Enanntiionithes
| Enanntiionithes | |
|---|---|

| |
| фоссил бохаорнитида Специфический | |
| Научная классификация | |
| Домен: | Эукариота |
| Королевство: | Животное |
| Филум: | Chordata |
| Клада : | Динозаурия |
| Клада : | Сауриския |
| Клада : | Теропода |
| Клада : | Avialae |
| Клада : | Ornithothoraces |
| Клада : | † Enanntiionithes Уокер , 1981 |
| Подгруппы | |
|
и см. Текст | |
Enantiornithes , также известные как Enantiornithines или Enantiornitheans в литературе, представляют собой группу вымерших авианов («птицы» в широком смысле), самая распространенная и разнообразная группа, известная из мезозойской эпохи . [ 3 ] [ 4 ] [ 5 ] Почти все удерживаемые зубы и когтистые пальцы на каждом крыле, но в остальном выглядели очень похоже на современные птицы снаружи. Более восьмидесяти видов Enantiornithes были названы, но некоторые имена представляют только одиночные кости, поэтому вполне вероятно, что не все действительны. Enantiornithes вымерли на границе мела-палеогена , наряду с Hesperornithe и всеми другими неавийскими динозаврами .
Открытие и именование
[ редактировать ]Первые обнаруженные Enantiornithes были неправильно направлены в современные группы птиц. Например, первые известные виды Enantiornithes, Gobipteryx Minuta , первоначально считали палеогном, связанным с страусами и Тинаму . [ 6 ] Enantiornithes были сначала признаны как отдельная линия, или «подкласс» птиц, Кирилом А. Уокером в 1981 году. Уокер сделал это открытие на основе некоторых частичных остатков из позднего мелового периода того, что сейчас является Аргентиной , которую он назначил Новый род, Enantiornis , дает всей группе свое название. С 1990 -х годов было обнаружено много более полных образцов энантиорнитов, и было установлено, что несколько ранее описанных «птиц» (например, iberomesornis , Cathayornis и Sinornis ) также были энантиорнинами.
Название «Enantiornithes» означает «противоположные птицы», от древнегреческого энантиоса ( ἐνάντιος ) «противоположные» + орниты ( ὄρνιθες ) "Птицы". Название было придумано Кирилом Александром Уокером в его знаменитой газете, которая основала группу. [ 7 ] В своей статье Уокер объяснил, что он имел в виду под «противоположностью»:
Возможно, наиболее фундаментальная и характерная разница между энантиорнитами и всеми другими птицами - это природа артикуляции между лопатой [...] и коракоидом , где «нормальное» состояние полностью обращено. [ 7 ]
Это относится к анатомической особенности-артикуляции костей плечевых костей , которая имеет вогнутую конъектную розетку между лопаткой (лопаточный лопаток) и коракоидом (первичная кость плечевого пояса у позвоночных, кроме млекопитающих), которая является наоборот это современных птиц. В частности, в Enantiornithes лопатка вогнута и в форме блюда в этом соединении, а коракоид выпуклый. [ 3 ] : 249–50 У современных птиц коракоскапулярный сустав имеет вогнутую коракоидную и выпуклую лопату. [ 8 ] [ 9 ]
Уокер не был ясен в своих причинах дать это имя в разделе этимологии своей статьи, и эта двусмысленность привела к некоторой путанице среди более поздних исследователей. Например, Алан Федучча заявил в 1996 году:
Птицы названы так, потому что, среди многих отличительных черт, существует уникальное образование триоссового канала, а плюсневые костюмы сливаются проксимально к дистальному, противоположное этому у современных птиц [ 10 ]
Точка зрения Федуччии о Tarsometatarsus (комбинированная верхняя и лодыжка) является правильной, но Уокер не использовал эту рассуждения в своей оригинальной статье. Уокер никогда не описывал слияние Tarsometatarsus как противоположного, а скорее как «только частичный». Кроме того, не уверен, что Enantiornithes имели триоссовые каналы, поскольку не сохраняет эту особенность. [ 3 ]
Как группа, энантиорниты часто называют «энантиорнитинами» в литературе. Тем не менее, несколько ученых отметили, что это неверно, потому что следуя стандартным правилам формирования имен животных групп, это подразумевает ссылку только на подсемейство Enantiornithinae . После соглашений об именах, используемых для современных птиц, а также вымерших групп, было отмечено, что правильный термин «Enantiornithean». [ 11 ] [ 12 ]
Происхождение и диапазон
[ редактировать ]Praeornis , из оксфордского - киммериджского из Казахстана , возможно, был самым ранним известным членом Enantiornithes, согласно Agnolin et al. (2017). [ 13 ]
Птицы с уверенными идентифицированными характеристиками энантиорнитов, найденных в Австралии , Альбиане маастрихтском Южной и кампании Мексиканском Америке ( Алексарнис [ 14 ] ), Монголия и Западный край доисторической Азии предполагают мировое распределение этой группы или в относительно теплых регионах, по крайней мере. [ 15 ] Enantiornithes были обнаружены на каждом континенте, кроме Антарктиды . Окаменелости, относящиеся к этой группе, являются исключительно меловыми по возрасту, и считается, что энантиорниты вымерли в то же время, что и их родственники динозавров без авиланцев . Самые ранние известные Enantiornithes взяты из раннего мела Испании Noguerornis (например, ) и Китая (например, Protopteryx ) и последних из покойного мела Северной и Южной Америки (например, Avisaurus и Enantiornis ). Широко распространенное вхождение этой группы предполагает, что, по крайней мере, некоторые энантиорниты смогли пересекать океаны под их собственной властью; Они являются первой известной линией Avialan с глобальным распределением.
Описание
[ редактировать ]
Многие окаменелости энантиорнитов очень фрагментарны, а некоторые виды известны только из куска одной кости. Почти все образцы, которые являются полными, в полной артикуляции и с сохранением мягких тканей известны из Лас Хойас в Куэнке , Испании и группе Ихол в Ляонинге ( Китай ). Чрезвычайные остатки энантиорнитов также были сохранены в бирманских янтарных месторождениях от 99 миллионов лет назад и включают в себя полученные из них, описанные в 2017 году. [ 16 ] и 2018, [ 17 ] а также изолированные части тела, такие как крылья [ 18 ] [ 19 ] [ 20 ] и ноги. [ 19 ] [ 21 ] Эти янтарные останки являются одними из самых хорошо сохранившихся из всех мезозозовых динозавров. Окаменелости этой клады были обнаружены как в внутренних, так и в морских отложениях, что позволяет предположить, что они были экологически разнообразной группой.
Enantiornithes, по -видимому, включали кулики, пловцы, граниворы, насекомоядные, рыбаки и хищники. Подавляющее большинство энантиорнитов были небольшими, между размером воробья и звездочкой , [ 22 ] Однако отображают значительные различия в размерах с некоторыми видами. Крупнейшим видом в этом кладе включают Pengornis Houi , [ 23 ] Сянгорнис Шенми , [ 24 ] Чжорнис Хани , [ 22 ] и Mirarce Eatoni , [ 25 ] (с последним видом, описываемыми как сходные по размеру, с современными индюками), хотя, по крайней мере, существует также несколько более крупных видов, в том числе потенциально виды размером с краны, известные только по следам в формировании Eumeralla (и, возможно, также представлены в Формирование Wonthaggi по одной фуркуле ). [ 26 ] Среди наименьших описанных образцов - неназванные мысли, хотя голотип образцов Parvavis Chuxiongensis [ 27 ] и Cratoavis cearensis [ 28 ] сопоставимы по размеру с небольшими сиськами или колибри.
