Ласса маммаренавирус
| Ласса маммаренавирус | |
|---|---|
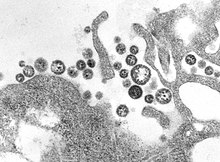
| |
| TEM- микрофотография маммаренавируса Ласса вирионов | |
| Классификация вирусов | |
| (без рейтинга): | Вирус |
| Область : | Рибовирия |
| Королевство: | Орторнавиры |
| Тип: | Негарнавирикота |
| Сорт: | Эллиовирицетес |
| Заказ: | Буньявирусы |
| Семья: | Аренавирусиды |
| Род: | маммаренавирус |
| Разновидность: | Ласса маммаренавирус
|
| Синонимы | |
| |
Вирус Ласса маммарена ( LASV ) — аренавирус , вызывающий геморрагическую лихорадку Ласса . [ 1 ] разновидность вирусной геморрагической лихорадки (ВГЛ) у человека и других приматов . Вирус Ласса маммарена — это новый вирус и избранный агент , требующий сдерживания, эквивалентного 4-му уровню биобезопасности . Он эндемичен в странах Западной Африки, особенно в Сьерра-Леоне , Гвинейской Республике , Нигерии и Либерии , где ежегодная заболеваемость инфекцией составляет от 300 000 до 500 000 случаев, что приводит к 5 000 смертей в год. [ 2 ]
По состоянию на 2012 год открытия в регионе реки Мано в Западной Африке расширили эндемическую зону между двумя известными эндемичными регионами Ласса, что указывает на то, что LASV более широко распространен в экозоне тропических лесистых саванн в Западной Африке. [ 3 ] Не существует одобренных вакцин против лихорадки Ласса для использования у людей. [ 4 ]
Открытие
[ редактировать ]В 1969 году медсестра-миссионерка Лаура Вайн заболела загадочной болезнью, которую она заразилась от акушерской пациентки в деревне Ласса в штате Борно , Нигерия. [ 5 ] [ 6 ] [ 1 ] Затем ее перевезли в Джос , где она умерла. Впоследствии заразились еще двое, одной из которых была пятидесятидвухлетняя медсестра Лили Пиннео, которая ухаживала за Лорой Вайн. [ 7 ] Образцы из Pinneo были отправлены в Йельский университет в Нью-Хейвене , где новый вирус, который позже будет известен как Lassa mammarenavirus , был впервые выделен Хорди Казальс-Ариет , Соней Бакли и другими. [ 8 ] [ 9 ] [ 10 ] Казальс заболел лихорадкой и чуть не погиб; от этого умер один техник. [ 8 ] К 1972 году было обнаружено, что многосоставная крыса Mastomys natalensis является основным резервуаром вируса в Западной Африке и способна выделять вирус с мочой и фекалиями, не проявляя видимых симптомов. [ 11 ] [ 12 ]
Вирусология
[ редактировать ]Структура и геном
[ редактировать ]
Вирусы Ласса [ 14 ] [ 15 ] представляют собой оболочечные одноцепочечные двусегментированные РНК- содержащие вирусы . Их геном [ 16 ] содержится в двух сегментах РНК, каждый из которых кодирует по два белка, по одному в каждом смысле, всего четыре вирусных белка. [ 17 ] Большой сегмент кодирует небольшой белок цинковых пальцев (Z), который регулирует транскрипцию и репликацию. [ 18 ] [ 19 ] и РНК-полимераза (L). Небольшой сегмент кодирует нуклеопротеин (NP) и предшественник поверхностного гликопротеина (GP, также известный как вирусный шип ), который протеолитически расщепляется на гликопротеины оболочки GP1 и GP2, которые связываются с рецептором альфа-дистрогликана и опосредуют проникновение в клетку-хозяина. [ 20 ]
Лихорадка Ласса вызывает геморрагическую лихорадку, часто проявляющуюся иммуносупрессией. Вирус Ласса маммарена размножается очень быстро и демонстрирует временной контроль репликации. [ 21 ] Первым этапом репликации является транскрипция копий мРНК отрицательного или минус-смыслового генома. Это обеспечивает адекватное снабжение вирусными белками для последующих этапов репликации, поскольку белки NP и L транслируются с мРНК. Геном с положительным или плюсовым смыслом затем создает вирусной комплементарной РНК (вкРНК) копии себя . Копии РНК представляют собой матрицу для производства потомства с отрицательным смыслом, но из них также синтезируется мРНК. мРНК, синтезированная из вкРНК, транслируется с образованием белков GP и Z. Этот временной контроль позволяет белкам-шипам производиться последними и, следовательно, задерживать распознавание иммунной системой хозяина. [ нужна ссылка ]
