нуклеоид
Нуклеоид ) представляет (что означает ядро -подобный собой область неправильной формы внутри прокариотической клетки , которая содержит весь или большую часть генетического материала . [1] [2] [3] Хромосома , типичного прокариота имеет круглую форму а ее длина очень велика по сравнению с размерами клетки, поэтому ее необходимо уплотнить, чтобы она поместилась. В отличие от ядра эукариотической клетки оно не окружено ядерной мембраной . Вместо этого нуклеоид формируется путем конденсации и функционального расположения с помощью хромосомных архитектурных белков и молекул РНК , а также сверхспирализации ДНК . Длина генома широко варьируется (обычно не менее нескольких миллионов пар оснований), и клетка может содержать несколько его копий.
Пока еще не известна структура бактериального нуклеоида с высоким разрешением, однако ключевые особенности были исследованы на Escherichia coli как на модельном организме . У E. coli хромосомная ДНК в среднем отрицательно сверхспиральна и свернута в плектонемические петли , которые ограничены разными физическими областями и редко диффундируют друг в друга. Эти петли пространственно организуются в области размером с мегабазу, называемые макродоменами, внутри которых участки ДНК часто взаимодействуют, но между которыми взаимодействия редки. Конденсированная и пространственно организованная ДНК образует спиральный эллипсоид, который радиально ограничен внутри клетки. Трехмерная структура ДНК в нуклеоиде, по-видимому, варьируется в зависимости от условий и связана с экспрессией генов , поэтому архитектура нуклеоида и транскрипция генов тесно взаимозависимы, взаимно влияя друг на друга.
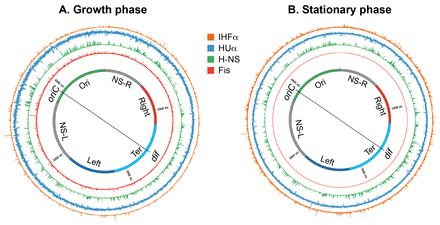
[[Файл: Нуклеоид Subhash 01.png|thumb|500px| Образование Escherichia coli нуклеоида A. Иллюстрация открытой конформации кольцевого генома Escherichia coli . Стрелки обозначают двунаправленную репликацию ДНК. генетическое положение начала двунаправленной репликации ДНК ( oriC ) и места декатенации хромосом ( dif ) в области терминации репликации ( ter Отмечено ). Цвета представляют собой определенные сегменты ДНК, как обсуждалось в CB . Иллюстрация случайной формы спирали, принятой чистой кольцевой ДНК Escherichia coli в состоянии теплового равновесия без суперспиралей и дополнительных стабилизирующих факторов. [4] [5] C. Карикатура на хромосому новорожденной клетки Escherichia coli . Геномная ДНК не только уплотнена в 1000 раз по сравнению с ее чистой случайной формой спирали, но также пространственно организована. oriC и dif локализованы в средней клетке, а определенные области ДНК, обозначенные цветом в A, организуются в пространственно различные домены.
Фон
[ редактировать ]У многих бактерий хромосома представляет собой одиночную ковалентно замкнутую (кольцевую) двухцепочечную молекулу ДНК, кодирующую генетическую информацию в гаплоидной форме. Размер ДНК варьируется от 500 000 до нескольких миллионов пар оснований (п.н.), кодирующих от 500 до нескольких тысяч генов в зависимости от организма. [2] Хромосомная ДНК присутствует в клетках в очень компактной, организованной форме, называемой нуклеоидом (что означает ядро ), которая не окружена ядерной мембраной , как в эукариотических клетках. [6] Выделенный нуклеоид содержит 80% ДНК, 10% белка и 10% РНК по массе. [7] [8]
Грамотрицательная бактерия Escherichia coli представляет собой модельную систему для исследования нуклеоида, посвященного тому, как хромосомная ДНК становится нуклеоидом, участвующим в этом факторам, что известно о ее структуре и как некоторые структурные аспекты ДНК влияют на экспрессию генов . [2] [3]
Есть два существенных аспекта формирования нуклеоида; конденсация большой ДНК в маленькое клеточное пространство и функциональная организация ДНК в трехмерной форме. Гаплоидная кольцевая хромосома E. coli состоит из ~ 4,6 x 10 6 б.п. Если ДНК расслаблена в форме B , ее окружность будет составлять ~1,5 миллиметра (0,332 нм x 4,6 x 10 6 ). Однако большая молекула ДНК, такая как хромосомная ДНК E. coli, не остается прямой жесткой молекулой в суспензии. [5] Броуновское движение приведет к искривлению и изгибам ДНК. Максимальная длина, до которой двухспиральная ДНК остается прямой, сопротивляясь изгибу, вызванному броуновским движением, составляет ~ 50 нм или 150 пар оснований, что называется длиной персистентности . Таким образом, чистая ДНК существенно конденсируется без каких-либо дополнительных факторов; при тепловом равновесии он принимает случайную форму клубка . [4] [5] Случайный клубок хромосомной ДНК E. coli будет занимать объем (4/3 π r 3 ) ~ 523 мкм 3 , рассчитанный по радиусу инерции ( R g = (√N a)/√6), где a — длина Куна (2 x персистентная длина), а N — количество сегментов длины Куна в ДНК (общая длина ДНК, разделенная на а ). [5] Хотя ДНК уже сконденсирована в виде случайного клубка, она еще не может принять объем нуклеоида менее микрона. Таким образом, присущих ДНК свойств недостаточно: дополнительные факторы должны способствовать дальнейшей конденсации ДНК порядка ~10 3 (объем случайного клубка, деленный на объем нуклеоида). Вторым важным аспектом образования нуклеоидов является функциональное расположение ДНК. Хромосомная ДНК не только конденсирована, но и функционально организована таким образом, который совместим с такими процессами транзакций ДНК, как репликация , рекомбинация , сегрегация и транскрипция . [9] [10] [11] Почти пять десятилетий исследований, начавшихся в 1971 году, [7] показал, что конечная форма нуклеоида возникает в результате иерархической организации ДНК. В наименьшем масштабе (1 т.п.н. или менее) архитектурные белки ДНК, связанные с нуклеоидами, конденсируют и организуют ДНК путем изгиба, образования петель, образования мостиков или обертывания ДНК. В более крупном масштабе (10 т.п.н. и более) ДНК образует плектонемические петли — сплетенную форму ДНК, возникающую в результате сверхспирализации. В мегабазовом масштабе плектонемические петли объединяются в шесть пространственно организованных доменов (макродоменов), которые определяются более частыми физическими взаимодействиями между участками ДНК внутри одного и того же макродомена, чем между разными макродоменами. [12] Длиннодействующие и короткодействующие связи ДНК-ДНК, образующиеся внутри и между макродоменами, способствуют конденсации и функциональной организации. Наконец, нуклеоид представляет собой спиральный эллипсоид с участками сильно конденсированной ДНК на продольной оси. [13] [14] [15]
Конденсация и организация
[ редактировать ]
Нуклеоид-ассоциированные белки (NAP)
[ редактировать ]У эукариот геномная ДНК конденсирована в виде повторяющегося массива ДНК-белковых частиц, называемых нуклеосомами . [16] [17] [18]
Нуклеосома состоит из ~146 п.н. ДНК, обернутой вокруг октамерного комплекса белков- гистонов . Хотя у бактерий нет гистонов, они обладают группой ДНК-связывающих белков, называемых нуклеоид-ассоциированными белками (NAP), которые функционально аналогичны гистонам в широком смысле. NAP очень распространены и составляют значительную часть белкового компонента нуклеоида. [19]
Отличительной характеристикой NAP является их способность связывать ДНК как специфическим (специфичным для последовательности или структуры), так и неспецифичным для последовательности образом. В результате NAP представляют собой белки с двойной функцией. [20] Специфическое связывание NAP в основном участвует в ген-специфической транскрипции , репликации ДНК , рекомбинации и репарации . [9] [10] [11] На пике их распространенности количество молекул многих NAP на несколько порядков превышает количество сайтов специфического связывания в геноме. [20] Таким образом, предполагается, что NAP связываются с хромосомной ДНК в основном неспецифичным для последовательности способом, и именно этот способ имеет решающее значение для компактизации хромосом. Примечательно, что так называемое неспецифическое связывание NAP не может быть полностью случайным. Может существовать низкая специфичность последовательности и/или структурная специфичность из-за конформации ДНК, зависящей от последовательности, или конформации ДНК, созданной другими NAP. [18]
Хотя молекулярные механизмы того, как NAP конденсируют ДНК in vivo , недостаточно изучены, на основании обширных исследований in vitro выяснилось, что NAP участвуют в уплотнении хромосом посредством следующих механизмов: NAP вызывают и стабилизируют изгибы ДНК, тем самым помогая конденсации ДНК за счет уменьшения длина персистентности. [20] NAP конденсируют ДНК путем образования мостиков, обертывания и группирования, которые могут возникать между близлежащими сегментами ДНК или удаленными сегментами ДНК хромосомы. Другой механизм, с помощью которого NAP участвуют в уплотнении хромосом, заключается в ограничении отрицательных суперспиралей в ДНК, что способствует топологической организации хромосомы. [20]
У E. coli идентифицировано не менее 12 NAP , [20] наиболее изученными из них являются HU, IHF, H-NS и Fis. Их распространенность, свойства связывания ДНК, а также влияние на конденсацию и организацию ДНК суммированы в таблицах ниже. [20]
| Белок | Молекулярная масса (кДа) | Родной функциональный блок | Избыток 1 в фазе роста | Избыток 1 в стационарной фазе |
|---|---|---|---|---|
| HUα и HUβ | ~ 9 | Гомо- и гетеродимеры | 55,000 (23) | 30,000 (12.5) |
| IHFα и IHFβ | ~ 11 | гетеродимер | 12,000 (5) | 55,000 (23) |
| Х-НС | ~ 15 | гомодимер | 20,000 (8) | 15,000 (6) |
| Фис | ~ 11 | гомодимер | 60,000 (25) | Необнаружимый |
| Дпс | ~ 19 | Додекамера | 6,000 (0.4) | 180,000 (12.5) |
1 Данные о численности (молекулы/клетку) были взяты из; [21] Число в скобках представляет собой микромолярную концентрацию, рассчитанную по следующей формуле: (количество нативных функциональных единиц/число Авогадро) x (1/объем клетки в литрах) x 10. 3 . Объем ячейки в литрах (2 х 10 −15 ) определяли, предполагая, что объем клетки E. coli составляет 2 мкм. 3 . [21]
| Белок | Обвязочный мотив | Специфическое сродство связывания ДНК 1 | Случайная аффинность связывания ДНК 1 |
|---|---|---|---|
| ХУ | Структурный мотив, определяемый изгибами и изломами ДНК. [22] [23] | 7,5 х 10 −9 [24] | 4,0 х 10 −7 [24] |
| ИХФ | ВАТКААНННТТР [25] | 1,5 х 10 −9 [26] | 1,7 х 10 −6 [26] |
| Х-НС | TCGATAAATT [27] | 10-15 х 10 −9 [28] | 6 х 10 −8 [28] |
| Фис | ГНТЯААВТТРАНК [29] | 0,2-1,0 х 10 −9 [29] [30] | >8,0 х 10 −6 [30] |
| Дпс | без даты | без даты | 1,65 х 10 −7 [31] |
| МатП | GTGACRNYGTCAC [32] | 8,0 х 10 −9 | без даты |
| МукБЭФ | без даты | без даты | без даты |
1 Сродство связывания относится к равновесной константе диссоциации (Kd) в молярных единицах (М). НД = не определено
ХУ
[ редактировать ]Гистоноподобный белок штамма E. coli U93 (HU) является эволюционно консервативным белком бактерий. [33] [34] HU существует в E. coli в виде гомо- и гетеродимеров двух субъединиц HUα и HUβ, имеющих 69% аминокислотной идентичности. [35] Хотя его называют гистоноподобным белком, близкими функциональными родственниками HU у эукариот являются белки группы высокой подвижности (HMG), а не гистоны. [36] [37] HU представляет собой неспецифический ДНК-связывающий белок. Он связывается с низким сродством к любой линейной ДНК. Однако он преимущественно связывается с высоким сродством со структурно искаженной ДНК. [38] [39] [40] [41] [42] [24] Примеры искаженных ДНК-субстратов включают крестообразную ДНК , выпуклую ДНК, дцДНК, содержащую одноцепочечные разрывы, такие как разрывы , разрывы или развилки . Более того, HU специфически связывает и стабилизирует петлю ДНК, опосредованную белком. [43] В структурно-специфичном режиме связывания ДНК HU распознает общий структурный мотив, определяемый изгибами или изломами, возникающими в результате искажения. [22] [44] [23] тогда как он связывается с линейной ДНК, блокируя фосфатный остов. [45] Хотя структурно-специфическое связывание с высоким сродством необходимо для специализированных функций HU, таких как сайт-специфическая рекомбинация , репарация ДНК , инициация репликации ДНК и регуляция генов, [9] [10] [11] похоже, что общее связывание с низким сродством участвует в конденсации ДНК. [45] При иммунопреципитации хроматина в сочетании с секвенированием ДНК ( ChIP-Seq ) HU не выявляет каких-либо специфических событий связывания. [46] Вместо этого он демонстрирует равномерное связывание по всему геному, что, по-видимому, отражает его в основном слабое, неспецифическое связывание, не специфичное для последовательности, таким образом маскируя высокоаффинное связывание in vivo . [46]
У штаммов, лишенных HU, нуклеоид «деконденсирован», что соответствует роли HU в уплотнении ДНК. [47] Следующие исследования in vitro предполагают возможные механизмы того, как HU может конденсировать и организовывать ДНК in vivo . HU не только стабильно связывается с искаженной ДНК с изгибами, но и индуцирует гибкие изгибы даже в линейной ДНК при концентрации менее 100 нМ. Напротив, HU демонстрирует противоположный архитектурный эффект на ДНК при более высоких физиологически значимых концентрациях. [45] [9] [10] [11] [47] [48] Он образует жесткие нуклеопротеиновые нити, вызывающие выпрямление ДНК, а не изгиб. Филаменты могут дополнительно образовывать сеть ДНК (группировку ДНК), расширяемую как латерально, так и медиально из-за мультимеризации HU-HU, запускаемой неспецифичным для последовательности связыванием ДНК. [45]
Насколько такое поведение HU актуально внутри клетки? Для формирования нитей требуется связывание HU с ДНК с высокой плотностью: один димер HU на ДНК длиной 9–20 п.н. Но на каждые ~150 п.о. хромосомной ДНК приходится только один димер HU, исходя из предполагаемого содержания 30 000 димеров HU на клетку (4600 000 п.н./30 000). [21] Это указывает на то, что гибкие изгибы с большей вероятностью происходят in vivo . Гибкий изгиб может вызвать конденсацию из-за уменьшения персистентной длины ДНК, как показали эксперименты с магнитным пинцетом , которые позволяют изучать конденсацию одной молекулы ДНК с помощью ДНК-связывающего белка. [48] [49] Однако из-за кооперативности в некоторых участках хромосомы могут образовываться жесткие нити и сети. Образование нитей само по себе не вызывает конденсацию. [48] но сплетение или группирование ДНК может существенно способствовать конденсации, объединяя отдаленные или близлежащие сегменты хромосом. [45]
ИХФ
[ редактировать ]Фактор хозяина интеграции (IHF) структурно практически идентичен HU. [51] но во многих аспектах ведет себя иначе, чем HU. В отличие от HU, который преимущественно связывается со структурным мотивом независимо от последовательности, IHF преимущественно связывается со специфической последовательностью ДНК, хотя специфичность возникает за счет зависимой от последовательности структуры ДНК и ее деформируемости. Специфическое связывание IHF в родственных сайтах резко изгибает ДНК более чем на 160 градусов. [51] Встречаемость мотива родственной последовательности составляет около 3000 в геноме E. coli . [46] Предполагаемое количество IHF в фазе роста составляет около 6000 димеров на клетку. Если предположить, что один димер IHF связывается с одним мотивом, а нуклеоид содержит более одного геномного эквивалента во время фазы экспоненциального роста, большинство молекул IHF будут занимать определенные участки в геноме и, вероятно, только конденсируют ДНК, вызывая резкий изгиб. [46]
Помимо преимущественного связывания со специфической последовательностью ДНК, IHF также связывается с ДНК неспецифичным для последовательности образом со сродством, аналогичным HU. Роль неспецифического связывания IHF в конденсации ДНК, по-видимому, имеет решающее значение в стационарной фазе, поскольку содержание IHF увеличивается в пять раз в стационарной фазе, а дополнительные димеры IHF, вероятно, будут неспецифически связывать хромосомную ДНК. [21] [52] [53] В отличие от HU, IHF не образует толстых жестких нитей при более высоких концентрациях. Вместо этого его неспецифическое связывание также вызывает изгиб ДНК, хотя степень изгиба намного меньше, чем в специфических сайтах, и аналогична гибкому изгибу, индуцированному HU в линейной ДНК при низких концентрациях. [54] In vitro изгиб, вызванный неспецифическим связыванием IHF, может вызывать конденсацию ДНК и способствовать образованию нуклеопротеиновых комплексов более высокого порядка в зависимости от концентрации хлорида калия и хлорида магния. [54] Организация ДНК высшего порядка с помощью IHF in vivo пока неясна. [54]
Х-НС
