Нейрофиламенты ( НФ ) классифицируются как промежуточные филаменты IV типа, в цитоплазме нейронов находящиеся . Это белковые полимеры диаметром 10 нм и длиной во многие микрометры. [1] Вместе с микротрубочками (~25 нм) и микрофиламентами (7 нм) они образуют цитоскелет нейрона . Считается, что они функционируют в первую очередь для обеспечения структурной поддержки аксонов и регулирования диаметра аксонов, что влияет на скорость нервной проводимости . Белки, образующие нейрофиламенты, являются членами семейства белков промежуточных филаментов, которое делится на шесть типов в зависимости от их генной организации и структуры белка. Типы I и II представляют собой кератины , экспрессирующиеся в эпителии. Тип III содержит белки виментин , десмин , периферин и глиальный фибриллярный кислый белок (GFAP). Тип IV состоит из белков нейрофиламентов NF-L, NF-M, NF-H и α-интернексина . Тип V состоит из ядерных ламинов , а тип VI — из белка нестина . Все гены промежуточной нити типа IV имеют два уникальных интрона, не обнаруженных в других последовательностях генов промежуточной нити, что позволяет предположить общее эволюционное происхождение от одного примитивного гена типа IV.
Белковый состав нейрофиламентов широко варьируется в зависимости от типа животных. Больше всего известно о нейрофиламентах млекопитающих. Исторически сложилось так, что первоначально считалось, что нейрофиламенты млекопитающих состоят всего из трех белков, называемых белком нейрофиламентов NF-L (низкомолекулярный; NF-L ), NF-M (средний молекулярный вес; NF-M ) и NF-H (высокомолекулярный). NF -H ). Эти белки были обнаружены в результате исследований аксонального транспорта , и их часто называют «триплетом нейрофиламентов». [3] Однако теперь ясно, что нейрофиламенты также содержат белок α-интернексин. [4] и что нейрофиламенты периферической нервной системы также могут содержать белок периферин. [5] (это отличается от периферина 2 , который экспрессируется в сетчатке ). Таким образом, нейрофиламенты млекопитающих представляют собой гетерополимеры, содержащие до пяти различных белков: NF-L, NF-M, NF-H, α-интернексина и периферина. Пять белков нейрофиламентов могут собираться в разных комбинациях в разных типах нервных клеток и на разных стадиях развития. Точный состав нейрофиламентов в любой конкретной нервной клетке зависит от относительных уровней экспрессии белков нейрофиламентов в клетке в данный момент. Например, экспрессия NF-H низкая в развивающихся нейронах и увеличивается постнатально в нейронах с миелинизированными аксонами. [6] Во взрослой нервной системе нейрофиламенты в мелких безмиелинизированных аксонах содержат больше периферина и меньше NF-H, тогда как нейрофиламенты в крупных миелинизированных аксонах содержат больше NF-H и меньше периферина. Субъединица промежуточных филаментов типа III, виментин , экспрессируется в развивающихся нейронах и нескольких очень необычных нейронах у взрослых в сочетании с белками типа IV, такими как нейроны сетчатки горизонтальные .
Белки субъединиц нейрофиламентов человека
Белок
Аминокислоты
NCBI Ref Seq
Прогнозируемая молекулярная масса
Кажущаяся молекулярная масса (SDS-PAGE)
Периферин
470
НП_006253.2
53,7 кДа
~56 кДа
α-Интернексин
499
НП_116116.1
55,4 кДа
~66 кДа
Белок нейрофиламентов L
543
НП_006149.2
61,5 кДа
~70 кДа
Белок нейрофиламентов М
916
НП_005373.2
102,5 кДа
~160 кДа
Белок нейрофиламентов H
1020
НП_066554.2
111,9 кДа
~200 кДа
Триплетные белки названы в зависимости от их относительного размера (низкий, средний, высокий). Кажущаяся молекулярная масса каждого белка, определенная с помощью SDS-PAGE, больше, чем масса, предсказанная по аминокислотной последовательности. Это связано с аномальной электрофоретической миграцией этих белков и особенно экстремально для белков нейрофиламентов NF-M и NF-H из-за их высокого содержания заряженных аминокислот и обширного фосфорилирования. Все три триплетных белка нейрофиламентов содержат длинные участки полипептидной последовательности, богатые остатками глутаминовой кислоты и лизина , а NF-M и особенно NF-H также содержат множественные тандемно повторяющиеся сайты фосфорилирования серина . Почти все эти сайты содержат пептид лизин-серин-пролин (KSP), а фосфорилирование обычно обнаруживается на аксональных, а не на дендритных нейрофиламентах. Человеческий NF-M имеет 13 таких сайтов KSP, тогда как человеческий NF-H экспрессируется двумя аллелями, один из которых производит 44, а другой 45 повторов KSP.