Нуклеотидные исследования генома показали, что Ласса имеет четыре линии: три обнаружены в Нигерии, а четвертая - в Гвинее, Либерии и Сьерра-Леоне. Нигерийские штаммы, вероятно, были предками других, но для подтверждения этого необходимы дополнительные исследования. [ 22 ]
Рецепторы
[ редактировать ]
Вирус Ласса маммарена проникает в клетку-хозяина посредством рецептора клеточной поверхности альфа-дистрогликана (альфа-DG). [ 20 ] универсальный рецептор белков внеклеточного матрикса . Он разделяет этот рецептор с прототипом аренавируса Старого Света, вируса лимфоцитарного хориоменингита . Распознавание рецептора зависит от специфической модификации сахара альфа-дистрогликана группой гликозилтрансфераз, известных как БОЛЬШИЕ белки. Конкретные варианты генов, кодирующих эти белки, по-видимому, находятся под положительным отбором в Западной Африке , где Ласса является эндемиком. [ 23 ] Альфа-дистрогликан также используется в качестве рецептора вирусами аренавирусов клады C Нового Света (вирусы Oliveros и Latino). Напротив, аренавирусы Нового Света клад A и B, которые включают важные вирусы Machupo , Guanarito , Junin и Sabia в дополнение к непатогенному вирусу Amapari, используют рецептор трансферрина 1 . Небольшая алифатическая аминокислота в положении 260 аминокислоты гликопротеина GP1 необходима для высокоаффинного связывания с альфа-DG. Кроме того, положение 259 аминокислоты GP1 также представляется важным, поскольку все аренавирусы, демонстрирующие высокоаффинное связывание с альфа-DG, содержат в этом положении объемистую ароматическую аминокислоту (тирозин или фенилаланин). [ нужна ссылка ]
В отличие от большинства вирусов с оболочкой, которые используют покрытые клатрином ямки для проникновения в клетку и связываются со своими рецепторами в зависимости от pH, вирус Ласса и лимфоцитарного хориоменингита вместо этого используют эндоцитотический путь, независимый от клатрина, кавеолина , динамина и актина . Оказавшись внутри клетки, вирусы быстро доставляются в эндосомы посредством везикулярного транспорта, хотя и в значительной степени независимого от малых ГТФаз Rab5 и Rab7 . При контакте с эндосомой рН-зависимое слияние мембран происходит при помощи гликопротеина оболочки, который при более низком рН эндосомы связывает лизосомальный белок LAMP1, что приводит к слиянию мембран и выходу из эндосомы. [ нужна ссылка ]
Жизненный цикл
[ редактировать ]
Жизненный цикл маммаренавируса Ласса аналогичен аренавирусам Старого Света. Вирус Ласса маммарена проникает в клетку путем рецептор-опосредованного эндоцитоза . Какой путь эндоцитоза используется, пока неизвестно, но, по крайней мере, проникновение в клетку чувствительно к истощению холестерина. Сообщалось, что интернализация вируса ограничивается при истощении холестерина. Рецептором, используемым для проникновения в клетку, является альфа- дистрогликан , высококонсервативный и повсеместно экспрессируемый рецептор клеточной поверхности для белков внеклеточного матрикса. Дистрогликан, который позже расщепляется на альфа-дистрогликан и бета-дистрогликан, первоначально экспрессируется в большинстве клеток до зрелых тканей и обеспечивает молекулярную связь между ЕСМ и цитоскелетом на основе актина. [ 24 ] После того, как вирус проникает в клетку путем эндоцитоза, опосредованного альфа-дистрогликаном, среда с низким pH вызывает pH-зависимое слияние мембран и высвобождает комплекс RNP (вирусный рибонуклеопротеин) в цитоплазму. Вирусная РНК распаковывается, и в цитоплазме инициируются репликация и транскрипция. [ 24 ] Когда начинается репликация, геномы S и L РНК синтезируют антигеномные S и L РНК, а из антигеномных РНК синтезируются геномные S и L РНК. Как геномные, так