Круговая РНК
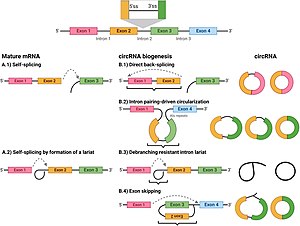
Б.1) прямая обратная склейка;
B.2) интронов ; Циркулизация, вызванная спариванием
Б.3) устойчивый к деветвлению лариат интрона ;
Б.4) циркуляризация, управляемая лариатом ( пропуск экзона ).
В молекулярной биологии кольцевая РНК (или циркРНК ) представляет собой тип одноцепочечной РНК , которая, в отличие от линейной РНК, образует ковалентно замкнутую непрерывную петлю. В кольцевой РНК 3'- и 5'-концы, обычно присутствующие в молекуле РНК, соединены вместе. Эта особенность придает кольцевой РНК многочисленные свойства, многие из которых были идентифицированы лишь недавно.
Многие типы кольцевых РНК возникают из генов , кодирующих белки . Было показано, что некоторые кольцевые РНК кодируют белки . [1] [2] Некоторые типы кольцевых РНК недавно показали потенциал в качестве регуляторов генов . Биологическая функция большинства кольцевых РНК неясна.
Поскольку кольцевая РНК не имеет 5'- или 3'-концов, они устойчивы к экзонуклеазой опосредованной деградации, , и предположительно более стабильны, чем большинство линейных РНК в клетках. [3] Циркулярная РНК связана с некоторыми заболеваниями, такими как рак . [4]
Сплайсинг РНК
[ редактировать ]В отличие от генов бактерий , гены эукариот разделены некодирующими последовательностями, называемыми интронами . У эукариот, когда ген транскрибируется с ДНК в транскрипт информационной РНК (мРНК), промежуточные интроны удаляются, в результате чего в зрелой мРНК остаются только экзоны , которые впоследствии могут транслироваться с образованием белкового продукта. [5] Сплайсосома , [5] Комплекс белок-РНК, расположенный в ядре, катализирует сплайсинг следующим образом:
- Сплайсосома . распознает интрон , который фланкирован специфическими последовательностями на его 5'- и 3'-концах, известных как донорный сайт сплайсинга (или 5'-сайт сплайсинга) и акцепторный сайт сплайсинга (или 3'-сайт сплайсинга) соответственно
- Последовательность 5'-сайта сплайсинга затем подвергается нуклеофильной атаке со стороны нижестоящей последовательности, называемой точкой ветвления, в результате чего образуется кольцевая структура, называемая лариатом.
- Затем свободный 5'-экзон атакует 3'-сайт сплайсинга, соединяя два экзона и высвобождая структуру, известную как интронный лариат . Впоследствии лариат интрона разветвляется и быстро деградирует. [5]

Альтернативный сплайсинг
[ редактировать ]Альтернативный сплайсинг — это явление, посредством которого один транскрипт РНК может давать разные белковые продукты в зависимости от того, какие сегменты считаются «интронами» и «экзонами» во время сплайсинга. [5] Хотя это и не специфично для человека, это частично объясняет тот факт, что люди и другие гораздо более простые виды (например, нематоды) имеют одинаковое количество генов (в диапазоне 20–25 тысяч). [6] Одним из наиболее ярких примеров альтернативного сплайсинга является дрозофилы ген DSCAM , который может дать начало примерно 30 тысячам различных альтернативно сплайсированных изоформ. [7]
Неканонический сплайсинг
[ редактировать ]Скремблирование экзонов
[ редактировать ]Скремблирование экзонов, также называемое перетасовкой экзонов, описывает событие, при котором экзоны соединяются в «неканоническом» (атипичном) порядке. Существует три способа скремблирования экзонов:
- Тандемное дублирование экзонов в геноме, которое часто встречается при раке.
- Транс-сплайсинг , при котором два транскрипта РНК сливаются, в результате чего образуется линейный транскрипт, содержащий экзоны, которые, например, могут происходить от генов, кодируемых на двух разных хромосомах. Транссплайсинг очень распространен у C. elegans.
- Донорный сайт сплайсинга присоединяется к акцепторному сайту сплайсинга, расположенному выше первичного транскрипта, образуя кольцевой транскрипт. [8]
Представление о том, что циклические транскрипты являются побочными продуктами несовершенного сплайсинга, подтверждается низкой распространенностью и отсутствием консервативности последовательности большинства циркРНК. [9] но был оспорен. [8] [10] [3]
Элементы Alu влияют на сплайсинг циркРНК
[ редактировать ]Повторяющиеся последовательности Alu составляют примерно 10% генома человека. [11] Присутствие элементов Alu во фланкирующих интронах белок-кодирующих генов, соседних с первым и последним экзонами, образующими циркРНК, влияет на образование циркРНК. [12] [13] [3] [14] Важно, чтобы фланкирующие интронные элементы Alu были комплементарными, поскольку это обеспечивает спаривание РНК, что, в свою очередь, облегчает синтез циркРНК. [15]
Влияние редактирования РНК на образование циркРНК
[ редактировать ]РНК могут подвергаться модификации оснований путем редактирования РНК после транскрипции. Редактирование РНК происходит главным образом в Alu-элементах генов, кодирующих белки. [16] Редактирование РНК A-to-I в верхних и нижних интронных элементах Alu, фланкирующих сайт обратного сплайсинга (BSS), может уменьшить образование циркРНК в сердце человека. [16] В поврежденном человеческом сердце преимущественное снижение редактирования РНК A-to-I приводит к повышенному образованию циркРНК, что, по-видимому, опосредовано лучшим комплементарным спариванием РНК элементов Alu, фланкирующих сайт обратного сплайсинга. [16]
Характеристики кольцевой РНК
[ редактировать ]Ранние открытия циркРНК
[ редактировать ]Ранние открытия кольцевых РНК привели к убеждению, что они не имеют значения из-за своей редкости. Эти ранние открытия включали анализ таких генов, как гены DCC и Sry , а также недавнее открытие человеческой некодирующей РНК ANRIL , каждая из которых экспрессирует кольцевые изоформы. человека Также были обнаружены гены, продуцирующие CircRNA, такие как ген ETS-1 , гены цитохрома P450 человека и крысы , ген крысиного андрогенсвязывающего белка ( Shbg ) и ген дистрофина человека. [17]
Полногеномная идентификация циркРНК
[ редактировать ]Скремблированные изоформы и циркРНК
[ редактировать ]В 2012 году в попытке первоначально идентифицировать специфичные для рака события скремблирования экзонов, скремблированные экзоны были обнаружены в больших количествах как в нормальных, так и в раковых клетках. Было обнаружено, что скремблированные изоформы экзонов составляют около 10% от общего количества изоформ транскриптов в лейкоцитах , при этом HeLa и H9 2748 скремблированных изоформ в эмбриональных стволовых клетках идентифицировано . Кроме того, примерно 1 из 50 экспрессируемых генов продуцирует изоформы скремблированного транскрипта по крайней мере в 10% случаев. Тесты, используемые для распознавания цикличности, включали обработку образцов РНКазой R , ферментом, который разрушает линейные, но не кольцевые РНК, и тестирование на наличие поли-А-хвостов , которых нет в кольцевых молекулах. В целом было обнаружено, что 98% скремблированных изоформ представляют собой circRNA, circRNA локализованы в цитоплазме, а circRNA обнаружены в большом количестве. [17] [8]
Открытие более высокого содержания циркРНК
[ редактировать ]В 2013 году было обнаружено более высокое содержание циркРНК. человека РНК фибробластов обрабатывали РНКазой R для обогащения кольцевыми РНК с последующей категоризацией кольцевых транскриптов на основе их численности (низкая, средняя, высокая). [3] Было обнаружено, что примерно 1 из 8 экспрессируемых генов продуцирует обнаруживаемые уровни циркРНК, в том числе с низкой распространенностью, которая была значительно выше, чем предполагалось ранее, и объяснялась большей глубиной секвенирования . [3] [8]
Тканевая специфичность и антагонистическая активность CircRNAs