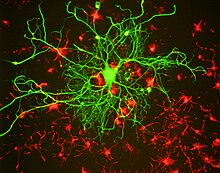
Клетки головного мозга крысы, выращенные в культуре тканей и окрашенные в зеленый цвет антителами к субъединице нейрофиламентов NF-L, что позволяет выявить крупный нейрон. Культуру окрашивали в красный цвет на альфа-интернексин, который в этой культуре обнаружен в нейрональных стволовых клетках, окружающих большой нейрон. Изображение предоставлено EnCor Biotechnology Inc. Фиксированный формалином и залитый парафином срез мозжечка человека , окрашенный антителами к свету нейрофиламентов, NF-L выявляется коричневым красителем, ядра клеток выявляются синим красителем. Богатая ядерными ядрами область слева — это зернистый слой, область справа — молекулярный слой. Антитело связывает отростки корзинчатых клеток, аксоны параллельных волокон, перикарию клеток Пуркинье и различные другие аксоны. Изображение предоставлено EnCor Biotechnology Inc.
Как и другие белки промежуточных филаментов, все белки нейрофиламентов имеют общую центральную альфа-спиральную область, известную как палочковидный домен из-за его палочковидной третичной структуры, окруженную амино-концевыми и карбокси-концевыми доменами, которые в значительной степени неструктурированы. Стержневые домены двух белков нейрофиламентов димеризуются с образованием альфа-спиральной спиральной спирали . Два димера соединяются в шахматном порядке антипараллельно, образуя тетрамер. Считается, что этот тетрамер является основной субъединицей (т.е. строительным блоком) нейрофиламента. Субъединицы тетрамера соединяются бок о бок, образуя нити единичной длины, которые затем отжигаются конец к концу, образуя зрелый полимер нейрофиламентов, но точная организация этих субъединиц внутри полимера неизвестна, в основном из-за гетерогенности белка. состав и неспособность кристаллизовать нейрофиламенты или белки нейрофиламентов. Структурные модели обычно предполагают наличие восьми тетрамеров (32 полипептида нейрофиламентов) в поперечном сечении нити, но измерения линейной массовой плотности предполагают, что это может варьироваться.
Аминоконцевые домены белков нейрофиламентов содержат многочисленные сайты фосфорилирования и, по-видимому, важны для взаимодействия субъединиц во время сборки филаментов. Карбокси-концевые домены представляют собой внутренне неупорядоченные домены, в которых отсутствует альфа-спираль или бета-лист. Различные размеры белков нейрофиламентов во многом обусловлены различиями в длине карбокси-концевых доменов. Эти домены богаты кислотными и основными аминокислотными остатками. Карбокси-концевые домены NF-M и NF-H являются самыми длинными и сильно модифицируются посттрансляционными модификациями, такими как фосфорилирование и гликозилирование in vivo. Они выступают радиально из основной нити, образуя плотную щеточную кайму из сильно заряженных и неструктурированных доменов, аналогичную щетине ершика для бутылок. Было предложено, чтобы эти энтропийно перемещающиеся домены определяли зону исключения вокруг каждой нити, эффективно отделяя нити от их соседей. Таким образом, выступы карбокси-концевых концов максимизируют свойства полимеров нейрофиламентов по заполнению пространства. При электронной микроскопии эти домены выглядят как выступы, называемые боковыми рукавами, которые, по-видимому, контактируют с соседними нитями.
