Арбускулярная микориза




Арбускулярная микориза ( АМ ) (множественная микориза ) — разновидность микоризы , при которой -симбионт гриб ( АМ-грибы , или АМФ) проникает в корковые клетки корней , сосудистого растения образуя арбускулы . Арбускулярная микориза — разновидность эндомикоризы наряду с эрикоидной микоризой и микоризой орхидей (не путать с эктомикоризой ). Для них характерно образование уникальных древовидных структур — арбускул. [ 1 ] Кроме того, часто встречаются шаровидные структуры хранения, называемые везикулами.
Арбускулярные микоризы образуются грибами подтипа Glomeromycotina . Этот подтип, наряду с Mortierellomycotina и Mucoromycotina, образует тип Mucoromycota , сестринскую кладу более известных и разнообразных дикаровых грибов. [ 2 ]
AM-грибы помогают растениям захватывать питательные вещества, такие как фосфор , сера , азот и микроэлементы из почвы . Считается, что развитие арбускулярного микоризного симбиоза сыграло решающую роль в первоначальном заселении суши растениями и в эволюции сосудистых растений. [ 3 ] Было сказано, что быстрее перечислить растения, которые не образуют эндомикоризу, чем те, которые образуют. [ 4 ] Этот симбиоз представляет собой высокоразвитые мутуалистические отношения, обнаруженные между грибами и растениями, наиболее распространенный из известных растительных симбиозов. [ 5 ] а АМФ обнаружен в 80% сосудистых растений . существующих сегодня семейств [ 6 ]
Раньше этот тип микоризных ассоциаций назывался «Везикулярная арбускулярная микориза (ВАМ)», но поскольку некоторые представители этих грибов не образуют пузырьков, например, представители Gigasporaceae ; для их включения термин был изменен на «Арбускулярные микоризы». [ 7 ] [ 8 ]
Достижения в исследованиях физиологии и экологии микоризы с 1970-х годов привели к лучшему пониманию многочисленных ролей АМФ в экосистеме. Примером может служить важный вклад клееподобного белка гломалина в структуру почвы (см. ниже). Эти знания применимы к человеческой деятельности по управлению экосистемами , восстановлению экосистем и сельскому хозяйству .
Эволюция микоризного симбиоза
[ редактировать ]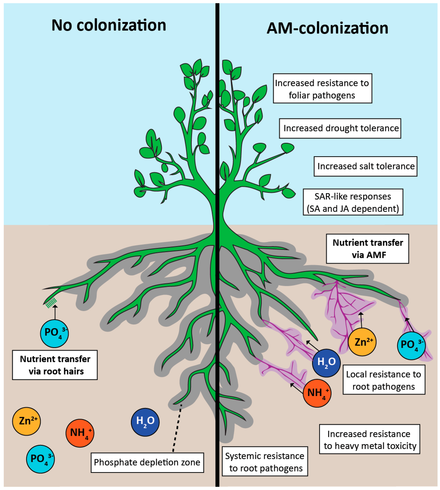
Палеобиология
[ редактировать ]Как палеобиологические , так и молекулярные данные указывают на то, что АМ — это древний симбиоз, возникший не менее 460 миллионов лет назад. Симбиоз АМ широко распространен среди наземных растений, что позволяет предположить, что микоризы присутствовали у ранних предков современных наземных растений. Эта положительная связь с растениями могла способствовать развитию наземных растений. [ 5 ]
В кремне Рини нижнего девона обнаружены окаменелости самых ранних наземных растений, в которых наблюдались AM-грибы. [ 10 ] Окаменелые растения, содержащие микоризные грибы, сохранились в кремнеземе.
В раннем девоне произошло развитие наземной флоры. В растениях райнского кремня нижнего девона (400 млн лет назад) обнаружены структуры, напоминающие пузырьки и споры современных видов Glomus . Колонизированные ископаемые корни наблюдались у Aglaophyton major и Rhynia , древних растений, обладающих характеристиками сосудистых растений и мохообразных с примитивными протостелическими корневищами . [ 10 ]
Внутрикорневой мицелий наблюдался во внутриклеточных пространствах корней, а арбускулы наблюдались в слоях тонкостенных клеток, подобных палисадной паренхиме . Ископаемые арбускулы очень похожи на арбускулы существующих АМФ. [ 10 ] Клетки, содержащие арбускулы, имеют утолщенные стенки, что наблюдается и в современных колонизированных клетках.
Микоризы миоцена имеют везикулярную морфологию, очень напоминающую морфологию современных Glomerales . Эта консервативная морфология может отражать доступность питательных веществ, обеспечиваемых растениями-хозяевами как в современных, так и в миоценовых мутуализмах. [ 11 ] Однако можно утверждать, что эффективность сигнальных процессов, вероятно, изменилась со времен миоцена, и это невозможно обнаружить в летописи окаменелостей. Тонкая настройка сигнальных процессов позволит улучшить координацию и обмен питательными веществами между симбионтами, одновременно повышая приспособленность как грибов, так и растительных симбионтов.
Характер взаимоотношений растений и предков арбускулярных микоризных грибов является спорным. Две гипотезы:
- Микоризный симбиоз развился из паразитарного взаимодействия, которое переросло во взаимовыгодные отношения.
- Микоризы развились из сапробных грибов, ставших эндосимбиотами. [ 10 ]
В Райни Черт были обнаружены как сапротрофы, так и биотрофы, но доказательств в поддержку какой-либо гипотезы мало.
Существуют некоторые ископаемые свидетельства, свидетельствующие о том, что паразитические грибы не убивали клетки-хозяева сразу после инвазии, хотя в клетках-хозяевах наблюдалась реакция на инвазию. Этот ответ, возможно, развился в химические сигнальные процессы, необходимые для симбиоза. [ 10 ]
Считается, что в обоих случаях симбиотическое взаимодействие растений и грибов превратилось из отношений, в которых грибы брали питательные вещества от растения, в симбиотические отношения, в которых растение и грибы обмениваются питательными веществами.
Древние растения не имели настоящих корней. Струллу-Дерриен и Струллу предложили термин «парамикориза» для микоризы, поражающей корневище, побег или слоевища, и термин «эвмикориза», поражающий настоящие корни. [ 12 ] [ 13 ] [ 14 ] Эти структуры были обнаружены как в спорофитах, так и в гаметофитах ранних наземных растений. [ 13 ]
Молекулярные доказательства
[ редактировать ]Возросший интерес к микоризному симбиозу и разработка сложных молекулярных методов привели к быстрому развитию генетических данных. Ван и др. (2010) исследовали гены растений, участвующие в коммуникации с грибами-партнерами порядка Glomales (DMI1, DMI3, IPD3). [ 15 ] [ 16 ] Эти три гена можно было секвенировать из всех основных клад современных наземных растений, включая печеночников , самую базальную группу, и оказалось, что филогения трех генов согласуется с нынешними филогениями наземных растений. Это означает, что микоризные гены должны были присутствовать у общего предка наземных растений и что они должны были быть вертикально унаследованы с тех пор, как растения колонизировали землю. [ 15 ]
Симбиоз АМ-грибов и цианобактерий
[ редактировать ]Выявлено, что АМ-грибы имеют коровый фермент бактериального типа (рибонуклеаза III) механизма процессинга мРНК, вероятно, в результате процесса горизонтального переноса генов от цианобактериального предка и, возможно, связанного с симбиозом. [ 17 ] Эта находка генетической окаменелости внутри AM-грибов повышает вероятность тесной связи между AM-грибами и предками цианобактерий. о подобном симбиозе геосифона и ностока . Ранее сообщалось [ 18 ]
Эволюция циркадных часов у AM-грибов
[ редактировать ]Удивительно, но, несмотря на свою длительную эволюцию в качестве подземного партнера корней растений, чья среда далека от света или колебаний температуры, у AMF все еще есть сохранившиеся циркадные часы, чей грибковый циркадный осциллятор ( frq ) активируется синим светом, подобно модельному циркадному генератору. гриб Neurospora crassa . [ 19 ] Доказанное сохранение циркадных часов и выходных генов у R. иррегулярного открывает двери для изучения циркадных часов у грибкового партнера симбиоза AM. В том же исследовании охарактеризован ген AMF frq , [ 19 ] Это первый frq, ген идентифицированный из внешней группы Дикарии, и он предполагает, что эволюция гена frq в грибном царстве намного старше, чем считалось ранее.
Физиология
[ редактировать ]Пресимбиоз
[ редактировать ]Развитие АМ-грибов до колонизации корней, известное как пресимбиоз, состоит из трех стадий: прорастание спор, рост гиф, распознавание хозяина и образование апрессория .
Прорастание спор
[ редактировать ]Споры АМ-грибов представляют собой толстостенные многоядерные покоящиеся структуры. [ 21 ] Прорастание спор не зависит от растения, так как споры прорастали в экспериментальных условиях в отсутствие растений как in vitro , так и в почве. Однако скорость прорастания может быть увеличена за счет выделений корня хозяина . [ 22 ] Споры грибов AM прорастают при подходящих условиях почвенной матрицы, температуры, концентрации углекислого газа, pH и концентрации фосфора. [ 21 ]
Рост гиф
[ редактировать ]Рост гифов AM через почву контролируется экссудатами корней хозяина, известными как стриголактоны , и концентрацией фосфора в почве. [ 23 ] Низкие концентрации фосфора в почве увеличивают рост и ветвление гиф, а также вызывают выделение растениями соединений, которые контролируют интенсивность ветвления гиф. [ 22 ] [ 24 ]
Ветвление гиф грибов АМ, выращенных в среде с концентрацией фосфора 1 мМ, значительно снижается, но длина зародышевой трубки и общий рост гиф не изменяются. Концентрация фосфора 10 мМ ингибировала как рост, так и ветвление гиф. Такая концентрация фосфора наблюдается в естественных почвенных условиях и, таким образом, может способствовать снижению микоризной колонизации. [ 24 ]
Распознавание хоста
[ редактировать ]Показано, что корневые экссудаты растений-хозяев AMF, выращенных в жидкой среде с фосфором и без него, влияют на рост гиф. Споры Gigaspora margarita выращивали в экссудатах растений-хозяев. Гифы грибов, выращенные в экссудате корней, лишенных фосфора, росли больше и давали третичные ветви по сравнению с таковыми, выращенными в экссудате растений, получавших достаточное количество фосфора. При добавлении корневых экссудатов, способствующих росту, в низкой концентрации грибы AM образовывали разбросанные длинные ветви. По мере увеличения концентрации экссудата грибы образовывали более плотно сгруппированные ветви. В арбускулах с наибольшей концентрацией формировались структуры фосфорного обмена АМФ. [ 24 ]
Считается, что эта хемотаксическая реакция грибов на экссудаты растений-хозяев повышает эффективность колонизации корней хозяина в почвах с низким содержанием фосфора. [ 22 ] Это приспособление грибов к эффективному исследованию почвы в поисках подходящего растения-хозяина. [ 24 ]
Еще одно доказательство того, что арбускулярные микоризные грибы проявляют специфичный для хозяина хемотаксис , который обеспечивает рост гиф по направлению к корням потенциального растения-хозяина: споры Glomus mosseae были отделены от корней растения-хозяина, растений-нехозяев и мертвого растения-хозяина с помощью проницаемой мембраны. только для гиф. При обработке растением-хозяином грибы пересекали мембрану и всегда выходили в пределах 800 мкм от корня, но не при обработке растениями-нехозяевами и мертвыми растениями. [ 25 ]
Молекулярные методы использовались для понимания сигнальных путей между арбускулярными микоризами и корнями растений. В 2003 г. было показано, как АМ претерпевает физиологические изменения в присутствии экссудатов потенциальных корней растения-хозяина, колонизируя его. Экссудаты корней растений-хозяев запускают и активируют гены грибов AM, необходимые для дыхания соединений углерода в спорах. В экспериментах скорость транскрипции 10 генов увеличивалась через полчаса после воздействия и еще быстрее через 1 час. после 4-часового воздействия АМ отвечают морфологическим ростом. Гены, выделенные в то время, участвуют в активности митохондрий и выработке ферментов. Скорость дыхания грибов, измеренная по скорости потребления O 2 , увеличивалась на 30% через 3 часа после воздействия корневых экссудатов, что указывает на то, что корневые экссудаты растения-хозяина стимулируют митохондриальную активность спор AMF. Это может быть частью грибкового регуляторного механизма, который сохраняет энергию спор для эффективного роста и разветвления гиф при получении сигналов от потенциального растения-хозяина. [ 26 ]
Благодарный
[ редактировать ]Когда арбускулярные микоризные грибные гифы встречаются с корнем растения-хозяина, аппрессорий на корневом эпидермисе формируется хозяина или «инфекционная структура». Благодаря этой структуре гифы могут проникать в кору паренхимы . [ 27 ] АМ не нуждается в химических сигналах растения для формирования аппрессорий. АМ-грибы могли образовывать аппрессории на клеточных стенках «призрачных» клеток, у которых был удален протопласт , чтобы исключить передачу сигналов между грибами и растением-хозяином. Однако гифы не проникали дальше в клетки и не росли в направлении коры корня, что указывает на то, что передача сигналов между симбионтами необходима для дальнейшего роста после образования аппрессорий. [ 22 ]
Симбиоз
[ редактировать ]Попав внутрь паренхимы, гриб образует сильно разветвленные структуры для обмена питательными веществами с растением, называемые арбускулами . [ 27 ] Это отличительные структуры арбускулярного микоризного гриба. Арбускулы являются местом обмена фосфора, углерода, воды и других питательных веществ. [ 21 ] Выделяют две формы: парижский тип характеризуется ростом гиф от одной клетки к другой; а Arum тип характеризуется ростом гиф в пространстве между растительными клетками. [ 28 ] Выбор между типом Paris и типом Arum в первую очередь определяется семейством растений-хозяев, хотя некоторые семейства или виды способны относиться к любому типу. [ 29 ] [ 30 ]
Растение-хозяин контролирует межклеточную пролиферацию гиф и образование арбускул. растения Происходит деконденсация хроматина растения , что свидетельствует об усилении транскрипции ДНК в клетках, содержащих арбускулы. [ 27 ] Для размещения арбускул в растительной клетке-хозяине требуются серьезные модификации. Вакуоли размножаются сжимаются, а другие клеточные органеллы . растительной клетки Цитоскелет реорганизуется вокруг арбускул.