[ редактировать ]Отличительная черта гистоноподобного или термостабильного нуклеоидного структурирующего белка (H-NS). [55] [56] [57] [58] от других НАП является способность переключаться с гомодимерной формы при относительно низких концентрациях (<1 х 10 −5 М) в олигомерное состояние на более высоких уровнях. [59] [60] Из-за свойств олигомеризации H-NS распространяется латерально вдоль богатой АТ ДНК в реакции нуклеации , где сайты с высоким сродством действуют как центры нуклеации. [61] [62] [28] Распространение H-NS на ДНК приводит к двум противоположным результатам в зависимости от концентрации магния в реакции. При низкой концентрации магния (< 2 мМ) H-NS образует жесткие нуклеопротеиновые нити, тогда как при более высоких концентрациях магния (> 5 мМ) он образует меж- и внутримолекулярные мостики. [63] [64] [65] [66] [67] Образование жестких нитей приводит к выпрямлению ДНК без конденсации, тогда как образование мостиков вызывает значительное сворачивание ДНК. [66] Анализ связывания H-NS в геноме с помощью анализа ChIP-Seq предоставил косвенные доказательства распространения H-NS по ДНК in vivo . H-NS избирательно связывается с 458 участками генома. [50] Хотя было продемонстрировано, что H-NS предпочитает изогнутую ДНК, образованную повторяющимися A-треками в последовательностях ДНК. [61] [68] В основе селективного связывания лежит наличие мотива консервативной последовательности, обнаруженного в богатых АТ регионах. [27] Что еще более важно, частое появление мотива последовательности в области связывания H-NS, который может усиливать кооперативные белок-белковые взаимодействия, и необычно большая длина области связывания согласуются с распространением белка. Преобладает ли in vivo образование нитей или образование мостиков ДНК , зависит от физиологической концентрации магния внутри клетки. [66] [69] Если концентрация магния равномерно низкая (<5 мМ), H-NS будет образовывать жесткие нуклеопротеиновые нити in vivo . [66] С другой стороны, если существует неравномерное распределение магния в клетке, это может способствовать как образованию мостиков ДНК, так и повышению жесткости, но в разных областях нуклеоида. [66]
Более того, H-NS наиболее известен как глобальный сайленсер генов, который преимущественно ингибирует транскрипцию горизонтально переносимых генов, и именно жесткая нить приводит к молчанию генов. [70] [71] В совокупности оказывается, что образование жестких филаментов является наиболее вероятным результатом взаимодействий H-NS-ДНК in vivo , что приводит к молчанию генов, но не индуцирует конденсацию ДНК. Соответственно, отсутствие H-NS не меняет объем нуклеоида. [72] Однако возможно, что в некоторых условиях окружающей среды E. coli испытывает высокую концентрацию магния. В таких условиях H-NS может переключаться с формы, индуцирующей филаменты, на форму, индуцирующую мостик, которая способствует конденсации и организации ДНК. [66]
Фис
[ редактировать ]Фактор стимуляции инверсии (Fis) представляет собой ДНК-связывающий белок, специфичный для последовательности, который связывается со специфическими последовательностями ДНК, содержащими симметричный мотив длиной 15 пар оснований. [29] [30] [73] Как и IHF, Fis индуцирует изгиб ДНК в родственных сайтах. Способность изгибать ДНК проявляется в структуре гомодимера Fis. Гомодимер Fis обладает двумя мотивами спираль-поворот-спираль (HTH), по одному от каждого мономера. Мотив HTH обычно распознает большую бороздку ДНК. Однако расстояние между спиралями распознавания ДНК двух мотивов HTH в гомодимере Fis составляет 25 Å , что на ~ 8 Å короче, чем шаг канонической B-ДНК , что указывает на то, что белок должен изгибать или скручивать ДНК для стабильного связывания. . [74] [75] Соответственно, кристаллическая структура комплексов Fis-ДНК показывает, что расстояние между спиралями узнавания остается неизменным, тогда как изгиб ДНК находится в диапазоне 60-75 градусов. [30] Существует 1464 области связывания Fis, распределенных по геному E. coli , и мотив связывания, идентифицированный компьютерным путем, соответствует известному мотиву длиной 15 п.н. [50] [76] Специфическое связывание Fis в таких сайтах будет вызывать изгибы ДНК и, таким образом, способствовать конденсации ДНК за счет уменьшения длины персистенции ДНК. Более того, многие сайты связывания Fis встречаются последовательно, например, в стабильных промоторах РНК, например, в промоторе P1 рРНК оперона rrnB . Когерентное изгибание Fis в тандемных участках, вероятно, создаст микропетлю ДНК, которая может в дальнейшем способствовать конденсации ДНК. [77]
Помимо высокоаффинного специфического связывания с родственными сайтами, Fis может связываться со случайной последовательностью ДНК. Неспецифическое связывание ДНК имеет важное значение, поскольку Fis в фазе роста так же распространен, как и HU . Следовательно, ожидается, что большинство молекул Fis будут связываться с ДНК неспецифичным для последовательности образом. Эксперименты с магнитным пинцетом показывают, что это неспецифическое связывание Fis может способствовать конденсации и организации ДНК. [78] [79] Fis вызывает легкую конденсацию одиночной молекулы ДНК при <1 мМ, но индуцирует существенную складку за счет образования петель ДНК среднего размера ~ 800 п.н. при >1 мМ. Петли в экспериментах с магнитным пинцетом отличаются от микропетлей, созданных в результате когерентного изгиба ДНК в родственных участках, поскольку они требуют образования комплексов ДНК-белок высокой плотности, достигаемых путем независимого от последовательности связывания. Хотя появление таких петель in vivo еще предстоит продемонстрировать, связывание Fis с высокой плотностью может происходить in vivo за счет согласованного действия как специфического, так и неспецифического связывания. Тандемное появление специфических сайтов может инициировать реакцию нуклеации, подобную реакции нуклеации H-NS, а затем неспецифическое связывание может привести к образованию локализованных массивов Fis высокой плотности. Мосты между этими локализованными областями могут создавать большие петли ДНК. [79] Fis присутствует исключительно в фазе роста , а не в стационарной фазе . [80] [81] Т.о., любая роль Fis в хромосомной конденсации должна быть специфична для растущих клеток. [81]
Нуклеоид-ассоциированные РНК (нРНК)
[ редактировать ]Ранние исследования по изучению влияния обработки РНКазой А на изолированные нуклеоиды показали, что РНК участвует в стабилизации нуклеоида в конденсированном состоянии. [82] Более того, обработка РНКазой А разрушала волокна ДНК на более тонкие волокна, как это наблюдалось с помощью атомно-силовой микроскопии нуклеоида с использованием «процедуры лизиса на подложке». [83] Эти результаты продемонстрировали участие РНК в структуре нуклеоида, но идентичность молекулы РНК до недавнего времени оставалась неизвестной. [47] Большинство исследований HU были сосредоточены на его связывании с ДНК. [83] Однако HU также связывается с дцРНК и гибридами РНК-ДНК с более низким сродством, как и с линейной дцДНК. [84] Более того, HU преимущественно связывается с РНК, содержащими вторичные структуры, и с гибридом РНК-ДНК, в котором РНК содержит разрыв или выступ. [84] [85] Сродство связывания HU с этими РНК-субстратами аналогично тому, с которым он связывается с искаженной ДНК. Иммунопреципитация HU-связанной РНК в сочетании с обратной транскрипцией и исследованием на микрочипах (RIP-Chip), а также анализ РНК из очищенных интактных нуклеоидов выявили молекулы РНК, связанные с нуклеоидами, которые взаимодействуют с HU. [47] Некоторые из них являются некодирующими РНК, и одна такая РНК, названная naRNA4 (нуклеоид-ассоциированная РНК 4), кодируется повторяющимся экстрагенным палиндромом ( REP325 ). У штамма, лишенного REP325 , нуклеоид деконденсирован, как и у штамма, лишенного HU. [47] naRNA4, скорее всего, участвует в конденсации ДНК, соединяя сегменты ДНК в присутствии HU. [86] Недавние исследования дают представление о молекулярном механизме того, как naRNA4 устанавливает связи ДНК-ДНК. РНК нацелена на области ДНК, содержащие крестообразные структуры, и образует комплекс РНК-ДНК, который имеет решающее значение для установления связей ДНК-ДНК. [87] Удивительно, но хотя HU помогает в образовании комплекса, он не присутствует в конечном комплексе, что указывает на его потенциальную роль катализатора (шаперона). Природа комплекса РНК-ДНК остается загадочной, поскольку образование комплекса не требует обширного спаривания оснований Уотсона/Крика, но чувствительно к РНКазе H, которая расщепляет РНК в гибриде РНК-ДНК, и комплекс связывается с антителом, специфичным для Гибриды РНК-ДНК. [47] [83] [84]
Суперспирализация
[ редактировать ]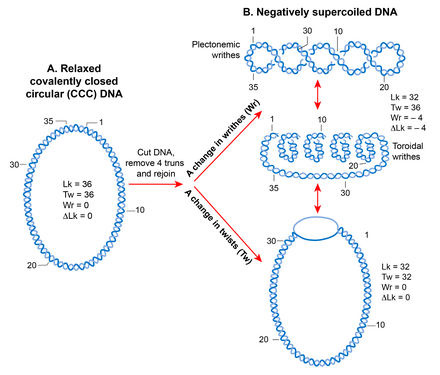
Из-за своей спиральной структуры двухцепочечная молекула ДНК становится топологически ограниченной в ковалентно замкнутой кольцевой форме, что исключает вращение свободных концов. [88] Количество раз, когда две цепи пересекают друг друга в топологически ограниченной ДНК, называется числом связывания (Lk), которое эквивалентно количеству спиральных витков или скручиваний в кольцевой молекуле. [89] Lk топологической ДНК остается инвариантной, независимо от того, как деформируется молекула ДНК, пока ни одна цепь не разорвана. [90] [91]
Lk ДНК в релаксированной форме определяется как Lk 0 . Для любой ДНК Lk 0 можно рассчитать, разделив длину (в п.о.) ДНК на количество п.н. на один виток спирали. Это соответствует 10,4 п.н. для релаксированной B-формы ДНК . Любое отклонение от Lk 0 вызывает сверхспирализацию ДНК. Уменьшение числа связывания (Lk<Lk 0 ) создает отрицательную сверхспирализацию, тогда как увеличение числа связывания (Lk>Lk 0 ) создает положительную сверхспирализацию. [92] [90]
Состояние сверхспирализации (когда Lk не равно Lk 0 ) приводит к переходу в структуре ДНК, который может проявляться в изменении количества витков (отрицательное <10,4 п.н./виток, положительное >10,4 п.н. за оборот) и/или в образование корчей , называемых суперспиралами. Таким образом, Lk математически определяется как знакозависимая сумма двух геометрических параметров: скручивания и скручивания. Количественной мерой сверхспирализации, не зависящей от размера молекул ДНК, является плотность суперспирализации (σ), где σ =∆Lk/Lk 0 . [91]
Писцы могут принимать две структуры; плектонема и соленоид или тороид. Плектонемическая структура возникает в результате переплетения винтовых осей. Тороидальные суперспирали возникают, когда ДНК образует несколько спиралей, расположенных вокруг оси и не пересекающихся друг с другом, как в телефонном шнуре. [90] Корчи в форме плектонем являются правосторонними и левосторонними в положительно или отрицательно сверхспиральной ДНК соответственно. Ориентальность тороидальных суперспиралей противоположна таковой у плектонем. И плектонемы, и тороидальные суперспирали могут находиться как в свободной форме, так и в связанной форме с белками. Лучшим примером связанной тороидальной суперспирализации в биологии является эукариотическая нуклеосома , в которой ДНК обертывается вокруг гистонов . [17]
Плектонемические суперспирали у E. coli
[ редактировать ]
У большинства бактерий ДНК присутствует в сверхспиральной форме. Круговая природа хромосомы E. coli делает ее топологически ограниченной молекулой, которая в основном имеет отрицательную сверхспираль с расчетной средней плотностью сверхспирализации (σ) -0,05. [93] В хроматине эукариот ДНК находится преимущественно в тороидальной форме, которая ограничена и определяется гистонами посредством образования нуклеосом. Напротив, в нуклеоиде E. coli около половины хромосомной ДНК организовано в виде свободных плектонемических суперспиралей. [94] [95] [96] Оставшаяся ДНК удерживается либо в плектонемической форме, либо в альтернативных формах, включая, помимо прочего, тороидальную форму, за счет взаимодействия с белками, такими как NAP. Таким образом, плектонемические суперспирали представляют собой эффективную суперспираль генома E. coli , отвечающую за его конденсацию и организацию. И плектонемическая, и тороидальная суперспирализация способствуют конденсации ДНК. Примечательно, что из-за разветвленности плектонемических структур она обеспечивает меньшую конденсацию ДНК, чем тороидальная структура. Молекула ДНК одинакового размера с одинаковой плотностью суперспирализации более компактна в тороидальной форме, чем в плектонемической форме. Помимо конденсации ДНК, сверхспирализация способствует организации ДНК. Он способствует распутыванию ДНК, уменьшая вероятность сцепления. [97] Суперспирализация также помогает сблизить два удаленных участка ДНК, тем самым способствуя потенциальному функциональному взаимодействию между различными сегментами ДНК. [91]
Источники сверхспирализации E. coli
[ редактировать ]Три фактора способствуют созданию и поддержанию сверхспирализации хромосомной ДНК в E. coli : (i) активность топоизомераз , (ii) акт транскрипции и (iii) NAP. [95]
Топоизомеразы
[ редактировать ]Топоизомеразы представляют собой особую категорию метаболических ферментов ДНК, которые создают или устраняют сверхспирализацию путем разрыва, а затем повторного лигирования нитей ДНК. [98] E. coli обладает четырьмя топоизомеразами. ДНК-гираза вызывает отрицательную сверхспирализацию в присутствии АТФ и устраняет положительную сверхспирализацию в отсутствие АТФ. [99] Во всех формах жизни ДНК-гираза является единственной топоизомеразой, которая может создавать отрицательную суперспираль, и именно из-за этой уникальной способности бактериальные геномы обладают свободными отрицательными суперспиралями; ДНК-гираза обнаружена у всех бактерий, но отсутствует у высших эукариот. Напротив, Topo I противостоит ДНК-гиразе, расслабляя отрицательно сверхспирализованную ДНК. [100] [101] Существуют генетические данные, позволяющие предположить, что баланс между противоположными активностями ДНК-гиразы и Topo I ответственен за поддержание устойчивого уровня средней отрицательной сверхспиральности в E. coli . [100] [102] Оба фермента необходимы для выживания E. coli . Нулевой штамм topA , гена, кодирующего Topo I, выживает только благодаря наличию супрессорных мутаций в генах, кодирующих ДНК-гиразу. [100] [102] Эти мутации приводят к снижению активности гиразы, что позволяет предположить, что избыточная отрицательная суперспирализация из-за отсутствия Topo I компенсируется снижением активности отрицательной суперспирализации ДНК-гиразы. Topo III необязателен для E. coli , и неизвестно, играет ли он какую-либо роль в суперспирализации E. coli. [103] Основная функция Topo IV — разделение сестринских хромосом. Однако было показано, что он также способствует устойчивому уровню отрицательной сверхспирализации за счет ослабления отрицательной сверхспирализации вместе с Топо I. [104] [105]
| топоизомераза | Тип | Функция | Одно- или двухцепочечное расщепление |
|---|---|---|---|
| Топоизомераза I | Я | Удаляет (-) суперспирализацию | SS |
| Топоизомераза III | Я | Удаляет (-) суперспирализацию | SS |
| Топоизомераза IV | ИМА | Удаляет (-) суперспирализацию | ДС |
| ДНК-гираза | ИМА | Создает (-) суперспирализацию и удаляет (+) суперспирализацию. | ДС |
Транскрипция
[ редактировать ]Модель двойного суперспирального домена, предложенная Лю и Вангом, утверждала, что раскручивание двойной спирали ДНК во время транскрипции вызывает сверхспирализацию ДНК, как показано на рисунке . [106] Согласно их модели, транскрибирующая РНК-полимераза (РНКП), скользящая по ДНК, заставляет ДНК вращаться вокруг своей спиральной оси. Препятствие свободному вращению ДНК может возникнуть из-за топологического ограничения, в результате чего ДНК перед РНКП станет чрезмерно перекрученной (положительно сверхспиральной), а ДНК позади РНКП станет недостаточно перекрученной (отрицательно сверхспиральной). Было обнаружено, что топологическое ограничение не требуется, поскольку RNAP генерирует достаточный крутящий момент, который вызывает суперспирализацию даже в линейной матрице ДНК. [107] Если ДНК уже имеет отрицательную суперспираль, это действие расслабляет существующие отрицательные суперспирали, прежде чем вызвать накопление положительных суперспиралей перед РНКП и вводить больше отрицательных суперспиралей позади РНКП. В принципе, ДНК-гираза и Топо I должны удалять избыток положительных и отрицательных суперспиралей соответственно, но если скорость элонгации РНКП превышает оборот двух ферментов, транскрипция способствует устойчивому уровню суперспирализации. [107]
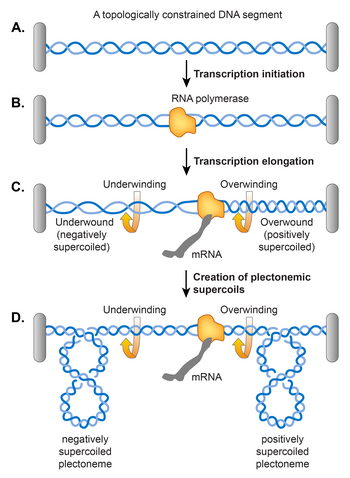
Контроль сверхспирализации с помощью NAP
[ редактировать ]В хроматине эукариот ДНК редко присутствует в свободной сверхспиральной форме, поскольку нуклеосомы сдерживают почти всю отрицательную суперспирализацию за счет прочного связывания ДНК с гистонами. Аналогично, в E. coli нуклеопротеиновые комплексы, образованные NAP, сдерживают половину плотности суперспирализации нуклеоида. [93] [96] Другими словами, если NAP диссоциирует от нуклеопротеинового комплекса , ДНК примет свободную плектонемическую форму. Экспериментально было показано, что связывание ДНК HU, Fis и H-NS сдерживает отрицательную сверхспирализацию в расслабленной, но топологически ограниченной ДНК. [108] [109] [110] [111] [112] Они могут сделать это либо путем изменения шага спирали ДНК, либо путем создания тороидальных извиваний путем изгибания и свертывания ДНК. Альтернативно, NAP могут преимущественно связываться и стабилизировать другие формы внутренней ДНК, такие как крестообразные структуры и разветвленные плектонемы. Сообщалось, что Fis организует разветвленные плектонемы посредством связывания с кроссоверными областями, а HU преимущественно связывается с крестообразными структурами. [112]
NAP также косвенно регулируют сверхспирализацию ДНК. Fis может модулировать суперспирализацию, подавляя транскрипцию генов, кодирующих ДНК-гиразу. [113] Существуют генетические данные, позволяющие предположить, что HU контролирует уровень суперспирализации путем стимуляции ДНК-гиразы и снижения активности Topo I. [114] [115] В подтверждение генетических исследований было показано, что HU стимулирует декатенацию ДНК, катализируемую ДНК-гиразой in vitro . [116] Механически неясно, как HU модулирует активность гиразы и Topo I. HU может физически взаимодействовать с ДНК-гиразой и Topo I, или деятельность HU по организации ДНК, такая как изгиб ДНК, может облегчать или ингибировать действие ДНК-гиразы и Topo I соответственно. [114] [116]
Плектонемические суперспирали организуются в несколько топологических доменов.