и антигеномные РНК необходимы для транскрипции и трансляции . S-РНК кодирует белки GP и NP (вирусный нуклеокапсидный белок), тогда как L-РНК кодирует белки Z и L. Белок L, скорее всего, представляет собой вирусную РНК-зависимую РНК-полимеразу. [ 25 ] При заражении клетки вирусом L-полимераза связывается с вирусным РНП и инициирует транскрипцию геномной РНК. 5'- и 3'-концевые 19-нуклеотидные области вирусного промотора обоих сегментов РНК необходимы для узнавания и связывания вирусной полимеразы . Первичная транскрипция сначала транскрибирует мРНК из геномных S- и L-РНК, которые кодируют белки NP и L соответственно. Транскрипция заканчивается на структуре «стебель-петля» (SL) внутри межгеномной области. Аренавирусы используют стратегию захвата кэпа для получения кэп-структур из клеточных мРНК, и это опосредовано эндонуклеазной активностью L-полимеразы и кэп-связывающей активностью NP. Антигеномная РНК транскрибирует вирусные гены GPC и Z, закодированные в геномной ориентации, из S- и L-сегментов соответственно. Антигеномная РНК также служит матрицей для репликации. [ 4 ] После трансляции GPC он посттрансляционно модифицируется в эндоплазматическом ретикулуме . GPC расщепляется на GP1 и GP2 на более поздней стадии секреторного пути. Сообщалось, что клеточная протеаза за это расщепление отвечает SKI-1/S1P. Расщепленные гликопротеины включаются в оболочку вириона , когда вирус отпочковывается, и высвобождаются из клеточной мембраны. [ 25 ]
Патогенез
[ редактировать ]
Лихорадка Ласса вызывается вирусом Ласса маммарена . Симптомы включают гриппоподобное заболевание, характеризующееся лихорадкой, общей слабостью, кашлем, болью в горле, головной болью и желудочно-кишечными проявлениями. Геморрагические проявления включают проницаемость сосудов. [ 4 ]
При проникновении маммаренавирус Ласса поражает почти все ткани человеческого организма. Он начинается со слизистой оболочки , кишечника, легких и мочевыделительной системы, а затем переходит в сосудистую систему. [ 5 ]
Основными мишенями вируса являются антигенпрезентирующие клетки , преимущественно дендритные клетки и эндотелиальные клетки. [ 26 ] [ 27 ] [ 28 ] В 2012 году сообщалось, что нуклеопротеин (NP) вируса маммарена Ласса хозяина саботирует реакцию врожденной иммунной системы . Как правило, когда патоген попадает в организм хозяина, врожденная защитная система распознает молекулярные паттерны, связанные с патогеном (PAMP), и активирует иммунный ответ. Один из механизмов обнаруживает двухцепочечную РНК (дцРНК), которая синтезируется только вирусами с отрицательным смыслом . В цитоплазме рецепторы дцРНК, такие как RIG-I (ген I, индуцируемый ретиноевой кислотой) и MDA-5 (ген 5, связанный с дифференцировкой меланомы), обнаруживают дцРНК и инициируют сигнальные пути , которые транслоцируют IRF-3 ( регуляторный фактор интерферона 3) и другие факторы транскрипции в ядро. Транслоцированные факторы транскрипции активируют экспрессию интерферонов 𝛂 и 𝛃, и они инициируют адаптивный иммунитет . NP, кодируемый вирусом Ласса маммаренавируса, важен для репликации и транскрипции вируса, но он также подавляет врожденный ответ IFN хозяина путем ингибирования транслокации IRF-3. НП маммаренавируса Ласса Сообщается, что он обладает экзонуклеазной активностью только в отношении дцРНК. [ 29 ] Активность экзонуклеазы NP dsRNA противодействует ответам IFN, расщепляя PAMP, что позволяет вирусу уклоняться от иммунных ответов хозяина. [ 30 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б Фрейм JD, Болдуин JM, Gocke DJ, Troup JM (июль 1970 г.). «Лихорадка Ласса, новое вирусное заболевание человека из Западной Африки. I. Клиническое описание и патологоанатомические данные». Американский журнал тропической медицины и гигиены . 19 (4): 670–6. дои : 10.4269/ajtmh.1970.19.670 . ПМИД 4246571 .
- ^ «Лихорадка Ласса» . www.who.int .