Есть два других типа гиф, которые происходят из колонизированного корня растения-хозяина. После того как произошла колонизация, недолговечные гифы побегов вырастают из корня растения в почву. Это гифы, которые поглощают фосфор и микроэлементы, поставляемые растению. Гифы грибов AM имеют высокое соотношение поверхности к объему, что делает их поглотительную способность большей, чем у корней растений. [ 31 ] Гифы АМФ также тоньше корней и могут проникать в недоступные для корней поры почвы. [ 32 ] Четвертый тип гиф АМФ растет из корней и колонизирует корни других растений-хозяев. Четыре типа гиф морфологически различны. [ 21 ]
Поглощение и обмен питательных веществ
[ редактировать ]АМ-грибы — облигатные симбионты . Они обладают ограниченной сапробной способностью и зависят от растений в плане углеродного питания. [ 33 ] АМ-грибы поглощают продукты фотосинтеза растения-хозяина в виде гексоз .
Перенос углерода от растения к грибам может происходить через арбускулы или внутрикорневые гифы. [ 34 ] Вторичный синтез из гексоз под действием АМ происходит во внутрикорневом мицелии . Внутри мицелия гексоза превращается в трегалозу и гликоген . Трегалоза и гликоген представляют собой формы хранения углерода, которые могут быстро синтезироваться и разлагаться и могут буферизировать внутриклеточные концентрации сахара. [ 34 ] Внутрирадикальная гексоза вступает в окислительный пентозофосфатный путь , который производит пентозу для нуклеиновых кислот.
Биосинтез липидов также происходит во внутрирадикальном мицелии. Затем липиды сохраняются или экспортируются в экстрарадикальные гифы, где они могут храниться или метаболизироваться. Распад липидов на гексозы, известный как глюконеогенез , происходит во внерадикальном мицелии. [ 34 ] Примерно 25% углерода, перенесенного из растения в грибы, хранится во внерадикальных гифах. [ 35 ] До 20% углерода растения-хозяина может быть передано АМ-грибам. [ 34 ] Это представляет собой значительные инвестиции углерода растениями-хозяевами в микоризную сеть и вклад в подземный пул органического углерода.
Увеличение поступления углерода от растения к AM-грибам увеличивает поглощение и перенос фосфора от грибов к растению. [ 36 ] Аналогичным образом, поглощение и перенос фосфора снижается, когда уменьшается количество фотосинтата, поставляемого грибам. Виды АМФ различаются по способности снабжать растение фосфором. [ 37 ] В некоторых случаях арбускулярные микоризы являются плохими симбионтами, обеспечивающими мало фосфора и поглощающими относительно большое количество углерода. [ 37 ]
Основная польза микоризы для растений объясняется увеличением поглощения питательных веществ, особенно фосфора. Это может быть связано с увеличением площади поверхности, контактирующей с почвой, увеличением перемещения питательных веществ в микоризу, изменением корневой среды и увеличением запасов питательных веществ. [ 32 ] Микориза может гораздо эффективнее поглощать фосфор, чем корни растений. Фосфор перемещается к корню или посредством диффузии, а гифы сокращают расстояние, необходимое для диффузии, тем самым увеличивая поглощение. Скорость поступления фосфора в микоризу может в шесть раз превышать скорость поступления в корневые волоски. [ 32 ] В некоторых случаях роль поглощения фосфора может полностью взять на себя микоризная сеть, и весь фосфор растения может иметь гифальное происхождение. [ 37 ] Меньше известно о роли азотного питания в арбускулярной микоризной системе и его влиянии на симбиоз и сообщество. Хотя в выяснении механизмов этого сложного взаимодействия были достигнуты значительные успехи, еще предстоит провести много исследований.
Микоризная активность увеличивает концентрацию фосфора, доступного в ризосфере . корневой зоны Микоризы снижают pH за счет избирательного поглощения NH 4. + ( -ионы аммония ) и высвобождая H + ионы. Снижение pH почвы увеличивает растворимость фосфорных осадков. Гифальный NH 4 + поглощение также увеличивает приток азота к растению, поскольку внутренние поверхности почвы поглощают аммоний и распределяют его путем диффузии. [ 35 ]
Мейоз и рекомбинация
[ редактировать ]AM-грибы считались бесполыми, поскольку у них отсутствуют заметные половые структуры. [ 38 ] Однако было обнаружено, что гомологи 51 гена, которые функционируют в мейозе , включая семь генов, специфичных для мейоза, консервативны в геномах нескольких видов AMF, что позволяет предположить, что эти предположительно древние бесполые грибы могут быть способны подвергаться обычному мейозу. [ 39 ] Кроме того, у Rhizophagus ignores генетический обмен, включающий реципрокную рекомбинацию, был обнаружен в дикарионах между гаплоидными геномами. [ 38 ]
Механизм колонизации
[ редактировать ]Недавние исследования показали, что AM-грибы выделяют диффузионный фактор, известный как фактор myc , который активирует индуцируемый ген фактора клубенька MtEnod11. Это тот же ген, который участвует в установлении симбиоза с ризобиальными азотфиксирующими бактериями (Косута и др., 2003). Этот фактор был впервые выявлен Фабьеной Майе и ее коллегами. [ 40 ] в новаторской работе, опубликованной в журнале Nature, они извлекли триста литров микоризованных корней моркови и экссудата из 40 миллионов прорастающих спор Rhizophagus нерегулярной и очистили активную фракцию. Они продемонстрировали, что это активное начало имеет липохитоолигосахаридную природу.
Распознавание факторов Myc запускает общий симбиотический сигнальный путь (CSSP), который в конечном итоге приводит к программе аккомодации растения, чтобы стать заложником арбускулярных микориз.

Общий симбиотический сигнальный путь (CSSP) представляет собой сигнальный каскад в растениях, который, как считается, активируется как при NOD-фактора (для клубенькообразующих восприятии ризобий ), так и при восприятии MYC-фактора , выделяемого арбускулярными микоризными грибами. Этот путь отличается от путей распознавания патогенов , но может иметь некоторые общие рецепторы, участвующие как в распознавании патогенов, так и в CSSP. Недавняя работа [ 41 ] Кевин Коуп и его коллеги показали, что, возможно, другие типы микоризы могут включать компоненты CSSP, такие как распознавание Myc-фактора.
Колонизация AMF требует следующей цепочки [ 42 ] событий, которые можно условно разделить на следующие этапы:
- 1. Предварительная контактная сигнализация,
- 2. CSSP
- 2.А. Восприятие
- 2.Б. Передача инфекции
- 2.С. Транскрипция
- 3. Программа проживания
Экология
[ редактировать ]Биогеография
[ редактировать ]Арбускулярные микоризные грибы чаще всего встречаются у растений, растущих на минеральных почвах, и имеют чрезвычайно важное значение для растений, растущих на субстратах с дефицитом питательных веществ, таких как вулканическая почва и среда песчаных дюн . Популяции AM-грибов наиболее велики в растительных сообществах с высоким разнообразием, таких как тропические леса и луга умеренного пояса, где они имеют много потенциальных растений-хозяев и могут воспользоваться своей способностью колонизировать широкий круг хозяев. [ 43 ] В очень засушливых или богатых питательными веществами почвах частота микоризной колонизации ниже. Микориза наблюдалась в водной среде обитания ; однако было показано, что заболоченные почвы уменьшают колонизацию некоторых видов. [ 43 ] Арбускулярные микоризные грибы встречаются у 80% видов растений. [ 44 ] и исследованы на всех континентах, кроме Антарктиды. [ 45 ] [ 46 ] На биогеографию glomeromycota влияют ограничения расселения, [ 47 ] факторы окружающей среды, такие как климат, [ 45 ] ряд почвы и pH почвы, [ 46 ] питательные вещества почвы [ 48 ] и растительное сообщество. [ 45 ] [ 49 ] Хотя данные 2000 года свидетельствуют о том, что AM-грибы не являются специалистами по своим видам-хозяевам, [ 50 ] исследования 2002 года показали, что по крайней мере некоторые таксоны грибов являются специалистами-хозяевами. [ 51 ] Экология мукоромикотиновых грибов, образующих арбускулярные микоризы с «тонкокорневыми эндофитами», в значительной степени неизвестна.
Реакция на растительные сообщества
[ редактировать ]Специфика, круг хозяев и степень колонизации микоризных грибов трудно анализировать в полевых условиях из-за сложности взаимодействий между грибами внутри корня и внутри системы. Нет четких доказательств того, что арбускулярные микоризные грибы проявляют специфичность к колонизации потенциальных видов растений-хозяев АМ, как и грибковые патогены для их растений-хозяев. [ 43 ] Это может быть связано с противоположным селективным давлением.
В патогенных отношениях растение-хозяин получает выгоду от мутаций, которые предотвращают колонизацию, тогда как при мутуалистических симбиотических отношениях растение получает выгоду от мутаций, которые позволяют колонизировать AMF. [ 43 ] Однако виды растений различаются по степени и зависимости от колонизации некоторыми АМ-грибами, причем некоторые растения могут быть факультативными микотрофами , а другие - облигатными микотрофами. [ 43 ] В последнее время статус микоризы связывают с распространением растений. [ 52 ] при этом облигатно-микоризные растения занимают более теплые и засушливые места обитания, тогда как факультативные микоризные растения занимают более обширные места обитания.