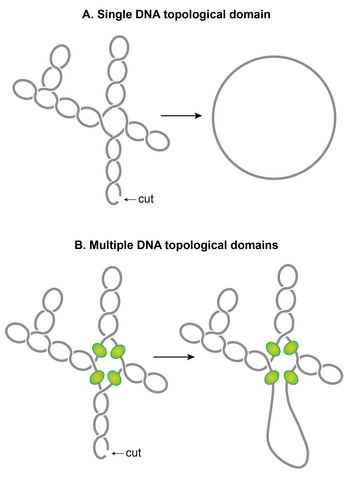
[ редактировать ]Одной из поразительных особенностей нуклеоида является то, что плектонемические суперспирали организованы в несколько топологических доменов. [117] Другими словами, единственный разрез в одном домене ослабит только этот домен, а не другие. Топологическая область образуется из-за барьера суперспирали-диффузии. Независимые исследования с использованием различных методов показали, что размер топологических доменов варьируется от 10 до 400 КБ. [95] [117] [118] Случайное размещение барьеров, обычно наблюдаемое в этих исследованиях, по-видимому, объясняет широкую вариабельность размеров доменов. [117]
Хотя идентичность доменных барьеров еще предстоит установить, возможные механизмы, ответственные за формирование барьеров, включают: (i) Доменный барьер может формироваться, когда белок, способный сдерживать суперспирали, одновременно связывается с двумя разными участками на хромосоме, образуя топологически изолированная петля или домен ДНК. Экспериментально было продемонстрировано, что опосредованное белком петлеобразование в суперспиральной ДНК может создавать топологический домен. [119] [120] NAP, такие как H-NS и Fis, являются потенциальными кандидатами, основываясь на их способностях к образованию петель ДНК и распределении их сайтов связывания. (ii) Элементы бактериальной мозаики (BIME) также являются потенциальными кандидатами на роль доменных барьеров. BIME — это палиндромные повторяющиеся последовательности, которые обычно встречаются между генами. Было показано, что BIME препятствует диффузии суперспирализации в синтетически созданной топологической кассете, встроенной в хромосому E. coli . [121] По геному распределено около 600 BIME, что, возможно, разделяет хромосому на 600 топологических доменов. [122] (iii) Барьеры также могут возникать в результате прикрепления ДНК к клеточной мембране посредством белка, который связывается как с ДНК, так и с мембраной, или в результате зарождающейся транскрипции и трансляции белков, закрепленных на мембране. (iv) Транскрипционная активность может создавать барьеры суперспирализации-диффузии. Было показано, что активно транскрибирующая РНКП блокирует диссипацию плектонемических суперспиралей, тем самым формируя диффузионно-суперспиральный барьер. [123] [124] [125]
Динамика нуклеоида, зависящая от фазы роста
[ редактировать ]Нуклеоид реорганизуется в клетках стационарной фазы, что позволяет предположить, что структура нуклеоида очень динамична и определяется физиологическим состоянием клеток. Сравнение карт контактов нуклеоида с высоким разрешением показало, что дальние контакты в макродомене Ter увеличились в стационарной фазе по сравнению с фазой роста . [126] Кроме того, границы CID в стационарной фазе отличались от границ, обнаруженных в фазе роста. Наконец, морфология нуклеоида претерпевает массивную трансформацию во время длительной стационарной фазы; [127] нуклеоид имеет упорядоченную тороидальную структуру. [128]
Специфические изменения в структуре нуклеоида в фазе роста могут быть вызваны изменением уровней архитектурных белков ДНК, связанных с нуклеоидами (NAP и субъединицы Muk), суперспирализацией и транскрипционной активностью. Содержание NAP и субъединиц Muk меняется в зависимости от цикла роста бактерий. Fis и индуцированный голоданием ДНК-связывающий белок Dps, еще один NAP, почти исключительно присутствуют в фазе роста и стационарной фазе соответственно. Уровни Fis повышаются при входе в экспоненциальную фазу, а затем быстро снижаются, пока клетки все еще находятся в экспоненциальной фазе, достигая уровней, которые невозможно обнаружить в стационарной фазе. [129] В то время как уровни Fis начинают снижаться, уровни Dps начинают расти и достигают максимума в стационарной фазе. [21] Резкий переход в структуре нуклеоида, наблюдаемый в длительной стационарной фазе, в основном объясняется Dps. Он образует ДНК/ кристаллические сборки, которые защищают нуклеоид от агентов, повреждающих ДНК, присутствующих во время голодания. [128]
HU, IHF и H-NS присутствуют как в фазе роста, так и в стационарной фазе. [21] Однако их численность значительно меняется, так что HU и Fis являются наиболее распространенными NAP в фазе роста, тогда как IHF и Dps становятся наиболее распространенными NAP в стационарной фазе. [21] HUαα является преобладающей формой на ранней экспоненциальной фазе, тогда как гетеродимерная форма преобладает в стационарной фазе с небольшим количеством гомодимеров. [130] Этот переход имеет функциональные последствия в отношении структуры нуклеоида, поскольку две формы, по-видимому, по-разному организуют и конденсируют ДНК; как гомо-, так и гетеродимеры образуют нити, но только гомодимер может объединять несколько сегментов ДНК, образуя сеть ДНК. [45] Число копий MukB увеличивается в два раза в стационарной фазе. [131] [132] Увеличение количества молекул MukB может влиять на процессивность комплекса MukBEF как фактора выдавливания петель ДНК, что приводит к увеличению или увеличению числа петель. [131] [132]
Суперспирализация может действовать согласованно с архитектурными белками ДНК, реорганизовывая нуклеоид. Общий уровень суперспирализации снижается в стационарной фазе, а на региональном уровне суперспирализация имеет иную картину. [133] Изменения в суперспирализации могут изменить топологическую организацию нуклеоида. Более того, поскольку хромосомная область с высокой транскрипционной активностью образует границу CID, изменения транскрипционной активности во время разных фаз роста могут изменять формирование границ CID и, следовательно, пространственную организацию нуклеоида. Возможно, изменения границ CID, наблюдаемые в стационарной фазе, могут быть связаны с высокой экспрессией другого набора генов в стационарной фазе по сравнению с фазой роста. [126]
Структура нуклеоида и экспрессия генов
[ редактировать ]NAP и экспрессия генов
[ редактировать ]Структура хромосом E. coli и экспрессия генов, по-видимому, взаимно влияют друг на друга. С одной стороны, корреляция границы CID с высокой транскрипционной активностью указывает на то, что организация хромосом регулируется транскрипцией. С другой стороны, трехмерная структура ДНК внутри нуклеоида на всех уровнях может быть связана с экспрессией генов. Во-первых, было показано, что реорганизация трехмерной архитектуры нуклеоида в E. coli может динамически модулировать паттерн клеточной транскрипции. [134] Мутант HUa сделал нуклеоид очень сильно конденсированным за счет повышенной положительной сверхспиральности хромосомной ДНК. Следовательно, многие гены были подавлены, а многие покоящиеся гены экспрессировались. Кроме того, существует множество конкретных случаев, когда белково-опосредованные локальные архитектурные изменения изменяют транскрипцию генов. Например, образование жестких нуклеопротеиновых нитей с помощью H-NS блокирует доступ РНКП к промотору, тем самым предотвращая транскрипцию гена. [135] Посредством подавления генов H-NS действует как глобальный репрессор, преимущественно ингибируя транскрипцию горизонтально переносимых генов. [50] [27] В другом примере специфическое связывание HU с опероном gal облегчает образование петли ДНК, которая поддерживает репрессию оперона gal в отсутствие индуктора. [136] Топологически отличная микропетля ДНК, созданная в результате когерентного изгиба ДНК Fis на стабильных промоторах РНК, активирует транскрипцию. [77] Изгиб ДНК с помощью IHF дифференциально контролирует транскрипцию двух тандемных промоторов оперона ilvGMEDA в E. coli . [137] [138] Специфические топологические изменения, вызванные NAP, не только регулируют транскрипцию генов, но также участвуют в других процессах, таких как инициация репликации ДНК, рекомбинация и транспозиция. [9] [10] [11] В отличие от специфической регуляции генов, еще предстоит выяснить, как структура хромосом более высокого порядка и ее динамика влияют на глобальную экспрессию генов на молекулярном уровне. [139]
Сверхспирализация ДНК и экспрессия генов
[ редактировать ]Между суперспирализацией ДНК и транскрипцией генов существует двусторонняя взаимосвязь. [139] Отрицательная сверхспирализация промоторной области может стимулировать транскрипцию, облегчая плавление промотора и увеличивая аффинность связывания ДНК белкового регулятора. Стохастические всплески транскрипции, по-видимому, являются общей характеристикой высокоэкспрессируемых генов, а уровни суперспирализации матрицы ДНК способствуют взрыву транскрипции. [140] Согласно модели двойного суперспирального домена, транскрипция гена может влиять на транскрипцию других близлежащих генов посредством реле суперспирализации. Одним из таких примеров является активация промотора leu-500 . [139] Суперспирализация не только опосредует специфичные для генов изменения, но также обеспечивает крупномасштабные изменения в экспрессии генов. Топологическая организация нуклеоида может обеспечить независимую экспрессию генов, чувствительных к суперспирализации, в разных топологических доменах. Карта неограниченной сверхспирализации в масштабе генома показала, что геномные регионы имеют разные плотности суперспирализации в стационарном состоянии, что указывает на то, что уровень суперспирализации различается в отдельных топологических доменах. [133] В результате изменение суперспирализации может привести к домен-специфической экспрессии генов, в зависимости от уровня суперспирализации в каждом домене. [133]
Эффект суперспирализации на экспрессию генов может быть опосредован NAP, которые прямо или косвенно влияют на суперспирализацию. Влияние HU на экспрессию генов, по-видимому, связано с изменением суперспирализации и, возможно, организацией ДНК более высокого порядка. Положительная корреляция между связыванием ДНК-гиразы и активацией генов, вызванной отсутствием HU, предполагает, что изменения в суперспирализации ответственны за дифференциальную экспрессию. Также было обнаружено, что HU отвечает за позиционный эффект на экспрессию генов путем изоляции транскрипционных единиц путем ограничения суперспирализации, индуцированной транскрипцией. [141] Точечные мутации в HUa резко изменили профиль экспрессии генов E. coli, изменив ее морфологию , физиологию и метаболизм . В результате мутантный штамм оказался более инвазивным по отношению к клеткам млекопитающих. [134] [142] Этот драматический эффект сопровождался уплотнением нуклеоида и усилением положительной сверхспирализации. [45] [143] Мутантный белок представлял собой октамер, в отличие от димера дикого типа. Он оборачивает ДНК на своей поверхности правосторонним образом, сдерживая положительные суперспирали, в отличие от HU дикого типа. [143] Эти исследования показывают, что аминокислотные замены в HU могут оказывать существенное влияние на структуру нуклеоида, что, в свою очередь, приводит к значительным фенотипическим изменениям. [143]
Поскольку MukB и HU стали ключевыми игроками во взаимодействиях ДНК на больших расстояниях, будет полезно сравнить влияние каждого из этих двух белков на глобальную экспрессию генов. [144] Хотя HU, по-видимому, контролирует экспрессию генов путем модуляции плотности суперспирализации, точный молекулярный механизм остается неизвестным, а влияние MukB на экспрессию генов еще предстоит проанализировать. [144] [145]
Пространственная организация
[ редактировать ]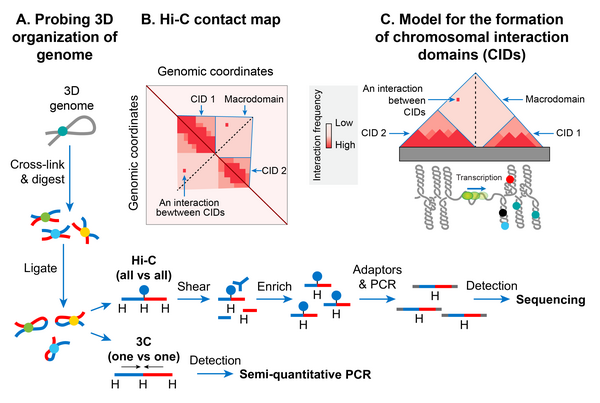
Домены хромосомного взаимодействия
[ редактировать ]В последние годы появление молекулярного метода, называемого захватом конформации хромосом (3C), позволило изучить пространственную организацию хромосом с высоким разрешением как у бактерий, так и у эукариот. [146] 3C и его версия, сочетающая глубокое секвенирование (Hi-C) [147] определить физическую близость, если таковая имеется, между любыми двумя геномными локусами в трехмерном пространстве. Карта контактов бактериальных хромосом с высоким разрешением, включая хромосому E. coli , показала, что бактериальная хромосома сегментирована на множество областей с высокой степенью самодействия, называемых доменами хромосомного взаимодействия (CID). [126] [148] [149] CID эквивалентны топологически ассоциированным доменам (TAD), наблюдаемым во многих хромосомах эукариот. [150] предполагая, что формирование CID является общим явлением организации генома. Две характеристики определяют CID или TAD. Во-первых, геномные области CID физически взаимодействуют друг с другом чаще, чем с геномными областями за пределами этого CID или с областями соседнего CID. Во-вторых, наличие границы между CID, препятствующей физическому взаимодействию геномных областей двух соседних CID. [126]
Было обнаружено, что хромосома E. coli состоит из 31 CID в фазе роста. Размер CID варьировался от 40 до ~300 КБ. Похоже, что барьер суперспирализации-диффузии, ответственный за разделение петель плектонемической ДНК на топологические домены, действует как граница CID у E. coli и многих других бактерий. Другими словами, наличие сверхспирационно-диффузионного барьера определяет формирование CID. Результаты зондирования Hi-C хромосом в E. coli , Caulobacter crescentus и Bacillus subtilis сходятся в модели, согласно которой формируются CID, поскольку плектонемическая петля вместе с деятельностью по организации ДНК NAP способствует физическому взаимодействию между геномными локусами, а граница CID состоит из область без плектонем (PFR), которая предотвращает эти взаимодействия. PFR создается из-за высокой транскрипционной активности, поскольку спиральное раскручивание ДНК за счет активной транскрипции РНКП сдерживает плектонемические суперспирали. В результате диссипация суперспиралей также блокируется, создавая барьер диффузии суперспиралей. Косвенным подтверждением этой модели является наблюдение, что CID бактериальных хромосом, включая Хромосома E. coli содержит на своих границах гены с высокой степенью транскрипции, что указывает на роль транскрипции в формировании границы CID. [126] [148] Более прямое доказательство было получено в результате открытия того, что размещение высоко транскрибируемого гена в положении, где не было границы, создавало новую границу CID в хромосоме C. crescentus . [148] Однако не все границы CID коррелируют с хорошо транскрибируемыми генами в хромосоме E. coli , что позволяет предположить, что другие неизвестные факторы также ответственны за формирование границ CID и диффузионных барьеров суперспирализации. [148]
Макродомены
[ редактировать ]Плектонемические петли ДНК, организованные в виде топологических доменов или CID, по-видимому, далее сливаются, образуя большие пространственно различные домены, называемые макродоменами (MD). В E. coli MD первоначально были идентифицированы как большие сегменты генома, чьи ДНК-маркеры локализовались вместе (совместно) в исследованиях флуоресцентной гибридизации in situ (FISH). [151] [152] Большая геномная область (~ 1 МБ), охватывающая локус oriC (начало репликации хромосом), колокализовалась и была названа макродоменом Ori. Аналогично, большая геномная область (~1-Mb), покрывающая область конца репликации ( ter ), локализовалась совместно и была названа макродоменом Ter. Позже MD были идентифицированы на основании того, как часто пары лямбда- атт -сайтов, вставленных в различные удаленные места хромосомы, рекомбинировали друг с другом. В этом методе, основанном на рекомбинации, MD определялся как большая область генома, сайты ДНК которой могут преимущественно рекомбинировать друг с другом, но не с участками за пределами этого MD. Метод, основанный на рекомбинации, подтвердил MD Ори и Тер, которые были идентифицированы в исследованиях FISH, и идентифицировал два дополнительных MD. [12] [153]
Два дополнительных MD были образованы дополнительными областями размером ~ 1 Мб, фланкирующими Ter, и назывались левым и правым. Эти четыре MD (Ori, Ter, Left и Right) составляют большую часть генома, за исключением двух геномных областей, фланкирующих Ori. Эти два региона (NS-L и NS-R) были более гибкими и неструктурированными по сравнению с МД, поскольку сайты ДНК в них рекомбинировали с сайтами ДНК, расположенными в МД с обеих сторон. Генетическое положение oriC , по-видимому, диктует формирование MD, поскольку изменение положения oriC с помощью генетических манипуляций приводит к реорганизации MD. Например, геномные регионы, ближайшие к oriC, всегда ведут себя как NS независимо от последовательности ДНК, а регионы, расположенные дальше, всегда ведут себя как MD. [154]