Нейрофиламенты обнаруживаются в нейронах позвоночных в особенно высоких концентрациях в аксонах, где все они расположены параллельно вдоль длинной оси аксона, образуя непрерывно перекрывающийся массив. Было предложено, чтобы они функционировали как структуры, заполняющие пространство и увеличивающие диаметр аксонов. Их вклад в диаметр аксона определяется количеством нейрофиламентов в аксоне и плотностью их упаковки. Считается, что количество нейрофиламентов в аксоне определяется экспрессией генов нейрофиламентов. [7] и аксональный транспорт. Плотность упаковки нитей определяется их боковыми ответвлениями, определяющими расстояние между соседними нитями. Считается, что фосфорилирование боковых ответвлений увеличивает их растяжимость, увеличивая расстояние между соседними нитями. [8] путем связывания двухвалентных катионов между боковыми ответвлениями соседних нитей [9] [10]
На ранних стадиях развития аксоны представляют собой узкие отростки, содержащие относительно мало нейрофиламентов. Те аксоны, которые становятся миелинизированными, накапливают больше нейрофиламентов, что приводит к увеличению их калибра. После того, как аксон вырос и соединился с клеткой-мишенью , диаметр аксона может увеличиться в пять раз. [11] Это вызвано увеличением количества нейрофиламентов, экспортируемых из тела нервной клетки, а также замедлением скорости их транспорта. В зрелых миелинизированных аксонах нейрофиламенты могут быть единственной наиболее распространенной цитоплазматической структурой и могут занимать большую часть площади поперечного сечения аксона. Например, большой миелинизированный аксон может содержать тысячи нейрофиламентов в одном поперечном сечении.
Помимо своей структурной роли в аксонах, нейрофиламенты также являются грузом аксонального транспорта . [3] Большинство белков нейрофиламентов в аксонах синтезируются в теле нервных клеток, где они быстро собираются в полимеры нейрофиламентов в течение примерно 30 минут. [12] Эти собранные полимеры нейрофиламентов транспортируются по аксону по микротрубочкам , питаемым моторными белками микротрубочек . [13] Нити движутся в двух направлениях, то есть как к кончику аксона (антеградно), так и к телу клетки (ретроградно), но общее направление является антероградным. Нити движутся со скоростью до 8 мкм/с в коротких временных масштабах (секунды или минуты) со средней скоростью примерно 1 мкм/с. [14] Однако средняя скорость в более длительных временных масштабах (часах или днях) низкая, поскольку движения очень редки и состоят из коротких спринтов, прерываемых длительными паузами. [15] [16] Таким образом, в долговременных масштабах нейрофиламенты движутся в медленном компоненте аксонального транспорта.
множество специфических антител Разработано и коммерчески доступно к белкам нейрофиламентов. Эти антитела можно использовать для обнаружения белков нейрофиламентов в клетках и тканях с помощью иммунофлуоресцентной микроскопии или иммуногистохимии . Такие антитела широко используются для идентификации нейронов и их отростков в гистологических срезах и культурах тканей . Белок промежуточных филаментов VI типа Нестин экспрессируется в развивающихся нейронах и глии. Нестин считается маркером нейрональных стволовых клеток, и наличие этого белка широко используется для определения нейрогенеза . Этот белок теряется по мере развития.
Антитела к нейрофиламентам также широко используются в диагностической невропатологии . Окрашивание этими антителами позволяет отличить нейроны (положительные на белки нейрофиламентов) от глии (отрицательные на белки нейрофиламентов).