Способность одних и тех же грибов АМ колонизировать многие виды растений имеет экологические последствия. Растения разных видов могут быть связаны под землей общей мицелиальной сетью. [ 43 ] Одно растение может обеспечивать фотосинтетический углерод для создания мицелиальной сети, которую другое растение другого вида может использовать для поглощения минералов. Это означает, что арбускулярные микоризы способны балансировать подземные внутри- и межвидовые взаимодействия растений. [ 43 ]
Поскольку грибы Glomeromycota живут внутри корней растений, они могут подвергаться существенному влиянию своего растения-хозяина и, в свою очередь, также влиять на растительные сообщества. Растения могут отдавать до 30% углерода фотосинтеза AM-грибам. [ 53 ] а взамен АМ-грибы могут приобретать до 80% растительного фосфора и азота. [ 44 ] Разнообразие грибковых сообществ AM положительно связано с разнообразием растений. [ 54 ] продуктивность растений [ 55 ] и травоядные. [ 56 ] На арбускулярные микоризные грибы можно влиять в результате мелкомасштабного взаимодействия с местным растительным сообществом. Например, соседство растений вокруг очагового растения может изменить грибковые сообщества AM. [ 57 ] как и порядок размещения растений на участках. [ 58 ]
AM-грибы и инвазия растений
[ редактировать ]В ходе инвазий видов растений грибное сообщество и биомасса АМ могут радикально изменяться. В большинстве случаев биомасса и разнообразие АМ-грибов уменьшаются по мере инвазий. [ 59 ] [ 60 ] [ 61 ] Однако некоторые виды микотрофных растений могут фактически увеличивать разнообразие грибов AM во время инвазии. [ 62 ]
Микоризный статус инвазивных видов растений часто варьируется в зависимости от региона. Например, в Великобритании и Центральной Европе в последнее время инвазивные растения чаще становятся облигатно микоризными, чем ожидалось. [ 52 ] [ 63 ] в то время как инвазивные растения в Калифорнии оказались микоризными реже, чем ожидалось. [ 64 ]
Взаимодействие АМ-грибов с другими растительными симбионтами
[ редактировать ]Все симбионты внутри растения-хозяина взаимодействуют, часто непредсказуемым образом. Метаанализ 2010 года показал, что растения, колонизированные как AM-грибами, так и вертикально передающимися эндофитами, часто крупнее, чем растения, независимо колонизированные этими симбионтами. [ 65 ] Однако эта взаимосвязь зависит от контекста, поскольку AM-грибы могут синергически взаимодействовать с грибковыми эндофитами, населяющими листья растения-хозяина. [ 66 ] [ 67 ] или антагонистически. [ 68 ] [ 69 ] [ 70 ] Аналогичный диапазон взаимодействий может происходить между AM-грибами, эктомикоризными грибами и эндофитами с темными перегородками . [ 71 ]
Реакция на градиенты окружающей среды
[ редактировать ]Арбускулярные микоризные грибы различаются в зависимости от многих градиентов окружающей среды . Известно, что их толерантность к замораживанию и высыханию варьируется в зависимости от таксона AM-грибов. [ 72 ] Грибы AM становятся менее распространенными и разнообразными при более высоких концентрациях питательных веществ и влаги в почве. [ 73 ] предположительно потому, что оба растения выделяют меньше углерода АМ-грибам, а АМ-грибы перераспределяют свои ресурсы внутрирадикальным гифам в этих условиях окружающей среды. [ 74 ] В долгосрочной перспективе эти условия окружающей среды могут даже создать локальную адаптацию между растениями-хозяевами, AM-грибами и местными концентрациями питательных веществ в почве. [ 75 ] Состав АМ часто становится менее разнообразным на вершинах гор, чем на более низких высотах, что обусловлено составом видов растений. [ 76 ]
Было показано, что AM-грибы улучшают толерантность растений к абиотическим факторам окружающей среды, таким как засоленность . Они облегчают солевой стресс и способствуют росту и продуктивности растений. [ 77 ]
Экология ризосферы
[ редактировать ]Ризосфера – это почвенная зона в непосредственной близости от корневой системы.
Арбускулярный микоризный симбиоз влияет на сообщество и разнообразие других организмов в почве. Это можно увидеть непосредственно по выделению экссудатов или косвенно по изменению вида растений, типа и количества растительных экссудатов. [ 78 ]
Было показано, что разнообразие микориз увеличивает разнообразие видов растений по мере увеличения потенциального числа ассоциаций. Доминирующие арбускулярные микоризные грибы могут предотвратить инвазию немикоризных растений на землю, где они установили симбиоз, и способствовать развитию своего микоризного хозяина. [ 79 ]
Когда в почве присутствуют бактерии-ризобии, микоризная колонизация увеличивается из-за увеличения концентрации химических сигналов, участвующих в установлении симбиоза (Xie et al., 2003). Молекулы, подобные факторам Nod, были выделены из AM-грибов и, как было показано, индуцируют MtEnod11, образование боковых корней и усиливают микоризацию. [ 80 ] Эффективная микоризная колонизация также может увеличить образование клубеньков и симбиотическую фиксацию азота у микоризных бобовых. [ 35 ]
Степень колонизации арбускулярной микоризы и виды влияют на популяцию бактерий в ризосфере. [ 81 ] Виды бактерий различаются по своей способности конкурировать за корневые экссудаты углеродных соединений. Изменение количества или состава корневых и грибных экссудатов вследствие существующей микоризной колонизации АМ определяет разнообразие и численность бактериального сообщества в ризосфере. [ 78 ]
Влияние АМ-грибов на рост корней и побегов растений может оказывать и косвенное влияние на ризосферные бактерии. АМФ вносит значительное количество углерода в ризосферу за счет роста и дегенерации гифальной сети. Имеются также данные, позволяющие предположить, что АМ-грибы могут играть важную роль в обеспечении специфического воздействия видов растений на бактериальный состав ризосферы. [ 78 ]
Glomeromycota и глобальное изменение климата
[ редактировать ]Глобальное изменение климата влияет на грибковые сообщества АМ и взаимодействие между грибами АМ и их растениями-хозяевами. Хотя общепринято, что взаимодействие между организмами повлияет на их реакцию на глобальное изменение климата, нам все еще не хватает возможности предсказать результат этих взаимодействий в будущих климатических условиях. [ 82 ] В недавних метаанализах было обнаружено, что AM-грибы увеличивают биомассу растений в условиях засухи и уменьшают биомассу растений в ходе исследований по моделированию отложения азота. [ 83 ] [ 84 ] Было показано, что арбускулярные микоризные грибы сами по себе увеличивают свою биомассу в ответ на повышенное содержание CO 2 в атмосфере . [ 85 ]
Растения без арбускулярной микоризы
[ редактировать ]Представители семейства горчичных ( Brassicaceae ), такие как капуста, цветная капуста, рапс и крамбе , не образуют на своих корнях арбускулярные микоризные грибы. [ 86 ]
Молекулярно-генетический анализ арбускулярных микоризных грибов
[ редактировать ]За последние десять лет произошел впечатляющий прогресс в области молекулярно-генетических технологий и инструментов. Эти достижения позволяют микробным и микоризным экологам задавать новые и интересные вопросы об экологической и эволюционной роли арбускулярных микоризных (АМ) грибов как отдельных особей, в сообществах и экосистемах. Генетический анализ AM-грибов использовался для изучения генетической структуры отдельных спор с использованием мультилокусного генотипирования. [ 87 ] Разнообразие и адаптация грибов AM в нескольких луговых сообществах, [ 88 ] вплоть до глобального исследования разнообразия грибов AM, которое значительно увеличило описанное молекулярное разнообразие внутри типа Glomeromycota. [ 89 ]
Все последние достижения молекулярной генетики явно позволяют анализировать микробные сообщества в гораздо более тонких и функциональных масштабах и потенциально с большей уверенностью, чем предыдущие методы. Классический АМ-метод идентификации грибов путем извлечения спор из почвы и дальнейшего морфологического анализа спор. [ 90 ] чревато усложнением проблем из-за различных стратегий и форм AM-грибов, например, отсутствие споруляции у определенных видов, сезонность, высокая некультивируемость, возможная ошибочная идентификация (человеческая ошибка) и новые данные о многоядерных спорах. [ 91 ] и высокая генетическая изменчивость внутри клональных видов AM. [ 92 ] Из-за этих различных проблем в прошлом исследователи, вероятно, искажали истинный состав грибковых сообществ AM, присутствующих в любой момент времени или в любом месте. Кроме того, следуя традиционным методам экстракции, культивирования и микроскопической идентификации, невозможно определить активные/функционирующие популяции грибов AM, которые, вероятно, являются наиболее важными при попытке связать симбиотические взаимодействия и механизмы растения-AM с экологическими или экосистемными функциями. Это особенно актуально в случае анализа колонизации корней, который может определить процент корней, колонизированных AM-грибами. Основная проблема с этим анализом возникает в полевых почвах, которые одновременно содержат несколько видов грибов АМ в ассоциации с целевым растением (см. Экология АМ). Идентификация ассоциированных грибных симбионтов невозможна без использования молекулярных методов. Хотя генетический анализ грибковых сообществ AM значительно продвинулся за последнее десятилетие, методология еще не полностью усовершенствована. Ниже представлен обзор методов, используемых в молекулярно-генетическом анализе AM-грибов, а также их применение в исследованиях, будущие направления и некоторые их проблемы.
Обзор методов
[ редактировать ]ДНК/РНК
[ редактировать ]Генетический анализ AM-грибов из образцов почвы и корней варьируется в зависимости от их применимости для ответа на экологические или филогенетические вопросы. В анализах ДНК используются различные ядерные маркеры для описания AM-грибов и представления различных областей ядерного рибосомного оперона ( 18S рРНК ), обнаруженного во всех эукариотических организмах. Анализ ДНК AM-грибов с использованием этих маркеров начался в начале 1990-х годов. [ 93 ] и продолжают развиваться по сей день. Ген рРНК малой субъединицы (SSU), ген внутренней транскрибируемой спейсера ( ITS ) и ген рРНК большой субъединицы (LSU) в настоящее время являются наиболее распространенными используемыми ДНК-маркерами. Регион СГУ чаще всего использовался в экологических исследованиях, [ 94 ] в то время как регионы ITS и LSU преимущественно использовались в таксономических конструкциях типа Glomeromycota. [ 95 ]
qPCR и qRT-PCR
[ редактировать ]ПЦР в реальном времени или количественная ПЦР (кПЦР) становится хорошо зарекомендовавшим себя методом быстрой амплификации и одновременного количественного определения целевой ДНК грибов AM из биологических образцов (корней растений или почвы). Сравнительно недавние разработки в области маркеров qPCR позволяют исследователям исследовать относительную численность видов грибов AM внутри корней в тепличных экспериментах, а также в полевых условиях для идентификации местных грибковых сообществ AM.
Маркеры кПЦР для арбускулярных микоризных грибов будут состоять из АМ-специфичных праймеров и флуоресцентно-меченных гидролизных зондов. Эти специфические праймеры для АМ (обсуждаемые выше) могут быть выбраны исследователем, и это решение обычно зависит от поставленного вопроса, имеющихся ресурсов и готовности устранять неполадки в лаборатории.
Микрочип
[ редактировать ]Анализ микрочипов ДНК в настоящее время используется в исследованиях грибов AM для одновременного измерения экспрессии многих генов целевых видов или экспериментальных образцов. Наиболее распространенным инструментом или методом является использование технологии функционального генного массива (FGA), специализированного микрочипа, который содержит зонды для генов, которые функционально важны в микробных процессах, таких как круговорот углерода, азота или фосфора. FGA обладают способностью одновременно исследовать множество функциональных генов. [ 96 ] Этот метод обычно используется для общего анализа функциональных микробных генов, но в сочетании с генетическим секвенированием можно сделать выводы о связи между составом грибкового сообщества и функциональностью микробов.