Череп
[ редактировать ]
Учитывая их широкий спектр мест обитания и диет, морфология черепа энантиорнитов значительно различалась между видами. Черепа Enantiornithes объединили уникальный набор примитивных и передовых функций. Как и в более примитивных авиаланах, таких как Archeopteryx , они сохранили несколько отдельных черепных костей, небольших предчексил (кости кончика морды), а у большинства видов были зубастые челюсти, а не беззубые клювы. Только несколько видов, таких как Gobipteryx Minuta , были полностью беззубыми и имели клювы. У них также были простые квадратные кости , полная стержня, отделяющая каждую орбиту (отверстие для глаз) от каждого анторбитального фенестра , и зубные цифры (основные зубчатые кости нижней челюсти) без раздвоенных задних кончиков. Сквамозальная кость сохраняется в неопределенном юношеском образце, в то время как посторбитал сохраняется в Shenqiornis и Pengornis . У современных птиц эти кости ассимилируются в череп. У некоторых Enantiornithes, возможно, их временные фенестра (отверстия в стороне головы) объединены на орбиты, как у современных птиц из -за постбибибитов либо отсутствия, либо недостаточно долго, чтобы разделить отверстия. [ 29 ] Квадратоджугальная кость , которая у современных птиц сливается с югалом, сохраняется в Птеригорнисе . [ 30 ] Присутствие этих примитивных особенностей черепа сделало бы энантиорниты, способных только к ограниченному черепному кинезису (способность перемещать челюсть независимо от черепа). [ 31 ]
Крыло
[ редактировать ]
Как очень большая группа птиц, энантиорниты демонстрировали большое разнообразие различных планов тела, основанные на различиях в экологии и кормлении, отраженных в равном разнообразии форм крыльев, многих параллельных адаптациях к различным образам жизни, наблюдаемым у современных птиц. В целом, крылья энантиорнитов были продвинуты по сравнению с более примитивными авиаланами, такими как Archeopteryx , и показали некоторые особенности, связанные с полетом, аналогичными тем, которые встречаются в линии, ведущей к современным птицам, Ornithuromorpha . В то время как у большинства Enantiornithes были когти, по крайней мере, некоторые из их пальцев, у многих видов были укороченные руки, очень подвижный плечевой сустав и пропорциональные изменения в костях крыла, похожих на современные птицы. Как и современные птицы, Enantiornithes имели алулы или «ублюдки крылья», небольшие направления первых перьев на первой цифре, которая обеспечивала более высокую маневренность в воздухе и помогала в точных приземлениях. [ 32 ]
Несколько крыльев с консервированными перьями были найдены в бирманской янтаре . Это первое полное мезозойское динозавр, остается сохраняемым таким образом (в противном случае известны несколько изолированных перьев, не назначенных любыми видами), и один из самых изысканно сохранившихся динозаврских окаменелостей. [ 33 ] Сохраненные крылья показывают различия в пигменте перьев и доказывают, что Enantiornithes имели полностью современные перья, включая колючки, шарюлы и шкулеты, а также современное расположение перьев с крыльями, включая перья с длинными полетами, короткие челюсти, большую алулу и нижнюю часть шар. [ 18 ]
Одно ископаемое из энантиорнитов показывает, как крылоподобные пучки перьев на ногах, похожие на Archeopteryx . Перья ноги также напоминает четырехкрытый микрораптор динозавров , однако отличаются тем, что перья короче, более дезорганизованы (они явно не образуют крыло) и только распространяются на лодыжку, а не вдоль ноги. [ 34 ]
Хвост
[ редактировать ]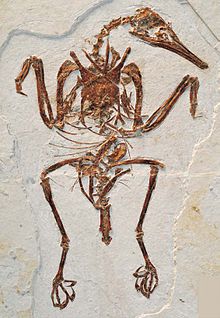
Clarke et al. (2006) обследовали все ископаемые энантиорнитов, которые затем известны и пришли к выводу, что ни одна из них не сохранила хвостовые перья, которые образовали вентилятор, создающий лифт, как у современных птиц. Они обнаружили, что все авиаланы за пределами Euornithes (клада, которую они называли Ornithurae ) с сохранившимися хвостовыми перьями, имели только короткие чехлы или удлиненные парные хвостовые перья. Они предположили, что развитие пигостила в энантиоорнитах, должно быть, было функцией укорочения хвоста, а не развития современной анатомии перья. Эти ученые предположили, что поклонник хвостовых перьев и связанной мускулатуры, необходимой для их контроля, известной как ректрическая луковица , развивалась вместе с коротким треугольным пигостилом, как у современных птиц, а не длинным, стержневым или кинжальным Пигостилы в более примитивных авиалянах, таких как энантиорниты. Вместо фаната перья у большинства Enantiornithes была пара длинных специализированных штифтов, аналогичных вымершим конфуциузинорнисам и некоторым существующим Птицы парадиса . [ 35 ]
Тем не менее, дальнейшие открытия показали, что, по крайней мере, среди базальных энантиорнитов, анатомия хвоста была более сложной, чем считалось ранее. Один род, Shanweiniao , был первоначально интерпретирован как по меньшей мере четыре длинных перья, которые перекрывали друг друга [ 36 ] и, возможно, сформировал подъемную поверхность, похожую на хвостовые вентиляторы Euronithes , [ 37 ] Хотя более позднее исследование указывает на то, что у Shanweiniao более вероятно, что перья с ручками с рахисом , похожие на перья, присутствующие в Paraprotopteryx . [ 38 ] У Chiappeavis , примитивного Pengornithid , был поклонник хвостовых перьев, похожий на более примитивные авиаланы, такие как Sapeornis , что позволяет предположить, что это могло быть наследственным состоянием, причем штифте -карницы были изготовлены несколько раз в ранних авианах для целей отображения. [ 38 ] У другого вида Enantiornithes, Feitianius , также был сложный поклонник перьев хвоста. Что еще более важно, мягкие ткани, сохранившиеся вокруг хвоста, были интерпретированы как останки ретрической луковицы, что позволяет предположить, что эта особенность на самом деле не ограничивалась видами с современными пигостилами, но могла развиваться гораздо раньше, чем предполагалось и присутствовало во многих Enantiornithes. [ 39 ] По крайней мере, один род Enantiornithes, Crolaispennia , имел современный пигостиль, но у него не было фаната хвоста. [ 40 ]
Биология
[ редактировать ]Диета
[ редактировать ]
Учитывая широкое разнообразие формы черепа среди Enantiornithe, многие различные диетические специализации должны были присутствовать среди группы. У некоторых, таких как Shenqiornis , были большие, крепкие челюсти, подходящие для еды с твердыми беспозвоночными. Короткие, тупые зубы Пенгорни, вероятно, использовались для питания членистоногих с мягким телом. [ 29 ] Сильные когтя Bohaiornithidae позволяют предположить, что они были хищниками из малых и средних позвоночных, но их надежные зубы вместо этого предполагают диету с жестким животным. [ 2 ]
Несколько образцов сохраняют фактическое содержание желудка. К сожалению, ни один из них не сохраняет череп, поэтому не может быть сделана прямая корреляция между их известной диетой и морской/формой зуба. Было обнаружено, что у эоалулависа останки экзоскелетов из водных ракообразных сохранились в пищеварительном тракте, [ 41 ] и Enantiophoenix сохранили корпускулы янтаря среди окаменелых костей, что позволяет предположить, что это животное, питаемое на сок деревьев, очень похоже на современные сапсеры и другие птицы. SAP был бы окаменело и стал янтарным. [ 42 ] Тем не менее, в последнее время было высказано предположение, что SAP двигался посмертным, поэтому не представляет истинное содержание желудка. В сочетании с предполагаемыми рыбными пеллетами Piscivorenantiornis яичниками оказались экскременты рыбы, странное содержание желудка некоторых видов оказалось , а предполагаемые гастролиты бохаорниса - случайные минеральные осадки, только эоалулавис демонстрирует фактическое содержание желудка. [ 43 ]
Исследование, проведенные паравивыми пищеварительными системами, показывает, что в известных Enantiornithes не было урожая и желудка, не использовало гастролиты и не выбрасывали гранулы. Это считается противоречивым с высоким разнообразием диет, которые подразумевают их различные зубы и форма черепа, [ 44 ] Хотя некоторые современные птицы потеряли желудок и полагаются исключительно на сильные желудочные кислоты. [ 45 ] Был обнаружен пример с тем, что предположительно, является гастролитами в том, что было бы желудком окаменелости, вновь открыв обсуждение использования гастролитов энантиорнитами. Рентгеновский и сканирующий инспекция пород пород определил, что они на самом деле были кристаллами халцедения, а не гастролитами. [ 46 ]
Longipterygidae является наиболее тщательно изученным семейством с точки зрения диеты из -за их довольно необычной ростральной анатомии, с длинными челюстями и несколькими зубами, расположенными на концах челюсти. Они по -разному интерпретировались как писковоры, [ 47 ] Прощатели, сродни береговым птицам [ 48 ] и как древесные коры. [ 49 ] Исследование 2022 года, однако, обнаруживает, что они, скорее всего, будут общими насекомоядными (без, возможно, Shengjingornis из -за его большего размера, плохо сохранившегося черепа и необычной анатомии педали), будучи слишком малым для специализированного плотоядного и травоядного; Атипичный рострам предварительно предполагается, что он не связан с питанием экологии. [ 50 ]
Хищничество
[ редактировать ]Окаменевает из Испании, сообщаемое Sanz et al. В 2001 году включали остатки четырех скелетов из трех различных видов энантионов. Они существенно полны, очень тесно связаны и показывают поверхностную ямку костей, что указывает на частичное пищеварение. Авторы пришли к выводу, что эта ассоциация представляла собой извергнутый гранул и, из деталей пищеварения и размера, в том, что птенцы были проглатываются целыми птерозавром или малым динозавром . Это было первым доказательством того, что мезозойские авиаляны были добычи животными, и что некоторые мезозойские пан-авианцы отрыгнули грануги, как Совы, делают сегодня. [ 51 ]
История жизни
[ редактировать ]
Известные ископаемые энантиорнит включают яйца , [ 52 ] [ 53 ] эмбрионы , [ 54 ] и детеныши . [ 55 ] Эмбрион, все еще скрученный в его яйце, сообщалось из формации Йикса . [ 56 ] Образцы несовершеннолетних могут быть идентифицированы по сочетанию факторов: грубая текстура их костей, указывающих на части, которые все еще были сделаны из хряща во время смерти, относительно небольших грудных костей, больших черепов и глаз, и костей, которые еще не слились друг с другом. Полем [ 57 ] Некоторым образцам хэтленда были даны официальные названия, в том числе « Liaoxiornis delicatus »; Тем не менее, Луис Чиаппе и его коллеги считали практику именования новых видов, основанных на несовершеннолетних, наносимых для изучения энантиорнитов, потому что практически невозможно определить, какие взрослые виды принадлежат данный юношеский образ Полем [ 57 ]
Вместе с образцами хэтла монгольского гобиптерикса [ 58 ] и Гобипип , [ 59 ] [ 60 ] Эти находки демонстрируют, что вылупление энантиорнитов имела скелетную окостенение, хорошо развитые крыло перья и большой мозг, которые коррелируют с предварительными или сверхпрочные паттерны развития у птиц сегодняшнего дня. Другими словами, Enantiornithes, вероятно, вылупились из яйца, уже хорошо развитого и готового к бегу, корму и, возможно, даже в возрасте всего несколько дней. [ 57 ]
Результаты предполагают, что энантиорниты, особенно зубчатые виды, имели более длительное время инкубации, чем современные птицы. [ 61 ] [ 62 ]
Анализ гистологии костей энантиорнита был проведен для определения скоростей роста этих животных. Исследование, проведенное в 2006 году костей , показало модель роста, отличную от современных птиц; Несмотря на то, что рост был быстрым в течение нескольких недель после вылупления, вероятно, до лечения , этот маленький вид долгое время не достигал размера взрослого, вероятно, несколько лет. [ 63 ] Все другие исследования подтвердили мнение о том, что рост до размера взрослого был медленным, как и у живых прекоциальных птиц (в отличие от пешеходных птиц, которые, как известно, быстро достигают размера взрослых). [ 41 ] Исследования скорости роста костей в различных энантиорнитах показали, что меньшие виды, как правило, расти быстрее, чем более крупные, противоположность схеме, наблюдаемому у более примитивных видов, таких как Jeholornis и у неавильских динозавров. [ 64 ] Некоторые анализы интерпретировали гистологию кости, чтобы указать, что Enantiornithes, возможно, не имели полной эндотермии птиц , вместо этого имея промежуточную скорость метаболизма . [ 65 ] Однако исследование 2021 года отвергает идею о том, что у них меньше эндотермических метаболизмов, чем современные птицы. [ 66 ]
Доказательства колониального гнездования были обнаружены в Enantiornithes, в отложениях позднего мела ( маастрихта ) Румынии . [ 67 ] Свидетельство от мест гнездования показывает, что Enantiornithes похоронили свои яйца, как современные мегаподы , что согласуется с их предполагаемыми суперпериальными адаптациями. [ 68 ]
Исследование, проведенное в 2020 году на перьях несовершеннолетних, еще больше подчеркивает онтологическое сходство с современными мегаподами, но предостерегает несколько различий, таких как древесная природа большинства энантиорнитов, в отличие от наземного образа жизни мегаподов. [ 69 ]
Предполагалось, что сверхпериальность в Enantiornithes мог бы помешать им разрабатывать специализированные аранжировки на ногах, которые можно увидеть у современных птиц, таких как Zygodactyly. [ 70 ]
Хотя подавляющее большинство исследований гистологии и известные остатки энантиорнитов указывают на то, что супернеплеальность является нормой, один из них, один из них, MPCM-LH-26189, по-видимому, представляет собой альтрический юношеский, что подразумевает, что, как и современные птицы, исследуют множественные репродуктивные стратегии. [ 71 ]
Полет
[ редактировать ]Поскольку у многих энантиорнитов не хватало сложных хвостов и обладали радикально другой анатомией крыла по сравнению с современными птицами, они были предметом нескольких исследований, проверяющих их возможности полета.