[ редактировать ]В то же время был разработан вычислительный метод обнаружения циркРНК, который привел к обнаружению циклРНК de novo у людей, мышей и C. elegans , а также к их тщательной проверке. Часто обнаруживалось, что экспрессия циркРНК зависит от ткани/стадии развития. Кроме того, было обнаружено, что циркулярные РНК обладают способностью действовать как антагонисты микроРНК, микроРНК, которые мешают трансляции мРНК, примером чего является циркулярная РНК CDR1as , которая имеет сайты связывания микроРНК (как показано ниже). [18]
Данные CircRNA и ENCODE Ribozero RNA-seq
[ редактировать ]В 2014 году человеческие циркРНК были идентифицированы и количественно оценены на основе данных ENCODE Ribozero RNA-seq . Было обнаружено, что большинство циркулярных РНК представляют собой минорные изоформы сплайсинга и экспрессируются только в нескольких типах клеток, при этом 7112 человеческих циркулярных РНК имеют кольцевые фракции (доля сходства изоформы с транскриптами одного и того же локуса) по меньшей мере 10%. Также было обнаружено, что CircRNA не более консервативны, чем их линейный контроль, и, согласно профилированию рибосом, не транслируются. [9] Как отмечалось ранее, циркРНК обладают способностью действовать как антагонисты микроРНК, что также известно как способность действовать как губки микроРНК. Помимо CDR1as, очень немногие циркРНК могут действовать как губки микроРНК. В целом было обнаружено, что большинство кольцевых РНК являются несущественными побочными продуктами несовершенного сплайсинга. [18] [9]
CircRNA и CIRCexplorer
[ редактировать ]В том же году был разработан CIRCexplorer, инструмент, используемый для идентификации тысяч circRNA у людей без секвенирования РНКазы данных R. Было обнаружено, что подавляющее большинство идентифицированных высокоэкспрессируемых экзонных кольцевых РНК процессируются из экзонов, расположенных в середине генов RefSeq , что позволяет предположить, что образование кольцевой РНК обычно связано со сплайсингом РНК. Установлено, что большинство кольцевых РНК содержат несколько, чаще всего два-три экзона. Было обнаружено, что экзоны из циркРНК только с одним циркулярным экзоном намного длиннее, чем экзоны из циркулярных экзонов, что указывает на то, что процессинг может предпочитать определенную длину для максимизации циркуляризации экзона(ов). Интроны кольцевых экзонов обычно содержат высокие плотности Alu , которые могут образовывать инвертированные повторяющиеся пары Alu (IRAlus). IRAlus, как конвергентные, так и дивергентные, располагаются параллельно фланкирующим интронам циркРНК на одинаковых расстояниях до соседних экзонов. Было также обнаружено, что IRAlus и другие неповторяющиеся, но дополняющие друг друга последовательности способствуют образованию кольцевой РНК. С другой стороны, было установлено, что на эффективность циркуляризации экзонов влияет конкуренция спаривания РНК, так что альтернативное спаривание РНК и ее конкуренция приводят к альтернативной циркуляризации. Наконец, было обнаружено, что как циркуляризация экзонов, так и ее регуляция являются эволюционно динамичными. [15]
Полногеномное обращение к circRNA в случаях болезни Альцгеймера
[ редактировать ]Лаборатория Кручаги провела первый крупномасштабный анализ циркРНК при болезни Альцгеймера (БА) и продемонстрировала роль циркРНК в здоровье и заболевании. В нескольких наборах данных было обнаружено, что в общей сложности 148 циклРНК значимо связаны со статусом болезни Альцгеймера и рейтингом клинической деменции (CDR) на момент смерти после коррекции частоты ложных открытий (FDR). Экспрессия circRNA не зависела от линейной формы, и эта экспрессия circRNA также корректировалась в зависимости от пропорции клеток. Также было обнаружено, что CircRNAs совместно экспрессируются с известными генами, вызывающими болезнь Альцгеймера, такими как APP и PSEN1 , что указывает на то, что некоторые circRNA также являются частью причинного пути. В целом было обнаружено, что экспрессия circRNA в мозге объясняет больше клинических проявлений болезни Альцгеймера, чем количество аллелей APOε4, что позволяет предположить, что circRNA могут быть использованы в качестве потенциального биомаркера болезни Альцгеймера. [19]
Классы CircRNA
[ редактировать ]
Кольцевые РНК можно разделить на пять классов: [20] [21]
| Классы кольцевых РНК | Описание |
| Вироиды и вирус гепатита дельта (HDV) | У вироидов и HDV одноцепочечные циклРНК жизненно важны для репликации РНК. Циркулярность позволяет одному событию инициации привести к созданию нескольких геномных копий в процессе, также известном как репликация РНК по вращающемуся кругу . [22] [23] [24] |
| ЦирРНК из интронов | Кольцевые молекулы образуются интронами, образующимися в результате сплайсингосомного сплайсинга, сплайсинга тРНК, а также интронов группы I и группы II (самосплайсинг рибозимов). Интроны группы I образуют циркРНК посредством автокаталитического рибозимального действия, и хотя их можно обнаружить in vivo, их функция еще предстоит определить. [22] [23] [24] Интроны группы II также генерируют циркРНК in vivo . Круглые интроны, образующиеся в результате сплайсосомного сплайсинга эукариот, представляют собой кольцевые интронные лариаты, известные как кольцевые интронные РНК (циРНК). Благодаря циркуляризации циРНК могут избежать деградации и, как полагают, в значительной степени представлены. Функция CiRNA в настоящее время неизвестна; однако предполагается, что они могут играть роль в усилении транскрипции генов, из которых они производятся, поскольку они взаимодействуют с РНК-полимеразой II. [21] |
| ЦирРНК из промежуточных продуктов реакций процессинга РНК | Сначала они соединяются из предшественников в виде линейных молекул, а затем циркуляризируются с помощью лигазы. Они необходимы для обеспечения перестройки порядка последовательностей РНК и жизненно важны для биогенеза пермутированных генов тРНК у некоторых водорослей и архей. [21] |
| Некодирующие циркРНК у архей | У некоторых видов архей есть циркРНК, которые образуются из вырезанных кольцевых интронов тРНК. Считается, что циркуляризация функциональных некодирующих РНК действует как защитный механизм против экзонуклеаз и способствует правильному сворачиванию. [21] [3] |
| ЦирРНК у эукариот, полученные путем обратного сплайсинга | Кольцевые РНК, продуцируемые обратным сплайсингом (форма скремблирования экзонов), возникают, когда 5'-сайт сплайсинга соединяется с расположенным выше 3'-сайтом сплайсинга. В настоящее время у человека идентифицировано более 25 000 различных циркРНК. [21] [3] |
Длина циркРНК
[ редактировать ]Недавнее исследование человеческих циркРНК показало, что эти молекулы обычно состоят из 1–5 экзонов. [18] Каждый из этих экзонов может быть в три раза длиннее среднего экспрессируемого экзона. [3] предполагая, что длина экзона может играть роль в принятии решения о том, какие экзоны циркулировать. 85% циркулярных экзонов перекрываются с экзонами, кодирующими белок . [18] хотя сами кольцевые РНК, по-видимому, не транслируются. Во время образования циркРНК экзон 2 часто является вышестоящим «акцепторным» экзоном. [8]
Интроны, окружающие экзоны, выбранные для циркуляризации, в среднем в три раза длиннее, чем те, которые не фланкируют экзоны перед кольцом. [8] [3] хотя пока не ясно, почему это так. По сравнению с областями, не имеющими кружков, эти интроны с гораздо большей вероятностью содержат комплементарные инвертированные повторы Alu , причем Alu является наиболее распространенным транспозоном в геноме. [3] Поскольку Alu повторяет спаривание оснований друг с другом, было высказано предположение, что это может позволить сайтам сплайсинга находить друг друга, тем самым способствуя циркуляризации. [10] [3]
Интроны внутри циркРНК сохраняются с относительно высокой частотой (~ 25%), [9] таким образом добавляя дополнительную последовательность к зрелым циркРНК.