ПЛФА / НЛФА
[ редактировать ]Специфические химические характеристики организмов можно использовать для обнаружения биомассы более загадочных организмов, таких как AM-грибы или почвенные бактерии. Липиды, точнее фосфолипиды и нейтральные липиды, содержат жирные кислоты, связанные с глицериновым остовом. Жирнокислотный состав организмов варьируется, и пропорции конкретных жирных кислот могут зависеть от организма. Например, у AM-грибов доли жирных кислот 16:1ω5 и 18:1ω7 в фосфолипидной части составляют примерно 58% общего состава жирных кислот. [ 97 ] Жирная кислота 16:1ω5 является наиболее часто используемой кислотой для характеристики AM-грибов в почвах и может использоваться в качестве надежного индикатора биомассы мицелия в образце почвы. [ 97 ]
Анализ нейтральных липидов и жирных кислот AM-грибов обычно рассматривается как метод определения запаса энергии, но, что наиболее важно, соотношение NLFA (16:1ω5) и PLFA (16:1ω5) потенциально может использоваться для определения пищевого статуса AM. грибы в почвах. Энергия в основном хранится у AM-грибов в виде нейтральных липидов в структурах хранения, таких как споры и везикулы. Благодаря этому NLFA достаточно хорошо коррелирует с количеством спор в данном объеме почвы. [ 97 ] Отношение концентрации NLFA к концентрации PLFA (активный мицелий) может затем дать долю углерода, выделенного в структуры хранения (споры, измеренные как NLFA).
Проблемы с анализом липидных жирных кислот включают неполную специфичность жирных кислот к АМ-грибам, видо- или родоспецифичные вариации в составе жирных кислот могут усложнить анализ в системах с несколькими видами АМ-грибов (например, полевая почва), высокие фоновые уровни определенная концентрация жирных кислот в почвах, а также то, что фосфолипиды коррелируют с площадью мембраны организма, а соотношение поверхности к объему может широко варьироваться у разных организмов, таких как бактерии и грибы. [ 98 ] Необходимо провести дополнительную работу для определения эффективности этого метода в полевых почвах со многими родами и видами грибов AM, чтобы выявить способность метода различать множество различных составов жирных кислот.
Будущие направления исследований AM-грибов
[ редактировать ]Одной из перспектив будущего анализа АМ-грибов является использование зондов со стабильными изотопами. Зондирование стабильными изотопами ( SIP ) — это метод, который можно использовать для определения активной метаболической функции отдельных таксонов в сложной системе микробов. Такой уровень специфичности, связывающий микробную функцию и филогенетику, ранее не был достигнут в микробной экологии. Этот метод также можно использовать независимо от классических методов культивирования в микробной экологии, позволяя проводить in situ анализ функциональных микробов . Применение секвенирования одного ядра из спор AM-грибов также было разработано недавно и также позволяет избежать необходимости использования методов культивирования. [ 99 ]
Зондирование стабильных изотопов (SIP)
[ редактировать ]SIP, точнее SIP на основе ДНК/РНК, использует субстраты, обогащенные стабильными изотопами, такие как 13 С, 15 Н, или Н 2 18 O, а затем анализирует «меченые» маркеры с использованием видоспецифичных маркеров ДНК или РНК. [ 100 ] Анализ меченой ДНК проводят путем разделения немеченой и меченой ДНК на градиенте хлорида цезия, сформированном в ультрацентрифуге. [ 101 ] Поскольку все микробные организмы способны импортировать воду в свои клетки, использование H 2 18 Зондирование стабильными изотопами — это очень интересный новый метод, который может пролить свет на вопросы, на которые микробные экологи и биологи пытались ответить в течение многих лет, в частности, какие микробные организмы являются активными в моей системе? Н 2 18 Метод O, или тяжелой воды, нацелен на все активно растущие организмы и мало влияет на сам рост. Это особенно верно для большинства тепличных экспериментов с арбускулярными микоризами, поскольку растения в любом случае необходимо поливать, а вода не отбирает напрямую организмы со специфическими метаболическими путями. [ 101 ] как это произойдет при использовании 13 С и 15 Н.
С этим методом мало что было сделано в экспериментах с арбускулярной микоризой, но если будет доказано, что он работает в контролируемом эксперименте, и при дальнейшем усовершенствовании методов анализа ДНК/РНК грибковых сообществ, это может стать жизнеспособным вариантом для очень специфичного определения активно растущей части грибкового сообщества. Виды грибов AM в разные вегетационные периоды, с разными растениями-хозяевами или обработками, а также в условиях изменения климата.
Механизм обработки мРНК и мРНК для понимания симбиоза AM
[ редактировать ]Сообщается, что мРНК играют решающую роль в перекрестных помехах между хозяином и симбионтом. [ 102 ] Таким образом, механизм процессинга мРНК важен для понимания симбиоза AM. Похоже, что AM-грибы обладают уникальными особенностями: они имеют основной фермент бактериального типа, а также большое количество белков Argonaute в их системе процессинга мРНК (или системе РНКи). [ 17 ] Исследование механизмов процессинга мРНК и мРНК также является интересной темой для понимания симбиоза AM-грибов.
Фиторемедиация
[ редактировать ]Нарушение местных растительных сообществ на территориях, находящихся под угрозой опустынивания , часто сопровождается деградацией физических и биологических свойств почвы, ее структуры, доступности питательных веществ и органических веществ. При восстановлении нарушенных земель важно заменить не только надземную растительность, но и биологические и физические свойства почвы. [ 103 ]
Относительно новый подход к восстановлению земель заключается в инокуляции почвы АМ-грибами при повторном внедрении растительности в проекты экологического восстановления (фиторемедиация). Это позволило растениям-хозяевам прижиться на деградированной почве и улучшить ее качество и здоровье. [ 104 ] Параметры качества почв значительно улучшились в долгосрочной перспективе при введении смеси аборигенных видов арбускулярных микоризных грибов по сравнению с неинокулированной почвой и почвой, инокулированной одним экзотическим видом АМ-грибов. [ 103 ] Преимущества заключались в увеличении роста растений и увеличении поглощения фосфора. [ 105 ] и содержание азота в почве, более высокое содержание органического вещества в почве и агрегацию почвы, что связано с более высоким клубнеобразованием бобовых в присутствии грибов AM, лучшей инфильтрацией воды и аэрацией почвы из-за агрегации почвы. [ 103 ] Нативные штаммы АМ-грибов усиливают извлечение тяжелых металлов из загрязненных почв и делают почву здоровой и пригодной для выращивания сельскохозяйственных культур. [ 106 ]
Сельское хозяйство
[ редактировать ]
на продуктивность растений и плодородие почвы
Справа: бактерии-помощники микоризы (MH) и бактерии, способствующие росту растений (PGP), синергически взаимодействующие с АМФ. [ 107 ]
Многие современные агрономические приемы разрушают микоризный симбиоз. Существует большой потенциал малозатратного сельского хозяйства для управления системой таким образом, чтобы способствовать микоризному симбиозу.
Традиционные методы ведения сельского хозяйства, такие как обработка почвы , тяжелые удобрения и фунгициды , плохие севообороты и отбор растений, выживающих в этих условиях, препятствуют способности растений образовывать симбиоз с арбускулярными микоризными грибами.
Большинство сельскохозяйственных культур могут расти лучше и продуктивнее, если они хорошо колонизированы AM-грибами. Симбиоз АМ увеличивает поглощение фосфора и микроэлементов и рост растения-хозяина (Джордж и др., 1992).
Борьба с АМ-грибами особенно важна для органических и малозатратных сельскохозяйственных систем, где фосфор в почве, как правило, низкий, хотя все агроэкосистемы могут получить выгоду от содействия формированию арбускулярной микоризы.
Некоторые культуры, которые плохо усваивают питательные вещества из почвы, очень зависят от поглощения фосфора АМ-грибами. Например, лен , обладающий плохой хемотаксической способностью, сильно зависит от АМ-опосредованного поглощения фосфора при низких и средних концентрациях фосфора в почве (Thingstrup et al. 1998).
Правильное управление АМФ в агроэкосистемах может улучшить качество почвы и продуктивность земли. Сельскохозяйственные методы, такие как минимальная обработка почвы, использование удобрений с низким содержанием фосфора и многолетние системы земледелия, способствуют функциональному микоризному симбиозу.
Обработка почвы
[ редактировать ]Обработка почвы снижает инокуляционный потенциал почвы и эффективность микоризы, разрушая внерадикальную сеть гиф (Miller et al. 1995, McGonigle & Miller 1999, Mozafar et al. 2000).
Разрушая макроструктуру почвы, гифальная сеть становится неинфекционной (Miller et al. 1995, McGonigle & Miller 1999). Нарушение сети гиф снижает поглотительные способности микоризы, поскольку площадь поверхности, охватываемая гифами, значительно уменьшается. Это, в свою очередь, снижает поступление фосфора в растения, подключенные к гифальной сети (рис. 3, McGonigle & Miller 1999).
В системе минимальной обработки почвы внесение тяжелых фосфорных удобрений может не потребоваться по сравнению с системами тяжелой обработки почвы. Это связано с увеличением микоризной сети, что позволяет микоризам обеспечивать растение достаточным количеством фосфора (Миллер и др., 1995).
Фосфорное удобрение
[ редактировать ]Преимущества AMF наиболее велики в системах с низкими затратами. Интенсивное использование фосфорных удобрений может препятствовать колонизации и росту микоризы.
По мере того, как уровень фосфора в почве, доступный растениям, увеличивается, количество фосфора также увеличивается в тканях растения, и утечка углерода из растения в результате симбиоза AM-грибов становится бесполезной для растения (Грант 2005).
Снижение микоризной колонизации из-за высокого уровня фосфора в почве может привести к дефициту у растений других микроэлементов, которые усваиваются микоризой, таких как медь (Timmer & Leyden 1980).
Многолетние системы земледелия
[ редактировать ]Покровные культуры выращивают осенью, зимой и весной, покрывая почву в периоды, когда она обычно остается без покрова растущих растений.
Микоризные покровные культуры можно использовать для улучшения потенциала микоризного инокулята и сети гиф (Кабир и Коиде, 2000, Босвелл и др., 1998, Соренсен и др., 2005).
Поскольку AM-грибы биотрофны, рост их гифальных сетей зависит от растений. Выращивание покровной культуры продлевает время роста AM до осени, зимы и весны. Стимулирование роста гиф создает более обширную сеть гиф. Увеличение микоризной колонизации, обнаруженное в системах покровных культур, можно в значительной степени объяснить увеличением экстрарадикальной сети гиф, которая может колонизировать корни новой культуры (Boswell et al. 1998). Экстрарадикальный мицелий способен пережить зиму, обеспечивая быструю весеннюю колонизацию и симбиоз в начале сезона (McGonigle and Miller 1999). Этот ранний симбиоз позволяет растениям использовать хорошо развитую сеть гифов и получать достаточное фосфорное питание во время раннего роста, что значительно повышает урожайность.
Качество почвы
[ редактировать ]Восстановление местных АМ-грибов увеличивает успех проектов экологического восстановления и скорость восстановления почвы. [ 103 ] АМ-грибы повышают стабильность почвенных агрегатов за счет производства экстрарадикальных гиф и почвенного белка, известного как гломалин .
Связанные с гломалином почвенные белки (GRSP) были идентифицированы с использованием моноклональных антител (Mab32B11), выработанных против измельченных спор AMF. Это определяется условиями его экстракции и реакцией с антителом Mab32B11.