Традиционно они считались низшими флаерами, из-за того, что анатомия плечевого пояса считается более примитивной и неспособной поддерживать наземный механизм запуска,, [ 72 ] а также из -за отсутствия ремешков во многих видах. [ 35 ] [ 37 ] [ 73 ]
Тем не менее, несколько исследований показали, что это были эффективные флаеры, такие как современные птицы, обладающие аналогично сложной нервной системой и связками с пером крыла. Кроме того, отсутствие сложного хвоста, по -видимому, не было очень актуальным для полета птиц в целом - у некоторых вымерших птиц, таких как Lithornids, также не хватало сложных хвостовых перьев, но были хорошими листовками, [ 74 ] И они, кажется, были способны к запуску на основе наземного. [ 75 ]
Enantiornithes напоминают орнитуроморфы сурового киля отсутствует в наиболее базально-базально- во многих анатомических особенностях летного аппарата , но у Современные перья для хвоста вовлечены в полете. [ 76 ] Хотя некоторые базальные энантиорниты демонстрируют исконные летные аппараты, к концу мезозоя у многих энантиорнитов было несколько особенностей, сходящихся с неорнирующими, включая глубоко укоренившуюся грудину, узкую фуркулу с коротким гипоклямидием и локлевые ручки перо, которые указывают на повышенные аэродинамические способности. [ 77 ] [ 78 ]
По крайней мере, Элсорни, похоже, стал вторым безлеточным . [ 79 ]
Классификация
[ редактировать ]Некоторые исследователи классифицируют Enantiornithes, наряду с настоящими птицами, в классе Aves . Другие используют более ограничительное группы определение AVES (которое включает только неорнитов , анатомически современных птиц) и поместите Enantiornithes в более инклюзивной группе Avialae . Enantiornithes были более продвинутыми, чем Archeopteryx , Confuciusornis и Sapeornis , но в некоторых отношениях они были более примитивными, чем современные птицы, возможно, по промежуточному эволюционному пути.
Консенсус научного анализа указывает на то, что Enantiornithes является одной из двух основных групп в большей группе Ornithothoraces . Другая орнитоторациновая группа - Euornithes или Ornithuromorpha , которая включает в себя всех живых птиц в качестве подмножества. Это означает, что Enantiornithes были успешной ветвью эволюции Avialan, но той, которая полностью диверсифицировалась отдельно от линии, ведущей к современным птицам. [ 3 ] Однако одно исследование показало, что общая стернальная анатомия была приобретена независимо, и такие отношения должны быть пересмотрены. [ 80 ]
Классификация Enantiornithes и таксономия исторически были осложнены рядом факторов. В 2010 году палеонтологи Цзингмай О'Коннор и Гарет Дайк обрисовали в общих чертах ряд критических замечаний против преобладающих практик ученых, которые не могут описать многие образцы в достаточных количествах, чтобы другие могли тщательно оценить. Некоторые виды были описаны на основе образцов, которые проводятся в частных коллекциях, что делает дальнейшее изучение или обзор предыдущих результатов невозможными. Поскольку для других ученых часто невозможно изучить каждый образцы лично, учитывая всемирное распределение энантионий, и из -за многих неинформативных описаний, которые были опубликованы на, возможно, важных образцах, многие из этих образцов становятся «функциональной номинальной дубией ». [ 81 ] Кроме того, многие виды были названы на основе чрезвычайно фрагментарных образцов, которые не были бы очень информативными научно, даже если бы они были описаны в достаточной степени. Более одной трети всех названных видов основаны только на фрагменте одной кости. О'Коннор и Дайк утверждали, что, хотя эти образцы могут помочь расширить знания о промежутке времени или географического диапазона энантиорнинов, и важно описать их, именование таких образцов «неоправданно». [ 81 ]
Отношения
[ редактировать ]Enantiornithes - это сестринская группа для Euornithes , и вместе они образуют кладу, называемую Ornithothoraces (хотя см. Выше). Большинство филогенетических исследований вызвали энантиорниты как монофилетическую группу, отличную от современных птиц и их ближайших родственников. Филогенетический анализ 2002 года от Кларка и Норелла, тем не менее, уменьшил количество аутапоморфий Enantiornithes всего до четырех. [ 82 ]
Систематика Enantiornithes очень предварительно и общеизвестно сложно изучать, из -за их небольшого размера [ 28 ] и тот факт, что энантиорниты имеют тенденцию быть чрезвычайно гомопластичными или очень похожими друг на друга в большинстве своих скелетных особенностей из -за сходящейся эволюции, а не общего происхождения. [ 38 ] То, что кажется довольно уверенным, так это то, что в Enantiornithes, возможно, были подразделения, включающие некоторые незначительные базальные линии в дополнение к более продвинутым эйенантиорнитам. Детали взаимосвязи всех этих линий, действительно, обоснованность большинства, оспариваются, хотя Avisauridae, например, кажется, вероятно, составляют действительную группу. Филогенетические таксономисты до сих пор очень неохотно предполагали разграничение клад энантиорнитов. [ 83 ]
Одно из таких разграничений названо Euenantiornithes , было определено Chiappe (2002) как составляющий все виды ближе к Синорнису, чем к iberomesornis . Поскольку iberomesornis часто оказывается наиболее примитивным или базальным членом Enantiornithes, Euenantiornithes могут быть чрезвычайно инклюзивной группой, состоящей из всех Enantiornithes, за исключением iberomesornis самого . Несмотря на то, что в соответствии с филогенетической номенклатурой , это определение эуэнантиорнитов подвергалось серьезной критике со стороны некоторых исследователей, таких как Пол Серено , который назвал это «плохо определенной кладкой [...] хорошим примером плохого выбора в филогенетическом определении» Полем [ 83 ]
Кладограмма et ниже была обнаружена анализом Wang al. В 2015 году обновлен из предыдущего набора данных, созданного Jingmai O'Connor. [ 30 ]
| Ornithothoraces |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||





Кладограмма ниже от Wang et al. , 2022, и включает в себя большинство названных таксонов и восстанавливает несколько ранее названных клад. Письма на ветвях указывают позиции таксона «подстановочного знака», которые были восстановлены в нескольких разрозненных позициях. [ 84 ]
| Enanntiionithes |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| л |
Ключ к буквам:
b = Болуохия
C = Cathayornis
e = Enantiophopenix
f = houornis
h = longipteryx
1 = Parabohaiornis
J = Pterygornis
L = птица
m = yuanjiawaornis
n = yungavolucris
Список родов
[ редактировать ]Таксономию Enantiornithes трудно оценить, и в результате несколько клад в группе постоянно обнаруживаются с помощью филогенетических анализов. Большинство Enantiornithes не включены в какую -либо конкретную семью, и, как таковые, перечислены здесь. Многие из них считались евэнантиорнитами, хотя противоречия, лежащие в основе этого имени, означает, что оно не используется последовательно в исследованиях энантиорнитов. [ Цитация необходима ]
| Имя | Год | Формация | Расположение | Примечания | Изображения |
|---|---|---|---|---|---|
| Абаворнис | 1998 | Биссекти ( позднее , туронс меловое Формирование | Один из многих фрагментарных биссекти -энантиорнитов, известный только по коракоидам | ||
| Алетоалорнис | 2007 | Формирование джиуфотанг ( ранний меловой , аптиан ) | Плохо известный | ||
| Алексарнис | 1976 | Красная Бокана Формирование ( поздний меловой , кампанский ) | Один из первых энантиорнинов известен. Когда -то считалось древним родственником роликов и дятлов | 
| |
| Авиджи | 2019 | Формирование Xiagou ( раннее меловое , аптиан ) | Один образец этого рода умер с яйцом в его теле. | 
| |
| Bauxitornis | 2010 | Формирование CSEHBánya ( покойный меловой , сантониан ) | Фрагментарный, но уникальный в структуре его Tarsometatarsus | 
| |
| BreviroStruavis | 2021 | Формирование джиуфотанг ( ранний меловой , аптиан ) | Обладал увеличенным подкоими , который предполагает специализацию по кормлению, похожая на колибри, медоносных и дятеров | 
| |
| Castignovolucris | 2023 | Безыменная формация ( поздний меловой , кампанский ) | Возможно, размер канадского гуся | ||
| Catenoleimus | 1998 | Биссекти ( позднее , туронс меловое Формирование | Один из многих фрагментарных биссекти -энантиорнитов, известный только по коракоиду | ||
| Cathayornis | 1992 | Формирование джиуфотанг ( ранний меловой , аптиан ) | Один из первых описанного Jehol Biota Enantiornithes. Известно у многих видов, хотя некоторые теперь помещаются в свои собственные роды. Возможно, была похожая внешность и образ жизни с питтой | 
| |
| Комманка | 1992 | Лас Хойас ( Ранний меловой , Барреманец ) | Один из самых полных лас -хойас -энантиорнитов | 
| |
| Кратовис [ 85 ] | 2015 | Формирование Сантаны ( Раннее меловое , Аптиан ) | Очень хорошо сохранившийся южноамериканский член группы, в комплекте с лентоподобными перьями хвоста | ||
| Crurispennia [ 40 ] | 2017 | Huajiying formation ( раннее меловое , хаутеривское ) | Был необычный орнитуроморф, похожий на пигостиль и кисти, похожие на бедра перья. Один из самых старых энантионов | 
| |
| CuspiroStrisornis | 1997 | Формирование джиуфотанг ( ранний меловой , аптиан ) | Первоначально ошибочно считал, что у него был острый клюв | 
| |
| Dalingheornis | 2006 | Формирование Йикса ( Раннее меловое , Аптиан ) | Был хорошо адаптирован для лазания из-за его гетеродактильных ног , как у Трогона | ||