Расположение циркРНК в клетке
[ редактировать ]В клетке циркРНК преимущественно обнаруживаются в цитоплазме , где количество транскриптов кольцевой РНК, полученных из гена, может быть до десяти раз больше, чем количество связанных линейных РНК, генерируемых из этого локуса . Неясно, как кольцевые РНК выходят из ядра через относительно небольшую ядерную пору . Поскольку ядерная оболочка разрушается во время митоза , одна из гипотез состоит в том, что молекулы покидают ядро во время этой фазы клеточного цикла . [3] Однако некоторые циркРНК, такие как CiRS-7/CDR1as, экспрессируются в тканях нейронов. [18] [25] где митотическое деление не преобладает.

ЦирРНК более стабильны, чем линейные РНК.
[ редактировать ]У CircRNA отсутствует полиаденилированный хвост, и, следовательно, предполагается, что они менее склонны к деградации экзонуклеазами. В 2015 году Энука и др. измерили периоды полураспада 60 циркулярных РНК и их линейных аналогов, экспрессируемых из одного и того же гена-хозяина, и обнаружили, что средний период полураспада циркулярных РНК клеток молочной железы (от 18,8 до 23,7 часов) по крайней мере в 2,5 раза превышает средний период полураспада их линейные аналоги (от 4,0 до 7,4 часа). [26] Обычно время жизни молекул РНК определяет время их реакции. [27] Соответственно, сообщалось, что циркРНК молочных желез медленно реагируют на стимуляцию факторами роста. [26]
Возможные функции кольцевой РНК
[ редактировать ]Эволюционное сохранение механизмов и сигналов циркуляризации
[ редактировать ]ЦирРНК были идентифицированы у различных видов в различных сферах жизни . В 2011 году Данан и др. секвенировали РНК архей . После расщепления общей РНК РНКазой R они смогли идентифицировать кольцевые виды, что указывает на то, что циркРНК не специфичны для эукариот. [28] Однако эти архейные круглые виды, вероятно, не образуются посредством сплайсинга, что позволяет предположить, что, вероятно, существуют другие механизмы генерации кольцевой РНК.
Было обнаружено, что CircRNA в значительной степени консервативны у человека и овцы. Анализируя данные тотального секвенирования РНК из коры теменных долей овцы и мононуклеарных клеток периферической крови, было показано, что 63% обнаруженных циркРНК гомологичны известным циркРНК человека. [29]
В более тесной эволюционной связи сравнение РНК из семенников мыши с РНК из клетки человека выявило 69 ортологичных циркРНК. Например, и люди, и мыши кодируют гены HIPK2 и HIPK3 , две паралогичные киназы, которые продуцируют большое количество циркРНК из одного конкретного экзона у обоих видов. [3] Эволюционная консервация усиливает вероятность значимой и важной роли циркуляризации РНК.
CDR1as/CiRS-7 как губка миР-7
[ редактировать ]МикроРНК (миРНК) представляют собой небольшие (около 21 нуклеотида) некодирующие РНК, которые подавляют трансляцию информационных РНК, участвующих в большом и разнообразном наборе биологических процессов. [30] Они напрямую соединяют пары оснований с целевыми информационными РНК (мРНК) и могут запускать расщепление мРНК в зависимости от степени комплементарности.
МикроРНК сгруппированы в «исходные семейства». Члены семьи имеют общие нуклеотиды 2–7, известные как семенная область. [31] Белки Argonaute являются «эффекторными белками», которые помогают микроРНК выполнять свою работу, в то время как губки микроРНК представляют собой РНК, которые «объедают» микроРНК определенного семейства, тем самым служа конкурентными ингибиторами , подавляющими способность микроРНК связывать свои мишени мРНК. благодаря наличию нескольких сайтов связывания, которые распознают конкретную зародышевую область. [31] Определенные кольцевые РНК имеют множество сайтов связывания микроРНК, что позволяет предположить, что они могут функционировать при губке. Две недавние статьи подтвердили эту гипотезу, подробно исследовав кольцевую губку под названием CDR1as/CiRS-7, в то время как другие группы не нашли прямых доказательств того, что кольцевые РНК действуют как губки микроРНК, путем анализа потенциального взаимодействия кольцевых РНК с белком Argonaut (AGO) с использованием данные высокопроизводительного секвенирования РНК, выделенной с помощью перекрестного связывания и иммунопреципитации (HITS-CLIP). [32]
CDR1as/CiRS-7 кодируется в геноме, антисмысловом человека по отношению к локусу (гену) CDR1 (отсюда и название CDR1as), [18] и нацелен на миР-7 (отсюда и название CiRS-7 — кольцевая РНК-губка для миР-7 ). [25] Он имеет более 60 сайтов связывания миР-7, что намного больше, чем у любой известной губки с линейной миРНК. [18] [25]
AGO2 представляет собой белок Argonaute, ассоциированный с миР-7 (см. выше). Хотя CDR1as/CiRS-7 могут расщепляться миР-671 и связанным с ней белком Argonaute, [25] он не может быть расщеплен миР-7 и AGO2. Активность расщепления микроРНК зависит от комплементарности за пределами 12-го положения нуклеотида; ни один из сайтов связывания CiRS-7 не отвечает этому требованию.