Существуют и другие косвенные доказательства того, что гломалин имеет грибковое происхождение АМ. Когда AM-грибы удаляются из почвы путем инкубации почвы без растений-хозяев, концентрация GRSP снижается. Аналогичное снижение GRSP также наблюдалось в инкубированных почвах лесных, лесных и сельскохозяйственных земель. [ 108 ] и луга, обработанные фунгицидами. [ 109 ]
Предполагается, что гломалин улучшает водостойкость почвенных агрегатов и уменьшает эрозию почвы . Была обнаружена сильная корреляция между GRSP и водоустойчивостью почвенных агрегатов в самых разных почвах, где органический материал является основным связующим агентом, хотя механизм неизвестен. [ 109 ] Белок гломалин еще не выделен и не описан, а связь между гломалином, GRSP и арбускулярными микоризными грибами еще не ясна. [ 109 ]
См. также
[ редактировать ]- Эктомикориза
- Эрикоидная микориза
- Микориза
- Микориза и изменение климата
- Микоризные грибы и хранение углерода в почве
- Прототакситы
- Международная коллекция (везикулярных) арбускулярных микоризных грибов (INVAM)
- Glomus iranicus var. тонкие гифы
Ссылки
[ редактировать ]- ^ Микоризный симбиоз . 2008. doi : 10.1016/b978-0-12-370526-6.x5001-6 . ISBN 978-0-12-370526-6 .
- ^ Спатафора, Джозеф В.; Чанг, Ин; Бенни, Джеральд Л.; Лазарь, Кэти; Смит, Мэтью Э.; Берби, Мэри Л.; Бонито, Грегори; Корради, Николас; Григорьев Игорь; Грыганский Андрей; Джеймс, Тимоти Ю.; О'Доннелл, Керри; Роберсон, Роберт В.; Тейлор, Томас Н.; Юлинг, Джесси (2016). «Филогенетическая классификация зигомицетных грибов на уровне типа, основанная на данных масштаба генома» . Микология . 108 (5): 1028–1046. дои : 10.3852/16-042 . ISSN 0027-5514 . ПМК 6078412 . ПМИД 27738200 .
- ^ Брундретт, MC (2002). «Коэволюция корней и микоризы наземных растений» . Новый фитолог . 154 (2): 275–304. дои : 10.1046/j.1469-8137.2002.00397.x . ПМИД 33873429 .
- ^ Барбур, Миннесота; Берк, Дж. Х.; Питтс, В.Д. (1980). Экология наземных растений . Границы в физике. Издательская компания Бенджамина/Каммингса. п. 118 . ISBN 978-0-8053-0540-1 .
- ^ Перейти обратно: а б Саймон, Л.; Буске, Ж.; Левеск, К.; Лалонд, М. (1993). «Происхождение и разнообразие эндомикоризных грибов и совпадение с сосудистыми наземными растениями». Природа . 363 (6424): 67–69. Бибкод : 1993Natur.363...67S . дои : 10.1038/363067a0 . S2CID 4319766 .
- ^ Шюслер, А.; и др. (2001). «Новый тип грибов Glomeromycota : филогения и эволюция» . Микол. Рез . 105 (12): 1413–1421. дои : 10.1017/S0953756201005196 .
- ^ Мур, Дэвид; Робсон, Джеффри Д.; Тринчи, Энтони П.Дж. (2020), Путеводитель по грибам 21 века , номер документа : 10.1017/9781108776387 , ISBN 9781108776387
- ^ Смит, Салли А; Рид, Дэвид (2008), «2 - Колонизация корней и анатомия арбускулярных микориз» , Микоризный симбиоз (Третье изд.): 42–90, номер документа : 10.1016/B978-012370526-6.50004-0 , ISBN 9780123705266
- ^ Жакотт, Кэтрин; Мюррей, Джереми; Ридаут, Кристофер (2017). «Компромиссы в арбускулярном микоризном симбиозе: устойчивость к болезням, реакция роста и перспективы селекции сельскохозяйственных культур» . Агрономия . 7 (4): 75. doi : 10.3390/agronomy7040075 .
 В эту статью включен текст из этого источника, доступного по лицензии CC BY 4.0 .
В эту статью включен текст из этого источника, доступного по лицензии CC BY 4.0 .
- ^ Перейти обратно: а б с д и Реми, В.; Тейлор, Т.; Хасс, Х.; Керп, Х. (1994). «Везикулярная арбускулярная микориза возрастом четыреста миллионов лет» . Труды Национальной академии наук Соединенных Штатов Америки . 91 (25): 11841–11843. Бибкод : 1994PNAS...9111841R . дои : 10.1073/pnas.91.25.11841 . ПМЦ 45331 . ПМИД 11607500 .
- ^ Кар, РК; Мандаокар, Б.Д.; Кар, Р. (2005). «Микоризные ископаемые грибы из миоценовых отложений Мирозама, Северо-Восточная Индия». Современная наука . 89 : 257–259.
- ^ Струллу-Дерриен, Кристина; Струллу, Дезире-Жорж (ноябрь 2007 г.), «Микоризация ископаемых и живых растений» , Comptes Rendus Palevol , Палеоботаника и эволюция растительного мира: Некоторые текущие проблемы, 6 (6–7): 483–494, doi : 10.1016/j.crpv.2007.09.006
- ^ Перейти обратно: а б Крингс, Майкл; Харпер, Карла Дж; Тейлор, Эдит Л. (2018), «Грибы и грибковые взаимодействия в кремне Райни: обзор доказательств с описанием Perexiflasca tayloriana gen. et sp. nov», Philosophical Transactions of the Royal Society of London. Серия B, Биологические науки , 373 (1739), doi : 10.1098/rstb.2016.0500 , PMC 5745336 , PMID 29254965
- ^ Струллу-Дерриен, Кристина; Селосс, Марк-Андре; Кенрик, Пол; Мартин, Фрэнсис М. (24 марта 2018 г.), «Происхождение и эволюция микоризных симбиозов: от палеомикологии к филогеномике», New Phytologies , 220 (4): 1012–1030, doi : 10.1111/nph.15076 , ISSN 1469-8137 , PMID 29573278
- ^ Перейти обратно: а б Ван, Б.; Юн, Л.Х.; Сюэ, Ю.; Лю, Ю.; Ане, Дж. М.; Цю, ЮЛ (2010). «Наличие трех микоризных генов у общего предка наземных растений предполагает ключевую роль микоризы в колонизации суши растениями». Новый фитолог . 186 (2): 514–525. дои : 10.1111/j.1469-8137.2009.03137.x . hdl : 2027.42/78704 . ПМИД 20059702 .
- ^ Ане, Дж. М.; и др. (ноябрь 2002 г.). «Генетическое и цитогенетическое картирование генов DMI1, DMI2 и DMI3 Medicago truncatula, участвующих в трансдукции, узелковании и микоризации Nod-фактора» . Молекулярные растительно-микробные взаимодействия . 15 (11): 1108–18. дои : 10.1094/MPMI.2002.15.11.1108 . ПМИД 12423016 .
- ^ Перейти обратно: а б Ли, Сун-Джэ; Конг, Мэнсюань; Харрисон, Пол; Хиджри, Мохамед (2018), «Консервативные белки системы РНК-интерференции в арбускулярном микоризном грибе Rhizoglomus ignoree дают новый взгляд на эволюционную историю Glomeromycota», Genome Biology and Evolution , 10 (1): 328–343, doi : 10.1093 /gbe/evy002 , PMC 5786227 , PMID 29329439
- ^ Алексополус С., Мимс С., Блэквелл М. 2004. Вводная микология, 4-е изд. Хобокен (Нью-Джерси): Джон Уайли и сыновья.
- ^ Перейти обратно: а б Ли С.Дж., Конг М., Морс Д. и др. Экспрессия предполагаемых компонентов циркадных часов у арбускулярного микоризного гриба Rhizoglomus иррегулярный. Микориза (2018) https://doi.org/10.1007/s00572-018-0843-y
- ^ Марло Дж; Дальпе Ю; Сен-Арно М; Хиджри М (2011). «Развитие спор и ядерная наследственность у арбускулярных микоризных грибов» . Эволюционная биология BMC . 11 (1): 51. Бибкод : 2011BMCEE..11...51M . дои : 10.1186/1471-2148-11-51 . ПМК 3060866 . ПМИД 21349193 .
- ^ Перейти обратно: а б с д Райт С.Ф. (2005). «Борьба с арбускулярными микоризными грибами». В Р.В. Зобеле; С. Ф. Райт (ред.). Управление корнями и почвой: взаимодействие между корнями и почвой . США: Американское общество агрономии. стр. 183–197.
- ^ Перейти обратно: а б с д Даудс Д.Д. и Нагахаши Г. 2000. События сигнализации и распознавания до колонизации корней арбускулярными микоризными грибами. В текущих достижениях в исследованиях микоризы. Эд. Подила, Г.К. , Даудс, Д.Д. Миннесота: APS Press. Стр. 11–18.
- ^ Акияма К; Мацузаки К; Хаяши Х (2005). «Растительные сесквитерпены вызывают ветвление гиф у арбускулярных микоризных грибов». Природа . 435 (7043): 824–827. Бибкод : 2005Natur.435..824A . дои : 10.1038/nature03608 . ПМИД 15944706 . S2CID 4343708 .
- ^ Перейти обратно: а б с д Нагахаши, Дж; Даудс, Д.Д.; Эбни, Джорджия (1996). «Фосфорная добавка прямо и косвенно ингибирует ветвление гиф гриба VAM Gigaspora margarita за счет его влияния на корневую экссудацию» . Микориза . 6 (5): 403–408. Бибкод : 1996Mycor...6..403N . дои : 10.1007/s005720050139 . S2CID 36014515 .
- ^ Сбрана, К.; Джованнетти, М. (2005). «Хемотропизм арбускулярного микоризного гриба Glomus mosseae». Микориза . 15 (7): 539–545. Бибкод : 2005Mycor..15..539S . дои : 10.1007/s00572-005-0362-5 . ПМИД 16133246 . S2CID 23648484 .
- ^ Тамаслоухт, М.; Сехалон-Дельмас, Н.; Клювер, А.; Жоно, А.; Ру, К.; Бекар, Г.; Франкен, П. (2003). «Корневые факторы индуцируют экспрессию митохондриальных генов и дыхание грибов во время переключения развития от асимбиоза к пресимбиозу у арбускулярного микоризного гриба Gigaspora rosea» . Физиология растений . 131 (3): 1468–1478. дои : 10.1104/стр.012898 . ПМК 166906 . ПМИД 12644696 .
- ^ Перейти обратно: а б с Джанинацци-Пирсон, В. (1996). «Реакция растительных клеток на арбускулярные микоризные грибы: добраться до истоков симбиоза» . Растительная клетка . 8 (10): 1871–1883. дои : 10.1105/tpc.8.10.1871 . JSTOR 3870236 . ПМК 161321 . ПМИД 12239368 .
- ^ Лара Армстронг; Р. Ларри Петерсон; Лара Армстронг; Р. Ларри Петерсон (2002). «Взаимодействие между арбускулярным микоризным грибом Glomus interradices и корневыми клетками Panax quinquefolius: микоризная ассоциация парижского типа» . Микология . 94 (4): 587–595. дои : 10.2307/3761710 . JSTOR 3761710 . ПМИД 21156532 .
- ^ Ямато, Масахидэ (2005). «Морфологические типы арбускулярных микориз у пионеров древесных растений, произрастающих на ферме по выращиванию масличных пальм на Суматре, Индонезия». Миконаука . 46 : 66–68. дои : 10.1007/s10267-004-0212-x . S2CID 83767351 .