| Дунхуангия [ 86 ] | 2015 | Формирование Xiagou ( раннее меловое , аптиан ) | Род энантиорнитов из бассейна Чанма, область, в которой необычайно преобладают орнитуроморфы | ||
| Эльбрерон | 2009 | Формирование лечо ( поздние меловые , маастрихтские ) | Известно только из костей крыла. Может быть синонимом других лече, энантиорнит | ||
| Электорат | 2019 | Бирманская янтарь ( покойный меховой , сеноманский ) | Известно, сохраненный в янтаре с удлиненным средним носком | ||
| Enantiornis | 1981 | Формирование лечо ( поздние меловые , маастрихтские ) | Несмотря на то, что этот род был известен только из нескольких костей, это тезка энантиорнитов. Это также был один из крупнейших и последних представителей группы до их исчезновения | ||
| Eoalulavis | 1996 | Лас Хойас ( Ранний меловой , Барреманец ) | Сохраняет перья, включая алюлу , специализированный тип пера, который управляет потоком воздуха над крылом | ||
| Eocathayornis | 2002 | Формирование джиуфотанг ( ранний меловой , аптиан ) | Когда -то считается базальным близким родственником Cathayornis , хотя в настоящее время считается более отдаленным связанным | ||
| Eoenantiatis | 1999 | Формирование Йикса ( Раннее меловое , Аптиан ) | Хорошо сохранившиеся, но непоследовательные в филогенетическом размещении | 
| |
| Евгенавис | 2014 | Формирование Ilek ( Раннее меловое , Барремаское ) | Известно только по Tarsometatarsus, который разделяет некоторые особенности с такими же энантиорнитами | ||
| Исследователь | 1998 | Биссекти ( позднее , туронс меловое Формирование | Один из многих фрагментарных биссекти -энантиорнитов, известный только по коракоидам | ||
| Falcatakely | 2020 | Формирование Маеварано ( покойный меловой , маастрихтский ) | Разработал массивную морду только с одним зубом, несмотря на то, что он сохранил «примитивное» расположение черепа в отличие от современных птиц | 
| |
| Фейтиан [ 39 ] | 2015 | Формирование Xiagou ( раннее меловое , аптиан ) | Обладал сложным набором хвостовых перьев, в отличие от парных лентоподобных перьев большинства энантионов | 
| |
| Flexomornis | 2010 | Формирование деревянной кожи ( поздние меловые , сеноманские ) | Один из старейших авиаланов Североамериканской авиаляны, хотя и известно только по фрагментарным останкам | ||
| Fortipeavis | 2021 | Бирманская янтарь ( покойный меховой , сеноманский ) | Имел увеличенный внешний носок, который мог быть адаптацией для окуняемости | ||
| Fortunguavis [ 87 ] | 2014 | Формирование джиуфотанг ( ранний меловой , аптиан ) | Были надежные кости, в том числе ноги и когти, которые могли быть адаптированы для лазания по деревьям | ||
| Grabauornnis [ 88 ] | 2015 | Формирование Йикса ( Раннее меловое , Барремаское ) | Пропорции крыльев этого рода энантиорнитов, а также наличие алюлы предполагают, что это был хороший линейка | ||
| Gracilornis | 2011 | Формирование джиуфотанг ( ранний меловой , аптиан ) | Возможный родственник катайорниса с характерно тонкими костями | ||
| Гурилиния | 1999 | Формирование Немегта ( поздний меловой , маастрихтский ) | Плохо известный род Enantiornithe, но, очевидно, большой и поздний зажигающий член группы | ||
| Голландия [ 89 ] | 2010 | Формирование Barun Goyot ( поздние меловые , кампании ) | Первоначально идентифицированный как орнитуроморф, но с тех пор переосмысливается как род энантиорнитов, тесно связанных с лектавитами . [ 90 ] | 
| |
| Холботина [ 91 ] | 2015 | Формирование Andaikhudag ( ранний меловой , аптиан ) | Считается небольшим птерозавром с момента его открытия в 1977 году, пока он не получил официальное описание в 2015 году. Одержимые уникальные позвонки и примитивное небо. | ||
| Хуорнис | 1997 | Формирование джиуфотанг ( ранний меловой , аптиан ) | После того, как он считался сомнительным или видом Cathayornis , хотя исследование 2015 года считало, что оно является достоверным родом [ 92 ] | ||
| Huoshanornis | 2010 | Формирование джиуфотанг ( ранний меловой , аптиан ) | Возможно, был очень маневрируемым фияком из -за структуры его руки и грудины | ||
| Iberomesornis | 1992 | Лас Хойас ( Ранний меловой , Барреманец ) | Одно из первых родов энантиорнитов, известных с достойных останков. Также один из старейших и самых примитивных членов группы | 
| |
| Imparavis [ 93 ] | 2024 | Формирование джиуфотанг ( ранний меловой , аптиан ) | Самый ранний известный энантиорнитин с беззубым клювом | ||
| Incolornis | 1998 | Биссекти ( позднее , туронс меловое Формирование | Один из многих фрагментарных биссекти -энантиорнитов, известный только по коракоидам . Однажды считалось, что один вид принадлежит к Enantiornis | ||
| Юнорнис [ 94 ] | 2017 | Формирование Йикса ( Раннее меловое , Аптиан ) | Настолько хорошо сохранился, что его схема полета может быть восстановлен с использованием пропорций перьев и крыльев | 
| |
| Кизилкумавис | 1984 | Биссекти ( позднее , туронс меловое Формирование | Один из многочисленных фрагментарных биссекти -энантиорнитов, известный только по плечевой кости фрагменту | 
| |
| LargiroStrornis | 1997 | Формирование джиуфотанг ( ранний меловой , аптиан ) | Возможно, связанный с CuspiroStrisornis или синонимом Cathayornis | ||
| Лекта. | 1993 | Формирование лечо ( поздние меловые , маастрихтские ) | Большой и длинноногий член группы, пропорционально похожий на современные береговые птицы |  | |
| Ленесорнис | 1996 | Биссекти ( позднее , туронс меловое Формирование | Один из многих фрагментарных биссекти -энантиорнитов, известный только по фрагменту синсакрума . Первоначально считается, что принадлежит ichthyornis | ||
| Liaoningornis | 1996 | Формирование Йикса ( Раннее меловое , Аптиан ) | Первоначально считается орнитуран, но теперь считается родственником Eoalulavis | ||
| Longchengornis | 1997 | Формирование джиуфотанг ( ранний меловой , аптиан ) | Мог быть синонимом Cathayornis | ||
| Мартинавис | 2007 | Песчаник с образованием рептилий , образованием лечо ( покойный меловой , маастрихтский ) | Хотя этот род был известен только у гуммеров, был большим и жил в широком диапазоне | ||
| Microenantiatis | 2017 | Формирование джиуфотанг ( ранний меловой , аптиан ) | Небольшой член группы, которая обладала несколькими примитивными и полученными признаками по сравнению с другими энантиорнитами | ||
| Мирусавис | 2020 | Формирование Йикса ( меловое , Барремавство Аптиан ) Раннее | Голотип был маленькой остеологически незрелой самкой, сохранившейся медуллярной костной тканью | ||
| Monoenantisti [ 95 ] | 2016 | Формирование Йикса ( Раннее меловое , Аптиан ) | Известно из юношеского образца, который изображает, как различные признаки развивались в Enantiornithes с возрастом | ||
| Мусивавис | 2022 | Формирование джиуфотанг ( ранний меловой , аптиан ) | Большинство похожи на бохаорнитиды, но также имеют особенности других групп энантиорнитов | ||
| Нанантий | 1986 | Формирование Toolebuc ( ранний меловой , альбиан ) | Фрагментарный, но, возможно, был морской птицей, потому что останки из этого рода были найдены как Ichthyosaur содержание кишечника | ||
| Noguerornis | 1989 | El Montsec ( Ранний меловой , Барреманский ) | Сохраняет впечатления от пропатагия , кожи на плече, который образует часть крыла | ||
| Ориенанья | 2018 | Huajiying formation ( раннее меловое , хаутеривское ) | Многие детали мягких тканей образцов из этого рода были выявлены с помощью ультрафиолетового света | ||
| Otogornis | 1993 | Формирование Иджинхолуо ( Раннее меловое ) | Плохо известный | ||
| Paraprotopteryx | 2007 | Член Qiaotou of the Huajiying ( Ранний меловой , Аптиан ?) | По-видимому, имели четыре лентоподобные хвостовые перья вместо двух, как у большинства энантиорнитов | ||
| Парвавис [ 96 ] | 2014 | Формирование ( меловой , туронс покойный Цзяндихе | Небольшой, но полностью зрелый во время его смерти. Один из нескольких китайских энантиорнитов, датированных поздним меловым | 
| |
| Piscivorenantiisnis [ 97 ] | 2017 | Формирование джиуфотанг ( ранний меловой , аптиан ) | Известно из рассеянного скелета, сохранившегося на куске содержания желудка, состоящего из рыбных костей, что, возможно, было его последним блюдом | ||
| Protopteryx | 2000 | Huajiying formation ( раннее меловое , хаутеривское ) | Один из старейших и самых примитивных членов группы | 
| |
| Pterygornis [ 30 ] | 2016 | Формирование джиуфотанг ( ранний меловой , аптиан ) | Один рассеянный скелет из этого рода обладает хорошо сохранившимися костями черепа, в том числе квадратоджугал | ||
| Цилиана | 2011 | Формирование Xiagou ( раннее меловое , аптиан ) | Некоторые из останков этого рода включают в себя хорошо сохранившиеся задние конечности. Названия видов, Q. Graffini , названы в честь Грега Граффина из группы Bad Religion | ||
| Сазавис | 1989 | Биссекти ( позднее , туронс меловое Формирование | Один из многих фрагментарных биссекти -энантиорнитов, известный только по тибиотарсу (голени) | 
| |
| Шанхай | 2019 | Формирование джиуфотанг ( ранний меловой , аптиан ) | Необычно, предназначные из этого рода были слиты | ||
| Синорнис | 1992 | Формирование джиуфотанг ( ранний меловой , аптиан ) | Один из первых описанного Jehol Biota Enantiornithes. Похоже на Cathayornis , но обычно считается отличным | 
| |
| Сянгорнис | 2012 | Формирование джиуфотанг ( ранний меловой , аптиан ) | Рука этого рода была похожа на руку орнитуроморфа, вероятно, посредством сходящейся эволюции . Крупный член группы | ||
| Ятенвис [ 98 ] | 2022 | Формирование Хоррильо ( поздний меловой , маастрихтский ) | Самый южный известный член Enantiornithes и один из самых молодых членов группы | ||