Существует также значительный клинический интерес к использованию белков нейрофиламентов в качестве биомаркеров аксонального повреждения при заболеваниях, поражающих центральную нервную систему. [17] [18] Когда нейроны или аксоны дегенерируют, белки нейрофиламентов высвобождаются в кровь или спинномозговую жидкость. Таким образом, иммуноанализ белков нейрофиламентов в спинномозговой жидкости и плазме может служить индикатором аксонального повреждения при неврологических расстройствах. [19] Таким образом, уровни NF-L в крови и спинномозговой жидкости являются полезными маркерами для мониторинга заболевания при боковом амиотрофическом склерозе . [20] рассеянный склероз , [21] и в последнее время болезнь Хантингтона . [22] Его также оценивали как прогностический маркер функционального исхода после острого ишемического инсульта. [23] Мутантные мыши с аномалиями нейрофиламентов имеют фенотипы, напоминающие боковой амиотрофический склероз . [24] Недавняя работа, выполненная в сотрудничестве между EnCor Biotechnology Inc. и Университетом Флориды, показала, что антитела к NF-L, используемые в наиболее широко используемых анализах NF-L, специфичны к расщепленным формам NF-L, образующимся в результате протеолиза, индуцированного гибелью клеток. . [25]
^ Йонссон М., Зеттерберг Х., ван Страатен Э., Линд К., Сиверсен С., Эдман А. и др. (март 2010 г.). «Биомаркеры спинномозговой жидкости поражений белого вещества - поперечные результаты исследования LADIS». Европейский журнал неврологии . 17 (3): 377–82. дои : 10.1111/j.1468-1331.2009.02808.x . ПМИД 19845747 . S2CID 31052853 .
^ Розенгрен Л.Е., Карлссон Дж.Э., Карлссон Дж.О., Перссон Л.И., Виккельсё К. (ноябрь 1996 г.). «У пациентов с боковым амиотрофическим склерозом и другими нейродегенеративными заболеваниями наблюдается повышенный уровень белка нейрофиламентов в спинномозговой жидкости». Журнал нейрохимии . 67 (5): 2013–8. дои : 10.1046/j.1471-4159.1996.67052013.x . ПМИД 8863508 . S2CID 36897027 .
^ Теуниссен С.Э., Якобеус Э., Хадеми М., Брундин Л., Норгрен Н., Коэль-Зиммелинк М.Дж. и др. (апрель 2009 г.). «Комбинация N-ацетиласпартата спинномозговой жидкости и нейрофиламентов при рассеянном склерозе». Неврология . 72 (15): 1322–9. дои : 10.1212/wnl.0b013e3181a0fe3f . ПМИД 19365053 . S2CID 22681349 . ,
↑ Шоу Дж., Мадорски И., Инь Ю, Ван Ю, Рана С., Йоргенсен М., Фуллер Д.Д. (апрель 2023 г.). «Световые антитела к нейрофиламентам уманского типа являются эффективными реагентами для визуализации нейродегенерации». мозговые коммуникации 10.1093/braincomms/fcad067
Arc.Ask3.Ru Номер скриншота №: 71bcff0d92247637c684ecd81cf57d27__1693468920 URL1:https://arc.ask3.ru/arc/aa/71/27/71bcff0d92247637c684ecd81cf57d27.html Заголовок, (Title) документа по адресу, URL1: Neurofilament - Wikipedia
Данный printscreen веб страницы (снимок веб страницы, скриншот веб страницы), визуально-программная копия документа расположенного по адресу URL1 и сохраненная в файл, имеет: квалифицированную, усовершенствованную (подтверждены: метки времени, валидность сертификата), открепленную ЭЦП (приложена к данному файлу), что может быть использовано для подтверждения содержания и факта существования документа в этот момент времени. Права на данный скриншот принадлежат администрации Ask3.ru, использование в качестве доказательства только с письменного разрешения правообладателя скриншота. Администрация Ask3.ru не несет ответственности за информацию размещенную на данном скриншоте. Права на прочие зарегистрированные элементы любого права, изображенные на снимках принадлежат их владельцам. Качество перевода предоставляется как есть. Любые претензии, иски не могут быть предъявлены. Если вы не согласны с любым пунктом перечисленным выше, вы не можете использовать данный сайт и информация размещенную на нем (сайте/странице), немедленно покиньте данный сайт. В случае нарушения любого пункта перечисленного выше, штраф 55! (Пятьдесят пять факториал, Денежную единицу (имеющую самостоятельную стоимость) можете выбрать самостоятельно, выплаичвается товарами в течение 7 дней с момента нарушения.)