Метод Hi-C дополнительно подтвердил иерархическую пространственную организацию CID в форме макродоменов. [126] Другими словами, CID макродомена физически взаимодействовали друг с другом чаще, чем с CID соседнего макродомена или с геномными локусами вне этого макродомена. Данные Hi-C показали, что хромосома E. coli разделилась на два отдельных домена. Область, окружающая ter, образует изолированный домен, который перекрывается с ранее идентифицированным Ter MD. Контакты ДНК-ДНК в этом домене происходили только в диапазоне до ~280 т.п.н. Остальная часть хромосомы образовывала один домен, геномные локусы которого имели контакты в диапазоне> 280 т.п.н. [126] Хотя большинство контактов в этом домене были ограничены максимальным расстоянием ~500 т.п.н., существовали две рыхлые области, геномные локусы которых образовывали контакты на еще больших расстояниях (до ~1 Мб). Эти рыхлые области соответствовали ранее выявленным гибким и менее структурированным областям (NS). Границы изолированного домена, охватывающего ter , и две рыхлые области, идентифицированные методом Hi-C, сегментировали всю хромосому на шесть областей, которые соответствуют четырем MD и двум NS областям, определенным с помощью анализов на основе рекомбинации. [126]
Белки, управляющие образованием макродоменов
[ редактировать ]МатП
[ редактировать ]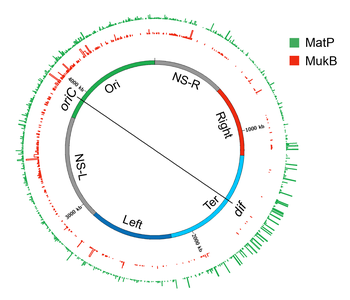
Поиск белка(ов), ответственного за образование макродоменов, привел к идентификации белка Macrodomain Ter (MatP). MatP почти исключительно связывается с Ter MD, распознавая мотив из 13 пар оснований, называемый макродоменовой последовательностью ter ( matS ). [32] В домене Ter присутствует 23 сайта matS , в среднем на каждые 35 КБ приходится один сайт. Дополнительные доказательства связывания MatP с доменом Ter получены при флуоресцентной визуализации MatP. Наблюдались дискретные фокусы MatP, которые локализовались совместно с маркерами ДНК домена Ter. [32] Сильное обогащение сигнала ChIP-Seq в Ter MD также подтверждает преимущественное связывание MatP с этим доменом. [32]
MatP конденсирует ДНК в домене Ter, поскольку отсутствие MatP увеличивает расстояние между двумя флуоресцентными маркерами ДНК, расположенными на расстоянии 100 т.п.н. друг от друга в домене Ter. Более того, MatP играет решающую роль в изоляции домена Ter от остальной части хромосомы. [126] Он способствует контактам ДНК-ДНК внутри домена Ter, но предотвращает контакты между локусами ДНК домена Ter и локусами фланкирующих областей. Как MatP конденсирует ДНК и способствует контактам ДНК-ДНК? Результаты экспериментов противоречивы. MatP может образовывать петлю ДНК между двумя matS сайтами in vitro , и его активность образования петли ДНК зависит от тетрамеризации MatP. Тетрамеризация происходит посредством спиральных взаимодействий между двумя молекулами MatP, связанными с ДНК. [156] Одна очевидная модель, основанная на результатах in vitro , заключается в том, что MatP способствует контактам ДНК-ДНК in vivo путем соединения сайтов matS . Однако, хотя MatP соединял удаленные сайты в исследованиях Hi-C, он не связывал конкретно сайты matS . Более того, мутант MatP, который не мог образовывать тетрамеры, вел себя как дикий тип. Эти результаты противоречат модели мостиков matS для организации Ter, оставляя механизм действия MatP неуловимым. Одна из возможностей состоит в том, что MatP распространяется на близлежащие сегменты ДНК от своего первичного сайта связывания matS и удаленных сайтов мостика по механизму, который не зависит от тетрамеризации. [156]
МукБЭФ
[ редактировать ]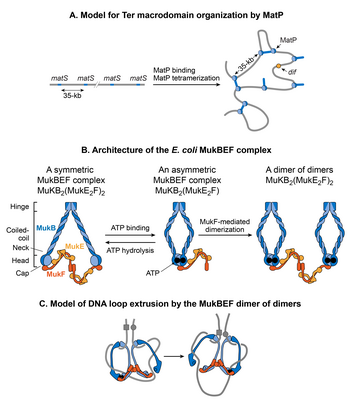
MukB принадлежит к семейству АТФаз, называемых структурным обеспечением белков хромосом (SMC), которые участвуют в организации хромосом высшего порядка у эукариот. [145] Два мономера MukB связываются посредством непрерывного антипараллельного спирального взаимодействия, образуя жесткий стержень длиной 100 нм. В середине стержня имеется гибкая шарнирная область. [162] [163] Благодаря гибкости шарнирной области MukB принимает характерную V-образную форму семейства SMC. Субъединицами, не относящимися к SMC, связанными с MukB, являются MukE и MukF. Ассоциация замыкает образование V, в результате чего образуются крупные кольцевые структуры. MukE и MukF кодируются вместе с MukB в одном опероне E. coli . [164] Делеция любой субъединицы приводит к одному и тому же фенотипу, что указывает на то, что комплекс MukBEF является функциональной единицей in vivo . [160] ДНК-связывающая активность комплекса находится в субъединице MukB, тогда как MukE и MukF модулируют активность MukB. [164]
Комплекс MukBEF вместе с Topo IV необходим для декатенации и репозиционирования вновь реплицированных oriC s. [165] [166] [167] [168] [155] Роль MukBEF не ограничивается во время репликации ДНК. Он организует и конденсирует ДНК даже в нереплицирующихся клетках. [131] Недавняя карта конформации хромосом высокого разрешения штамма E. coli , обедненного MukB , показывает, что MukB участвует в формировании взаимодействий ДНК-ДНК на всей хромосоме, за исключением домена Ter. [126] Как MukB не может действовать в домене Ter? MatP физически взаимодействует с MukB, тем самым предотвращая локализацию MukB в домене Ter. [155] Это проявляется в связывании ДНК MatP и MukB в домене Ter. ДНК-связывание MatP обогащено доменом Ter, тогда как ДНК-связывание MukB снижено по сравнению с остальной частью генома. Более того, у штамма, уже лишенного MatP, отсутствие MukB вызывает уменьшение контактов ДНК по всей хромосоме, включая домен Ter. [126] Этот результат согласуется с мнением, что MatP вытесняет MukB из домена Ter. [126]
Как комплекс MukBEF организует хромосому E. coli ? Согласно современным представлениям, комплексы SMC организуют хромосомы путем вытягивания петель ДНК. [169] Комплексы SMC перемещаются вдоль ДНК, образуя петли цис-способом (на одной и той же молекуле ДНК), при этом размер петель зависит от процессивности комплекса. Комплексы ГМК разных организмов различаются механизмом экструзии петель. [169] Флуоресцентная микроскопия одиночных молекул MukBEF в E. coli позволяет предположить, что минимальной функциональной единицей in vivo является димер из димеров. [160] Эта единица образуется путем соединения двух АТФ-связанных комплексов MukBEF посредством димеризации, опосредованной MukF. МукБЕФ локализуется в клетке в виде 1-3 кластеров, вытянутых параллельно длинной оси клетки. Каждый кластер содержит в среднем ~8-10 димеров. Согласно текущей модели, MukBEF выдавливает петли ДНК методом «скалолазания». [160] [170] Димер из димеров высвобождает один сегмент ДНК и захватывает новый сегмент ДНК, не отделяясь от хромосомы. Помимо образования петель ДНК, связь между отрицательной суперспирализацией и функцией MukBEF in vivo вместе со способностью субъединицы MukB ограничивать отрицательные суперспирали in vitro позволяет предположить, что MukBEF организует ДНК путем создания суперспиралей. [171] [172] [173]
Роль NAP и naRNA
[ редактировать ]Помимо вклада в уплотнение хромосом за счет изгиба, соединения и образования петель ДНК в меньшем масштабе (~ 1 т.п.н.), NAP участвуют в конденсации и организации ДНК, способствуя дальним контактам ДНК-ДНК. Два NAP, Fis и HU, стали ключевыми игроками в обеспечении дальних контактов ДНК-ДНК, которые происходят по всей хромосоме. [126] Еще предстоит изучить, как деятельность по организации ДНК Fis и HU, которая хорошо понята в меньшем масштабе (~ 1 т.п.н.), приводит к формированию дальнодействующих взаимодействий ДНК-ДНК. Тем не менее, некоторые взаимодействия ДНК, опосредованные HU, требуют присутствия naRNA4. [86] naRNA4 также участвует в установлении дальних контактов ДНК. HU катализирует некоторые контакты, а не все, что позволяет предположить, что РНК участвует вместе с другими NAP в формировании контактов ДНК. HU также, по-видимому, действует вместе с MukB, способствуя долгосрочным взаимодействиям ДНК-ДНК. Эта точка зрения основана на наблюдениях о том, что отсутствие HU или MukB вызывало уменьшение одних и тех же контактов ДНК-ДНК. Неясно, как MukB и HU потенциально действуют вместе, способствуя взаимодействиям ДНК-ДНК. Вполне возможно, что два белка взаимодействуют физически. Альтернативно, в то время как MukBEF выдавливает большие петли ДНК, HU конденсирует и организует эти петли. [169] [48]
Роль функционального родства генов
[ редактировать ]Есть сообщения о том, что функционально связанные гены E. coli физически находятся вместе в трехмерном пространстве внутри хромосомы, хотя они находятся далеко друг от друга на генетическом расстоянии. Пространственная близость функционально связанных генов не только делает биологические функции более разделенными и эффективными, но также способствует сворачиванию и пространственной организации нуклеоида. Недавнее исследование с использованием флуоресцентных маркеров для обнаружения специфических локусов ДНК изучило попарные физические расстояния между семью оперонами рРНК, которые генетически отделены друг от друга (на целых два миллиона пар оснований). В нем сообщалось, что все опероны, кроме rrn C, находились в физической близости. [174] [175] Удивительно, но исследования 3C-seq не выявили физической кластеризации оперонов rrn , что противоречит результатам исследования, основанного на флуоресценции. [126] Поэтому для разрешения этих противоречивых наблюдений необходимы дальнейшие исследования. В другом примере GalR образует сеть взаимодействия сайтов связывания GalR, разбросанных по хромосоме. [176] GalR представляет собой регулятор транскрипции галактозного регулона, состоящий из генов, кодирующих ферменты транспорта и метаболизма сахара D-галактозы. [177] GalR существует только в одном-двух очагах в клетках. [176] и могут самособираться в большие упорядоченные структуры. [178] Таким образом, оказывается, что связанный с ДНК GalR мультимеризуется с образованием взаимодействий на больших расстояниях. [176] [178]
Глобальная форма и структура
[ редактировать ]Традиционная трансмиссионная электронная микроскопия (ПЭМ) химически фиксированных клеток E. coli показала нуклеоид как органеллу неправильной формы . Однако широкопольная флуоресцентная визуализация живых нуклеоидов в 3D выявила дискретную эллипсоидную форму. [3] [14] [15] Наложение фазоконтрастного изображения клетки и флуоресцентного изображения нуклеоида показало близкое сопоставление только в радиальном измерении по всей длине нуклеоида до периферии клетки. Эта находка указывает на радиальное ограничение нуклеоида. [13] Детальное изучение трехмерного флуоресцентного изображения после поперечного сечения перпендикулярно его длинной оси дополнительно выявило две глобальные особенности нуклеоида: кривизну и продольные области с высокой плотностью. Исследование киральности центральной линии нуклеоида путем соединения центра интенсивности каждого поперечного сечения показало, что общая форма нуклеоида изогнута. [15] Распределение интенсивности флуоресценции в поперечных сечениях выявило субструктуру плотности, состоящую из изогнутых областей или пучков высокой плотности в центральном ядре и областей низкой плотности на периферии. [13] [14] Одним из следствий радиального ограничения является то, что оно определяет изогнутую форму нуклеоида. Согласно одной модели, нуклеоид вынужден изгибаться, поскольку он заключен в цилиндрическую клетку E. coli , радиус которой меньше ее сгибаемой длины (персистенции). [13] Эта модель была подтверждена наблюдениями о том, что удаление клеточной стенки или ингибирование синтеза клеточной стенки увеличивает радиус клетки и приводит к сопутствующему увеличению радиуса спирали и уменьшению шага спирали в нуклеоиде. [13]
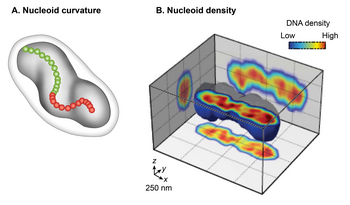
Нуклеоидно-мембранные связи
[ редактировать ]Сила расширения, обусловленная связями ДНК-мембрана, по-видимому, действует в противовес силам конденсации, поддерживая оптимальный уровень конденсации нуклеоида. Исследования клеточного фракционирования и электронной микроскопии впервые указали на возможность связей ДНК с мембраной. [179] [180] В настоящее время известно несколько примеров связей ДНК с мембраной. Трансерция — это механизм одновременной транскрипции, трансляции и вставки возникающих мембранных белков, который образует временные контакты ДНК-мембрана. [181] Было продемонстрировано, что трансерция двух мембранных белков LacY и TetA вызывает перемещение хромосомных локусов по направлению к мембране. [182] Другой механизм нуклеоидно-мембранных связей заключается в прямом контакте между мембранно-заякоренными регуляторами транскрипции и их целевыми участками в хромосоме. Одним из примеров такого регулятора транскрипции в E.coli является CadC. CadC содержит периплазматический сенсорный домен и цитоплазматический ДНК-связывающий домен. Ощущение кислой среды периплазматическим сенсорным доменом стимулирует ДНК-связывающую активность CadC, которая затем активирует транскрипцию его генов-мишеней. [183] Мембранная локализация генов, регулируемых мембранно-заякоренным регулятором транскрипции, еще не продемонстрирована. Тем не менее, ожидается, что активация генов-мишеней в хромосоме этими регуляторами приведет к контакту нуклеоид-мембрана, хотя это будет динамический контакт. Помимо этих примеров, хромосома также специфически закрепляется на клеточной мембране посредством белок-белкового взаимодействия между ДНК-связанными белками, например, SlmA и MatP, и дивисомой . [184] [185] Поскольку гены, кодирующие мембранные белки, распределены по всему геному, динамические контакты ДНК-мембраны посредством трансерции могут действовать как сила расширения нуклеоида. Эта сила расширения будет действовать в противовес силам конденсации для поддержания оптимального уровня конденсации. Образование высококонденсированных нуклеоидов при воздействии на клетки E. coli хлорамфеникола, который блокирует трансляцию, обеспечивает поддержку силы расширения временных контактов ДНК-мембрана, образующихся в результате трансерции. [186] [187] Круглая форма чрезмерно конденсированных нуклеоидов после обработки хлорамфениколом также предполагает роль опосредованных трансерцией контактов ДНК-мембраны в определении эллипсоидной формы нуклеоида. [187]
Визуализация
[ редактировать ]Нуклеоид можно четко визуализировать на электронной микрофотографии при очень большом увеличении , где, хотя его внешний вид может отличаться, он отчетливо виден на фоне цитозоля . [188] даже нити того, что принято считать ДНК Иногда видны . Окрашивая , который специфически окрашивает ДНК , красителем Фельгена нуклеоид также можно увидеть под световым микроскопом . [189] красители Интеркалирующие ДНК DAPI и бромистый этидий широко используются для флуоресцентной микроскопии нуклеоидов. Он имеет неправильную форму и встречается в прокариотических клетках. [13] [14]
Повреждение и восстановление ДНК
[ редактировать ]Изменения в структуре нуклеоида бактерий и архей наблюдаются после воздействия на ДНК повреждающих условий. Нуклеоиды бактерий Bacillus subtilis и Escherichia coli становятся значительно более компактными после УФ-облучения. [190] [191] Формирование компактной структуры в E. coli требует активации RecA посредством специфических взаимодействий RecA-ДНК. [192] Белок RecA играет ключевую роль в гомологичной рекомбинационной репарации повреждений ДНК.
Подобно B. subtilis и E. coli, описанным выше, воздействие на архей Haloferax volcanii стрессов, повреждающих ДНК, вызывает уплотнение и реорганизацию нуклеоида. [193] Компактизация зависит от белкового комплекса Mre11-Rad50, который катализирует раннюю стадию гомологичной рекомбинационной репарации двухцепочечных разрывов ДНК. Было высказано предположение, что уплотнение нуклеоида является частью реакции на повреждение ДНК, которая ускоряет восстановление клеток, помогая белкам репарации ДНК определять местонахождение мишеней и облегчая поиск интактных последовательностей ДНК во время гомологичной рекомбинации. [193]
См. также
[ редактировать ]Ссылки
[ редактировать ]![]() Эта статья была адаптирована из следующего источника под лицензией CC BY 4.0 ( 2019 ) ( отчеты рецензента ): Субхаш Верма; Чжун Цянь; Санкар Л. Адхья (декабрь 2019 г.). «Архитектура нуклеоида Escherichia coli» . ПЛОС Генетика . 15 (12): e1008456. doi : 10.1371/JOURNAL.PGEN.1008456 . ISSN 1553-7390 . ПМК 6907758 . ПМИД 31830036 . Викиданные Q84825966 .
Эта статья была адаптирована из следующего источника под лицензией CC BY 4.0 ( 2019 ) ( отчеты рецензента ): Субхаш Верма; Чжун Цянь; Санкар Л. Адхья (декабрь 2019 г.). «Архитектура нуклеоида Escherichia coli» . ПЛОС Генетика . 15 (12): e1008456. doi : 10.1371/JOURNAL.PGEN.1008456 . ISSN 1553-7390 . ПМК 6907758 . ПМИД 31830036 . Викиданные Q84825966 .
- ^ Танбихлер М., Ван С.К., Шапиро Л. (октябрь 2005 г.). «Бактериальный нуклеоид: высокоорганизованная и динамичная структура» . Журнал клеточной биохимии . 96 (3): 506–21. дои : 10.1002/jcb.20519 . ПМИД 15988757 .
- ^ Jump up to: а б с Дама RT, Тарк-Дам М (июнь 2016 г.). «Бактериальный хроматин: сходящиеся взгляды в разных масштабах». Современное мнение в области клеточной биологии . 40 :60–65. дои : 10.1016/j.ceb.2016.02.015 . ПМИД 26942688 .
- ^ Jump up to: а б с Клекнер Н., Фишер Дж.К., Стоуф М., Уайт М.А., Бейтс Д., Витц Дж. (декабрь 2014 г.). «Бактериальный нуклеоид: природа, динамика и сестринская сегрегация» . Современное мнение в микробиологии . 22 : 127–37. дои : 10.1016/j.mib.2014.10.001 . ПМЦ 4359759 . ПМИД 25460806 .