| Yuanjiawaornis [ 99 ] | 2015 | Формирование джиуфотанг ( ранний меловой , аптиан ) | Один из крупнейших энантиорнинов, известных по достойным останкам | ||
| Юнгаволукрис | 1993 | Формирование лечо ( поздние меловые , маастрихтские ) | Имел большой и необычайно широкий тарсометатарсальный (кость нагнетателя) | 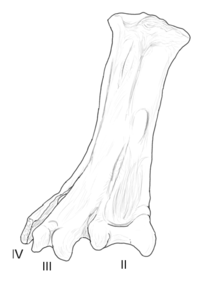
| |
| Юорнис | 2021 | Формирование Qiupa ( поздний меловой , маастрихтский ) | Большой, беззубые род Enantiornithes, один из самых жесток сохранившихся членов из позднего мела. | 
|
Longipterygidae
[ редактировать ]Longipterygidae . было семейством давних ранних мела-энантиорнитов, с зубами только на кончиках морды Как правило, они считаются довольно базальными членами группы. [ 36 ]
| Имя | Год | Формация | Расположение | Примечания | Изображения |
|---|---|---|---|---|---|
| Болуохия | 1995 | Формирование джиуфотанг ( ранний меловой , аптиан ) | Первоначально по ошибке считал, что обладал подсековым клювом | ||
| Camptodontornis | 2010 | Формирование джиуфотанг ( ранний меловой , аптиан ) | Первоначально называется Camptodontus , хотя это название рода занято жуком | ||
| Dapingfangornis | 2006 | Формирование джиуфотанг ( ранний меловой , аптиан ) | Возможно, на его лбу была структура, похожая на шипку | ||
| Longipteryx | 2001 | Формирование джиуфотанг ( ранний меловой , аптиан ) | Самый распространенный и известный член семьи | 
| |
| Лонгростравис | 2004 | Формирование Йикса ( Раннее меловое , Аптиан ) | Как и другие лонгиптеригиды, она обладала тонкой мордой, которая могла быть использована для зондирования беспозвоночных в грязи или коре | 
| |
| Rapaxavis | 2009 | Формирование джиуфотанг ( ранний меловой , аптиан ) | Специализирован на уход из -за структуры его ног | 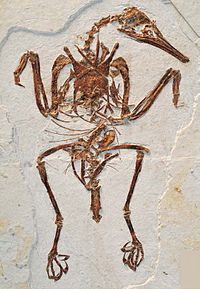
| |
| Shanweiniao | 2009 | Формирование Йикса ( Раннее меловое , Аптиан ) | Приобретенные несколько перьев хвоста, которые могли быть способны генерировать подъемник, как у современных птиц | 
| |
| Шелгорнис | 2012 | Формирование джиуфотанг ( ранний меловой , аптиан ) | Большой член семьи |
Pengornithidae
[ редактировать ]Pengornithidae . была семейством больших ранних энантионов У них было множество маленьких зубов и многочисленные примитивные особенности, которые теряются в большинстве других энантиорнинов. [ 1 ] В основном известный из раннего мела Китая, с предполагаемым поздним меловым таксоном от Мадагаскара. [ 100 ]
| Имя | Год | Формация | Расположение | Примечания | Изображения |
|---|---|---|---|---|---|
| Chiappeavis [ 38 ] | 2015 | Формирование джиуфотанг ( ранний меловой , аптиан ) | Обладал хвостом в форме вентилятора, состоящим из многих перьев | 
| |
| Eopengornis | 2014 | Huajiying formation ( раннее меловое , хаутеривское ) | Самый старый известный член семьи и одно из самых старых предполагаемых родов энантиорнитов известных. Обладают чрезвычайно хорошо сохранившимися хвостовыми лентами | ||
| Парапунгорнис [ 101 ] | 2015 | Формирование джиуфотанг ( ранний меловой , аптиан ) | Предполагается, что иметь стиль жизни, похожий на дятел, из-за черт ноги и хвоста | 
| |
| Pengornis | 2008 | Формирование джиуфотанг ( ранний меловой , аптиан ) | Первый Pengornithid обнаружил, а также один из крупнейших членов энантиорнитов, известных с приличных останков | ||
| Юанчуавис | 2021 | Формирование джиуфотанг ( ранний меловой , аптиан ) | Обладает тщательно продуманным хвостовым вентилятором «пинтах» дольше, чем его тело, у которого, возможно, была функция отображения | 
|
Bohaiornithidae
[ редактировать ]Bohaiornithids были большими, но геологически недолговечными ранними Enantiornithes, с длинными, зацепленными когтями и крепкими зубами с изогнутыми кончиками. Возможно, они были эквивалентны птицам добычи, хотя эта интерпретация открыта для больших дебатов. [ 2 ] Монофилия эволюционной этой группы сомнительна, и она может быть оценкой . [ 102 ]
| Имя | Год | Формация | Расположение | Примечания | Изображения |
|---|---|---|---|---|---|
| Бейгурнис | 2022 | Формирование Лонгцзяна ( Раннее меловое , Аптиан ) | Имел короткий, но надежный ручной Ungual | ||
| Bohaiornis | 2011 | Формирование джиуфотанг ( ранний меловой , аптиан ) | Первоначально считался сохраненным гастролитами , хотя позже было обнаружено, что они были обнаружены минеральными конкрециями | 
| |
| Гретхениао | 2019 | Формирование Йикса ( Раннее меловое , Барремаское ) | Адаптирован для хлопья, а не парящего, полета. Его описатели предлагают парафили или полифили из Bohaiornithidae | 
| |
| Linyiornis [ 103 ] | 2016 | Формирование джиуфотанг ( ранний меловой , аптиан ) | Возможный член семьи, известный из хорошо сохранившегося скелета, в комплекте со структурами, которые, как считается, развиваются яйца | ||
| Longusungguis | 2014 | Формирование джиуфотанг ( ранний меловой , аптиан ) | Довольно типичный член семьи | 
| |
| Parabohiornis | 2014 | Формирование джиуфотанг ( ранний меловой , аптиан ) | Близкий родственник Bohaiornis | ||
| Shenqiornis | 2010 | Член Qiaotou of the Huajiying ( Ранний меловой , Аптиан ?) | Первый известный член семьи, хотя и не считался близким родственником Bohaiornis до нескольких лет спустя. Сохраняет большую посторбитальную кость | ||
| Сулька | 2013 | Формирование Йикса ( Раннее меловое , Аптиан ) | Близкий родственник Shenqiornis с рифленой эмаль на зубах, уникальный среди ископаемых птиц | 
| |
| Чжорнис | 2013 | Формирование джиуфотанг ( ранний меловой , аптиан ) | Крупный член семьи с хорошо сохранившимся мозговым мозгом | 
|
Gobipterygidae
[ редактировать ]Некоторые члены группы неясны или плохо описаны и могут быть синонимом его видов типа Gobipteryx Minuta .
| Имя | Год | Формация | Расположение | Примечания | Изображения |
|---|---|---|---|---|---|
| Gobipteryx | 1974 | Формирование Barun Goyot ( поздние меловые , кампании ) | Бесзубитый усовершенствованный род энантиорнитов, обладающий надежным клювом, который сходятся с учетом современных птиц | ||
| Jibeinia | 1997 | Член Qiaotou of the Huajiying ( Ранний меловой , Аптиан ?) | Плохо известный и описанный со скелета, который сейчас потерян. Мог быть синонимом Vescornis | ||
| Вескорнис | 2004 | Член Qiaotou of the Huajiying ( Ранний меловой , Аптиан ?) | Небольшой и короткий род энантиорнитов, который может быть синонимом Jibeinia |
Avisauridae
[ редактировать ]Avisauridae подвергается двум различным определениям различной инклюзивности. Более инклюзивное определение, которое следует за Cau & Arduini (2008), используется здесь. Авизауриды были долговечным и широко распространенным семейством энантиорнитов, которые в основном различаются конкретными особенностями их тарсометатаралов (кости лодыжки). Крупнейшие и самые продвинутые члены группы выжили в Северной и Южной Америке до конца мела, но очень фрагментарны по сравнению с некоторыми более ранними таксонами.
| Имя | Год | Формация | Расположение | Примечания | Изображения |
|---|---|---|---|---|---|
| Ависавр | 1985 | Формирование Hell Creek ( покойный мелочный , маастрихтский ) | Одноименный авизаурид, а также один из крупнейших членов семьи. Первоначально считался динозавром без авиалана | 
| |
| Элсорн | 2007 | Формирование Джадохты ( поздний меловой , кампанский ) | Несмотря на неполное, его скелет обладает трехмерным сохранением. Возможно безлететь из -за своих пропорций крыла | ||
| EnantioPhoenix | 2008 | Формирование Уади Аль -Габур ( покойный меловой , сеноманский ) | что он питался на SAP деревье Когда -то считалось , | ||
| Gettyia | 2018 | Две медицины ( поздние меловые , кампанианские ) | Новый класс для славы Avisaurus | ||
| Халиморнис | 2002 | Формирование мела Мурвилля ( поздний меловой , кампанский ) | Жил в прибрежной среде | ||
| Интенсивно | 2010 | Формирование кожевенных заводов ( поздние меловые , кампании ) | Несмотря на то, что он тесно связан с некоторыми из крупнейших авизавридов, члены этого рода были очень маленькими птицами | 
| |
| Монкэнс | 2018 | Кайпаровитская формация ( поздний меловой , кампанский ) | Самый полный известный североамериканский авизаурид | 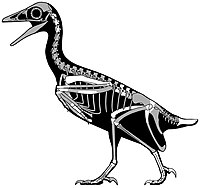
| |
| Mysiornis | 2011 | Формирование Ilek ( меловой , Барреманец Аптиан ) ранний | Обладает множеством особенностей различных групп в параве , хотя наиболее близко напоминает авизавриды среди выборки | ||
| Neuquenornis | 1994 | Формирование Bajo de la Carpa ( поздние меловые , сантониан ) | Обладают длинными крыльями и обратным ходом, указывающим на хороший рейс и способности | ||
| Soroavisaurus | 1993 | Формирование лечо ( поздние меловые , маастрихтские ) | Очень близкий родственник авизавра |
Сомнительные роды и заметные неназванные образцы
[ редактировать ]- Гобипип Решетови : описано в 2013 году из образцов эмбрионов в рамках яичной скорлупы из формирования Barun Goyot в Монголии . Эти образцы были очень похожи на образцы Embronic Gobipteryx , хотя описатели Gobipipus (набор противоречивых палеонтологов, включая Евгений Курочкин и Санкар Чаттерджи ) считают его различными. [ 104 ]
- Hebeiornis fengningensis : синоним Vescornis из -за того, что был описан из того же образца. Несмотря на то, что он был описан в 1999 году, за 5 лет до описания Vescornis , описание было настолько плохим по сравнению с описанием Vescornis , что последнее имя считается приоритетным большинством авторов. В результате имя Hebeiornis считается номенем Nudum («обнаженное имя»).
