Сома (биология)
Эта статья нуждается в дополнительных цитатах для проверки . ( декабрь 2008 г. ) |

В клеточной нейробиологии сома « тело ( мн .: сомата или сомы ; от греческого σῶμα (sôma) »), перикарион ( мн .: перикария ), нейроцитон или тело клетки — это луковицеобразная, не отростчатая часть нейрона или другой тип клеток головного мозга , содержащий клеточное ядро . Хотя его часто используют для обозначения нейронов, оно также может относиться и к другим типам клеток, включая астроциты . [1] олигодендроциты , [2] и микроглия . [3] Существует много различных специализированных типов нейронов, и их размеры варьируются от 5 микрометров до более 10 миллиметров для некоторых самых маленьких и самых больших нейронов беспозвоночных соответственно.
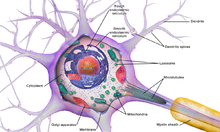
Сома нейрона (т.е. основная часть нейрона, от которой дендриты отходят ) содержит множество органелл , в том числе гранулы, называемые гранулами Ниссля , которые состоят в основном из шероховатой эндоплазматической сети и свободных полирибосом . [4] Ядро клетки является ключевой особенностью сомы. Ядро является источником большей части РНК , вырабатываемой в нейронах. В общем, большинство белков производятся из мРНК , которые не уходят далеко от ядра клетки. Это создает проблему доставки новых белков к окончаниям аксонов , которые могут находиться на расстоянии метра и более от сомы. Аксоны содержат микротрубочками связанные с моторные белки, , которые транспортируют белоксодержащие везикулы между сомой и синапсами на окончаниях аксонов . Такой транспорт молекул к соме и от нее поддерживает критические функции клеток. В случае нейронов сома получает большое количество тормозных синапсов, [5] которые могут регулировать активность этих клеток. Также было показано, что микроглиальные процессы постоянно контролируют функции нейронов через соматические соединения и при необходимости оказывают нейропротекцию. [6]
Аксонный холмик представляет собой специализированный участок тела нейрональной клетки, из которого происходит аксон. В этой области происходит большой синтез белка, поскольку она содержит множество гранул Ниссля (которые представляют собой рибосомы, обернутые RER ) и полирибосомы. Внутри аксонного бугорка материалы сортируются как элементы, которые войдут в аксон (например, компоненты цитоскелетной архитектуры аксона, митохондрии и т. д.) или останутся в соме. Кроме того, аксонный холмик также имеет специализированную плазматическую мембрану, содержащую большое количество потенциалзависимых ионных каналов, поскольку именно здесь чаще всего инициируется и запускается потенциал действия . [4]
Выживание некоторых сенсорных нейронов зависит от того, вступают ли окончания аксонов в контакт с источниками факторов выживания, которые предотвращают апоптоз . Факторами выживания являются нейротрофические факторы , включая такие молекулы, как фактор роста нервов (NGF). NGF взаимодействует с рецепторами на окончаниях аксона, и это производит сигнал, который должен быть транспортирован по аксону к ядру. Современная теория того, как такие сигналы выживания передаются от окончаний аксонов к соме, включает идею о том, что рецепторы NGF эндоцитозируются с поверхности кончиков аксонов и что такие эндоцитозные везикулы транспортируются вверх по аксону. [7]
Промежуточные филаменты широко распространены как в перикариях, так и в аксональных и дендритных отростках и называются нейрофиламентами . Нейрофиламенты сшиваются определенными фиксаторами и, пропитываясь серебром, образуют нейрофибриллы, видимые в световой микроскоп. [8]
Ссылки
[ редактировать ]- ^ Базаргани, Н; Эттвелл, Д. (февраль 2016 г.). «Передача сигналов кальция астроцитами: третья волна» . Природная неврология . 19 (2): 182–9. дои : 10.1038/nn.4201 . ПМИД 26814587 . S2CID 205438341 .
- ^ Бауманн, Н; Фам-Динь, Д. (апрель 2001 г.). «Биология олигодендроцитов и миелина в центральной нервной системе млекопитающих». Физиологические обзоры . 81 (2): 871–927. дои : 10.1152/physrev.2001.81.2.871 . ПМИД 11274346 .
- ^ Козловский, К; Веймер, Р.М. (2012). «Автоматизированный метод количественной оценки морфологии микроглии и его применение для продольного мониторинга состояния активации in vivo» . ПЛОС ОДИН . 7 (2): e31814. Бибкод : 2012PLoSO...731814K . дои : 10.1371/journal.pone.0031814 . ПМЦ 3294422 . ПМИД 22457705 .
- ^ Jump up to: а б Сквайр, Ларри; Берг, Дарвин; Блум, Флойд; дю Лак, Саша; Гош, Анирван; Спитцер, Николас, ред. (2008). Фундаментальная нейронаука (3-е изд.). Академическая пресса. ISBN 978-0-12-374019-9 .
- ^ Фрейнд и Катона, Neuron, том 56, выпуск 1, 4 октября 2007 г., страницы 33–42, https://doi.org/10.1016/j.neuron.2007.09.012
- ^ Череп С., Посфаи Б., Ленарт Н., Фекете Р., Ласло З.И., Леле З. и др. (январь 2020 г.). «Микроглия контролирует и защищает функцию нейронов через специализированные соматические пуринергические соединения» (PDF) . Наука . 367 (6477): 528–537. Бибкод : 2020Sci...367..528C . doi : 10.1126/science.aax6752 . ПМИД 31831638 . S2CID 209343260 .
- ^ Делькруа Дж.Д., Валлетта Дж., Ву С. и др. (2004). «Торговля сигналом NGF: последствия для нормальных и дегенерирующих нейронов». NGF и родственные молекулы в здоровье и болезнях . Прогресс в исследованиях мозга. Том. 146. стр. 3–23. дои : 10.1016/s0079-6123(03)46001-9 . ISBN 9780444514721 . ПМИД 14699953 .
- ^ Жункейра, 12-е издание .
Внешние ссылки
[ редактировать ]- Гистологическое изображение: 3_09 в Центре медицинских наук Университета Оклахомы - «Слайд 3 Спинной мозг ».