- ^ Jump up to: а б Блумфилд, Вирджиния (1997). «Конденсация ДНК многовалентными катионами». Биополимеры . 44 (3): 269–82. doi : 10.1002/(SICI)1097-0282(1997)44:3<269::AID-BIP6>3.0.CO;2-T . ПМИД 9591479 .
- ^ Jump up to: а б с д Трун, Нью-Джерси, Марко Дж. Ф. (1998). «Архитектура бактериальной хромосомы» (PDF) . Новости Американского общества микробиологии . 64 (5): 276–283.
- ^ Суровцев Иван Васильевич; Джейкобс-Вагнер, Кристина (март 2018 г.). «Субклеточная организация: важнейшая особенность репликации бактериальных клеток» . Клетка . 172 (6): 1271–1293. дои : 10.1016/j.cell.2018.01.014 . ПМК 5870143 . ПМИД 29522747 .
- ^ Jump up to: а б Стонингтон О.Г., Петтиджон Д.Э. (январь 1971 г.). «Свернутый геном Escherichia coli, выделенный в комплекс белок-ДНК-РНК» . Труды Национальной академии наук Соединенных Штатов Америки . 68 (1): 6–9. Бибкод : 1971ПНАС...68....6С . дои : 10.1073/pnas.68.1.6 . ПМК 391088 . ПМИД 4924971 .
- ^ Ворсель А., Бурги Э. (ноябрь 1972 г.). «О строении свернутой хромосомы кишечной палочки». Журнал молекулярной биологии . 71 (2): 127–47. дои : 10.1016/0022-2836(72)90342-7 . ПМИД 4564477 .
- ^ Jump up to: а б с д и Кано Ю., Госима Н., Вада М., Имамото Ф. (1989). «Участие продукта гена hup в репликативной транспозиции фага Mu в Escherichia coli». Джин . 76 (2): 353–8. дои : 10.1016/0378-1119(89)90175-3 . ПМИД 2666261 .
- ^ Jump up to: а б с д и Огура Т., Ники Х., Кано Ю., Имамото Ф., Хирага С. (январь 1990 г.). «Поддержание плазмид в мутантах HU и IHF Escherichia coli». Молекулярная и общая генетика . 220 (2): 197–203. дои : 10.1007/bf00260482 . ПМИД 2183003 . S2CID 10701528 .
- ^ Jump up to: а б с д и Хван Д.С., Корнберг А. (ноябрь 1992 г.). «Открытие начала репликации Escherichia coli белком DnaA с белком HU или IHF» . Журнал биологической химии . 267 (32): 23083–6. дои : 10.1016/S0021-9258(18)50059-4 . ПМИД 1429655 .
- ^ Jump up to: а б Валенс М., Пено С., Россиньоль М., Корне Ф., Боккар Ф. (октябрь 2004 г.). «Макродоменная организация хромосомы Escherichia coli» . Журнал ЭМБО . 23 (21): 4330–41. дои : 10.1038/sj.emboj.7600434 . ПМК 524398 . ПМИД 15470498 .
- ^ Jump up to: а б с д и ж г Фишер Дж.К., Бурникель А., Витц Г., Вайнер Б., Прентисс М., Клекнер Н. (май 2013 г.). «Четырехмерная визуализация организации и динамики нуклеоидов E. coli в живых клетках» . Клетка . 153 (4): 882–95. дои : 10.1016/j.cell.2013.04.006 . ПМЦ 3670778 . ПМИД 23623305 .
- ^ Jump up to: а б с д и Ле Галль А., Каттони Д.И., Гильяс Б., Матье-Демазиер С., Уджеди Л., Фиш Ж.Б. и др. (июль 2016 г.). «Бактериальные перегородочные комплексы разделяются внутри объема нуклеоида» . Природные коммуникации . 7 : 12107. Бибкод : 2016NatCo...712107L . дои : 10.1038/ncomms12107 . ПМЦ 4935973 . ПМИД 27377966 .
- ^ Jump up to: а б с Хадизаде Язди Н., Гет К.С., Джонсон Р.К., Марко Дж.Ф. (декабрь 2012 г.). «Изменение сворачивания и динамики хромосомы Escherichia coli в зависимости от условий роста» . Молекулярная микробиология . 86 (6): 1318–33. дои : 10.1111/mmi.12071 . ПМК 3524407 . ПМИД 23078205 .
- ^ Олинс А.Л., Олинс Д.Э. (январь 1974 г.). «Единицы сфероидного хроматина (v тельца)». Наука . 183 (4122): 330–2. Бибкод : 1974Sci...183..330O . дои : 10.1126/science.183.4122.330 . ПМИД 4128918 . S2CID 83480762 .
- ^ Jump up to: а б Люгер К., Мэдер А.В., Ричмонд Р.К., Сарджент Д.Ф., Ричмонд Т.Дж. (сентябрь 1997 г.). «Кристаллическая структура ядра нуклеосомы при разрешении 2,8 А». Природа . 389 (6648): 251–60. Бибкод : 1997Natur.389..251L . дои : 10.1038/38444 . ПМИД 9305837 . S2CID 4328827 .
- ^ Jump up to: а б Хорасанизаде С. (январь 2004 г.). «Нуклеосома: от геномной организации к геномной регуляции» . Клетка . 116 (2): 259–72. дои : 10.1016/s0092-8674(04)00044-3 . ПМИД 14744436 . S2CID 15504162 .
- ^ Талукдер А., Исихама А. (сентябрь 2015 г.). «Фазовые изменения структуры и белкового состава нуклеоида Escherichia coli» . Наука Китай Науки о жизни . 58 (9): 902–11. дои : 10.1007/s11427-015-4898-0 . ПМИД 26208826 .
- ^ Jump up to: а б с д и ж Азам Т.А., Исихама А. (ноябрь 1999 г.). «Двенадцать видов нуклеоид-ассоциированного белка из Escherichia coli. Специфичность распознавания последовательностей и аффинность связывания ДНК» . Журнал биологической химии . 274 (46): 33105–13. дои : 10.1074/jbc.274.46.33105 . ПМИД 10551881 . S2CID 9807664 .
- ^ Jump up to: а б с д и ж г Али Азам Т., Ивата А., Нишимура А., Уэда С., Исихама А. (октябрь 1999 г.). «Зависимые от фазы роста изменения белкового состава нуклеоида Escherichia coli» . Журнал бактериологии . 181 (20): 6361–70. дои : 10.1128/JB.181.20.6361-6370.1999 . ПМЦ 103771 . ПМИД 10515926 .
- ^ Jump up to: а б Свингер К.К., Лемберг К.М., Чжан Ю., Райс П.А. (июль 2003 г.). «Гибкий изгиб ДНК в сокристаллических структурах HU-ДНК» . Журнал ЭМБО . 22 (14): 3749–60. дои : 10.1093/emboj/cdg351 . ПМК 165621 . ПМИД 12853489 .
- ^ Jump up to: а б Го Ф, Адхья С (март 2007 г.). «Спиральная структура Escherichia coli HUalphabeta обеспечивает основу для сверхспирализации ДНК» . Труды Национальной академии наук Соединенных Штатов Америки . 104 (11): 4309–14. Бибкод : 2007PNAS..104.4309G . дои : 10.1073/pnas.0611686104 . ПМЦ 1838598 . ПМИД 17360520 .
- ^ Jump up to: а б с Пинсон В., Такахаши М., Рувьер-Янив Дж. (апрель 1999 г.). «Дифференциальное связывание Escherichia coli HU, гомодимерных форм и гетеродимерных форм с линейной, разрывной и крестообразной ДНК». Журнал молекулярной биологии . 287 (3): 485–97. дои : 10.1006/jmbi.1999.2631 . ПМИД 10092454 .
- ^ Крейг Н.Л., Нэш Х.А. (декабрь 1984 г.). «Фактор хозяина интеграции E. coli связывается с определенными участками ДНК». Клетка . 39 (3, часть 2): 707–16. дои : 10.1016/0092-8674(84)90478-1 . ПМИД 6096022 . S2CID 26758055 .
- ^ Jump up to: а б Оу Х.Д., Фан С., Диринк Т.Дж., Тор А., Эллисман М.Х., О'Ши CC (июль 2017 г.). «ChromEMT: визуализация трехмерной структуры и уплотнения хроматина в интерфазных и митотических клетках» . Наука . 357 (6349): eaag0025. дои : 10.1126/science.aag0025 . ПМК 5646685 . ПМИД 28751582 .
- ^ Jump up to: а б с Ланг Б., Блот Н., Буффартиг Э., Бакл М., Гирц М., Гуалерзи К.О. и др. (сентябрь 2007 г.). «Высокоаффинные сайты связывания ДНК для H-NS обеспечивают молекулярную основу для избирательного молчания в геномах протеобактерий» . Исследования нуклеиновых кислот . 35 (18): 6330–7. дои : 10.1093/нар/gkm712 . ПМК 2094087 . ПМИД 17881364 .
- ^ Jump up to: а б с Гульвади Р., Гао Ю., Кенни Л.Дж., Ян Дж. (ноябрь 2018 г.). «Анализ одной молекулы H-NS отделяет аффинность связывания ДНК от специфичности ДНК» . Исследования нуклеиновых кислот . 46 (19): 10216–10224. дои : 10.1093/nar/gky826 . ПМК 6212787 . ПМИД 30239908 .
- ^ Jump up to: а б с Шао Ю., Фельдман-Коэн Л.С., Осуна Р. (февраль 2008 г.). «Функциональная характеристика связывающей последовательности Fis-ДНК Escherichia coli» . Журнал молекулярной биологии . 376 (3): 771–85. дои : 10.1016/j.jmb.2007.11.101 . ПМК 2292415 . ПМИД 18178221 .
- ^ Jump up to: а б с д Стелла С., Касио Д., Джонсон Р.К. (апрель 2010 г.). «Форма малой бороздки ДНК управляет связыванием ДНК-изгибающего белка Fis» . Гены и развитие . 24 (8): 814–26. дои : 10.1101/gad.1900610 . ПМЦ 2854395 . ПМИД 20395367 .
- ^ Нараян К., Субраманиам С. (ноябрь 2015 г.). «Сфокусированные ионные пучки в биологии» . Природные методы . 12 (11): 1021–31. дои : 10.1038/nmeth.3623 . ПМК 6993138 . ПМИД 26513553 .
- ^ Jump up to: а б с д Мерсье Р., Пети М.А., Шбат С. , Робин С., Эль Каруи М., Боккар Ф., Эспели О. (октябрь 2008 г.). «Сайт-специфическая система MatP/matS организует терминальную область хромосомы E. coli в макродомен» (PDF) . Клетка . 135 (3): 475–85. дои : 10.1016/j.cell.2008.08.031 . ПМИД 18984159 . S2CID 3582710 .
- ^ Рувьер-Янив Ж, Грос Ф (сентябрь 1975 г.). «Характеристика нового низкомолекулярного ДНК-связывающего белка из Escherichia coli» . Труды Национальной академии наук Соединенных Штатов Америки . 72 (9): 3428–32. Бибкод : 1975PNAS...72.3428R . дои : 10.1073/pnas.72.9.3428 . ПМК 433007 . ПМИД 1103148 .
- ^ Сурьянараяна Т., Субраманиан, Арканзас (сентябрь 1978 г.). «Специфическая ассоциация двух гомологичных ДНК-связывающих белков с нативными 30-S рибосомальными субъединицами Escherichia coli». Biochimica et Biophysical Acta (BBA) - Нуклеиновые кислоты и синтез белка . 520 (2): 342–57. дои : 10.1016/0005-2787(78)90232-0 . ПМИД 213117 .
- ^ Менде Л., Тимм Б., Субраманиан Р. (декабрь 1978 г.). «Первичные структуры двух гомологичных рибосом-ассоциированных ДНК-связывающих белков Escherichia coli» . Письма ФЭБС . 96 (2): 395–8. Бибкод : 1978FEBSL..96..395M . дои : 10.1016/0014-5793(78)80446-3 . ПМИД 215461 . S2CID 39245157 .
- ^ Меграу Т.Л., Че CB (июнь 1993 г.). «Функциональная комплементарность между HMG1-подобным митохондриальным гистоном дрожжей HM и бактериальным гистоноподобным белком HU» . Журнал биологической химии . 268 (17): 12758–63. дои : 10.1016/S0021-9258(18)31453-4 . ПМИД 8509411 .
- ^ Полл Т.Т., Джонсон Р.К. (апрель 1995 г.). «Петлеобразование ДНК с помощью белков группы высокой подвижности Saccharomyces cerevisiae NHP6A/B. Последствия для сборки нуклеопротеинового комплекса и конденсации хроматина» . Журнал биологической химии . 270 (15): 8744–54. дои : 10.1074/jbc.270.15.8744 . ПМИД 7721780 .
- ^ Камашев Д., Рувьер-Янив Ж (декабрь 2000 г.). «Гистоноподобный белок HU специфически связывается с промежуточными продуктами рекомбинации и репарации ДНК» . Журнал ЭМБО . 19 (23): 6527–35. дои : 10.1093/emboj/19.23.6527 . ПМК 305869 . ПМИД 11101525 .
- ^ Шиндо Х., Фурубаяси А., Симидзу М., Мияке М., Имамото Ф. (апрель 1992 г.). «Преимущественное связывание гистоноподобного белка HU альфа E.coli с отрицательно сверхспиральной ДНК» . Исследования нуклеиновых кислот . 20 (7): 1553–8. дои : 10.1093/нар/20.7.1553 . ПМК 312237 . ПМИД 1579448 .
- ^ Понтиджа А., Негри А., Бельтрам М., Бьянки М.Е. (февраль 1993 г.). «Белок HU специфически связывается с изогнутой ДНК». Молекулярная микробиология . 7 (3): 343–50. дои : 10.1111/j.1365-2958.1993.tb01126.x . ПМИД 8459763 . S2CID 39362449 .
- ^ Боннефой Э., Такахаши М., Янив-младший (сентябрь 1994 г.). «Параметры ДНК-связывания белка HU Escherichia coli с крестообразной ДНК». Журнал молекулярной биологии . 242 (2): 116–29. дои : 10.1006/jmbi.1994.1563 . ПМИД 8089835 .
- ^ Кастан Б., Зельвер С., Лаваль Дж., Боите С. (апрель 1995 г.). «Белок HU Escherichia coli специфически связывается с ДНК, которая содержит одноцепочечные разрывы или пробелы» . Журнал биологической химии . 270 (17): 10291–6. дои : 10.1074/jbc.270.17.10291 . ПМИД 7730334 .
- ^ Любченко Ю.Л., Шляхтенко Л.С., Аки Т., Адья С. (февраль 1997 г.). «Атомно-силовая микроскопия, демонстрирующая образование петель ДНК с помощью GalR и HU» . Исследования нуклеиновых кислот . 25 (4): 873–6. дои : 10.1093/нар/25.4.873 . ПМК 146491 . ПМИД 9016640 .
- ^ Свингер К.К., Райс П.А. (январь 2007 г.). «Структурный анализ связывания HU-ДНК» . Журнал молекулярной биологии . 365 (4): 1005–16. дои : 10.1016/j.jmb.2006.10.024 . ЧВК 1945228 . ПМИД 17097674 .
- ^ Jump up to: а б с д и ж г Хаммел М., Амланджьоти Д., Рейес Ф.Е., Чен Дж.Х., Парпана Р., Тан Х.И. и др. (июль 2016 г.). «Сдвиг мультимеризации HU контролирует уплотнение нуклеоида» . Достижения науки . 2 (7): е1600650. Бибкод : 2016SciA....2E0650H . дои : 10.1126/sciadv.1600650 . ПМЦ 4966879 . ПМИД 27482541 .
- ^ Jump up to: а б с д и Прието А.И., Кахраманоглу С., Али Р.М., Фрейзер Г.М., Сешасаи А.С., Ласкомб Н.М. (апрель 2012 г.). «Геномный анализ связывания ДНК и регуляции генов гомологичными нуклеоид-ассоциированными белками IHF и HU в Escherichia coli K12» . Исследования нуклеиновых кислот . 40 (8): 3524–37. дои : 10.1093/nar/gkr1236 . ПМЦ 3333857 . ПМИД 22180530 .
- ^ Jump up to: а б с д и ж Макванин М., Эдгар Р., Кюи Ф., Тростел А., Журкин В., Адья С. (ноябрь 2012 г.). «Некодирующие РНК, связывающиеся с нуклеоидным белком HU в Escherichia coli» . Журнал бактериологии . 194 (22): 6046–55. дои : 10.1128/JB.00961-12 . ПМЦ 3486375 . ПМИД 22942248 .
- ^ Jump up to: а б с д и ван Ноорт Дж., Вербрюгге С., Гусен Н., Деккер С., Дам Р.Т. (май 2004 г.). «Двойная архитектурная роль HU: формирование гибких шарниров и жестких нитей» . Труды Национальной академии наук Соединенных Штатов Америки . 101 (18): 6969–74. Бибкод : 2004PNAS..101.6969V . дои : 10.1073/pnas.0308230101 . ПМК 406450 . ПМИД 15118104 .
- ^ Саркар Р., Рыбенков В.В. (06.12.2016). «Руководство по магнитным пинцетам и их применению» . Границы в физике . 4 : 48. Бибкод : 2016FrP.....4...48S . дои : 10.3389/fphy.2016.00048 . S2CID 44183628 .
- ^ Jump up to: а б с д Кахраманоглу С., Сешасаи А.С., Прието А.И., Ибберсон Д., Шмидт С., Циммерманн Дж. и др. (март 2011 г.). «Прямое и косвенное влияние H-NS и Fis на глобальный контроль экспрессии генов в Escherichia coli» . Исследования нуклеиновых кислот . 39 (6): 2073–91. дои : 10.1093/нар/gkq934 . ПМК 3064808 . ПМИД 21097887 .
- ^ Jump up to: а б Райс П.А., Ян С., Мизуучи К., Нэш Х.А. (декабрь 1996 г.). «Кристаллическая структура комплекса IHF-ДНК: индуцированный белком разворот ДНК» . Клетка . 87 (7): 1295–306. дои : 10.1016/s0092-8674(00)81824-3 . ПМИД 8980235 . S2CID 9291279 .
