К.А., Майтра А., Фельдманн Г. (2006). «Мутации митохондриальной ДНК при раке поджелудочной железы». Int J Gastrointest Рак . 37 (2–3): 57–64. дои : 10.1007/s12029-007-0008-2 . ПМИД 17827523 . S2CID 9716204 .
- ^ Хунг Вайоминг, Ву CW, Инь П.Х., Чанг CJ, Ли А.Ф., Чи CW, Вэй Ю.Х., Ли ХК (март 2010 г.). «Соматические мутации в митохондриальном геноме и их потенциальная роль в прогрессировании рака желудка человека». Биохим. Биофиз. Акта . 1800 (3): 264–70. дои : 10.1016/j.bbagen.2009.06.006 . ПМИД 19527772 .
- ^ Ву CW, Инь П.Х., Хун Вайоминг, Ли А.Ф., Ли Ш., Чи CW, Вэй Ю.Х., Ли Х.К. (сентябрь 2005 г.). «Мутации митохондриальной ДНК и истощение митохондриальной ДНК при раке желудка». Гены Хромосомы Рак . 44 (1): 19–28. дои : 10.1002/gcc.20213 . ПМИД 15892105 . S2CID 11009518 .
- ^ Ю, Джей-Джей; Т. Ян (2010). «Влияние мутации мтДНК на степень злокачественности опухоли у больных раком простаты». Стареющий мужчина . 13 (3): 159–165. дои : 10.3109/13685530903536668 . ПМИД 20136572 . S2CID 22017823 .
- ^ Гомес-Заера, М.; Дж. Абриль; Л. Гонсалес; и др. (2006). «Идентификация вариантов последовательностей соматической и зародышевой митохондриальной ДНК у больных раком простаты». Мутационные исследования . 595 (1–2): 42–51. дои : 10.1016/j.mrfmmm.2005.10.012 . ПМИД 16472830 .
- ^ Людвиг Дж. А., Вайнштейн Дж. Н. (ноябрь 2005 г.). «Биомаркеры в определении стадии рака, прогнозе и выборе лечения» . Нат. Преподобный Рак . 5 (11): 845–56. дои : 10.1038/nrc1739 . ПМИД 16239904 . S2CID 25540232 .
- ^ Куриан (3 июля 2023 г.). «Генетическое тестирование зародышевой линии после диагностики рака» . ДЖАМА . 330 (1): 43–51. дои : 10.1001/jama.2023.9526 . ПМЦ 10242510 . ПМИД 37276540 . Проверено 4 июля 2023 г.