Эксперимент с рыбками данио , в геноме которых нет локуса CDR1, доказывает активность губок CiRS-7. Во время развития миР-7 сильно экспрессируется в мозге рыбок данио. Чтобы заглушить экспрессию miR-7 у рыбок данио, Мемчак и его коллеги воспользовались инструментом под названием морфолино , который может связывать пары оснований и изолировать целевые молекулы. [33] Лечение морфолино имело такой же серьезный эффект на развитие среднего мозга, как и эктопическая экспрессия CiRS-7 в мозге рыбок данио с использованием инъецированных плазмид . Это указывает на значительное взаимодействие между CiRS-7 и миР-7 in vivo. [18]
Другая известная круглая губка микроРНК — SRY . SRY, который высоко экспрессируется в семенниках мышей, действует как губка миР-138 . [25] [34] В геноме SRY окружен длинными инвертированными повторами (IR) длиной более 15,5 тысяч оснований (т.п.н.). При удалении одного или обоих IR циклизация не происходит. Именно это открытие привело к появлению идеи инвертированных повторов, обеспечивающих циркуляризацию. [35]
Поскольку губки с кольцевой РНК характеризуются высоким уровнем экспрессии, стабильностью и большим количеством сайтов связывания микроРНК, они, вероятно, являются более эффективными губками, чем линейные. [10]
Другие возможные функции циркРНК
[ редактировать ]Хотя в последнее время внимание было сосредоточено на «губчатых» функциях circRNA, ученые рассматривают также несколько других функциональных возможностей. Например, в некоторых областях гиппокампа взрослых мышей наблюдается экспрессия CiRS-7, но не miR-7, что указывает на то, что CiRS-7 может выполнять роли, независимые от взаимодействия с miRNA. [18]
Потенциальные роли включают в себя следующее:
- Связывание с РНК-связывающими белками (RBP) и РНК, помимо микроРНК, с образованием комплексов РНК-белок. [10] Эти комплексы могут регулировать взаимодействия RBP и РНК, например, с каноническим линейным транскриптом гена или вирусной инфекцией. [8] [36]
- Производство белка
- Chen и Sarnow 1995 показали, что синтетическая циркРНК, содержащая IRES (внутренний сайт входа в рибосому), производит белковый продукт in vitro , тогда как без IRES этого не происходит. Хотя протестированная циркРНК была чисто искусственной конструкцией, Чен и Сарноу заявили в своей статье, что им было бы интересно посмотреть, содержат ли круги естественным образом элементы IRES. [37]
- Джек и др. 2013: Протестированы природные циркРНК, содержащие «старт-кодон» трансляции. Однако ни одна из этих молекул не связалась с рибосомами, что позволяет предположить, что многие циркРНК не могут транслироваться in vivo . [3]
- Транспорт микроРНК внутри клетки. Тот факт, что CiRS-7 может быть разрезана миР-671, может указывать на существование системы, позволяющей высвобождать «нагрузку» миРНК в подходящее время. [38]
- Регулирование мРНК в клетке посредством ограниченного спаривания оснований. Формально возможно, что миР-7 смягчает регуляторную активность CiRS-7, а не наоборот! [18] [38]
Циркулярные интронные длинные некодирующие РНК (циРНК)
[ редактировать ]Обычно интронные лариаты (см. выше) разветвлены и быстро деградируют. Однако нарушение разветвления может привести к образованию кольцевых интронных длинных некодирующих РНК, также известных как циРНК. [39] Образование CiRNA, по-видимому, зависит не от случайного процесса, а от присутствия специфических элементов вблизи 5'-сайта сплайсинга и сайта точки ветвления (см. выше).
ЦиРНК отличаются от циркРНК тем, что они преимущественно обнаруживаются в ядре, а не в цитоплазме . Кроме того, эти молекулы содержат мало (если таковые имеются) сайтов связывания микроРНК. Вместо того, чтобы действовать как губки, циРНК, по-видимому, регулируют экспрессию своих родительских генов. Например, относительно распространенная циРНК, называемая ci-ankrd52, положительно регулирует транскрипцию Pol II . Многие циРНК остаются в своих «местах синтеза» в ядре. Однако циРНК могут выполнять и другие функции, помимо простого регулирования своих родительских генов, поскольку циРНК действительно локализуются в дополнительных участках ядра, отличных от «мест их синтеза». [39]
Циркулярная РНК и болезни
[ редактировать ]Как и в большинстве тем молекулярной биологии , важно учитывать, как кольцевую РНК можно использовать в качестве инструмента помощи человечеству. Учитывая ее распространенность, эволюционную консервативность и потенциальную регуляторную роль, стоит изучить, как кольцевую РНК можно использовать для изучения патогенеза и разработки терапевтических вмешательств. Например:
- Циркулярный ANRIL (cANRIL) представляет собой кольцевую форму ANRIL, длинной некодирующей РНК (нкРНК). Экспрессия канрила коррелирует с риском развития атеросклероза — заболевания, при котором артерии становятся твердыми. Было высказано предположение, что канрил может изменять экспрессию INK4 /ARF, что, в свою очередь, увеличивает риск атеросклероза. [40] Дальнейшее изучение экспрессии cANRIL потенциально может быть использовано для профилактики или лечения атеросклероза.
- миР-7 играет важную регуляторную роль при некоторых видах рака и болезни Паркинсона , заболевании головного мозга неизвестного происхождения. [25] Возможно, активность губки CiRS-7 могла бы помочь в противодействии активности миР-7. Если активность кольцевой губки действительно может помочь в противодействии вредной активности микроРНК, ученым необходимо будет найти лучший способ обеспечить экспрессию губки, возможно, с помощью трансгена , который представляет собой синтетический ген, который передается между организмами. Также важно учитывать, что трансгены могут экспрессироваться только в определенных тканях или экспрессироваться только при индуцировании. [31]
- Было обнаружено, что кольцевые РНК регулируются гипоксией, особенно было обнаружено, что циркРНК cZNF292 обладает проангиогенной активностью в эндотелиальных клетках. [32]
- Было обнаружено, что кольцевая РНК помогает ВИЧ-1 уклоняться от иммунной защиты организма. Белок ВИЧ, известный как Vpr, индуцирует выработку кольцевой молекулы РНК, называемой ciTRAN. Впоследствии было обнаружено связывание ciTRAN со специфическим белком под названием SRSF1, тем самым эффективно нарушая способность SRSF1 ингибировать транскрипцию ВИЧ-1. [41]
Циркулярные РНК играют роль в патогенезе болезни Альцгеймера
[ редактировать ]Дубе и др ., [19] впервые продемонстрировали, что кольцевые РНК головного мозга (циркулярные РНК) являются частью патогенных событий, которые приводят к болезни Альцгеймера , выдвинув гипотезу о том, что специфическая циркРНК будет дифференциально экспрессироваться в случаях AD по сравнению с контрольной группой, и что эти эффекты могут быть обнаружены на ранних стадиях заболевания. Они оптимизировали и утвердили новый конвейер анализа кольцевых РНК (circRNA). Они провели трехэтапное исследование, используя данные секвенирования РНК мозга Knight ADRC в качестве открытия (этап 1), используя данные с горы Синай в качестве репликации (этап 2) и метаанализа (этап 3) для выявления наиболее значительная циркРНК дифференциально экспрессируется при болезни Альцгеймера. Используя его конвейер, они обнаружили 3547 circRNA, прошедших строгий контроль качества в когорте Knight ADRC, которая включает секвенирование РНК из 13 контрольных групп и 83 случаев болезни Альцгеймера, а также 3924 circRNA, прошедших строгий контроль качества в наборе данных MSBB. Метаанализ результатов открытия и репликации выявил в общей сложности 148 циркРНК, которые значительно коррелировали с CDR после коррекции FDR. Кроме того, 33 циркРНК прошли строгий основанный на генах множественный тест Бонферрони с коррекцией 5×10-6, включая circHOMER1 (P =2,21×10). −18 ) и circCDR1-AS (P = 2,83 × 10 −8 ), среди других. Они также провели дополнительные анализы, чтобы продемонстрировать, что экспрессия циркРНК не зависит от линейной формы, а также от доли клеток, что может исказить анализ секвенирования РНК головного мозга в исследованиях болезни Альцгеймера. Они провели анализ совместной экспрессии всех circRNA вместе с линейными формами и обнаружили, что circRNA, включая те, которые дифференциально экспрессируются при болезни Альцгеймера по сравнению с контролем, коэкспрессируются совместно с известными генами, вызывающими болезнь Альцгеймера, такими как APP и PSEN1, что указывает на то, что некоторые циркРНК также являются частью причинного пути. Они также продемонстрировали, что экспрессия circRNA в мозге объясняет больше клинических проявлений болезни Альцгеймера, чем количество аллелей APOε4, что позволяет предположить, что их можно использовать в качестве потенциального биомаркера болезни Альцгеймера. Это важное исследование в этой области, поскольку впервые циркРНК количественно определяют и подтверждают (с помощью ПЦР в реальном времени) в образцах человеческого мозга в полногеномном масштабе и в больших и хорошо охарактеризованных когортах. Это также демонстрирует, что эти формы РНК, вероятно, связаны с сложными признаками, включая болезнь Альцгеймера, и поможет понять биологические события, которые приводят к заболеванию.