- ^ Муртин С., Энгельхорн М., Гейзельманн Дж., Боккар Ф. (декабрь 1998 г.). «Количественный УФ-лазерный анализ взаимодействия IHF со специфическими сайтами связывания: повторная оценка эффективной концентрации IHF в клетке». Журнал молекулярной биологии . 284 (4): 949–61. дои : 10.1006/jmbi.1998.2256 . ПМИД 9837718 .
- ^ То же самое, доктор медицинских наук, Робертс Д., Вейсберг Р.А. (июнь 1994 г.). «Фазовые изменения уровня фактора интеграции хозяина в Escherichia coli» . Журнал бактериологии . 176 (12): 3738–48. дои : 10.1128/jb.176.12.3738-3748.1994 . ПМК 205563 . ПМИД 8206852 .
- ^ Jump up to: а б с Линь Дж., Чен Х., Дрёге П., Ян Дж. (2012). «Физическая организация ДНК с помощью множественных неспецифических способов связывания ДНК с интеграцией фактора хозяина (IHF)» . ПЛОС ОДИН . 7 (11): e49885. Бибкод : 2012PLoSO...749885L . дои : 10.1371/journal.pone.0049885 . ПМЦ 3498176 . ПМИД 23166787 .
- ^ Жаке М., Кукье-Кан Р., Пла Дж., Грос Ф. (декабрь 1971 г.). «Термостабильный белковый фактор, действующий на транскрипцию ДНК in vitro». Связь с биохимическими и биофизическими исследованиями . 45 (6): 1597–607. дои : 10.1016/0006-291x(71)90204-x . ПМИД 4942735 .
- ^ Кукьер-Кан Р., Жаке М., Грос Ф (декабрь 1972 г.). «Два термостойких низкомолекулярных белка Escherichia coli, которые стимулируют ДНК-направленный синтез РНК» . Труды Национальной академии наук Соединенных Штатов Америки . 69 (12): 3643–7. Бибкод : 1972PNAS...69.3643C . дои : 10.1073/pnas.69.12.3643 . ПМЦ 389839 . ПМИД 4566454 .
- ^ Спасский А, Бук ХК (ноябрь 1977 г.). «Физико-химические свойства ДНК-связывающего белка: фактор H1 Escherichia coli» . Европейский журнал биохимии . 81 (1): 79–90. дои : 10.1111/j.1432-1033.1977.tb11929.x . ПМИД 338303 .
- ^ Варшавский А.Я., Недоспасов С.А., Бакаев В.В., Бакаева Т.Г., Георгиев Г.П. (август 1977 г.). «Гистоноподобные белки в очищенном дезоксирибонуклеопротеине Escherichia coli» . Исследования нуклеиновых кислот . 4 (8): 2725–45. дои : 10.1093/нар/4.8.2725 . ПМК 342604 . ПМИД 333393 .
- ^ Фалькони М., Гуальтьери М.Т., Ла Теана А., Лоссо М.А., Пон К.Л. (май 1988 г.). «Белки прокариотического нуклеоида: первичная и четвертичная структура ДНК-связывающего белка H-NS Escherichia coli массой 15 кДа». Молекулярная микробиология . 2 (3): 323–9. дои : 10.1111/j.1365-2958.1988.tb00035.x . ПМИД 3135462 . S2CID 36215353 .
- ^ Уэгути С., Сузуки Т., Ёсида Т., Танака К., Мизуно Т. (октябрь 1996 г.). «Систематический мутационный анализ, выявляющий организацию функционального домена нуклеоидного белка H-NS Escherichia coli». Журнал молекулярной биологии . 263 (2): 149–62. дои : 10.1006/jmbi.1996.0566 . ПМИД 8913298 .
- ^ Jump up to: а б Римский С., Зубер Ф., Бакл М., Бук Х. (декабрь 2001 г.). «Молекулярный механизм репрессии транскрипции белком H-NS» . Молекулярная микробиология . 42 (5): 1311–23. дои : 10.1046/j.1365-2958.2001.02706.x . ПМИД 11886561 . S2CID 32623528 .
- ^ Буффартиг Э., Бакл М., Бадо С., Траверс А., Римский С. (май 2007 г.). «Кооперативное связывание H-NS с сайтами с высоким сродством в регуляторном элементе приводит к подавлению транскрипции». Структурная и молекулярная биология природы . 14 (5): 441–8. дои : 10.1038/nsmb1233 . ПМИД 17435766 . S2CID 43768346 .
- ^ Дама RT, Вайман С., Гусен Н. (сентябрь 2000 г.). «H-NS-опосредованное уплотнение ДНК, визуализированное с помощью атомно-силовой микроскопии» . Исследования нуклеиновых кислот . 28 (18): 3504–10. дои : 10.1093/нар/28.18.3504 . ПМК 110753 . ПМИД 10982869 .
- ^ Амит Р., Оппенгейм А.Б., Ставанс Дж. (апрель 2003 г.). «Повышение жесткости изгиба отдельных молекул ДНК с помощью H-NS, датчика температуры и осмолярности» . Биофизический журнал . 84 (4): 2467–73. Бибкод : 2003BpJ....84.2467A . дои : 10.1016/S0006-3495(03)75051-6 . ПМК 1302812 . ПМИД 12668454 .
- ^ Dame RT, Noom MC, Wuite GJ (ноябрь 2006 г.). «Организация бактериального хроматина белком H-NS, раскрытая с помощью манипуляций с двойной ДНК». Природа . 444 (7117): 387–90. Бибкод : 2006Natur.444..387D . дои : 10.1038/nature05283 . ПМИД 17108966 . S2CID 4314858 .
- ^ Jump up to: а б с д и ж Лю Ю, Чен Х, Кенни Л.Дж., Ян Дж. (февраль 2010 г.). «Двухвалентный переключатель управляет конформациями связывания H-NS / ДНК между режимами жесткости и мостика» . Гены и развитие . 24 (4): 339–44. дои : 10.1101/gad.1883510 . ПМК 2816733 . ПМИД 20159954 .
- ^ ван дер Валк Р.А., Вриде Дж., Цинь Л., Муленаар Г.Ф., Хофманн А., Гусен Н., Дам Р.Т. (сентябрь 2017 г.). «Механизм конформационных изменений, обусловленных окружающей средой, которые модулируют активность мостиков ДНК H-NS» . электронная жизнь . 6 . дои : 10.7554/eLife.27369 . ПМЦ 5647153 . ПМИД 28949292 .
- ^ Ямада Х., Мурамацу С., Мизуно Т. (сентябрь 1990 г.). «Белок Escherichia coli, который преимущественно связывается с резко изогнутой ДНК». Журнал биохимии . 108 (3): 420–5. doi : 10.1093/oxfordjournals.jbchem.a123216 . ПМИД 2126011 .
- ^ Мартин-Ороско Н., Туре Н., Захарик М.Л., Парк Э., Копельман Р., Миллер С. и др. (январь 2006 г.). «Визуализация транскрипции генов патогенов, индуцированной вакуолярным закислением внутри макрофагов» . Молекулярная биология клетки . 17 (1): 498–510. дои : 10.1091/mbc.e04-12-1096 . ПМЦ 1345685 . ПМИД 16251362 .
- ^ Винарди Р.С., Ян Дж., Кенни Л.Дж. (октябрь 2015 г.). «H-NS регулирует экспрессию генов и уплотняет нуклеоид: результаты экспериментов с одиночными молекулами» . Биофизический журнал . 109 (7): 1321–9. Бибкод : 2015BpJ...109.1321W . дои : 10.1016/j.bpj.2015.08.016 . ПМК 4601063 . ПМИД 26445432 .
- ^ Уолтерс Д., Ли Й., Лю Й., Ананд Дж., Ян Дж., Кенни Л.Дж. (январь 2011 г.). «Регулятор ответа Salmonella enterica SsrB ослабляет подавление H-NS, вытесняя H-NS, связанный в режиме полимеризации, и напрямую активирует транскрипцию» . Журнал биологической химии . 286 (3): 1895–902. дои : 10.1074/jbc.M110.164962 . ПМК 3023485 . ПМИД 21059643 .
- ^ Гао Ю, Фу ЮХ, Винарди Р.С., Тан Кью, Ян Дж., Кенни Л.Дж. (ноябрь 2017 г.). «Заряженные остатки в линкере H-NS управляют связыванием ДНК и подавлением генов в отдельных клетках» . Труды Национальной академии наук Соединенных Штатов Америки . 114 (47): 12560–12565. Бибкод : 2017PNAS..11412560G . дои : 10.1073/pnas.1716721114 . ПМК 5703333 . ПМИД 29109287 .
- ^ Хэнкок С.П., Стелла С., Касио Д., Джонсон Р.К. (2016). «Детерминанты последовательности ДНК, контролирующие сродство, стабильность и форму комплексов ДНК, связанных нуклеоидным белком Fis» . ПЛОС ОДИН . 11 (3): e0150189. Бибкод : 2016PLoSO..1150189H . дои : 10.1371/journal.pone.0150189 . ПМЦ 4784862 . ПМИД 26959646 .
- ^ Кострева Д., Гранзин Дж., Кох С., Чой Х.В., Рагунатан С., Вольф В. и др. (январь 1991 г.). «Трехмерная структура ДНК-связывающего белка FIS E. coli». Природа . 349 (6305): 178–80. Бибкод : 1991Natur.349..178K . дои : 10.1038/349178a0 . ПМИД 1986310 . S2CID 4318862 .
- ^ Кострева Д., Гранзин Дж., Сток Д., Чой Х.В., Лабан Дж., Сенгер В. (июль 1992 г.). «Кристаллическая структура фактора инверсионной стимуляции FIS при разрешении 2,0 А». Журнал молекулярной биологии . 226 (1): 209–26. дои : 10.1016/0022-2836(92)90134-6 . ПМИД 1619650 .
- ^ Чо Б.К., Найт Э.М., Барретт К.Л., Палссон Б.О. (июнь 2008 г.). «Полногеномный анализ связывания Fis в Escherichia coli указывает на причинную роль A-/AT-трактов» . Геномные исследования . 18 (6): 900–10. дои : 10.1101/гр.070276.107 . ПМК 2413157 . ПМИД 18340041 .
- ^ Jump up to: а б Трэверс А., Мусхелишвили Г (июнь 1998 г.). «Микропетли ДНК и микродомены: общий механизм активации транскрипции посредством торсионной передачи». Журнал молекулярной биологии . 279 (5): 1027–43. дои : 10.1006/jmbi.1998.1834 . ПМИД 9642081 .
- ^ Скоко Д., Ян Дж., Джонсон Р.К., Марко Дж.Ф. (ноябрь 2005 г.). «Конденсация ДНК с низкой силой и прерывистая деконденсация с высокой силой обнаруживают стабилизирующую петлю функцию белка Fis» . Письма о физических отзывах . 95 (20): 208101. Бибкод : 2005PhRvL..95t8101S . doi : 10.1103/PhysRevLett.95.208101 . ПМИД 16384101 . S2CID 37405026 .
- ^ Jump up to: а б Скоко Д., Ю Д., Бай Х., Шнурр Б., Ян Дж., МакЛеод С.М. и др. (декабрь 2006 г.). «Механизм уплотнения хромосом и образования петель нуклеоидным белком Fis Escherichia coli» . Журнал молекулярной биологии . 364 (4): 777–98. дои : 10.1016/j.jmb.2006.09.043 . ЧВК 1988847 . ПМИД 17045294 .
- ^ Джонсон Р.К., Саймон М.И. (июль 1985 г.). «Hin-опосредованная сайт-специфическая рекомбинация требует двух сайтов рекомбинации по 26 п.н. и рекомбинационного энхансера на 60 п.н.». Клетка . 41 (3): 781–91. дои : 10.1016/s0092-8674(85)80059-3 . ПМИД 2988787 . S2CID 34572809 .
- ^ Jump up to: а б Кахманн Р., Рудт Ф., Кох К., Мертенс Г. (июль 1985 г.). «Инверсия G в ДНК бактериофага Mu стимулируется участком гена инвертазы и фактором хозяина» . Клетка . 41 (3): 771–80. дои : 10.1016/S0092-8674(85)80058-1 . ПМИД 3159478 .
- ^ Петтиджон Д.Е., Хехт Р. (1974). «Молекулы РНК, связанные со свернутым бактериальным геномом, стабилизируют складки ДНК и разделяют домены суперспирализации». Симпозиумы Колд-Спринг-Харбор по количественной биологии . 38 : 31–41. дои : 10.1101/sqb.1974.038.01.006 . ПМИД 4598638 .
- ^ Jump up to: а б с Онива Р.Л., Морикава К., Такешита С.Л., Ким Дж., Охта Т., Вада С., Такэясу К. (октябрь 2007 г.). «Транскрипционно-связанная архитектура нуклеоидов у бактерий» . Гены в клетки . 12 (10): 1141–52. дои : 10.1111/j.1365-2443.2007.01125.x . ПМИД 17903174 .
- ^ Jump up to: а б с Баландина А., Камашев Д., Рувьер-Янов Ж (август 2002 г.). «Бактериальный гистонподобный белок HU специфически распознает сходные структуры во всех нуклеиновых кислотах: ДНК, РНК и их гибридах» . Журнал биологической химии . 277 (31): 27622–8. дои : 10.1074/jbc.M201978200 . ПМИД 12006568 .
- ^ Баландина А., Кларет Л., Хенгге-Аронис Р., Рувьер-Янив Дж. (февраль 2001 г.). «Гистоноподобный белок HU Escherichia coli регулирует трансляцию rpoS» . Молекулярная микробиология . 39 (4): 1069–79. дои : 10.1046/j.1365-2958.2001.02305.x . ПМИД 11251825 .
- ^ Jump up to: а б Цянь З., Макванин М., Димитриадис Е.К., Хе Х, Журкин В., Адхья С. (август 2015 г.). «Новая некодирующая РНК организует бактериальную хромосомную организацию» . мБио . 6 (4): e00998-15. дои : 10.1128/mBio.00998-15 . ПМК 4550694 . ПМИД 26307168 .
- ^ Цянь З., Журкин В.Б., Адхья С. (ноябрь 2017 г.). «Эшерихия коли» . Труды Национальной академии наук Соединенных Штатов Америки . 114 (46): 12225–12230. дои : 10.1073/pnas.1711285114 . ПМК 5699063 . ПМИД 29087325 .
- ^ Бауэр В.Р., Крик Ф.Х., Уайт Дж.Х. (июль 1980 г.). «Сверхспиральная ДНК». Научный американец . 243 (1): 100–13. ПМИД 6256851 .
- ^ «Коэффициент связи» . www.энциклопедияofmath.org . Энциклопедия математики . Проверено 22 февраля 2020 г.
- ^ Jump up to: а б с Бейтс А.Д., Максвелл А. (2005). Топология ДНК (2-е изд.). Оксфорд: Издательство Оксфордского университета. ISBN 978-0-19-856709-7 . OCLC 56793994 .
- ^ Jump up to: а б с Вологодский Александр Вадимович. (1992). Топология и физика кольцевой ДНК . ЦРК Пресс. ISBN 0-8493-4228-7 . OCLC 635611152 .
- ^ Синден Р.Р. (2006). Структура и функции ДНК . Эльзевир. ISBN 978-0-12-645750-6 . OCLC 698461259 .
- ^ Jump up to: а б Синден Р.Р., Карлсон Дж.О., Петтиджон Д.Е. (октябрь 1980 г.). «Крученое напряжение двойной спирали ДНК, измеренное с помощью триметилпсоралена в живых клетках E. coli: аналогичные измерения в клетках насекомых и человека». Клетка . 21 (3): 773–83. дои : 10.1016/0092-8674(80)90440-7 . ПМИД 6254668 . S2CID 2503376 .
- ^ Гриффит Дж. Д. (февраль 1976 г.). «Визуализация прокариотической ДНК в регулярно конденсированном хроматиноподобном волокне» . Труды Национальной академии наук Соединенных Штатов Америки . 73 (2): 563–7. Бибкод : 1976ПНАС...73..563Г . дои : 10.1073/pnas.73.2.563 . ПМК 335950 . ПМИД 1108025 .
- ^ Jump up to: а б с Постоу Л., Харди К.Д., Арсуага Дж., Козцарелли Н.Р. (июль 2004 г.). «Топологическая доменная структура хромосомы Escherichia coli» . Гены и развитие . 18 (14): 1766–79. дои : 10.1101/gad.1207504 . ПМК 478196 . ПМИД 15256503 .
- ^ Jump up to: а б Блиска Дж. Б., Коццарелли Н. Р. (март 1987 г.). «Использование сайт-специфической рекомбинации в качестве исследования структуры и метаболизма ДНК in vivo». Журнал молекулярной биологии . 194 (2): 205–18. дои : 10.1016/0022-2836(87)90369-х . ПМИД 3039150 .
- ^ Холмс В.Ф., Коцарелли Н.Р. (февраль 2000 г.). «Замыкание кольца: связи между белками SMC и разделением хромосом, конденсацией и суперспирализацией» . Труды Национальной академии наук Соединенных Штатов Америки . 97 (4): 1322–4. Бибкод : 2000PNAS...97.1322H . дои : 10.1073/pnas.040576797 . ПМК 34294 . ПМИД 10677457 .
- ^ Шампу Джей-Джей (2001). «ДНК-топоизомеразы: структура, функции и механизм». Ежегодный обзор биохимии . 70 : 369–413. doi : 10.1146/annurev.biochem.70.1.369 . ПМИД 11395412 . S2CID 18144189 .
- ^ Геллерт М., Мизуучи К., О'Ди М.Х., Нэш Х.А. (ноябрь 1976 г.). «ДНК-гираза: фермент, который вводит в ДНК суперспиральные повороты» . Труды Национальной академии наук Соединенных Штатов Америки . 73 (11): 3872–6. Бибкод : 1976PNAS...73.3872G . дои : 10.1073/pnas.73.11.3872 . ПМЦ 431247 . ПМИД 186775 .