- ^ Согоба Н., Фельдманн Х., Сафронец Д. (сентябрь 2012 г.). «Лихорадка Ласса в Западной Африке: свидетельства расширения региона эндемичности» . Зоонозы и общественное здравоохранение . 59 Приложение 2 (59): 43–7. дои : 10.1111/j.1863-2378.2012.01469.x . ПМИД 22958249 . S2CID 27063493 .
- ^ Jump up to: а б с Юн Н.Э., Уокер Д.Х. (октябрь 2012 г.). «Патогенез лихорадки Ласса» . Вирусы . 4 (10): 2031–48. дои : 10.3390/v4102031 . ПМК 3497040 . ПМИД 23202452 .
- ^ Jump up to: а б Дональдсон Р.И. (2009). Палата Ласса: Борьба одного человека с одной из самых смертоносных болезней в мире . Пресса Святого Мартина. ISBN 978-0-312-37700-7 .
- ^ «Лихорадка Ласса | CDC» . www.cdc.gov . Проверено 23 сентября 2016 г.
- ^ Фрейм JD (май 1992 г.). «История лихорадки Ласса. Часть I: Открытие болезни». Медицинский журнал штата Нью-Йорк . 92 (5): 199–202. ПМИД 1614671 .
- ^ Jump up to: а б Проно Л (9 января 2008 г.). Чжан Ю (ред.). Энциклопедия глобального здравоохранения Том. 1. МУДРЕЦ. п. 354. ИСБН 978-1-4129-4186-0 . OCLC 775277696 .
- ^ Бакли С.М. , Казальс Дж., Даунс В.Г. (июль 1970 г.). «Выделение и антигенная характеристика вируса Ласса» . Природа . 227 (5254): 174. Бибкод : 1970Natur.227..174B . дои : 10.1038/227174a0 . ПМИД 5428406 . S2CID 4211129 .
- ^ Оранский I (12 марта 2005 г.). «Соня Бакли» . Ланцет . 365 (9463): 932. doi : 10.1016/S0140-6736(05)71068-0 . ПМИД 15786567 . S2CID 5188080 .
- ^ Фрейзер Д.В., Кэмпбелл CC, Монат Т.П., Гофф П.А., Грегг М.Б. (ноябрь 1974 г.). «Лихорадка Ласса в Восточной провинции Сьерра-Леоне, 1970–1972 годы. I. Эпидемиологические исследования». Американский журнал тропической медицины и гигиены . 23 (6): 1131–9. дои : 10.4269/ajtmh.1974.23.1131 . ПМИД 4429182 .
- ^ Монат Т.П., Махер М., Казальс Дж., Кисслинг Р.Э., Каччапуоти А. (ноябрь 1974 г.). «Лихорадка Ласса в Восточной провинции Сьерра-Леоне, 1970–1972 годы. II. Клинические наблюдения и вирусологические исследования отдельных больничных случаев». Американский журнал тропической медицины и гигиены . 23 (6): 1140–9. дои : 10.4269/ajtmh.1974.23.1140 . ПМИД 4429183 .
- ^ Jump up to: а б Фелинг С.К., Леннарц Ф., Стрекер Т. (ноябрь 2012 г.). «Многофункциональная природа белка Z RING-пальца аренавируса» . Вирусы . 4 (11): 2973–3011. дои : 10.3390/v4112973 . ПМК 3509680 . ПМИД 23202512 .
- ↑ Джейми Дьял и Бен Фонер Вирус Ласса, курс «Человек и вирусы» Стэнфордского университета, 2005 г., по состоянию на 9 мая 2018 г.
- ^ Лэшли, Фелисса Р. и Джерри Д. Дарем. Новые инфекционные заболевания: тенденции и проблемы. Нью-Йорк: Springer Pub., 2002. Печать.
- ^ Ридли, Мэтт. Геном: автобиография вида в 23 главах. Нью-Йорк: HarperCollins, 1999. Печать.
- ^ «Геном RefSeq вируса Ласса» .
- ^ Корню Т.И., де ла Торре Дж.К. (октябрь 2001 г.). «Белок Z RING Finger вируса лимфоцитарного хориоменингита (LCMV) ингибирует транскрипцию и репликацию РНК минигенома S-сегмента LCMV» . Журнал вирусологии . 75 (19): 9415–26. doi : 10.1128/JVI.75.19.9415-9426.2001 . ПМК 114509 . ПМИД 11533204 .