- « Проорнис »-неофициально названная птица из Северной Кореи. Это может не быть членом Enantiornithes.
- Liaoxiornis delicatus : описан в 1999 году из образца энантиорнитов, обнаруженных в формации Йикса . Первоначально этот образец считался крошечным взрослым, но позже обнаружил, что он вынужден. Другие образцы отныне были назначены роду. Из -за отсутствия отличительной черты многие палеонтологи считают этот род неагностированным номенем Дубий .
- "Wasaibpanchi": предполагаемый член Enantiornithes из Пакистана; Описанная статья имеет сомнительный статус.
- LP-4450 : несовершеннолетний неопределенный образец энантиорнитов из образования Эль-Монтесека в Испании . В его описании 2006 года изучалась гистология скелета, в то время как в более поздних исследованиях сообщалось о сквамозной кости, присутствующей в образце, но неизвестной в других энантиорнитах.
- IVPP V 13939 : Кратко описанный в 2004 году, этот экземпляр Einantiornithes был продвинутым на ногах, похожих на (хотя и короче), чем у других паравцев, таких как микрораптор и анкиорнис . [ 34 ]
- DIP-V-15100 и DIP-V-15101 : два разных крыла из образцов хэтленда, которые были описаны в 2015 году. Они привлекли значительное количество внимания среды при их описании. Они были сохранены в исключительных деталях из -за того, что они оказались в ловушке в бирманской янтаре в течение приблизительно 99 миллионов лет. [ 18 ]
- HPG-15-1 : частичный труп из хэтлиза Enantiornithes также сохранился в бирманском янтаре. Несмотря на неопределенное, он привлекло еще больше внимания средств массовой информации, чем два крыла при его описании в 2017 году. [ 16 ]
- CUGB P1202 : Неопределенный ювенильный бохаорнитид из формирования Jiufotang . Анализ его перья 2016 года обнаружил удлиненные предполагаемые меланосомы , предполагая, что большая часть его перья была радужными .
- DIP-V-15102 : Еще один труп неопределенного хэтла, сохранившийся в бирманской янтаре. Описано в начале 2018 года. [ 17 ]
- MPCM-LH-26189 A/B : частичный скелет хэтленда из Лас Хойа в Испании, включая компоненты как плиты, так и контр-солаб. В его описании 2018 года показано, как различные функции развивались в Enantiornithes по мере их возраста. особенности включают в себя окостенение грудины Такие из различных меньших костей и слияние хвостовых позвонков в пигостиль .
Ссылки
[ редактировать ]- ^ Jump up to: а беременный в Ван, х.; О'Коннор, JK; Zheng, x.; Ван, М.; Хм.; Zhou, Z. (2014). «Понимание эволюции rachis доминировало на хвостовых перьях из нового базального энантиорнитина (Aves: Ornithothoraces)» . Биологический журнал Линневого общества . 113 (3): 805–819. doi : 10.1111/bij.12313 .
- ^ Jump up to: а беременный в Ван, мин; Чжоу, Чжун-Хе; O'Connor, Jingmai K.; Зеленков, Никита В. (2014). «Новое разнообразное семейство Enantiornithine (Bohaiornithidae Fam. Nov.) Из нижнего мела Китая с информацией от двух новых видов» (PDF) . Повертеблат Паласиатика . 52 (1): 31–76.
- ^ Jump up to: а беременный в дюймовый Chiappe, Luis M.; Уокер, Кирилл А. (2002). «Скелетная морфология и систематика меловых эйентиорнитов (Ornithothotorces: Enantiornithes)» . В Чиаппе, Луис М.; Витмер, Лоуренс М. (ред.). Мезозойские птицы: над головами динозавров . Калифорнийский университет. С. 240–67. ISBN 978-0-520-20094-4 .
- ^ Chiappe, Luis M. (2007). Прославленные динозавры: происхождение и ранняя эволюция птиц . Хобокен, Нью -Джерси: Джон Уайли и сыновья. ISBN 978-0-471-24723-4 . [ страница необходима ]
- ^ O'Connor, Jingmai K.; Chiappe, Luis M.; Гао, Чунлинг; Чжао, Бо (сентябрь 2011 г.). «Анатомия раннего мела энантиорнитина птицы Rapaxavis pani» . Acta Palaeontologica Polonica . 56 (3): 463–475. doi : 10.4202/app.2010.0047 . S2CID 55311115 .
- ^ Эльзановский, Анджедж (1974). «Предварительная примечание о палеогнитной птице из верхнего мела Монголии» (PDF) . Palaeontologia Polonica . 29 : 103–9. S2CID 53487062 .
- ^ Jump up to: а беременный Уокер, Калифорния (1981). «Новый подкласс птиц из мелового периода Южной Америки». Природа . 292 (5818): 51–3. Bibcode : 1981natur.292 ... 51W . doi : 10.1038/292051A0 . S2CID 4340858 .
- ^ Надежда, Сильвия (2002). «Мезозойское излучение неорнитов». В Чиаппе, Луис М.; Витмер, Лоуренс М. (ред.). Мезозойские птицы: над головами динозавров . Калифорнийский университет. С. 339–88. ISBN 978-0-520-20094-4 .
- ^ Mayr, G. (2021). «Коракоскапулярный сустав птиц неорнитина - экстенская гомоплазия на широко забытой суставной поверхности птичьего грудного пояса и его возможных функциональных корреляций» . Zoomorphology . 140 (2): 217–28. doi : 10.1007/s00435-021-00528-2 . S2CID 236397491 .
- ^ Feduccia, Alan (1996). Происхождение и эволюция птиц . Нью -Хейвен: издательство Йельского университета. ISBN 978-0-300-06460-5 . [ страница необходима ]
- ^ Харрис, Джеральд Д.; Ламанна, Мэтью С.; Ты, Хай-Лу; Джи, Шу-Ан; Джи, Цянь (18 мая 2006 г.). «Второе Enantiornithean (Aves: Ornithothoraces) крыло раннего мелового образования Xiagou возле Чангмы, провинция Гансу, Китайская Народная Республика». Канадский журнал наук о Земле . 43 (5): 547–554. Bibcode : 2006cajes..43..547h . doi : 10.1139/e06-007 .
- ^ Ты, Хай-Лу; Ламанна, Мэтью С.; Харрис, Джеральд Д.; Chiappe, Luis M.; О'Коннор, Цзингмай; Джи, Шу-Ан; Лю, Джун Чанг; Юань, Чонг-Xi; Ли, Да-Цин; Чжан, Син; Лаковара, Кеннет Дж.; Додсон, Питер; Джи, Цянь (16 июня 2006 г.). «Почти современная амфибийная птица с раннего мела северо -западного Китая». Наука . 312 (5780): 1640–1643. Bibcode : 2006sci ... 312.1640y . doi : 10.1126/science.1126377 . PMID 16778053 . S2CID 42723583 .
- ^ Агнолин, Флорида, Розадилья, Св. И Мексэм Карвало (2017). , Юрский год Казахстана Историческая биология . doi : 10.1080/08912963.2017.1413102
- ^ Пирс Бродкорб (1976). Открытие меловой птицы, по -видимому, предков для приказов Coraciiformes и Picformes (Aves: Carinatae) [размер файла 70 МБ] (PDF) . Тол. 27. Смитсоновский вклад в палеобиологию. С. 67–73. Архивировано из оригинала (PDF) 19 октября 2014 года.
- ^ L. A. Nesov, A. A. Yarkov (1989). Новые птицы мел-палеогена СССР и некоторые замечания по истории возникновения и эволюции класса (New Cretaceous-Paleogene birds of USSR and some remarks about history of the origin and evolution of class) (PDF) (in Russian). Vol. 197. USSR Academy of scienses. p. 82. Archived (PDF) from the original on 27 March 2019.
- ^ Jump up to: а беременный Син, Лида; O'Connor, Jingmai K.; МакКеллар, Райан С.; Chiappe, Luis M.; Ценг, Куоуэй; Ли, банда; Бай, Мин (сентябрь 2017 г.). «Средне-мозговой энантиорнитин (авес) хэтланг, сохранившийся в бирманском янтаре с необычным оперением». Gondwana Research . 49 : 264–277. Bibcode : 2017gondr..49..264x . doi : 10.1016/j.gr.2017.06.001 .
- ^ Jump up to: а беременный Син, Лида; O'Connor, Jingmai K.; МакКеллар, Райан С.; Chiappe, Luis M.; Бай, Мин; Ценг, Куоуэй; Чжан, Цзе; Ян, Хайдонг; Клык, июнь; Ли, банда (февраль 2018 г.). «Уплощенный энантиорнитин в середине мозговой бирманской янтаря: морфология и сохранение» . Научный бюллетень . 63 (4): 235–243. Bibcode : 2018scibu..63..235x . doi : 10.1016/j.scib.2018.01.019 . PMID 36659012 .
- ^ Jump up to: а беременный в Син, Лида; МакКеллар, Райан С.; Ван, мин; Бай, Мин; O'Connor, Jingmai K.; Бентон, Майкл Дж.; Чжан, Цзянпинг; Ван, Ян; Ценг, Куоуэй; Локли, Мартин Дж.; Ли, банда; Чжан, Вейвей; Сюй, Син (28 июня 2016 г.). «Мумифицированные предварительные крылья птиц в середине мозговой бирманской янтарь» . Природная связь . 7 (1): 12089. Bibcode : 2016natco ... 712089x . doi : 10.1038/ncomms12089 . PMC 4931330 . PMID 27352215 .