- ^ Jump up to: а б с Депью Р.Э., Лю Л.Ф. , Ван Дж.К. (январь 1978 г.). «Взаимодействие ДНК с белком омега Escherichia coli. Образование комплекса одноцепочечной ДНК с белком омега» . Журнал биологической химии . 253 (2): 511–8. дои : 10.1016/S0021-9258(17)38239-X . ПМИД 338610 .
- ^ Киркегор К., Ван Дж. К. (октябрь 1978 г.). «ДНК-топоизомераза I Escherichia coli катализирует связывание одноцепочечных колец комплементарных последовательностей оснований» . Исследования нуклеиновых кислот . 5 (10): 3811–20. дои : 10.1093/нар/5.10.3811 . ПМЦ 342711 . ПМИД 214763 .
- ^ Jump up to: а б Раджи А., Забель DJ, Лауфер К.С., Депью Р.Э. (июнь 1985 г.). «Генетический анализ мутаций, компенсирующих потерю ДНК-топоизомеразы I Escherichia coli» . Журнал бактериологии . 162 (3): 1173–9. дои : 10.1128/JB.162.3.1173-1179.1985 . ПМК 215900 . ПМИД 2987184 .
- ^ Дин Ф., Краснов М.А., Оттер Р., Мацук М.М., Шпенглер С.Дж., Коццарелли Н.Р. (1983). «Топоизомеразы Escherichia coli типа 1: идентификация, механизм и роль в рекомбинации». Симпозиумы Колд-Спринг-Харбор по количественной биологии . 47 (2): 769–77. дои : 10.1101/sqb.1983.047.01.088 . ПМИД 6305585 .
- ^ Зехидрих Э.Л., Ходурский А.Б., Башелье С., Шнайдер Р., Чен Д., Лилли Д.М., Коцарелли Н.Р. (март 2000 г.). «Роль топоизомераз в поддержании стационарной сверхспирализации ДНК в Escherichia coli» . Журнал биологической химии . 275 (11): 8103–13. дои : 10.1074/jbc.275.11.8103 . ПМИД 10713132 .
- ^ Като Дж., Нисимура Ю., Имамура Р., Ники Х., Хирага С., Сузуки Х. (октябрь 1990 г.). «Новая топоизомераза, необходимая для разделения хромосом в E. coli». Клетка . 63 (2): 393–404. дои : 10.1016/0092-8674(90)90172-б . ПМИД 2170028 . S2CID 42122316 .
- ^ Лю Л.Ф., Ван Дж.К. (октябрь 1987 г.). «Сверхспирализация матрицы ДНК во время транскрипции» . Труды Национальной академии наук Соединенных Штатов Америки . 84 (20): 7024–7. Бибкод : 1987PNAS...84.7024L . дои : 10.1073/pnas.84.20.7024 . ПМК 299221 . ПМИД 2823250 .
- ^ Jump up to: а б Кузин Ф., Лю Дж., Сэнфорд С., Чунг Х.Дж., Левенс Д. (ноябрь 2004 г.). «Динамическая реакция вышележащей ДНК на торсионный стресс, вызванный транскрипцией» . Структурная и молекулярная биология природы . 11 (11): 1092–100. дои : 10.1038/nsmb848 . ПМИД 15502847 . S2CID 28956216 .
- ^ Рувьер-Янив Ж, Янив М, Жермон ЖЕ (июнь 1979 г.). «ДНК-связывающий белок HU E. coli образует нуклеосомоподобную структуру с кольцевой двухцепочечной ДНК». Клетка . 17 (2): 265–74. дои : 10.1016/0092-8674(79)90152-1 . ПМИД 222478 . S2CID 28092421 .
- ^ Бройлс СС, Петтиджон Д.Э. (январь 1986 г.). «Взаимодействие белка HU Escherichia coli с ДНК. Доказательства образования нуклеосомоподобных структур с измененным шагом спирали ДНК». Журнал молекулярной биологии . 187 (1): 47–60. дои : 10.1016/0022-2836(86)90405-5 . ПМИД 3514923 .
- ^ Кундукад Б., Конг П., ван дер Маарел-младший, Дойл П.С. (сентябрь 2013 г.). «Зависящая от времени жесткость изгиба и спиральное скручивание ДНК за счет перестройки связанного белка HU» . Исследования нуклеиновых кислот . 41 (17): 8280–8. дои : 10.1093/нар/gkt593 . ПМЦ 3783175 . ПМИД 23828037 .
- ^ Таппер А.Э., Оуэн-Хьюз Т.А., Уссери Д.В., Сантос Д.С., Фергюсон Д.Д., Сайдботэм Дж.М. и др. (январь 1994 г.). «Связанный с хроматином белок H-NS изменяет топологию ДНК in vitro» . Журнал ЭМБО . 13 (1): 258–68. дои : 10.1002/j.1460-2075.1994.tb06256.x . ПМК 394800 . ПМИД 8306968 .
- ^ Jump up to: а б Шнайдер Р., Лурц Р., Людер Г., Толксдорф С., Трэверс А., Мусхелишвили Г. (декабрь 2001 г.). «Архитектурная роль белка хроматина FIS Escherichia coli в организации ДНК» . Исследования нуклеиновых кислот . 29 (24): 5107–14. дои : 10.1093/нар/29.24.5107 . ПМЦ 97572 . ПМИД 11812843 .
- ^ Шнайдер Р., Трэверс А., Кутателадзе Т., Мусхелишвили Г. (декабрь 1999 г.). «Архитектурный белок ДНК сочетает клеточную физиологию и топологию ДНК в Escherichia coli» . Молекулярная микробиология . 34 (5): 953–64. дои : 10.1046/j.1365-2958.1999.01656.x . ПМИД 10594821 .
- ^ Jump up to: а б Бенсаид А., Алмейда А., Дрлица К., Рувьер-Янив Дж. (февраль 1996 г.). «Перекрестная связь между топоизомеразой I и HU в Escherichia coli». Журнал молекулярной биологии . 256 (2): 292–300. дои : 10.1006/jmbi.1996.0086 . ПМИД 8594197 .
- ^ Малик М., Бенсаид А., Рувьер-Янив Дж., Дрлица К. (февраль 1996 г.). «Гистоноподобный белок HU и топология бактериальной ДНК: подавление дефицита HU за счет мутаций гиразы». Журнал молекулярной биологии . 256 (1): 66–76. дои : 10.1006/jmbi.1996.0068 . ПМИД 8609614 .
- ^ Jump up to: а б Марианс К.Дж. (июль 1987 г.). «ДНК-гираза-катализируемая декатенация многосвязных димеров ДНК» . Журнал биологической химии . 262 (21): 10362–8. дои : 10.1016/S0021-9258(18)61121-4 . ПМИД 3038875 .
- ^ Jump up to: а б с Синден Р.Р., Петтиджон Д.Э. (январь 1981 г.). «Хромосомы в живых клетках Escherichia coli разделены на домены суперспирализации» . Труды Национальной академии наук Соединенных Штатов Америки . 78 (1): 224–8. Бибкод : 1981PNAS...78..224S . дои : 10.1073/pnas.78.1.224 . ПМК 319024 . ПМИД 6165987 .
- ^ Хиггинс Н. П., Ян X, Фу Кью, Рот-младший (май 1996 г.). «Обследование домена суперспирали с использованием системы разрешения гамма-дельта в Salmonella typhimurium» . Журнал бактериологии . 178 (10): 2825–35. дои : 10.1128/jb.178.10.2825-2835.1996 . ПМК 178017 . ПМИД 8631670 .
- ^ Ян Ю, Дин Ю, Ленг Ф, Данлэп Д, Финзи Л (май 2018 г.). «Белковые петли в суперспиральной ДНК создают большие топологические домены» . Исследования нуклеиновых кислот . 46 (9): 4417–4424. дои : 10.1093/nar/gky153 . ПМК 5961096 . ПМИД 29538766 .
- ^ Ленг Ф., Чен Б., Данлэп Д.Д. (декабрь 2011 г.). «Деление сверхспиральной молекулы ДНК на два независимых топологических домена» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (50): 19973–8. Бибкод : 2011PNAS..10819973L . дои : 10.1073/pnas.1109854108 . ПМК 3250177 . ПМИД 22123985 .
- ^ Мулен Л., Рахмуни А.Р., Боккар Ф. (январь 2005 г.). «Топологические изоляторы ингибируют диффузию индуцированных транскрипцией положительных суперспиралей в хромосоме Escherichia coli» . Молекулярная микробиология . 55 (2): 601–10. дои : 10.1111/j.1365-2958.2004.04411.x . ПМИД 15659173 .
- ^ Димри Г.П., Радд К.Е., Морган М.К., Баят Х., Эймс Г.Ф. (июль 1992 г.). «Физическое картирование повторяющихся экстрагенных палиндромных последовательностей в Escherichia coli и филогенетическое распределение среди штаммов Escherichia coli и других кишечных бактерий» . Журнал бактериологии . 174 (14): 4583–93. дои : 10.1128/jb.174.14.4583-4593.1992 . ПМК 206253 . ПМИД 1624447 .
- ^ Дэн С., Штейн Р.А., Хиггинс Н.П. (март 2004 г.). «Вызванные транскрипцией барьеры для диффузии суперспирали в хромосоме Salmonella typhimurium» . Труды Национальной академии наук Соединенных Штатов Америки . 101 (10): 3398–403. Бибкод : 2004PNAS..101.3398D . дои : 10.1073/pnas.0307550101 . ПМЦ 373473 . ПМИД 14993611 .
- ^ Дэн С., Стейн Р.А., Хиггинс Н.П. (сентябрь 2005 г.). «Организация доменов суперспиралей и их реорганизация посредством транскрипции» . Молекулярная микробиология . 57 (6): 1511–21. дои : 10.1111/j.1365-2958.2005.04796.x . ПМК 1382059 . ПМИД 16135220 .
- ^ Букер Б.М., Дэн С., Хиггинс Н.П. (декабрь 2010 г.). «Топология ДНК высокотранскрибируемых оперонов серовара Salmonella enterica Typhimurium» . Молекулярная микробиология . 78 (6): 1348–64. дои : 10.1111/j.1365-2958.2010.07394.x . ПМИД 21143310 .
- ^ Jump up to: а б с д и ж г час я дж к л м н Лиой В.С., Курнак А., Марбути М., Дуигу С., Моцциконаччи Дж., Эспели О. и др. (февраль 2018 г.). «Многомасштабное структурирование хромосомы E. coli с помощью нуклеоид-ассоциированных и конденсиновых белков» . Клетка . 172 (4): 771–783.e18. дои : 10.1016/j.cell.2017.12.027 . ПМИД 29358050 .
- ^ Альмирон М., Линк Эй.Дж., Ферлонг Д., Колтер Р. (декабрь 1992 г.). «Новый ДНК-связывающий белок, играющий регуляторную и защитную роль в голодающей Escherichia coli» . Гены и развитие . 6 (12Б): 2646–54. дои : 10.1101/gad.6.12b.2646 . ПМИД 1340475 . S2CID 10308477 .
- ^ Jump up to: а б Френкиль-Криспин Д., Бен-Авраам И., Ингландер Дж., Шимони Э., Вольф С.Г., Мински А. (январь 2004 г.). «Реструктуризация нуклеоида у стационарных бактерий» . Молекулярная микробиология . 51 (2): 395–405. дои : 10.1046/j.1365-2958.2003.03855.x . ПМИД 14756781 .
- ^ Болл К.А., Осуна Р., Фергюсон К.К., Джонсон Р.К. (декабрь 1992 г.). «Драматические изменения уровней Fis при повышении уровня питательных веществ в Escherichia coli» . Журнал бактериологии . 174 (24): 8043–56. дои : 10.1128/jb.174.24.8043-8056.1992 . ПМК 207543 . ПМИД 1459953 .
- ^ Кларет Л., Рувьер-Янив Ж (октябрь 1997 г.). «Изменение состава HU во время роста Escherichia coli: гетеродимер необходим для длительного выживания». Журнал молекулярной биологии . 273 (1): 93–104. дои : 10.1006/jmbi.1997.1310 . ПМИД 9367749 .
- ^ Jump up to: а б с Бадринараянан А., Лестерлин С., Рейес-Ламот Р., Шерратт Д. (сентябрь 2012 г.). «Комплекс SMC Escherichia coli, MukBEF, формирует организацию нуклеоида независимо от репликации ДНК» . Журнал бактериологии . 194 (17): 4669–76. дои : 10.1128/JB.00957-12 . ПМЦ 3415497 . ПМИД 22753058 .
- ^ Jump up to: а б с Петрушенко З.М., Лай Ч., Рыбенков В.В. (ноябрь 2006 г.). «Антагонистические взаимодействия клейзинов и ДНК с бактериальным конденсином МукБ» . Журнал биологической химии . 281 (45): 34208–17. дои : 10.1074/jbc.M606723200 . ПМЦ 1634889 . ПМИД 16982609 .
- ^ Jump up to: а б с Лал А., Дхар А., Тростел А., Кузин Ф., Сешасаи А.С., Адхья С. (март 2016 г.). «Модель суперспирализации бактериальной хромосомы в масштабе генома» . Природные коммуникации . 7 : 11055. Бибкод : 2016NatCo...711055L . дои : 10.1038/ncomms11055 . ПМЦ 4820846 . ПМИД 27025941 .
- ^ Jump up to: а б Кар С., Эдгар Р., Адхья С. (ноябрь 2005 г.). «Ремоделирование нуклеоида измененным белком HU: реорганизация программы транскрипции» . Труды Национальной академии наук Соединенных Штатов Америки . 102 (45): 16397–402. Бибкод : 2005PNAS..10216397K . дои : 10.1073/pnas.0508032102 . ПМЦ 1283455 . ПМИД 16258062 .
- ^ Лим Си-Джей, Ли С.И., Кенни Л.Дж., Ян Дж. (2012). «Формирование нуклеопротеиновых нитей является структурной основой подавления гена бактериального белка H-NS» . Научные отчеты . 2 : 509. Бибкод : 2012NatSR...2E.509L . дои : 10.1038/srep00509 . ПМК 3396134 . ПМИД 22798986 .
- ^ Аки Т., Чой Х.Э., Адхья С. (февраль 1996 г.). «Гистоноподобный белок HU как специфический регулятор транскрипции: роль кофактора в репрессии транскрипции gal репрессором GAL» . Гены в клетки . 1 (2): 179–88. дои : 10.1046/j.1365-2443.1996.d01-236.x . ПМИД 9140062 .
- ^ Пейгель Дж. М., Винкельман Дж. В., Адамс К. В., Хэтфилд Г. В. (апрель 1992 г.). «Регуляция инициации транскрипции, опосредованная топологией ДНК, от тандемных промоторов оперона ilvGMEDA Escherichia coli». Журнал молекулярной биологии . 224 (4): 919–35. дои : 10.1016/0022-2836(92)90460-2 . ПМИД 1569580 .
- ^ Парех Б.С., Хэтфилд Г.В. (февраль 1996 г.). «Активация транскрипции за счет изгиба ДНК, индуцированного белком: доказательства модели структурной передачи ДНК» . Труды Национальной академии наук Соединенных Штатов Америки . 93 (3): 1173–7. Бибкод : 1996PNAS...93.1173P . дои : 10.1073/pnas.93.3.1173 . ПМК 40051 . ПМИД 8577735 .
- ^ Jump up to: а б с Дорман CJ, Дорман MJ (сентябрь 2016 г.). «Сверхспирализация ДНК является фундаментальным регуляторным принципом контроля экспрессии бактериальных генов» . Биофизические обзоры . 8 (3): 209–220. дои : 10.1007/s12551-016-0205-y . ПМЦ 5425793 . ПМИД 28510224 .
- ^ Чонг С., Чен С., Гэ Х., Се XS (июль 2014 г.). «Механизм транскрипционного разрыва у бактерий» . Клетка . 158 (2): 314–326. дои : 10.1016/j.cell.2014.05.038 . ПМК 4105854 . ПМИД 25036631 .
- ^ Бергер М., Герганова В., Бергер П., Рапитяну Р., Лишиковас В., Добриндт У. (август 2016 г.). «Гены на проводе: нуклеоид-ассоциированный белок HU изолирует единицы транскрипции в Escherichia coli» . Научные отчеты . 6 : 31512. Бибкод : 2016NatSR...631512B . дои : 10.1038/srep31512 . ПМЦ 4992867 . ПМИД 27545593 .
- ^ Коли П., Судан С., Фицджеральд Д., Адья С., Кар С. (2011). «Превращение комменсальной Escherichia coli K-12 в инвазивную форму посредством экспрессии мутантного гистоноподобного белка» . мБио . 2 (5). дои : 10.1128/mBio.00182-11 . ПМК 3172693 . ПМИД 21896677 .
- ^ Jump up to: а б с Кар С., Чой Э.Дж., Го Ф., Димитриадис Е.К., Котова С.Л., Адхья С. (декабрь 2006 г.). «Правосторонняя суперспирализация ДНК октамерной формой гистонподобного белка HU: модуляция клеточной транскрипции» . Журнал биологической химии . 281 (52): 40144–53. дои : 10.1074/jbc.M605576200 . ПМИД 17062578 .
- ^ Jump up to: а б Кар С., Чой Э.Дж., Го Ф., Димитриадис Е.К., Котова С.Л., Адхья С. (декабрь 2006 г.). «Правосторонняя суперспирализация ДНК октамерной формой гистонподобного белка HU: модуляция клеточной транскрипции» . Журнал биологической химии . 281 (52): 40144–53. дои : 10.1074/jbc.M605576200 . ПМИД 17062578 .
- ^ Jump up to: а б Ульманн Ф (июль 2016 г.). «Комплексы SMC: от ДНК к хромосомам». Nature Reviews Молекулярно-клеточная биология . 17 (7): 399–412. дои : 10.1038/номер.2016.30 . ПМИД 27075410 . S2CID 20398243 .
- ^ Деккер Дж., Риппе К., Деккер М., Клекнер Н. (февраль 2002 г.). «Захват конформации хромосомы». Наука . 295 (5558): 1306–11. Бибкод : 2002Sci...295.1306D . дои : 10.1126/science.1067799 . ПМИД 11847345 . S2CID 3561891 .