- ^ Матеквор, Ахулу Э; Наката, М; Нонака, М. (март 2005 г.). «Арбускулярные микоризы типа Arum и Paris в смешанном сосновом лесу на почве песчаных дюн в префектуре Ниигата, центральный Хонсю, Япония». Микориза . 15 (2): 129–36. Бибкод : 2005Mycor..15..129M . дои : 10.1007/s00572-004-0310-9 . ISSN 0940-6360 . ПМИД 15290409 . S2CID 25476630 .
- ^ Туоми, Дж.; Китовиита, М.; Хардлинг, Р. (2001). «Экономичность приобретения питательных веществ при микоризном симбиозе для растения-хозяина» . Ойкос . 92 : 62–70. дои : 10.1034/j.1600-0706.2001.920108.x .
- ^ Перейти обратно: а б с Болан, Н.С. (1991). «Критический обзор роли микоризных грибов в поглощении фосфора растениями». Растение и почва . 134 (2): 189–207. Бибкод : 1991ПлСой.134..189Б . дои : 10.1007/BF00012037 . S2CID 44215263 .
- ^ Харли, Дж. Л., Смит, С. Е., 1983. Микоризный симбиоз. Академическая пресса: Лондон.
- ^ Перейти обратно: а б с д Пфеффер, П.; Даудс Д.; Бекар, Г.; Шачар-Хилл, Ю. (1999). «Поглощение углерода, метаболизм и транспорт липидов в арбускулярной микоризе» . Физиология растений . 120 (2): 587–598. дои : 10.1104/стр.120.2.587 . ПМК 59298 . ПМИД 10364411 .
- ^ Перейти обратно: а б с Хамель, К. (2004). «Влияние грибов арбускулярной микоризы на круговорот N и P в корневой зоне» . Канадский журнал почвоведения . 84 (4): 383–395. дои : 10.4141/S04-004 .
- ^ Х. Букинг и Ю.Шачар-Хилл (2005). Поглощение, транспорт и перенос фосфатов арбускулярным микоризным грибом Glomus Intraradices стимулируется увеличением доступности углеводов. Новый Phytologies 165: 899–912
- ^ Перейти обратно: а б с Смит, С.; Смит, А.; Якобсен, И. (2003). «Микоризные грибы могут доминировать в снабжении растений фосфатами независимо от реакции роста» . Физиология растений . 133 (1): 16–20. дои : 10.1104/стр.103.024380 . ПМЦ 1540331 . ПМИД 12970469 .
- ^ Перейти обратно: а б Матеус, И.Д.; Осье, Б.; Ндиайе ММС; Круз, Дж.; Ли, С.Дж.; Сандерс, ИК (2022 г.). «Геномные признаки реципрокной рекомбинации у симбиотических арбускулярных микоризных грибов Rhizophagus ignores» . ПЛОС ОДИН . 17 (7): e0270481. Бибкод : 2022PLoSO..1770481M . дои : 10.1371/journal.pone.0270481 . ПМК 9249182 . PMID 35776745 .
- ^ Халари, Себастьян; Малик, Шере-Бану; Лилдхар, Леванния; Сламовиц, Клаудио Х.; Хиджри, Мохамед; Корради, Николя (2011). «Консервативный мейотический механизм у Glomus SPP., предположительно древней бесполой линии грибов» . Геномная биология и эволюция . 3 : 950–958. дои : 10.1093/gbe/evr089 . ПМЦ 3184777 . ПМИД 21876220 .
- ^ Майе, Фабьен; Пуансо, Верена; Андре, Оливье; Пуэх-Пажес, Вирджиния; Хауи, Александра; Генье, Моник; Кромер, Лоуренс; Жироде, Дельфин; Форми, Дэмиен; Нибель, Андреас; Мартинес, Эдуардо Андрес; Дригес, Хьюг; Бекар, Гийом; Денари, Жан (январь 2011 г.). «Симбиотические сигналы грибковых липохитоолигосахаридов при арбускулярной микоризе». Природа . 469 (7328): 58–63. Бибкод : 2011Природа.469...58М . дои : 10.1038/nature09622 . ПМИД 21209659 . S2CID 4373531 .
- ^ Коуп, Кевин Р.; Баскюль, Аделина; Ирвинг, Томас Б.; Венкатешваран, Мутусубраманиан; Маэда, Джунко; Гарсия, Кевин; Раш, Томас А.; Ма, Кэтлин; Лаббе, Джесси; Джоуди, Сара; Штайгервальд, Эдвард; Сецке, Джонатан; Фунг, Эммелин; Шнелл, Кимберли Г.; Ван, Юньцянь; Шлейф, Натаниэль; Бюкинг, Хайке; Штраус, Стивен Х.; Майе, Фабьен; Джаргит, Патрисия; Бекар, Гийом; Пуэх-Пажес, Вирджиния; Ане, Жан-Мишель (октябрь 2019 г.). «Эктомикоризный гриб Laccaria bicolor продуцирует липохитоолигосахариды и использует общий путь симбиоза для колонизации корней Populus» . Растительная клетка . 31 (10): 2386–2410. дои : 10.1105/tpc.18.00676 . ПМК 6790088 . ПМИД 31416823 .
- ^ Маклин, Эллисон М.; Браво, Армандо; Харрисон, Мария Дж. (октябрь 2017 г.). «Сигнальные сигналы растений и метаболические пути, обеспечивающие арбускулярный микоризный симбиоз» . Растительная клетка . 29 (10): 2319–2335. дои : 10.1105/tpc.17.00555 . ПМК 5940448 . ПМИД 28855333 .
- ^ Перейти обратно: а б с д и ж г Смит, С.Э., Рид DJ «Микоризный симбиоз». 2002. Академическая пресса: Лондон.
- ^ Перейти обратно: а б Смит, Рид, Салли, диджей (2008). Микоризный симбиоз . Нью-Йорк: Академическая пресса.
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Перейти обратно: а б с Опик, М; Ванатоа А; Ванатоа Э; Мура М; Дэвидсон Дж; Калвидж Дж.М.; Рейер У; Зобель М (2010). «Онлайн-база данных MaarjAM раскрывает глобальные и экосистемные закономерности распространения арбускулярных микоризных грибов (Glomeromycota)» . Новый фитолог . 188 (1): 233–241. дои : 10.1111/j.1469-8137.2010.03334.x . ПМИД 20561207 .
- ^ Перейти обратно: а б Кивлин, Стефани; Кристин В. Хоукс; Кэтлин К. Треседер (2011). «Глобальное разнообразие и распространение арбускулярных микоризных грибов» . Биология и биохимия почвы . 43 (11): 2294–2303. Бибкод : 2011SBiBi..43.2294K . doi : 10.1016/j.soilbio.2011.07.012 . S2CID 85135958 .
- ^ Лекберг, Ю; Койде РТ; Рор-младший; Олдирч-Вульф Л; Мортон Дж. Б. (2007). «Роль нишевых ограничений и расселения в составе арбускулярных микоризных грибных сообществ» . Журнал экологии . 95 (1): 95–100. Бибкод : 2007JEcol..95...95L . дои : 10.1111/j.1365-2745.2006.01193.x .
- ^ Лефф, Джонатан В.; Джонс, Стюарт Э.; Пробер, Сюзанна М.; Барберан, Альберт; Борер, Элизабет Т .; Фирн, Дженнифер Л.; Харпол, В. Стэнли; Хобби, Сара Э.; Хофмокель, Кирстен С. (1 сентября 2015 г.). «Последовательная реакция почвенных микробных сообществ на повышенное поступление питательных веществ на луга по всему миру» . Труды Национальной академии наук . 112 (35): 10967–10972. Бибкод : 2015PNAS..11210967L . дои : 10.1073/pnas.1508382112 . ISSN 0027-8424 . ПМЦ 4568213 . ПМИД 26283343 .
- ^ Аллен, Э.Б.; Аллен М.Ф.; Хелм-диджей; Траппе Дж.М.; Молина Р; Ринкон Э (1995). «Закономерности и регулирование разнообразия микоризных растений и грибов». Растение и почва . 170 (1): 47–62. Бибкод : 1995ПлСой.170...47А . дои : 10.1007/bf02183054 . S2CID 25097125 .
- ^ Клирономос, Джон (2000). Специфичность хозяина и функциональное разнообразие арбускулярных микоризных грибов . Галифакс, Канада: Микробные биосистемы: новые рубежи. Материалы VIII Международного симпозиума по микробной экологии. Атлантическое канадское общество микробной экологии. стр. 845–851.
- ^ Муж, Р; Герре Э.А.; Тернер С.Л.; Галерея Р; Молодой JPW (2002). «Молекулярное разнообразие арбускулярных микоризных грибов и закономерности ассоциаций во времени и пространстве в тропическом лесу» (PDF) . Молекулярная экология . 11 (12): 2669–2678. дои : 10.1046/j.1365-294x.2002.01647.x . ПМИД 12453249 . S2CID 10279037 .
- ^ Перейти обратно: а б Хемпель, Стефан; Готценбергер Л; Кун I; Михальский С.Г.; Риллиг М; Зобель М; Мура М (2013). «Микоризы во флоре Центральной Европы – связь с особенностями истории жизни растений и экологией». Экология . 94 (6): 1389–1399. Бибкод : 2013Ecol...94.1389H . дои : 10.1890/12-1700.1 . ПМИД 23923502 .
- ^ Дриго, Б; Пийл, А.С.; Дуйц, Х; Килак, AM; Гампер, штат Ха; Хоукамер, MJ; Бошкер, HTS; Боделье, PLE; Уайтли, А.С.; Вин, ЯВ; Ковальчук Г.А. (2010). «Смещение потока углерода от корней к связанным микробным сообществам в ответ на повышенное содержание CO 2 в атмосфере » . Труды Национальной академии наук Соединенных Штатов Америки . 107 (24): 10938–10942. Бибкод : 2010PNAS..10710938D . дои : 10.1073/pnas.0912421107 . ПМЦ 2890735 . ПМИД 20534474 .
- ^ ван дер Хейден, MG; Боллер А.Т.; Вимкен А; Сандерс И.Р. (1998). «Различные виды арбускулярных микоризных грибов являются потенциальными детерминантами структуры растительного сообщества». Экология . 79 (6): 2082–2091. doi : 10.1890/0012-9658(1998)079[2082:damfsa]2.0.co;2 .
- ^ ван дер Хейден, MGA; Барджетт Р.Д.; Ван Страален, Н.М. (2008). «Невидимое большинство: почвенные микробы как движущая сила разнообразия и продуктивности растений в наземных экосистемах» . Экологические письма . 11 (3): 296–310. Бибкод : 2008EcolL..11..296V . дои : 10.1111/j.1461-0248.2007.01139.x . ПМИД 18047587 .
- ^ Ваннетт, РЛ; Расманн С (2012). «Арбускулярные микоризные грибы опосредуют взаимодействие подземных растений и травоядных: филогенетическое исследование» . Функциональная экология . 26 (5): 1033–1042. Бибкод : 2012FuEco..26.1033V . дои : 10.1111/j.1365-2435.2012.02046.x .
- ^ Хауманн, Н.; Резюме Хоукса (2009). «Контроль соседства растений за составом арбускулярного микоризного сообщества» . Новый фитолог . 183 (4): 1188–1200. дои : 10.1111/j.1469-8137.2009.02882.x . ПМИД 19496954 .
- ^ Хаусманн, Н; Резюме Хоукса (2010). «Порядок установления растений-хозяев меняет состав арбускулярных микоризных сообществ». Экология . 91 (8): 2333–23343. Бибкод : 2010Ecol...91.2333H . дои : 10.1890/09-0924.1 . ПМИД 20836455 .
- ^ Баттен, КМ; Скоу К.М.; Дэвис К.Ф.; Харрисон С.П. (2006). «Два инвазивных растения изменяют состав микробного сообщества почвы на змеевидных лугах». Биологические инвазии . 8 (2): 217–230. Бибкод : 2006BiInv...8..217B . дои : 10.1007/s10530-004-3856-8 . S2CID 24969103 .