Патогенез циркулярной РНК при заболеваниях сердца, почек, печени, аутоиммунных заболеваниях и развитии рака
[ редактировать ]Сердечное заболевание
[ редактировать ]Недавние исследования показали, что циркРНК связана с сердечной недостаточностью и заболеваниями сердца. circFOXO3, гены тайтина, circSLC8A1-1 и circAmotl1 играют важную роль в сердечной функции посредством повышения или ингибирования, связанного с сердечными заболеваниями. Сверхэкспрессия circFOXO3 и его подавление связываются с факторами транскрипции E2F1, HIF1α и белком ID1, FAK, вызывая кардиомиопатию, индуцированную DOX. ЦирРНК, производная гена титина, индуцирует кардиотоксичность в кардиомиоцитах . Сверхэкспрессия circSLC8A1-1 вызывает размывание регулятора сердечной гипертрофии миР-133 и приводит к сердечной недостаточности. Помимо заболеваний сердца, опосредованных circRNA, некоторые circRNA играют роль в восстановлении повреждений сердца. Например, сверхэкспрессия circAmotl1 увеличивает продолжительность жизни кардиомиоцитов за счет связывания и транслокации AKT, который регулирует восстановление сердца. Циркулярная РНК CDR1 играет важную роль при инфекции миокарда . Сердечная дисфункция возникает после инфекции миокарда из-за снижения регуляции CircNfix. Поскольку различные типы кольцевой РНК связаны с сердечно-сосудистыми заболеваниями, ее можно использовать в качестве потенциального биомаркер и терапевтическая мишень. Например, послеоперационная фибрилляция предсердий наблюдалась у некоторых пациентов после операции на сердце, где в качестве биомаркера использовалась circRNA_025016. Хотя значимость сверхэкспрессии и подавления кольцевой РНК для заболеваний сердца была обнаружена в различных исследованиях, она до сих пор неясна. Таким образом, необходимы дальнейшие исследования для отслеживания прогрессирования заболевания на различных стадиях сердечной дисфункции с использованием кольцевой РНК в качестве биомаркера, которую можно использовать для доставки генов в клетки. [42]
Заболевания почек
[ редактировать ]Различные исследования показали, что кольцевая РНК действует как прогностический агент и биомаркер при заболеваниях почек, включая почечно-клеточный рак , острое повреждение почек, диабетическую нефропатию и волчаночный нефрит . Хроническое заболевание почек связано с миР-150, которая отрицательно регулируется circHLA-C у пациентов с волчаночным нефритом. Имеются также доказательства того, что кольцевая РНК участвует в остром повреждении почек. В этих обстоятельствах кольцевая РНК оказывается новым биомаркером, а также используется для таргетной терапии заболеваний почек, поскольку ее псевдоген может изменять состав ДНК. [43]
Заболевания печени
[ редактировать ]Данные показали, что кольцевая РНК играет роль в хронических заболеваниях печени и регуляции гомеостаза , что приводит к фиброзу печени и аутоиммунным заболеваниям по эпигенетическому механизму. [44]
Рак
[ редактировать ]Циркулярная РНК выполняет как положительные, так и отрицательные функции при раке. Например, было обнаружено, что ciRS-7 является онкогеном в ткани колоректального рака, который регулирует заболевание. Сверхэкспрессия этой ciRS-7 приводит к нарушению регуляции экспрессии генов, что приводит к злокачественным фенотипическим особенностям. С другой стороны, некоторые CircRNA демонстрируют положительные эффекты, такие как circ-ITCH, который регулирует рак легких, связанный с онкогенными губками miR7 и miR214, а сверхэкспрессия circ-ITCH ингибирует пролиферацию клеток при раке легких. В результате различных исследований было обнаружено, что F-circM-9, F-circPR и F-circEA, FcircEA-2 участвуют в развитии лейкемии и рака. В клетках остеосаркомы circ-0016347 индуцирует опухоль и снижает регуляцию мишени каспазы-1 . Другая кольцевая РНК hsa_circRNA_002178 приводит к раку молочной железы , когда она сверхэкспрессирует и подавляет функцию белка COL1A1. Напротив, подавление hsa_circRNA_002178 снижало выработку IL-6 и TNF-α, что ингибировало рост опухоли и воспаление. Некоторые вирусы, такие как Вирус Эпштейна-Барра и вирус папилломы человека могут кодировать кольцевые РНК, такие как circEBNA_W1_C1 (EBV) и circE7 (HPV), которые играют роль в онкогенезе у инфицированных людей. Поскольку циркРНК участвуют в процессе развития или регуляции рака, поэтому ее можно использовать в качестве биомаркера в процессе наблюдения и идентификации рака. [45] Циркулярная РНК обладает преимуществом стабильности, тканевой специфичности, ее можно обнаружить в крови, слюне, моче, спинномозговой жидкости, а секреция жидкости организма человека, которая имеет большое количество экзосом, хорошо подходит для использования в качестве биомаркера рака. [44] [45]
Аутоиммунное заболевание
[ редактировать ]Циркулярная РНК играет роль в прогрессировании аутоиммунных заболеваний, действуя как губка микроРНК, которая регулирует метилирование ДНК, адаптивную иммунную активацию и секрецию костимулирующих молекул. [44]
Циркулярная РНК в иммунном ответе
[ редактировать ]Циркулярная РНК играет значительную роль в иммунной регуляции и индукции Т-клеточных ответов. circRNA100783 участвует в иммунитете и старении CD8+ Т-клеток. circRNA-003780 и circRNA-010056 также играют важную роль в дифференцировке и поляризации макрофагов. [45]