- ^ Либерман-Эйден Э., ван Беркум Н.Л., Уильямс Л., Имакаев М., Рагочи Т., Теллинг А. и др. (октябрь 2009 г.). «Комплексное картирование долгосрочных взаимодействий раскрывает принципы свертывания человеческого генома» . Наука . 326 (5950): 289–93. Бибкод : 2009Sci...326..289L . дои : 10.1126/science.1181369 . ПМЦ 2858594 . ПМИД 19815776 .
- ^ Jump up to: а б с д Ле ТБ, Имакаев М.В., Мирный Л.А., Лауб М.Т. (ноябрь 2013 г.). «Картирование пространственной организации бактериальной хромосомы высокого разрешения» . Наука . 342 (6159): 731–4. Бибкод : 2013Sci...342..731L . дои : 10.1126/science.1242059 . ПМЦ 3927313 . ПМИД 24158908 .
- ^ Ван X, Ле ТБ, Ладжуа БР, Деккер Дж, Лауб МТ, Руднер ДЗ (август 2015 г.). «Конденсин способствует расположению ДНК, фланкирующей место его загрузки в Bacillus subtilis» . Гены и развитие . 29 (15): 1661–75. дои : 10.1101/gad.265876.115 . ПМЦ 4536313 . ПМИД 26253537 .
- ^ Деккер Дж., Херд Э. (октябрь 2015 г.). «Структурное и функциональное разнообразие топологически ассоциированных областей» . Письма ФЭБС . 589 (20 частей А): 2877–84. Бибкод : 2015FEBSL.589.2877D . дои : 10.1016/j.febslet.2015.08.044 . ПМК 4598308 . ПМИД 26348399 .
- ^ Ники Х., Хирага С. (апрель 1998 г.). «Полярная локализация начала и конца репликации в нуклеоидах Escherichia coli во время разделения хромосом» . Гены и развитие . 12 (7): 1036–45. дои : 10.1101/gad.12.7.1036 . ПМК 316681 . ПМИД 9531540 .
- ^ Ники Х., Ямаичи Ю., Хирага С. (январь 2000 г.). «Динамическая организация хромосомной ДНК кишечной палочки» . Гены и развитие . 14 (2): 212–23. дои : 10.1101/gad.14.2.212 . ПМК 316355 . ПМИД 10652275 .
- ^ Боккар Ф, Эсно Э, Валенс М (июль 2005 г.). «Пространственное расположение и макродоменная организация бактериальных хромосом» . Молекулярная микробиология . 57 (1): 9–16. дои : 10.1111/j.1365-2958.2005.04651.x . ПМИД 15948945 .
- ^ Дуйгу С., Боккар Ф. (май 2017 г.). «Организация хромосом дальнего действия в Escherichia coli: положение начала репликации определяет неструктурированные области, а также правый и левый макродомены» . ПЛОС Генетика . 13 (5): e1006758. дои : 10.1371/journal.pgen.1006758 . ПМЦ 5441646 . ПМИД 28486476 .
- ^ Jump up to: а б с Ноливос С., Аптон А.Л., Бадринараянан А., Мюллер Дж., Завадска К., Виктор Дж. и др. (январь 2016 г.). «MatP регулирует скоординированное действие топоизомеразы IV и MukBEF при сегрегации хромосом» . Природные коммуникации . 7 : 10466. Бибкод : 2016NatCo...710466N . дои : 10.1038/ncomms10466 . ПМЦ 4738335 . ПМИД 26818444 .
- ^ Jump up to: а б Дюпейн П., Тонтат Н.К., Эспели О., Уитфилл Т., Боккар Ф., Шумахер М.А. (ноябрь 2012 г.). «Молекулярная основа белково-опосредованного механизма соединения ДНК, который функционирует при конденсации хромосомы E. coli» . Молекулярная клетка . 48 (4): 560–71. doi : 10.1016/j.molcel.2012.09.009 . ПМЦ 7505563 . ПМИД 23084832 .
- ^ Ван С., Косстик Р., Гарднер Дж. Ф., Гампорт Р. И. (октябрь 1995 г.). «Специфическое связывание фактора хозяина интеграции Escherichia coli включает как большие, так и малые бороздки ДНК». Биохимия . 34 (40): 13082–90. дои : 10.1021/bi00040a020 . ПМИД 7548068 .
- ^ Карась В.О., Вестерлакен И., Мейер А.С. (октябрь 2015 г.). «ДНК-связывающий белок голодающих клеток (Dps) использует двойные функции для защиты клеток от многочисленных стрессов» . Журнал бактериологии . 197 (19): 3206–15. дои : 10.1128/JB.00475-15 . ПМК 4560292 . ПМИД 26216848 .
- ^ Jump up to: а б Ву Дж.С., Лим Дж.Х., Шин Х.К., Су М.К., Ку Б., Ли К.Х. и др. (январь 2009 г.). «Структурные исследования бактериального конденсинового комплекса выявили АТФ-зависимое нарушение межсубъединичных взаимодействий» (PDF) . Клетка . 136 (1): 85–96. дои : 10.1016/j.cell.2008.10.050 . ПМИД 19135891 . S2CID 4608756 .
- ^ Jump up to: а б с д и Бадринараянан А., Рейес-Ламот Р., Упхофф С., Лик MC, Шерратт DJ (октябрь 2012 г.). «Архитектура in vivo и действие бактериального структурного поддержания хромосомных белков» . Наука . 338 (6106): 528–31. Бибкод : 2012Sci...338..528B . дои : 10.1126/science.1227126 . ПМЦ 3807729 . ПМИД 23112333 .
- ^ Кумар Р., Гросбарт М., Медсестра П., Банг С., Вайман К.Л., Марианс К.Дж. (октябрь 2017 г.). «Бактериальный конденсин MukB уплотняет ДНК путем секвестрации суперспиралей и стабилизации топологически изолированных петель» . Журнал биологической химии . 292 (41): 16904–16920. дои : 10.1074/jbc.M117.803312 . ПМК 5641887 . ПМИД 28842486 .
- ^ Ники Х., Имамура Р., Китаока М., Яманака К., Огура Т., Хирага С. (декабрь 1992 г.). «Белок MukB E.coli, участвующий в разделении хромосом, образует гомодимер со структурой стержня и шарнира, обладающий активностью связывания ДНК и связывания АТФ / ГТФ» . Журнал ЭМБО . 11 (13): 5101–9. дои : 10.1002/j.1460-2075.1992.tb05617.x . ПМК 556988 . ПМИД 1464330 .
- ^ Мелби Т.Э., Чампальо К.Н., Бриско Дж., Эриксон Х.П. (сентябрь 1998 г.). «Симметричная структура структурного обеспечения хромосом (SMC) и белков MukB: длинные антипараллельные спиральные спирали, сложенные гибким шарниром» . Журнал клеточной биологии . 142 (6): 1595–604. дои : 10.1083/jcb.142.6.1595 . ПМК 2141774 . ПМИД 9744887 .
- ^ Jump up to: а б Ямадзоэ М., Оноги Т., Сунако Ю., Ники Х., Яманака К., Ичимура Т., Хирага С. (ноябрь 1999 г.). «Комплексное образование белков MukB, MukE и MukF, участвующих в разделении хромосом у Escherichia coli» . Журнал ЭМБО . 18 (21): 5873–84. дои : 10.1093/emboj/18.21.5873 . ПМЦ 1171653 . ПМИД 10545099 .
- ^ Ли Ю, Стюарт Н.К., Бергер А.Дж., Вос С., Шеффлер А.Дж., Бергер Дж.М. и др. (ноябрь 2010 г.). «Конденсин MukB Escherichia coli стимулирует активность топоизомеразы IV путем прямого физического взаимодействия» . Труды Национальной академии наук Соединенных Штатов Америки . 107 (44): 18832–7. дои : 10.1073/pnas.1008678107 . ПМЦ 2973889 . ПМИД 20921377 .
- ^ Вос С.М., Стюарт Н.К., Окли М.Г., Бергер Дж.М. (ноябрь 2013 г.). «Структурная основа взаимодействия MukB-топоизомеразы IV и его функциональные последствия in vivo» . Журнал ЭМБО . 32 (22): 2950–62. дои : 10.1038/emboj.2013.218 . ПМЦ 3832749 . ПМИД 24097060 .
- ^ Николас Э., Аптон А.Л., Упхофф С., Генри О, Бадринараянан А., Шерратт Д. (февраль 2014 г.). «Комплекс SMC MukBEF привлекает топоизомеразу IV к месту начала репликации в живой Escherichia coli» . мБио . 5 (1): e01001-13. дои : 10.1128/mBio.01001-13 . ПМЦ 3950513 . ПМИД 24520061 .
- ^ Завадски П., Стрейси М., Гинда К., Завадска К., Лестерлин С., Капанидис А.Н., Шерратт DJ (декабрь 2015 г.). «Локализация и действие топоизомеразы IV в сегрегации хромосом Escherichia coli координируется комплексом SMC, MukBEF» . Отчеты по ячейкам . 13 (11): 2587–2596. дои : 10.1016/j.celrep.2015.11.034 . ПМК 5061553 . ПМИД 26686641 .
- ^ Jump up to: а б с Бакстер Дж., Оливер А.В., Шалбеттер С.А. (январь 2019 г.). «Являются ли комплексы SMC факторами выдавливания петли? Связь теории с фактом» . Биоэссе . 41 (1): e1800182. doi : 10.1002/bies.201800182 . ПМИД 30506702 .
- ^ Завадска К., Завадски П., Бейкер Р., Раджасекар К.В., Вагнер Ф., Шерратт Дж., Арчишевска Л.К. (январь 2018 г.). «АТФазы MukB регулируются независимо N- и C-концевыми доменами клейзина MukF» . электронная жизнь . 7 . дои : 10.7554/eLife.31522 . ПМК 5812716 . ПМИД 29323635 .
- ^ Савицке Дж. А., Остин С. (февраль 2000 г.). «Подавление дефектов сегрегации хромосом мутантов Escherichia coli muk мутациями топоизомеразы I» . Труды Национальной академии наук Соединенных Штатов Америки . 97 (4): 1671–6. Бибкод : 2000PNAS...97.1671S . дои : 10.1073/pnas.030528397 . ПМК 26494 . ПМИД 10660686 .
- ^ Адачи С., Хирага С. (июль 2003 г.). «Мутанты, подавляющие гиперчувствительность к новобиоцину нулевой мутации mukB» . Журнал бактериологии . 185 (13): 3690–5. дои : 10.1128/jb.185.13.3690-3695.2003 . ПМК 161581 . ПМИД 12813060 .
- ^ Петрушенко З.М., Лай Ч., Рай Р., Рыбенков В.В. (февраль 2006 г.). «Реформирование ДНК МукБ. Правостороннее завязывание, левостороннее суперспирализация» . Журнал биологической химии . 281 (8): 4606–15. дои : 10.1074/jbc.M504754200 . ПМЦ 1633270 . ПМИД 16368697 .
- ^ Гаал Т., Брэттон Б.П., Санчес-Васкес П., Сливицки А., Сливицки К., Вегель А. и др. (октябрь 2016 г.). «Колокализация отдаленных хромосомных локусов в пространстве у E. coli: бактериальное ядрышко» . Гены и развитие . 30 (20): 2272–2285. дои : 10.1101/gad.290312.116 . ПМК 5110994 . ПМИД 27898392 .
- ^ Джин DJ, Кабрера Дж. Э. (ноябрь 2006 г.). «Связь распределения РНК-полимеразы с глобальной регуляцией генов и динамической структурой бактериального нуклеоида в Escherichia coli». Журнал структурной биологии . 156 (2): 284–91. дои : 10.1016/j.jsb.2006.07.005 . ПМИД 16934488 .
- ^ Jump up to: а б с Цянь З., Димитриадис Е.К., Эдгар Р., Эсварамурти П., Адхья С. (июль 2012 г.). «Репрессор галактозы опосредует межсегментарные хромосомные связи в Escherichia coli» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (28): 11336–41. Бибкод : 2012PNAS..10911336Q . дои : 10.1073/pnas.1208595109 . ПМЦ 3396475 . ПМИД 22733746 .
- ^ Вейкерт М.Дж., Адхья С. (октябрь 1993 г.). «Галактозный регулон кишечной палочки» . Молекулярная микробиология . 10 (2): 245–51. дои : 10.1111/j.1365-2958.1993.tb01950.x . ПМИД 7934815 . S2CID 6872903 .
- ^ Jump up to: а б Агершу Э.Д., Кристиансен Г., Шафер Н.П., Мэдсен Д.Д., Бродерсен Д.Е., Семси С., Отцен Д.Е. (июнь 2016 г.). «Регулятор транскрипции GalR самособирается с образованием очень регулярных трубчатых структур» . Научные отчеты . 6 : 27672. Бибкод : 2016NatSR...627672A . дои : 10.1038/srep27672 . ПМЦ 4899725 . ПМИД 27279285 .
- ^ Кавенофф Р., Райдер О.А. (март 1976 г.). «Электронная микроскопия мембранассоциированных складчатых хромосом Escherichia coli». Хромосома . 55 (1): 13–25. дои : 10.1007/bf00288323 . ПМИД 767075 . S2CID 31879250 .
- ^ Ворсель А., Бурги Э. (январь 1974 г.). «Свойства мембраносвязанной формы свернутой хромосомы Escherichia coli». Журнал молекулярной биологии . 82 (1): 91–105. дои : 10.1016/0022-2836(74)90576-2 . ПМИД 4594427 .
- ^ Роджиани М., Гулиан М. (2015). «Хромосомно-мембранные взаимодействия у бактерий». Ежегодный обзор генетики . 49 : 115–29. doi : 10.1146/annurev-genet-112414-054958 . ПМИД 26436460 .
- ^ Либби Э.А., Роджиани М., Гулиан М. (май 2012 г.). «Экспрессия мембранных белков вызывает репозицию хромосомных локусов у бактерий» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (19): 7445–50. Бибкод : 2012PNAS..109.7445L . дои : 10.1073/pnas.1109479109 . ПМЦ 3358875 . ПМИД 22529375 .
- ^ Брамейер С., Рёш Т.С., Эль Андари Дж., Хойер Э., Шварц Дж., Грауманн П.Л., Юнг К. (2019). «ДНК-связывание управляет локализацией встроенного в мембрану рецептора семейства ToxR» . Коммуникационная биология . 2 : 4. дои : 10.1038/s42003-018-0248-7 . ПМК 6320335 . ПМИД 30740540 .
- ^ Эспели О, Борн Р, Дюпейн П, Тиль А, Гигант Э, Мерсье Р, Боккар Ф (май 2012 г.). «Взаимодействие MatP-дивизомы координирует сегрегацию хромосом с делением клеток в E. coli» . Журнал ЭМБО . 31 (14): 3198–211. дои : 10.1038/emboj.2012.128 . ПМК 3400007 . ПМИД 22580828 .
- ^ Бернхардт Т.Г., де Бур П.А. (май 2005 г.). «SlmA, нуклеоид-ассоциированный белок, связывающий FtsZ, необходимый для блокирования сборки перегородочного кольца на хромосомах в E. coli» . Молекулярная клетка . 18 (5): 555–64. doi : 10.1016/j.molcel.2005.04.012 . ПМЦ 4428309 . ПМИД 15916962 .
- ^ Волдринг К.Л., Йенсен П.Р., Вестерхофф Х.В. (сентябрь 1995 г.). «Структура и разделение бактериальной ДНК: определяются балансом сил сжатия и расширения?» (PDF) . Письма FEMS по микробиологии . 131 (3): 235–42. дои : 10.1111/j.1574-6968.1995.tb07782.x . ПМИД 7557335 .
- ^ Jump up to: а б Ван Хельворт Дж. М., Халс П.Г., Вишер Н.О., Уолдринг К.Л. (май 1998 г.). «Слитые нуклеоиды ресегрегируют быстрее, чем удлинение клеток в нитях pbpB (Ts) Escherichia coli после освобождения от ингибирования хлорамфениколом» . Микробиология . 144 (5): 1309–17. дои : 10.1099/00221287-144-5-1309 . ПМИД 9611806 .
- ^ Ельцов Михаил; Зубер, Бенуа (2006). «Просвечивающая электронная микроскопия бактериального нуклеоида». Журнал структурной биологии . 156 (2): 246–254. дои : 10.1016/j.jsb.2006.07.007 . ISSN 1047-8477 . ПМИД 16978880 .
- ^ Робинов, К; Келленбергер, Э. (1994). «Возвращение к бактериальному нуклеоиду» . Микробиологические обзоры . 58 (2): 211–232. doi : 10.1128/mmbr.58.2.211-232.1994 . ISSN 0146-0749 . ПМК 372962 . ПМИД 7521510 .
- ^ Смит Б.Т., Гроссман А.Д., Уокер Г.С. (январь 2002 г.). «Локализация UvrA и влияние повреждения ДНК на хромосому Bacillus subtilis» . Журнал бактериологии . 184 (2): 488–93. дои : 10.1128/jb.184.2.488-493.2002 . ПМК 139587 . ПМИД 11751826 .
- ^ Одсбу I, Скарстад К (октябрь 2009 г.). «Снижение активности рибонуклеотидредуктазы замедляет репликационную вилку хромосомы, но не меняет ее локализацию» . ПЛОС ОДИН . 4 (10): е7617. Бибкод : 2009PLoSO...4.7617O . дои : 10.1371/journal.pone.0007617 . ПМЦ 2773459 . ПМИД 19898675 .
- ^ Левин-Зайдман С., Френкиль-Криспин Д., Шимони Е., Сабанай И., Вольф С.Г., Мински А. (июнь 2000 г.). «Упорядоченные внутриклеточные сборки RecA-ДНК: потенциальное место активности, опосредованной RecA in vivo» . Труды Национальной академии наук Соединенных Штатов Америки . 97 (12): 6791–6. Бибкод : 2000PNAS...97.6791L . дои : 10.1073/pnas.090532397 . ЧВК 18741 . ПМИД 10829063 .
- ^ Jump up to: а б Дельмас С., Даггин И.Г., Аллерс Т. (январь 2013 г.). «Повреждение ДНК вызывает уплотнение нуклеоида посредством комплекса Mre11-Rad50 у археи Haloferax volcanii» . Молекулярная микробиология . 87 (1): 168–79. дои : 10.1111/mmi.12091 . ПМЦ 3565448 . ПМИД 23145964 .