- ^ Хоукс, резюме; Белнап Дж; Д'Антонио С; Файерстоун М (2006). «Арбускулярные микоризные комплексы в корнях местных растений изменяются в присутствии инвазивных экзотических трав». Растение и почва . 281 (1–2): 369–380. Бибкод : 2006PlSoi.281..369H . дои : 10.1007/s11104-005-4826-3 . S2CID 16801874 .
- ^ Кивлин, Стефани; Кристин В. Хоукс (2011). «Дифференциация эффектов инвазии и разнообразия: влияние надземных растительных сообществ на подземные грибковые сообщества» . Новый фитолог . 189 (2): 526–535. дои : 10.1111/j.1469-8137.2010.03494.x . ПМИД 20958304 .
- ^ Лекберг, Ю; Гиббонс С.М.; Розендаль С; Рэмси П.В. (2013). «Сильное вторжение растений может увеличить численность и разнообразие микоризных грибов» . Журнал ИСМЕ . 7 (7): 1424–1433. Бибкод : 2013ISMEJ...7.1424L . дои : 10.1038/ismej.2013.41 . ПМЦ 3695300 . ПМИД 23486251 .
- ^ Фиттер, АХ (2005). «Тьма видна: размышления о подземной экологии» (PDF) . Журнал экологии . 93 (2): 231–243. Бибкод : 2005JEcol..93..231F . дои : 10.1111/j.0022-0477.2005.00990.x .
- ^ Прингл, А; Бевер, Джей Ди; Гард, М; Паррент, Дж. Л.; Риллиг, MC; Клирономос, Дж. Н. (2009). «Микоризные симбиозы и инвазии растений». Ежегодный обзор экологии, эволюции и систематики . 40 : 699–715. doi : 10.1146/annurev.ecolsys.39.110707.173454 .
- ^ Лаример, Алабама; Бевер Дж.Д.; Клэй К. (2010). «Интерактивные эффекты растительных микробных симбионтов: обзор и метаанализ». Симбиоз . 51 (2): 139–148. Бибкод : 2010Symbi..51..139L . дои : 10.1007/s13199-010-0083-1 . S2CID 11569239 .
- ^ Новас, М.В.; Янноне LJ; Годеас AM; Кабрал Д. (2009). «Положительная связь между микоризой и эндофитами листьев у Poa bonariensis». Микологический прогресс . 8 : 75–81. дои : 10.1007/s11557-008-0579-8 . hdl : 11336/27622 . S2CID 2206235 .
- ^ Лаример, Алабама; Бевер Дж.Д.; Клэй К. (2012). «Последствия одновременного взаимодействия грибных эндофитов и арбускулярных микоризных грибов с общей травой-хозяином». Ойкос . 121 (12): 2090–2096. Бибкод : 2012Oikos.121.2090L . дои : 10.1111/j.1600-0706.2012.20153.x .
- ^ Омачини, М; Эггерс Т; Бонковский М; Ганг переменного тока; Джонс Т.Х. (2006). «Эндофиты листьев влияют на микоризный статус и рост коинфицированных и соседних растений». Функциональная экология . 20 (2): 226–232. Бибкод : 2006FuEco..20..226O . дои : 10.1111/j.1365-2435.2006.01099.x .
- ^ Мак, КМЛ; Радджерс Дж. А. (2008). «Балансирование нескольких мутуалистов: асимметричные взаимодействия между растениями, арбускулярными микоризными грибами и грибковыми эндофитами». Ойкос . 117 (2): 310–320. Бибкод : 2008Оикос.117..310М . CiteSeerX 10.1.1.722.4169 . дои : 10.1111/j.2007.0030-1299.15973.x .
- ^ Лю, QH; Парсонс Эй Джей; Сюэ Х; Фрейзер К; Райан Дж.Д.; Ньюман Дж.А.; Расмуссен С (2011). «Конкуренция между лиственными эндофитами Neotyphodium lolii и микоризными грибами Glomus spp. в Lolium perenne зависит от наличия ресурсов и содержания углеводов в хозяине». Функциональная экология . 25 (4): 910–920. Бибкод : 2011FuEco..25..910L . дои : 10.1111/j.1365-2435.2011.01853.x . hdl : 10214/3316 .
- ^ Райнингер, В; Зибер Т.Н. (2012). «Микориза уменьшает неблагоприятное воздействие темных перегородчатых эндофитов (DSE) на рост хвойных деревьев» . ПЛОС ОДИН . 7 (8): 1–10. Бибкод : 2012PLoSO...742865R . дои : 10.1371/journal.pone.0042865 . ПМК 3416760 . ПМИД 22900058 .
- ^ Клирономос, Дж. Н.; Харт ММ; Герни Дж. Э.; Мутоглис П. (2001). «Межвидовые различия в толерантности арбускулярных микоризных грибов к замораживанию и высыханию». Канадский журнал ботаники . 79 (10): 1161–1166. дои : 10.1139/cjb-79-10-1161 .
- ^ Оге, Р.М. (2001). «Водные отношения, засуха и везикулярно-арбускулярный микоризный симбиоз». Микориза . 11 (1): 3–42. Бибкод : 2001Mycor..11....3A . дои : 10.1007/s005720100097 . S2CID 29379395 .
- ^ Джонсон, Северная Каролина; Роуленд Д.Л.; Коркиди Л; Эгертон-Уорбертон LM; Аллен Э.Б. (2003). «Обогащение азота изменяет распределение микоризы на пяти мезических и полузасушливых лугах». Экология . 84 (7): 1895–1908. doi : 10.1890/0012-9658(2003)084[1895:neamaa]2.0.co;2 .
- ^ Джонсон, Северная Каролина; Уилсон GWT; Боукер М.А.; Уилсон Дж.А.; Миллер Р.М. (2010). «Ограничение ресурсов является движущей силой местной адаптации в микоризных симбиозах» . Труды Национальной академии наук Соединенных Штатов Америки . 107 (5): 2093–2098. Бибкод : 2010PNAS..107.2093J . дои : 10.1073/pnas.0906710107 . ПМЦ 2836645 . ПМИД 20133855 .
- ^ Гай, Япония; Тянь Х; Ян Ф.Ю.; Кристи П; Ли XL; Клирономос Ю.Н. (2012). «Разнообразие арбускулярных микоризных грибов вдоль градиента высот Тибета». Педобиология . 55 (3): 145–151. Бибкод : 2012Педоб..55..145Г . дои : 10.1016/j.pedobi.2011.12.004 .
- ^ Порсель, Р; Арока, Р; Руис-Лозано, ЖМ (2012). «Снятие стресса с помощью арбускулярных микоризных грибов. Обзор» (PDF) . Агрономия для устойчивого развития . 32 : 181–200. дои : 10.1007/s13593-011-0029-x . S2CID 8572482 .
- ^ Перейти обратно: а б с Маршнер, П.; Тимонен, С. (2004). «Взаимодействие между видами растений и микоризной колонизацией на состав бактериального сообщества в ризосфере». Прикладная экология почв . 28 : 23–36. дои : 10.1016/j.apsoil.2004.06.007 .
- ^ Эрикссон, А. (2001). «Арбускулярная микориза в связи с историей управления, питательными веществами почвы и разнообразием растений». Экология растений . 155 (2): 129–137. дои : 10.1023/А:1013204803560 . S2CID 42097761 .
- ^ «Липохитоолигосахариды, стимулирующие арбускулярный микоризный симбиоз» . Архивировано из оригинала 5 августа 2012 г.
- ^ Шрей, Сильвия Д.; и др. (2012). «Продукция вторичных метаболитов, модулирующих рост грибков и бактерий, широко распространена среди стрептомицетов, ассоциированных с микоризой» . БМК Микробиология . 12 (1): 164. дои : 10.1186/1471-2180-12-164 . ПМЦ 3487804 . ПМИД 22852578 .
- ^ Ван дер Путтен, WH (2012). «Изменение климата, взаимодействие надземных и подземных территорий и сдвиги ареала видов». Ежегодный обзор экологии, эволюции и систематики . 43 : 365–383. doi : 10.1146/annurev-ecolsys-110411-160423 . hdl : 20.500.11755/0f7d61b9-e435-4da3-b3d6-3ca115bdbdb5 . S2CID 85941864 .
- ^ Уорчел, Элиза; Джаок, Ханна Э.; Кивлин, Стефани Н. (2013). «Грибные симбионты изменяют реакцию растений на засуху». Микробная экология . 65 (3): 671–678. Бибкод : 2013MicEc..65..671W . дои : 10.1007/s00248-012-0151-6 . ПМИД 23250115 . S2CID 122731 .
- ^ Кивлин, С.Н.; Эмери С.М.; Радджерс Дж. А. (2013). «Грибные симбионты изменяют реакцию растений на глобальные изменения». Американский журнал ботаники . 100 (7): 1445–1457. дои : 10.3732/ajb.1200558 . ПМИД 23757444 .
- ^ Треседер, К.К. (2004). «Метаанализ реакций микоризы на азот, фосфор и атмосферный CO 2 в полевых исследованиях» . Новый фитолог . 164 (2): 347–355. дои : 10.1111/j.1469-8137.2004.01159.x . ПМИД 33873547 .
- ^ Гломалин. «Тайник трети мирового запаса почвенного углерода». Журнал сельскохозяйственных исследований .
- ^ Стукенброк, Ева; Розендаль, Сорен (2005). «Клональное разнообразие и популяционно-генетическая структура арбускулярных микоризных грибов (Glomus spp.), изученных методом мультилокусного генотипирования одиночных спор». Молекулярная экология . 14 (3): 743–752. Бибкод : 2005MolEc..14..743S . дои : 10.1111/j.1365-294x.2005.02453.x . ПМИД 15723666 . S2CID 30799196 .
- ^ Баомин, Цзи; и др. (2013). «Модели разнообразия и адаптации Glomeromycota из трех лугов прерий». Молекулярная экология . 22 (9): 2573–2587. Бибкод : 2013MolEc..22.2573J . дои : 10.1111/mec.12268 . ПМИД 23458035 . S2CID 6769986 .
- ^ Опик, Маарья; и др. (2013). «Глобальный отбор проб корней растений расширяет описанное молекулярное разнообразие арбускулярных микоризных грибов». Микориза . 23 (5): 411–430. Бибкод : 2013Mycor..23..411O . дои : 10.1007/s00572-013-0482-2 . ПМИД 23422950 . S2CID 17162006 .
- ^ Джонсон, Нэнси; Зак, доктор медицинских наук; Тилман, Д.; Пфлегер, Флорида (1991). «Динамика везикулярных арбускулярных микориз в ходе старой полевой сукцессии». Экология . 86 (3): 349–358. Бибкод : 1991Oecol..86..349J . дои : 10.1007/bf00317600 . ПМИД 28312920 . S2CID 31539360 .
- ^ Марло, Джули; Дальпе, Ю .; Сен-Арно, М.; Хиджри, М. (2011). «Развитие спор и ядерная наследственность у арбускулярных микоризных грибов» . Эволюционная биология BMC . 11 (1): 51. Бибкод : 2011BMCEE..11...51M . дои : 10.1186/1471-2148-11-51 . ПМК 3060866 . ПМИД 21349193 .
- ^ Эхингер, Мартина О.; Дэниел Кролл; Александр М. Кох; Ян Р. Сандерс (2012). «Значительные генетические и фенотипические изменения, возникающие в результате клонального роста одной споры арбускулярного микоризного гриба на протяжении нескольких поколений» . Новый фитолог . 196 (3): 853–861. дои : 10.1111/j.1469-8137.2012.04278.x . ПМИД 22931497 .