Циркулярная РНК действует как очень активный иммунный агент, когда она объединяется с растворимыми белковыми антигенами и индуцирует адаптивный иммунитет, не требующий определенного пути введения. Циркулярная РНК плазмы и комбинированная циркулярная РНК имеют более высокую эффективность в диагностике, чем тканеспецифическое лечение и одиночная кольцевая РНК. Обработка кольцевой РНК активирует дифференцировку и созревание дендритных клеток, которые затем секретируют большое количество различных цитокинов и хемокинов путем экспрессии генов IL-1β, IL-6 и TNFα. После иммунизации кольцевой РНК, кодирующей последовательность антигена, Т-клеточные ответы медиатора CD8+ на целевой антиген усиливаются. Циклическая РНК обладает очень выгодными свойствами стабильности и длительного срока хранения, поэтому ее можно использовать в качестве биомаркеров и плазмид для экспрессии интересующих генов. [46]
Циркулярная РНК в регуляции миогенеза
[ редактировать ]Циркулярная РНК играет важную роль в таких механизмах миогенеза , как circRBFOX2, circLMO7 действует как отрицательный регулятор, а CircSVIL действует как положительный регулятор. [47] circRBFOX2 регулирует экспрессию миР-206 и индуцирует пролиферацию миобластов , оказывая негативное влияние на процесс миогенеза. circLMO7 участвует в сверхэкспрессии HDAC4 и подавляет экспрессию MEF2A за счет повышения регуляции миР-378a-3p, что приводит к дифференцировке миобластов. [47] CircSVIL, положительный регулятор, индуцирует активность миР-203, которая является ингибитором продукции и дифференцировки миобластов. circFUT10 участвует в ингибировании пролиферации миобластов, но усиливает дифференцировку за счет усиления экспрессии SRF. [47] Губки circSNX29 миР-744 и губки circFGFR2 миР-133a-5p и миР-29b-1-5p, которые способствуют дифференцировке миобластов. circSNX29 активирует пути Wnt путем усиления экспрессии Wnt5a и CaMKIId, которые участвуют в регуляции миогенеза. [47]
Вироиды как кольцевые РНК
[ редактировать ]Вироиды — это в основном патогены растений, которые состоят из коротких участков (несколько сотен нуклеиновых оснований) высококомплементарных кольцевых одноцепочечных и некодирующих РНК без белковой оболочки. По сравнению с другими инфекционными возбудителями растений вироиды имеют чрезвычайно малые размеры: от 246 до 467 нуклеиновых оснований; таким образом, они состоят менее чем из 10 000 атомов. Для сравнения: длина генома самых маленьких известных вирусов, способных вызвать инфекцию, составляет около 2000 азотистых оснований. [48]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Давуд А., Ихаб Закария З., Хишам Рашван Х., Браудаки М., Юнесс Р.А. (март 2023 г.). «Циркулярные РНК: новый уровень сложности, позволяющий избежать гетерогенности рака молочной железы» . Исследование некодирующих РНК . 8 (1): 60–74. дои : 10.1016/j.ncrna.2022.09.011 . ПМЦ 9637558 . ПМИД 36380816 . S2CID 252890235 .
- ^ Памудурти Н.Р., Барток О., Йенс М., Ашвал-Флусс Р., Стоттмайстер С., Руэ Л. и др. (апрель 2017 г.). «Трансляция CircRNAs» . Молекулярная клетка . 66 (1): 9–21.e7. doi : 10.1016/j.molcel.2017.02.021 . ПМЦ 5387669 . ПМИД 28344080 .

- ^ Перейти обратно: а б с д и ж г час я дж к л м н Джек В.Р., Соррентино Дж.А., Ван К., Слевин М.К., Берд К.Э., Лю Дж. и др. (февраль 2013 г.). «Кольцевые РНК широко распространены, консервативны и связаны с повторами ALU» . РНК . 19 (2): 141–157. дои : 10.1261/rna.035667.112 . ПМК 3543092 . ПМИД 23249747 .
- ^ Вромман М., Вандесомпель Дж., Волдерс П.Дж. (январь 2021 г.). «Замыкание круга: современное состояние и перспективы баз данных кольцевых РНК» . Брифинги по биоинформатике . 22 (1): 288–297. дои : 10.1093/нагрудник/bbz175 . ПМЦ 7820840 . ПМИД 31998941 .
- ^ Перейти обратно: а б с д Рис Дж.Б. (2010). Кэмпбелл Биология (9-е изд.). Сан-Франциско: Бенджамин Каммингс.
- ^ Ю Дж., Ху С., Ван Дж., Вонг Г.К., Ли С., Лю Б. и др. (апрель 2002 г.). «Проект последовательности генома риса (Oryza sativa L. ssp. indica)». Наука . 296 (5565): 79–92. Бибкод : 2002Sci...296...79Y . дои : 10.1126/science.1068037 . ПМИД 11935017 . S2CID 208529258 .
- ^ Селото А.М., Грейвли Б.Р. (октябрь 2001 г.). «Альтернативный сплайсинг пре-мРНК Dscam дрозофилы регулируется как во времени, так и в пространстве» . Генетика . 159 (2): 599–608. дои : 10.1093/генетика/159.2.599 . ПМЦ 1461822 . ПМИД 11606537 .
- ^ Перейти обратно: а б с д и ж г Зальцман Дж., Гавад С., Ван П.Л., Лакайо Н., Браун П.О. (2012). «Кольцевые РНК являются преобладающей изоформой транскриптов сотен человеческих генов в различных типах клеток» . ПЛОС ОДИН . 7 (2): e30733. Бибкод : 2012PLoSO...730733S . дои : 10.1371/journal.pone.0030733 . ПМК 3270023 . ПМИД 22319583 .
- ^ Перейти обратно: а б с д Го Цзюй, Агарвал В., Го Х, Бартель Д.П. (июль 2014 г.). «Расширенная идентификация и характеристика кольцевых РНК млекопитающих» . Геномная биология . 15 (7): 409. doi : 10.1186/s13059-014-0409-z . ПМК 4165365 . ПМИД 25070500 .
- ^ Перейти обратно: а б с д Вилуш Дж. Э., Sharp PA (апрель 2013 г.). «Молекулярная биология. Окружной путь к некодирующей РНК» . Наука . 340 (6131): 440–441. Бибкод : 2013Sci...340..440W . дои : 10.1126/science.1238522 . ПМК 4063205 . ПМИД 23620042 .
- ^ Дэниел С., Зильберберг Г., Бем М., Оман М. (февраль 2014 г.). «Элементы Alu формируют транскриптом приматов посредством цис-регуляции редактирования РНК» . Геномная биология . 15 (2): С28. дои : 10.1186/gb-2014-15-2-r28 . ПМК 4053975 . ПМИД 24485196 .
- ^ Иванов А., Мемчак С., Уайлер Э., Торти Ф., Порат Х.Т., Орехуэла М.Р. и др. (январь 2015 г.). «Анализ последовательностей интронов выявляет признаки биогенеза кольцевой РНК у животных» . Отчеты по ячейкам . 10 (2): 170–177. дои : 10.1016/j.celrep.2014.12.019 . ПМИД 25558066 .