- ^ Jump up to: а беременный Син, Лида; МакКеллар, Райан С.; O'Connor, Jingmai K.; Бай, Мин; Ценг, Куоуэй; Chiappe, Luis M. (30 января 2019 г.). «Полностью пернатый энантиорнитин ног и фрагмент крыла сохранились в середине межздорового бирманского янтаря» . Научные отчеты . 9 (1): 927. Bibcode : 2019natsr ... 9..927x . doi : 10.1038/s41598-018-37427-4 . PMC 6353931 . PMID 30700773 .
- ^ Син, Лида; МакКеллар, Райан С.; О'Коннор, Цзингмай К. (июнь 2020 г.). «Необычно большое птичье крыло в середине-мозговом бирманском янтаре». Мерашные исследования . 110 : 104412. BIBCODE : 2020CRRES.11004412X . doi : 10.1016/j.cretres.2020.104412 . S2CID 213510021 .
- ^ Син, Лида; O'Connor, Jingmai K.; Chiappe, Luis M.; МакКеллар, Райан С.; Кэрролл, Натан; Ху, Хан; Бай, Мин; Лей, Фумин (2019-07-22). «Новая птица энантиорнитина с необычными пропорциями педали, найденными в янтаре» . Текущая биология . 29 (14): 2396–2401.e2. doi : 10.1016/j.cub.2019.05.077 . ISSN 0960-9822 . PMID 31303484 . S2CID 195887085 .
- ^ Jump up to: а беременный Чжан, Зихуи; Chiappe, Luis M.; Хан, банда; Chinsamy, Anusuya (2013). «Большая птица из раннего мела Китая: новая информация о черепе энантиорнитинов». Журнал палеонтологии позвоночных . 33 (5): 1176–89. Bibcode : 2013jvpal..33.1176z . doi : 10.1080/02724634.2013.762708 . S2CID 84677039 .
- ^ Чжоу, Чжонге; Кларк, Джулия; Чжан, Фученг (май 2008 г.). «Понимание разнообразия, размера тела и морфологической эволюции от самой большой ранней меловой птицы энантиорнитина» . Журнал анатомии . 212 (5): 565–77. doi : 10.1111/j.1469-7580.2008.00880.x . PMC 2409080 . PMID 18397240 .
- ^ Ху, Донгу; Сюй, Син; Хоу, Лианхай; Салливан, Корвин (2012). «Новая писательница энантиорнитина из нижнего мела Западного Ляонинг, Китай, и ее последствия для ранней эволюции птиц». Журнал палеонтологии позвоночных . 32 (3): 639–45. Bibcode : 2012jvpal..32..639H . doi : 10.1080/02724634.2012.652321 . S2CID 85942925 .
- ^ Аттерхолт, Джесси; Хатчисон, Дж. Говард; О'Коннор, Цзингмай К. (13 ноября 2018 г.). «Наиболее полный энантиорнитин из Северной Америки и филогенетический анализ Avisauridae» . ПЕРЕЙ . 6 : E5910. doi : 10.7717/peerj.5910 . PMC 6238772 . PMID 30479894 .
- ^ Мартин, Энтони Дж.; Виккерс-богатая, Патриция; Рич, Томас Х.; Холл, Майкл; Angielczyk, Kenneth (январь 2014 г.). «Самые старые известные следы птиц из Австралии: формация Eumeralla (Albian), Dinosaur Cove, Victoria» . Палеонтология . 57 (1): 7–19. Bibcode : 2014palgy..57 .... 7m . doi : 10.1111/pala.12082 .
- ^ Ван, мин; Чжоу, Чжонге; Сюй, Гуанхуй (2014-01-01). «Первая писательница энантиорнитина из верхнего мела Китая». Журнал палеонтологии позвоночных . 34 (1): 135–145. Bibcode : 2014jvpal..34..135W . doi : 10.1080/02724634.2013.794814 . ISSN 0272-4634 . S2CID 85255564 .
- ^ Jump up to: а беременный Дуб, Исмар; Новости, Фернандо; Агнолин, Федерико; Исаси, Марсело; Фрейтас, Франциско; Андраде, Хосе (2015-06-05). «Новый род и виды птицы энантиорнитина из раннего мела Бразилии » . Бразильский журнал геологии . 45 (2): 161–171. Doi : 10.1590/23174889201500020001 .
- ^ Jump up to: а беременный O'Connor, Jingmai K.; Chiappe, Luis M. (28 февраля 2011 г.). «Пересмотр энантиорнитина (Aves: Ornithothoraces) морфология черепа». Журнал систематической палеонтологии . 9 (1): 135–157. Bibcode : 2011jspal ... 9..135o . doi : 10.1080/14772019.2010.5266639 . S2CID 86503357 .
- ^ Jump up to: а беременный в Ван, мин; Ху, Хан; Ли, Чжихенг (21 августа 2015 г.). «Новая маленькая птица энантиорнитина из биоты Джехола, что имеет значение для ранней эволюции морфологии птичьего черепа». Журнал систематической палеонтологии . 14 (6): 481–497. doi : 10.1080/14772019.2015.1073801 . S2CID 83603202 .
- ^ Ван, мин; Ху, Хан (январь 2017 г.). «Сравнительное морфологическое исследование Jugal и Quadratojugal у ранних птиц и их родственников динозавров» . Анатомическая запись . 300 (1): 62–75. doi : 10.1002/ar.23446 . PMID 28000410 . S2CID 3649504 .
- ^ Chiappe, Luis M. (2009). «Разрешенные динозавры: эволюционный переход к современным птицам» . Эволюция: образование и охват . 2 (2): 248–56. doi : 10.1007/s12052-009-0133-4 .
- ^ Беккер, Рэйчел (28 июня 2016 г.). «Птичьи крылья, пойманные в янтарь, являются ископаемым первым с эпохи динозавров». Природа . doi : 10.1038/nature.2016.20162 . S2CID 88601510 .
- ^ Jump up to: а беременный Чжан, Фученг; Чжоу, Чжонге (октябрь 2004 г.). «Палеонтология: перья ноги у ранней меловой птицы» . Природа . 431 (7011): 925. BIBCODE : 2004NATR.431..925Z . doi : 10.1038/431925a . PMID 15496911 . S2CID 4322054 .
- ^ Jump up to: а беременный Кларк, Джулия А.; Чжоу, Чжонге; Чжан, Фученг (март 2006 г.). «Понимание эволюции полета птичьего перелета от новой клады ранних мела -орнитуринов из Китая и морфологии Yixianornis Grabaui » . Журнал анатомии . 208 (3): 287–308. doi : 10.1111/j.1469-7580.2006.00534.x . PMC 2100246 . PMID 16533313 .
- ^ Jump up to: а беременный O'Connor, Jingmai K.; Ван, Сюри; Chiappe, Luis M.; Гао, Чунлинг; Мэн, Циндзин; Ченг, Сяодон; Лю, Цзиньюань (12 марта 2009 г.). «Филогенетическая поддержка специализированной клады меловых птиц энантиорнитина с информацией от нового вида». Журнал палеонтологии позвоночных . 29 (1): 188–204. Bibcode : 2009jvpal..29..188o . doi : 10.1080/02724634.2009.10010371 . S2CID 196607241 .
- ^ Jump up to: а беременный Chiappe, Luis M.; Бо, Чжао; O'Connor, Jingmai K.; Чунлинг, Гао; Xuri, Wang; Хабиб, Майкл; Маруган-Лобон, Иисус; Циндзин, Мэн; Xiaodong, Cheng (2014). «Новый образец ранней меловой птицы Hongshanornis longicresta : понимание аэродинамики и диеты базального орнитуроморфа» . ПЕРЕЙ . 2 : E234. doi : 10.7717/peerj.234 . PMC 3898307 . PMID 24482756 .
- ^ Jump up to: а беременный в дюймовый O'Connor, Jingmai K.; Ван, Сяоли; Чжэн, Сяотинг; Ху, Хан; Чжан, Xiaomei; Чжоу, Чжонге (январь 2016 г.). «Enantiornithine с хвостом в форме вентилятора и эволюция рекрического комплекса у ранних птиц» . Текущая биология . 26 (1): 114–119. doi : 10.1016/j.cub.2015.11.036 . PMID 26748849 .
- ^ Jump up to: а беременный O'Connor, Jingmai K.; Ли, Да-Цин; Ламанна, Мэтью С.; Ван, мин; Харрис, Джеральд Д.; Аттерхолт, Джесси; Вы, Хай-Лу (30 декабря 2015 г.). «Новый ранний меловой энантиорнитин (Aves, Ornithothoraces) из северо -западного Китая с тщательно продуманной хвостовой орнаментом». Журнал палеонтологии позвоночных . 36 (1): E1054035. doi : 10.1080/02724634.2015.1054035 . S2CID 85800831 .
- ^ Jump up to: а беременный Ван, мин; O'Connor, Jingmai K.; Пан, Янхон; Zhou, Zhonghe (2017-01-31). «Странная ранняя меловая птица энантиорнитина с уникальными перьями и пигостилом в форме орнитуроморфа» . Природная связь . 8 : 14141. Bibcode : 2017natco ... 814141W . doi : 10.1038/ncomms14141 . PMC 5290326 . PMID 28139644 .
- ^ Jump up to: а беременный Санз, Хосе Л.; Chiappe, Luis M.; Buscalioni, Angela D. (1995). «Остеология Concornis lacustris (Aves: Enantiornithes) из нижнего мела Испании и пересмотр его филогенетических отношений». Американский музей новичок (3133): 1–23. HDL : 2246/3667 .
- ^ Далла Веккия, Фабио М.; Chiappe, Luis M. (2003). «Первый птичий скелет из мезозоя северной гондваны». Журнал палеонтологии позвоночных . 22 (4): 856–60. doi : 10.1671/0272-4634 (2002) 022 [0856: fasftm] 2.0.co; 2 . JSTOR 4524284 . S2CID 130143737 .
- ^ О'Коннор, Цзингмай К. (1 января 2019 г.). «Трофические привычки ранних птиц». Палеогеография, палеоклиматология, палеоэкология . 513 : 178–195. Bibcode : 2019ppp ... 513..178o . doi : 10.1016/j.palaeo.2018.03.006 . S2CID 133781513 .