- ^ Саймон, Л; Лалонд, М.; Брунс, Т.Д. (1992). «Специфическая амплификация 18S грибковых рибосомальных генов из везикулярно-арбускулярных эндомикоризных грибов, колонизирующих корни» . Прил. Окружающая среда. Микробиол . 58 (1): 291–295. Бибкод : 1992ApEnM..58..291S . дои : 10.1128/АЕМ.58.1.291-295.1992 . ПМК 195206 . ПМИД 1339260 .
- ^ Хельгасон, Торунн; Фиттер, АХ; Янг, JPW (1999). «Молекулярное разнообразие арбускулярных микоризных грибов, колонизирующих Hyacinthoides non-scripta (колокольчик) в полуестественных лесах» (PDF) . Молекулярная экология . 8 (4): 659–666. Бибкод : 1999MolEc...8..659H . дои : 10.1046/j.1365-294x.1999.00604.x . S2CID 85991904 .
- ^ Редекер, Дирк; Артур Шюслер; Герберт Стокингер; Сидни Л. Штюрмер; Джозеф Б. Мортон; Кристофер Уокер (2013). «Обоснованный консенсус по классификации арбускулярных микоризных грибов (Glomeromycota)». Микориза . 23 (7): 515–531. Бибкод : 2013Mycor..23..515R . дои : 10.1007/s00572-013-0486-y . ПМИД 23558516 . S2CID 16495856 .
- ^ Он, Чжили; Джой Д. ВАН НОСТРАНД; Йе ДЭН; Цзичжун ЧЖОУ (2011). «Разработка и применение функциональных генных микрочипов для анализа функционального разнообразия, состава и структуры микробных сообществ». Передний. Окружающая среда. наук. англ. Китай . 5 (1): 1–20. дои : 10.1007/s11783-011-0301-y .
- ^ Перейти обратно: а б с Олссон, Пал Аксель (1999). «Сигнатурные жирные кислоты служат инструментом для определения распределения и взаимодействия микоризных грибов в почве» . ФЭМС Микробиология Экология . 29 (4): 303–310. Бибкод : 1999FEMME..29..303O . дои : 10.1111/j.1574-6941.1999.tb00621.x .
- ^ Дж., Шнурер; Клархольм, М.; Россволл, Т. (1985). «Микробная биомасса и активность в сельскохозяйственной почве с различным содержанием органического вещества». Биология и биохимия почвы . 17 (5): 611–618. Бибкод : 1985SBiBi..17..611S . дои : 10.1016/0038-0717(85)90036-7 .
- ^ Монтолиу-Нерин, Мерсе; Санчес-Гарсия, Марисоль; Бергин, Клаудия; Грабхерр, Манфред; Эллис, Барбара; Кучера, Верена Эстер; Кирчак, Марцин; Йоханнессон, Ханна; Рослинг, Анна (28 января 2020 г.). «Создание de novo эталонных геномных сборок сложных эукариотических микроорганизмов из одиночных ядер» . Научные отчеты . 10 (1): 1303. Бибкод : 2020НатСР..10.1303М . дои : 10.1038/s41598-020-58025-3 . ISSN 2045-2322 . ПМК 6987183 . ПМИД 31992756 .
- ^ Дюмон, Марк Г.; Мюррелл, Колин Дж. (2005). «Зондирование стабильными изотопами - связь микробной идентичности с функцией». Обзоры природы Микробиология . 3 (6): 499–504. дои : 10.1038/nrmicro1162 . ПМИД 15886694 . S2CID 24051877 .
- ^ Перейти обратно: а б Шварц, Эгберт (2007). «Характеристика растущих микроорганизмов в почве методом зондирования стабильными изотопами H218O» . Прикладная и экологическая микробиология . 73 (8): 2541–2546. Бибкод : 2007ApEnM..73.2541S . дои : 10.1128/АЕМ.02021-06 . ПМЦ 1855593 . ПМИД 17322324 .
- ^ Форми Д. и др. 2016. Регуляция малых РНК и соответствующих мишеней в волосковых клетках корня Phaseolus vulgaris, индуцированных фактором Nod. Int J Mol Sci. 176:887.
- ^ Перейти обратно: а б с д Джеффрис, П.; Джанинацци, С.; Перотто, С.; Турнау, К.; Бареа, Дж. (2003). «Вклад арбускулярных микоризных грибов в устойчивое поддержание здоровья растений и плодородия почвы». Биология и плодородие почв . 37 (1): 1–16. Бибкод : 2003BioFS..37....1J . дои : 10.1007/s00374-002-0546-5 . S2CID 20792333 .
- ^ Ахтар, Овайд; Мишра, Рани; Кери, Харбанс Каур (01 марта 2019 г.). «Ассоциация арбускулярной микоризы способствует накоплению хрома и устойчивости у растений, растущих на загрязненных хромом почвах». Труды Национальной академии наук Индии. Раздел B: Биологические науки . 89 (1): 63–70. дои : 10.1007/s40011-017-0914-4 . ISSN 2250-1746 . S2CID 46007322 .
- ^ Филлион, М.; Бриссон, Дж.; Гуиди В.; Лабрек, М. (2011). «Повышение удаления фосфора в фильтрах растительности ивы и тополя с использованием арбускулярных микоризных грибов». Экологическая инженерия . 37 (2): 199–205. Бибкод : 2011EcEng..37..199F . дои : 10.1016/j.ecoleng.2010.09.002 .
- ^ Ахтар, Овайд; Кери, Харбанс Каур; Зуми, Ифра (15 сентября 2020 г.). «Инокуляция арбускулярной микоризы и Aspergillus terreus вместе с добавкой компоста усиливает фиторемедиацию технозоля, богатого хромом, с помощью Solanum lycopersicum в полевых условиях» . Экотоксикология и экологическая безопасность . 201 : 110869. Бибкод : 2020EcoES.20110869A . дои : 10.1016/j.ecoenv.2020.110869 . ISSN 0147-6513 . ПМИД 32585490 . S2CID 220073862 .
- ^ Джованнини, Лука; Болл, Микела; Аньолуччи, Моника; Авио, Лучано; Сбрана, Кристиана; Туррини, Алессандра; Джованнетти, Мануэла (2020). «Арбускулярные микоризные грибы и связанная с ними микробиота как биостимуляторы растений: стратегии исследований для выбора наиболее эффективной инокуляты» . Агрономия . 10 : 106. doi : 10.3390/agronomy10010106 . В эту статью включен текст из этого источника, доступного по лицензии CC BY 4.0 .
- ^ Риллиг, М.; Рэмси, П.; Моррис, С.; Пол, Э. (2003). «Гломалин, арбускулярно-микоризный грибковый почвенный белок, реагирует на изменения в землепользовании». Растение и почва . 253 (2): 293–299. дои : 10.1023/А:1024807820579 . S2CID 11007821 .
- ^ Перейти обратно: а б с Риллиг, М. (2004). «Арбускулярные микоризы, гломалин и почвенная агрегация» . Канадский журнал почвоведения . 84 (4): 355–363. дои : 10.4141/S04-003 .
Примечания
[ редактировать ]- Босуэлл, EP; РТ Койде; Д.Л. Шамуэй; HD Адди. (1998). «Посев озимой пшеницы, рост и урожайность микоризных грибов ВА, кукурузы» . Сельское хозяйство, экосистемы и окружающая среда . 67 (1): 55–65. Бибкод : 1998AgEE...67...55B . дои : 10.1016/S0167-8809(97)00094-7 .
- Бюкинг Х.; Шачар-Хилл Ю. (2005). «Поглощение, транспорт и перенос фосфатов арбускулярными микоризными грибами увеличиваются при наличии углеводов» . Новый фитолог . 165 (3): 889–912. дои : 10.1111/j.1469-8137.2004.01274.x . ПМИД 15720701 .
- Джордж Э., К. Хаусслер, С.К. Котари, С.Л. Ли и Х. Маршнер, 1992. Вклад микоризных гиф в поглощение питательных веществ и воды растениями. В книге «Микоризы в экосистемах», под ред. DJ Рид, Д.Х. Льюис, А.Х. Фиттер, И.Дж. Александер. Соединенное Королевство: CAB International, стр. 42–47.
- Грант, К.; Битман, С.; Монреаль, М.; Пленчетт, К.; Морель, К. (2005). «Фосфор в почве и удобрениях: влияние на питание растений и развитие микоризы» . Канадский журнал науки о растениях . 85 : 3–14. дои : 10.4141/P03-182 .
- Косута, С.; Шабо, М.; Луньон, Г.; Гоф, К.; Денари, Дж.; Баркер, Д.; Бакар, Г. (2003). «Диффузионный фактор арбускулярных микоризных грибов индуцирует симбиоз-специфическую экспрессию MtENOD11 в корнях Medicago truncatula» . Физиология растений . 131 (3): 952–962. дои : 10.1104/стр.011882 . ПМК 166861 . ПМИД 12644648 .
- Кабир, З.; РТ Койде (2000). «Влияние одуванчика или покровной культуры на потенциал инокуляции микоризы, агрегацию почвы и урожайность кукурузы». Сельское хозяйство, экосистемы и окружающая среда . 78 (2): 167–174. Бибкод : 2000AgEE...78..167K . дои : 10.1016/S0167-8809(99)00121-8 .
- МакГонигл, Т.П.; М. Х. Миллер (1999). «Зимняя выживаемость экстрарадикальных гиф и спор арбускулярных микоризных грибов в полевых условиях». Прикладная экология почв . 12 (1): 41–50. Бибкод : 1999AppSE..12...41M . дои : 10.1016/S0929-1393(98)00165-6 .
- Миллер, Миннесота; Макгонигл Т.П.; Адди, HD (1995). «Функциональная экология везикулярных арбускулярных микориз под влиянием внесения фосфатных удобрений и обработки почвы в сельскохозяйственной экосистеме». Критические обзоры по биотехнологии . 15 (3–4): 241–255. дои : 10.3109/07388559509147411 .
- Мозафар, А.; Анкен, Т.; Рух, Р.; Фроссар, Э. (2000). «Интенсивность обработки почвы, микоризные и немикоризные грибы и концентрация питательных веществ в кукурузе, пшенице и рапсе». Агрономический журнал . 92 (6): 1117–1124. Бибкод : 2000AgrJ...92.1117M . дои : 10.2134/agronj2000.9261117x .
- Соренсен, Дж. Н.; Дж. Ларсен; И. Якобсен (2005). «Образование микоризы и концентрация питательных веществ в луке-порей ( Allium porrum ) в зависимости от предыдущей культуры и управления покровными культурами на почвах с высоким содержанием фосфора». Растение и почва . 273 (1–2): 101–114. Бибкод : 2005ПлСой.273..101С . дои : 10.1007/s11104-004-6960-8 . S2CID 30777851 .
- Вингструп, И.; Г. Рубаек; Э. Сиббенсен; И. Якобсен (1999). «Рост льна ( Linum usitatissimum L.) зависит от арбускулярных микоризных грибов и поглощения фосфора при промежуточных, но не высоких уровнях фосфора в почве в поле». Растение и почва . 203 : 37–46. дои : 10.1023/A:1004362310788 . S2CID 27345855 .
- Тиммер, Л.; Лейден, Р. (1980). «Связь микоризной инфекции с дефицитом меди, вызванным фосфором, в проростках кислого апельсина» . Новый фитолог . 85 : 15–23. дои : 10.1111/j.1469-8137.1980.tb04443.x . S2CID 85946706 .
- Се, З.; Штелин, К.; Фиерхейлиг, Х.; Веймкен, А.; Джаббури, С.; Бротон В.; Вогели-Ланге, Р.; Томас Б. (1995). «Ризобиальные факторы клубенька стимулируют микоризную колонизацию клубеньковых и неклубеньковых соевых бобов» . Физиология растений . 108 (4): 1519–1525. дои : 10.1104/стр.108.4.1519 . ПМК 157531 . ПМИД 12228558 .