- ^ Вилуш Дж. Э. (04 мая 2015 г.). «Повторяющиеся элементы регулируют биогенез кольцевой РНК» . Мобильные генетические элементы . 5 (3): 39–45. дои : 10.1080/2159256X.2015.1045682 . ПМЦ 4588227 . ПМИД 26442181 .
- ^ Лян Д., Вилуш Дж.Э. (октябрь 2014 г.). «Короткие интронные повторы облегчают производство кольцевой РНК» . Гены и развитие . 28 (20): 2233–2247. дои : 10.1101/gad.251926.114 . ПМК 4201285 . ПМИД 25281217 .
- ^ Перейти обратно: а б Чжан СО, Ван ХБ, Чжан Ю, Лу Х, Чен ЛЛ, Ян Л (сентябрь 2014 г.). «Комплементарная циркуляризация экзонов, опосредованная последовательностью» . Клетка . 159 (1): 134–147. дои : 10.1016/j.cell.2014.09.001 . ПМИД 25242744 . S2CID 18390400 .
- ^ Перейти обратно: а б с Кокот К.Э., Кнойер Дж.М., Джон Д., Ребс С., Мёбиус-Винклер М.Н., Эрбе С. и др. (июнь 2022 г.). «Уменьшение редактирования РНК A-to-I в поврежденном сердце человека регулирует образование кольцевых РНК» . Фундаментальные исследования в кардиологии . 117 (1): 32. дои : 10.1007/s00395-022-00940-9 . ПМК 9226085 . ПМИД 35737129 .
- ^ Перейти обратно: а б Барретт С.П., Зальцман Дж. (июнь 2016 г.). «Циркулярные РНК: анализ, экспрессия и потенциальные функции» . Разработка . 143 (11): 1838–1847. дои : 10.1242/dev.128074 . ПМЦ 4920157 . ПМИД 27246710 .
- ^ Перейти обратно: а б с д и ж г час я дж Мемчак С., Йенс М., Элефсиниоти А., Торти Ф., Крюгер Дж., Рыбак А. и др. (март 2013 г.). «Циркулярные РНК представляют собой большой класс животных РНК с регуляторной активностью». Природа . 495 (7441): 333–338. Бибкод : 2013Natur.495..333M . дои : 10.1038/nature11928 . ПМИД 23446348 . S2CID 4416605 .
- ^ Перейти обратно: а б Дюбе У., Дель-Агила Дж.Л., Ли З., Бадде Дж.П., Цзян С., Сюй С. и др. (ноябрь 2019 г.). «Атлас экспрессии кортикальной кольцевой РНК в мозге при болезни Альцгеймера демонстрирует клинические и патологические ассоциации» . Природная неврология . 22 (11): 1903–1912. дои : 10.1038/s41593-019-0501-5 . ПМК 6858549 . ПМИД 31591557 .
- ^ Эгер Н., Шоппе Л., Шустер С., Лауфс У., Бекель Й.Н. (2018). «Кольцевой сплайсинг РНК». В Сяо Дж (ред.). Круглые РНК . Достижения экспериментальной медицины и биологии. Том. 1087. Сингапур: Springer Singapore. стр. 41–52. дои : 10.1007/978-981-13-1426-1_4 . ISBN 978-981-13-1425-4 . ПМИД 30259356 .
- ^ Перейти обратно: а б с д и Нисар С., Бхат А.А., Сингх М., Каредат Т., Ризван А., Хашем С. и др. (05.02.2021). «Понимание роли CircRNA: биогенез, характеристика, функциональное и клиническое влияние на злокачественные новообразования человека» . Границы клеточной биологии и биологии развития . 9 : 617281. doi : 10.3389/fcell.2021.617281 . ПМЦ 7894079 . ПМИД 33614648 .
- ^ Перейти обратно: а б Грабовский П.Я., Зауг А.Я., Чех Т.Р. (февраль 1981 г.). «Промежуточная последовательность предшественника рибосомальной РНК превращается в кольцевую РНК в изолированных ядрах тетрахимены». Клетка . 23 (2): 467–476. дои : 10.1016/0092-8674(81)90142-2 . ПМИД 6162571 . S2CID 41905828 .
- ^ Перейти обратно: а б Крюгер К., Грабовски П.Дж., Зауг А.Дж., Сэндс Дж., Готтшлинг Д.Е., Чех Т.Р. (ноябрь 1982 г.). «Самосплайсинг РНК: аутоиссечение и автоциклизация промежуточной последовательности рибосомальной РНК тетрахимены». Клетка . 31 (1): 147–157. дои : 10.1016/0092-8674(82)90414-7 . ПМИД 6297745 . S2CID 14787080 .
- ^ Перейти обратно: а б Зауг А.Я., Грабовский П.Я., Чех Т.Р. (17–23 февраля 1983 г.). «Автокаталитическая циклизация вырезанной промежуточной последовательности РНК представляет собой реакцию расщепления-лигирования». Природа . 301 (5901): 578–583. Бибкод : 1983Natur.301..578Z . дои : 10.1038/301578a0 . ПМИД 6186917 . S2CID 4348876 .
- ^ Перейти обратно: а б с д и ж Хансен Т.Б., Йенсен Т.И., Клаузен Б.Х., Брамсен Дж.Б., Финсен Б., Дамгаард К.К., Кьемс Дж. (март 2013 г.). «Природные кольца РНК действуют как эффективные губки микроРНК». Природа . 495 (7441): 384–388. Бибкод : 2013 Природа.495..384H . дои : 10.1038/nature11993 . ПМИД 23446346 . S2CID 205233122 .
- ^ Перейти обратно: а б Энука Ю., Лауриола М., Фельдман М.Е., Сас-Чен А., Улицкий И., Ярден Ю. (февраль 2016 г.). «Циркулярные РНК долговечны и демонстрируют лишь минимальные ранние изменения в ответ на фактор роста» . Исследования нуклеиновых кислот . 44 (3): 1370–1383. дои : 10.1093/nar/gkv1367 . ПМЦ 4756822 . ПМИД 26657629 .
- ^ Снеппен К. (2014). Модели динамики и регуляции жизни в биологических системах . Сан-Франциско: Издательство Кембриджского университета. ISBN 978-1-107-06190-3 .
- ^ Данан М., Шварц С., Эдельхейт С., Сорек Р. (апрель 2012 г.). «Общетранскриптомное открытие кольцевых РНК у архей» . Исследования нуклеиновых кислот . 40 (7): 3131–3142. дои : 10.1093/nar/gkr1009 . ПМЦ 3326292 . ПМИД 22140119 .
- ^ Варела-Мартинес Э., Корси Дж.И., Антон С., Городкин Дж., Юго Б.М. (январь 2021 г.). «Новое открытие циркРНК у овец свидетельствует о высокой консервативности обратного сращивания» . Научные отчеты . 11 (1): 427. doi : 10.1038/s41598-020-79781-2 . ПМК 7801505 . ПМИД 33432020 .
- ^ Дин XC, Вейлер Дж., Гроссханс Х. (январь 2009 г.). «Регуляция регуляторов: механизмы, контролирующие созревание микроРНК». Тенденции в биотехнологии . 27 (1): 27–36. дои : 10.1016/j.tibtech.2008.09.006 . ПМИД 19012978 .