- ^ Джавани М., Лукашевич И.С., Санчес А., Николь С.Т., Сальвато М.С. (сентябрь 1997 г.). «Завершение последовательности вируса лихорадки Ласса и идентификация открытой рамки считывания безымянного пальца на 5'-конце L-РНК» . Вирусология . 235 (2): 414–8. дои : 10.1006/viro.1997.8722 . ПМИД 9281522 .
- ^ Jump up to: а б Цао В., Генри М.Д., Борроу П., Ямада Х., Элдер Дж.Х., Равков Е.В. и др. (декабрь 1998 г.). «Идентификация альфа-дистрогликана как рецептора вируса лимфоцитарного хориоменингита и вируса лихорадки Ласса». Наука . 282 (5396): 2079–81. Бибкод : 1998Sci...282.2079C . дои : 10.1126/science.282.5396.2079 . ПМИД 9851928 .
- ^ Лэшли Ф (2002). Тенденции и проблемы новых инфекционных заболеваний . Издательская компания Спрингер.
- ^ Боуэн М.Д., Роллин П.Е., Ксиазек Т.Г., Хустад Х.Л., Бауш Д.Г., Демби А.Х. и др. (август 2000 г.). «Генетическое разнообразие штаммов вируса Ласса» . Журнал вирусологии . 74 (15): 6992–7004. doi : 10.1128/JVI.74.15.6992-7004.2000 . ПМК 112216 . ПМИД 10888638 .
- ^ «Эндемик: Медицинская энциклопедия MedlinePlus» .
- ^ Jump up to: а б Ройек Дж. М., Кунц С. (апрель 2008 г.). «Проникновение в клетку патогенных аренавирусов человека» . Клеточная микробиология . 10 (4): 828–35. дои : 10.1111/j.1462-5822.2007.01113.x . ПМИД 18182084 .
- ^ Jump up to: а б Дростен С., Кюммерер Б.М., Шмитц Х., Гюнтер С. (январь 2003 г.). «Молекулярная диагностика вирусных геморрагических лихорадок». Противовирусные исследования . 57 (1–2): 61–87. дои : 10.1016/s0166-3542(02)00201-2 . ПМИД 12615304 .
- ^ Левен М.И., Гибсон Н.А., Фентон А.С., Папатома Э., Барнетт Д. (июль 1990 г.). «Использование блокатора кальциевых каналов никардипина у новорожденных с тяжелой асфиксией». Медицина развития и детская неврология . 32 (7): 567–74. дои : 10.1111/j.1469-8749.1990.tb08540.x . ПМИД 2391009 . S2CID 38807245 .
- ^ Маханти С., Хатчинсон К., Агарвал С., Макрей М., Роллин П.Е., Пулендран Б. (март 2003 г.). «Авангард: повреждение дендритных клеток и адаптивного иммунитета вирусами Эбола и Ласса» . Журнал иммунологии . 170 (6): 2797–801. дои : 10.4049/jimmunol.170.6.2797 . ПМИД 12626527 .
- ^ Байз С., Каплон Дж., Фор С., Паннетье Д., Жорж-Курбо MC, Дойбель В. (март 2004 г.). «Инфекция вирусом Ласса дендритных клеток и макрофагов человека продуктивна, но не активирует клетки» . Журнал иммунологии . 172 (5): 2861–9. дои : 10.4049/jimmunol.172.5.2861 . ПМИД 14978087 .
- ^ Хасти К.М., Кинг Л.Б., Зандонатти М.А., Сапфир Э.О. (август 2012 г.). «Структурная основа специфичности дцРНК экзонуклеазы NP вируса Ласса» . ПЛОС ОДИН . 7 (8): е44211. Бибкод : 2012PLoSO...744211H . дои : 10.1371/journal.pone.0044211 . ПМЦ 3429428 . ПМИД 22937163 .
- ^ Хасти К.М., Бэйл С., Кимберлин С.Р., Сапфир Э.О. (апрель 2012 г.). «Сокрытие доказательств: две стратегии уклонения от врожденного иммунитета вирусами геморрагической лихорадки» . Современное мнение в вирусологии . 2 (2): 151–6. дои : 10.1016/j.coviro.2012.01.003 . ПМЦ 3758253 . ПМИД 22482712 .