- ^ Перейти обратно: а б с Эберт М.С., Шарп, Пенсильвания (ноябрь 2010 г.). «Губки микроРНК: прогресс и возможности» . РНК . 16 (11): 2043–2050. дои : 10.1261/rna.2414110 . ПМК 2957044 . ПМИД 20855538 .
- ^ Перейти обратно: а б Бекель Дж.Н., Джаэ Н., Хоймюллер А.В., Чен В., Бун Р.А., Стеллос К. и др. (октябрь 2015 г.). «Идентификация и характеристика эндотелиальной циркулярной РНК, регулируемой гипоксией» . Исследование кровообращения . 117 (10): 884–890. дои : 10.1161/CIRCRESAHA.115.306319 . ПМИД 26377962 .
- ^ Саммертон Дж (декабрь 1999 г.). «Морфолино-антисмысловые олигомеры: случай структурного типа, независимого от РНКазы H». Biochimica et Biophysical Acta (BBA) – Структура и экспрессия генов . 1489 (1): 141–158. дои : 10.1016/S0167-4781(99)00150-5 . ПМИД 10807004 .
- ^ Кэпел Б., Суэйн А., Николис С., Хакер А., Уолтер М., Купман П. и др. (июнь 1993 г.). «Циркулярные транскрипты гена, определяющего семенники Sry, в семенниках взрослой мыши» . Клетка . 73 (5): 1019–1030. дои : 10.1016/0092-8674(93)90279-у . ПМИД 7684656 .
- ^ Дубин Р.А., Казми М.А., Острер Х. (декабрь 1995 г.). «Инвертированные повторы необходимы для циркуляризации транскрипта Sry семенников мыши». Джин . 167 (1–2): 245–248. дои : 10.1016/0378-1119(95)00639-7 . PMID 8566785 .
- ^ Бхардвадж В., Сингх А., Далави Р., Ралте Л., Чаунгту Р.Л., Сентил Кумар Н. и др. (04.11.2022). «Vpr ВИЧ-1 индуцирует ciTRAN, чтобы предотвратить подавление транскрипции провируса» . bioRxiv : 2022.11.04.515166. дои : 10.1101/2022.11.04.515166 . S2CID 253371318 .
- ^ Чен С.И., Сарнов П. (апрель 1995 г.). «Инициация синтеза белка эукариотическим трансляционным аппаратом на кольцевых РНК». Наука . 268 (5209): 415–417. Бибкод : 1995Sci...268..415C . дои : 10.1126/science.7536344 . ПМИД 7536344 .
- ^ Перейти обратно: а б Хентце М.В., Прейсс Т. (апрель 2013 г.). «Кольцевые РНК: варианты загадки сплайсинга» . Журнал ЭМБО . 32 (7): 923–925. дои : 10.1038/emboj.2013.53 . ПМЦ 3616293 . ПМИД 23463100 .
- ^ Перейти обратно: а б Чжан Ю., Чжан С.О., Чен Т., Сян Ю.Ф., Инь Ц.Ф., Син Ю.Х. и др. (сентябрь 2013 г.). «Циркулярные интронные длинные некодирующие РНК» . Молекулярная клетка . 51 (6): 792–806. doi : 10.1016/j.molcel.2013.08.017 . ПМИД 24035497 .
- ^ Берд С.Э., Джек В.Р., Лю Ю., Санофф Х.К., Ван З., Шарплесс Н.Е. (декабрь 2010 г.). «Экспрессия линейных и новых кольцевых форм некодирующей РНК, связанной с INK4/ARF, коррелирует с риском атеросклероза» . ПЛОС Генетика . 6 (12): e1001233. дои : 10.1371/journal.pgen.1001233 . ПМЦ 2996334 . ПМИД 21151960 .
- ^ Бхардвадж В., Сингх А., Чоудхари А., Далави Р., Ралте Л., Чаунгту Р.Л. и др. (сентябрь 2023 г.). «Vpr ВИЧ-1 индуцирует ciTRAN для предотвращения репрессии транскрипции провируса» . Достижения науки . 9 (36): eadh9170. Бибкод : 2023SciA....9H9170B . дои : 10.1126/sciadv.adh9170 . ПМЦ 10482341 . ПМИД 37672576 .
- ^ Ван, Ин; Лю, Бинь (17 мая 2020 г.). «Циркулярная РНК в больном сердце» . Клетки . 9 (5): 1240. doi : 10.3390/cells9051240 . ISSN 2073-4409 . ПМЦ 7290921 . ПМИД 32429565 .
- ^ Цзинь, Хуан; Ши, Ян, Хуэй; Ли, Ваньхай, Юй-ханг; Цай, Мэн, Сяо-мин (июнь 2020 г.) . / Журнал клеточной и молекулярной медицины . 24 (12): 6523–6533. doi : jcmm.15295 . ISSN 1582-1838 . PMC 7299708. 10.1111 PMID 32333642 .
- ^ Перейти обратно: а б с Цзэн, Сяньхуэй; Юань, Сянлэй; Цай, Цюй; Тан, Ченгвэй; Гао, Цзиньхан (август 2021 г.). «Циркулярная РНК как эпигенетический регулятор при хронических заболеваниях печени» . Клетки . 10 (8): 1945. doi : 10.3390/cells10081945 . ISSN 2073-4409 . ПМЦ 8392363 . ПМИД 34440714 .
- ^ Перейти обратно: а б с Ли, Вэйчжэнь; Лю, Цзя-Цян; Чен, Мин; Сюй, Цзян; Чжу, Ди (март 2022 г.). «Циркулярная РНК в развитии рака и иммунной регуляции» . Журнал клеточной и молекулярной медицины . 26 (6): 1785–1798. дои : 10.1111/jcmm.16102 . ISSN 1582-1838 . ПМЦ 8918416 . ПМИД 33277969 .
- ^ Амайя, Лаура; Григорян, Лилит; Ли, Чжицзянь; Ли, Одри; Вендер, Пол А.; Пулендран, Бали; Чанг, Ховард Ю. (16 мая 2023 г.). «Циркулярная РНК-вакцина индуцирует мощные Т-клеточные реакции» . Труды Национальной академии наук . 120 (20): e2302191120. Бибкод : 2023PNAS..12002191A . дои : 10.1073/pnas.2302191120 . ISSN 0027-8424 . ПМЦ 10193964 . ПМИД 37155869 .
- ^ Перейти обратно: а б с д Чжан, Пэнпэн; Чжан, Жуй; Ван, Ялин; Хан, Цюнь; Сюй, Хайся; Сюй, Юнцзе (2019-08-13). . «Циркулярная РНК-регуляция миогенеза» . Клетки . 8 ): 885. : 10.3390 / 8080885. ISSN 2073-4409 . PMC 6721685 . ( 8 клетки doi
- ^ Сэнгер Х.Л., Клотц Г., Риснер Д., Гросс Х.Дж., Кляйншмидт А.К. (ноябрь 1976 г.). «Вироиды представляют собой одноцепочечные ковалентно замкнутые кольцевые молекулы РНК, существующие в виде стержнеобразных структур с большим количеством спаренных оснований» . Труды Национальной академии наук Соединенных Штатов Америки . 73 (11): 3852–3856. Бибкод : 1976PNAS...73.3852S . дои : 10.1073/pnas.73.11.3852 . ПМЦ 431239 . ПМИД 1069269 .