- ^ O'Connor, Jingmai K.; Чжоу, Чжонге; Смит, Эндрю (6 ноября 2019 г.). «Эволюция современной пищеварительной системы птиц: понимание от паравийских окаменелостей из биотов Янльяо и Джехола» . Палеонтология . 63 (1): 13–27. doi : 10.1111/pala.12453 .
- ^ Хьюстон, Дэвид С.; Copsey, JA (1994). «Пищеварение кости и морфология кишечника бородатого стервятника» . Журнал Raptor Research . 28 (2): 73–78.
- ^ «Короткий кристалл: кварц и окаменечная птица | Джоррии» . 2021-03-12 . Получено 2021-05-06 .
- ^ O'Connor, Jingmai K.; Чжоу, Чжонге; Чжан, Фученг (28 февраля 2011 г.). «Переоценка Болуохии Чженги (Aves: Enantiornithes) и обсуждение внутрикладического разнообразия в Jehol Avifauna, Китай». Журнал систематической палеонтологии. 9 (1): 51–63. doi: 10.1080/14772019.2010.512614. S2CID 84817636.
- ^ Lianhai Hou; Луизм. Chiappe; Фученг Чжан; Ченг-Мин Чуонг (2004). «Новая ранняя меловая ископаемость из Китая документирует новую трофическую специализацию для мезозойских птиц». Naturwissenschaften. 91 (1): 22–25. Bibcode: 2004nw ..... 91 ... 22h. doi: 10.1007/s00114-003-0489-1. PMC 4382005. PMID 14740099.
- ^ Morschhauser, Em; Варриккио, диджей; Гао, C.; Лю, Дж.; Wang, Z.; Cheng, X. & Meng, Q. (2009). «Анатомия ранней меловой птицы Rapaxavis pani, нового вида из провинции Ляонинг, Китай». Журнал палеонтологии позвоночных. 29 (2): 545–554. doi: 10.1671/039.029.0210. S2CID 84643293.
- ^ Миллер, Кейс Винсент; Питман, Майкл; Ван, Сяоли; Чжэн, Сяотинг; Яркая, Джен А. (2022). «Диета мезозойских зубчатых птиц (Longipterygidae), выведенная из количественного анализа существующих прокси -диетических прокси -животных» . BMC Biology . 20 (1): 101. doi : 10.1186/s12915-022-01294-3 . PMC 9097364 . PMID 35550084 .
- ^ Санз, Хосе Л.; Chiappe, Luis M.; Ферназ-Ялво, Иоланда; Ортега, Франциско; Санчес-Чилл, Бегона; Поято-Ариза1, Франциско Дж.; Перес-Морено, Бернардино П. (февраль 2001 г.). "У них ранний меховой пеллет" Природа 409 (6823): 998–1000. Bibcode : 2001natur . два : 1038/3 10 11234054PMID 663531S2CID
{{cite journal}}: CS1 Maint: числовые имена: список авторов ( ссылка ) - ^ Михайлов, Константин Э. (1991). «Классификация ископаемых яичных скорлупы амниотических позвоночных» (PDF) . Acta Palaeontologica Polonica . 36 (2): 193–238.
- ^ Михайлов, Константин Э. (1996). «Новые роды ископаемых яиц из верхнего мела Монголии». Палеонтологический журнал . 30 (2): 246–8.
- ^ Elżanowski, Andrzej (1981). «Скелеты эмбриональных птиц из покойного мела Монголии» (PDF) . Palaeontologia Polonica . 42 : 147–79.
- ^ Санз, Хосе Л.; Chiappe, Luis M.; Браун Перес, Бернардино П.; Мораталла, Хосе Дж.; Эрнандес-корабль, Франциско; Поиск, Анжела Д.; Ортега, Франциско; Поято-Ариза, Франциско Дж.; Расскин-Гутман, Диего; Martānez-Delclos, Ксавье (6 июня 1997 г.). Подразумеваемое Наука 276 (5318): 1543–6 Два : 10,1126/наука .
- ^ Чжоу, Чжонге; Чжан, Фученг (22 октября 2004 г.). «Предварительный птичий эмбрион из нижнего мела Китая». Наука . 306 (5696): 653. doi : 10.1126/science.1100000 . PMID 15499011 . S2CID 34504916 .
- ^ Jump up to: а беременный в Chiappe, Luis M.; Шуан, Джи; Цянь, Джи (2007). «Ювенильные птицы из раннего мела Китая: последствия для онтогенеза энантиорнитин» . Американский музей новичок (3594): 1–46. doi : 10.1206/0003-0082 (2007) 3594 [1: jbftec] 2.0.co; 2 . HDL : 2246/5890 . S2CID 85871695 .
- ^ Elżanowski, Andrzej (1995). «Переячные птицы и птичья филогения». Курьерский научно -исследовательский институт Сенкенберг . 181 : 37–53.
- ^ Курочкин, en; Chatterjee, S.; Михайлов, К.Е. (декабрь 2013 г.). «Эмбриональная птица энантиорнитина и связанные яйца из мела Монголии». Палеонтологический журнал . 47 (11): 1252–1269. Bibcode : 2013palj ... 47.1252K . doi : 10.1134/s0031030113110087 . S2CID 86747842 .
- ^ Курочкин, en; Chatterjee, S.; Михайлов, К.Е. (19 декабря 2013 г.). «Эмбриональная птица энантиорнитина и связанные яйца из мела Монголии». Палеонтологический журнал . 47 (11): 1252–1269. Bibcode : 2013palj ... 47.1252K . doi : 10.1134/s0031030113110087 . S2CID 86747842 .
- ^ Инкубационные периоды динозавра, непосредственно определяемые по количеству линий роста у эмбриональных зубов, показывают развитие рептилий-PNAS
- ^ Происхождение птичьего клюва: новые идеи инкубации динозавров
- ^ House-Moo, Оскар; Buscalioni, Deligated Angel; Куб, Хорхе; Каштан, Жак; Лот, Мари-Маделен; Дейзи, Эммануэль; Ricqlès, Armand (2006). Нижний меловой из Лас Хойас (Испания) Рендус учетные записи Palevol 5 (5): 685–91. Bibcode : 2006crpal . два 10.1016/j.crpv.2005.12.018:
- ^ O'Connor, Jingmai K.; Ван, мин; Чжэн, Ся-Тин; Ван, Сяо-Ли; Чжоу, Чжун-Хе (2014). «Гистология двух женских ранних меловых птиц» (PDF) . Повертеблат Паласиатика . 52 (1): 112–28.
- ^ Chiappe, LM (1995). «Филогенетическое положение меловых птиц Аргентины: Enantiornithes и Patagopteryx deferrariisi ». Курьер Forschungsinstitut Senckenberg . 181 : 55–63.
- ^ Кубо, Хорхе; Buscalioni, Angela D.; Legendre, Lucas J.; Бурдон, Эстель; Санз, Хосе Л.; Рикклес, Арманд (2021). «Палеогистологические выводы о скорости метаболизма в состоянии покоя у контактов и iberomesornis (Enantiornithes, Ornithothotoraces) из нижнего мелового из Лас -Хойаса (Испания)» . Палеонтология . 65 doi : 10.1111/pala.12583 . S2CID 245082389 .
- ^ Дайк, Гарет; Времир, Матиас; Кайзер, Гэри; Найш, Даррен (июнь 2012 г.). «Утопленная колония размножения мезозойских птиц из покойного мела Трансильвании». Умирайте натуральный . 99 (6): 435–42. Bibcode : 2012nw ..... 99..435d . Citeseerx 10.1.1.394.9006 . doi : 10.1007/s00114-012-0917-1 . PMID 22575918 . S2CID 1396792 .
- ^ Фернандес, Мариела С.; Гарсия, Родольфо А.; Фиорелли, Лукас; Сколаро, Алехандро; Сальвадор, Родриго Б.; Котаро, Карлос Н.; Кайзер, Гэри У.; Дайк, Гарет Дж.; Фарке, Эндрю А. (17 апреля 2013 г.). «Большое накопление птичьих яиц из позднего мела Патагонии (Аргентина) раскрывает новую стратегию гнездования в мезозойских птицах» . Plos один . 8 (4): E61030. BIBCODE : 2013PLOSO ... 861030F . doi : 10.1371/journal.pone.0061030 . PMC 3629076 . PMID 23613776 .
- ^ O'Connor, Jingmai K.; Фальк, Аманда; Ван, мин; Чжэн, Ся-Тин (2020). «Первый отчет о незрелых перьях у несовершеннолетних энантиорнитов раннего мелового Джехола Авифауны». Повертеблат Паласиатика . 58 : 24–44. doi : 10.19615/j.cnki.1000-3118.190823 .
- ^ Кларк, Александр Д.; О'Коннор, Цзингмай К. (15 июня 2021 года). «Изучение экоморфологии двух меловых энантиорнитинов с уникальной морфологией педали» . Границы в экологии и эволюции . 9 : 654156. DOI : 10.3389/fevo.2021.654156 .
- ^ Кэй, Томас Г.; Питман, Майкл; Марк-Лобон, Хесус; Мартин-Абад, Хьюго; Санз, Хосе Луис; Buscallioni, Анжела Д. (21 марта 2019 г.). «Полностью фледортин хэтлент, выявленная лазерной стимулированной флуоресценцией, подтверждает предварительное поведение гнездования » Научные отчеты 9 (1): 5006. Bibcode : 2019natsr ... 9.5006K Doi : 10.1038/ s41598-019-41423-7 PMC 64288442 PMID 30899080
- ^ Падиан, Кевин; Chiappe, Luis M. (11 января 2007 г.). «Происхождение и ранняя эволюция птиц» (PDF) . Биологические обзоры . 73 (1): 1–42. doi : 10.1111/j.1469-185x.1997.tb00024.x . S2CID 86007060 .
- ^ Чжоу, Шуан; Чжоу, Чжун-Хе; О'Коннор, Цзингмай К. (2012). «Новая базальная орниториновая птица из нижнего мела Западного Ляонинг, Китай» (PDF) . Повертеблат Паласиатика . 50 (1): 9–24.
- «Новая базальная орниториновая птица, обнаруженная из нижнего мела западного Ляонинг, Китай» . Phys.org . 13 февраля 2012 года.
- ^ Houde, Peter W. (1988). «Палеогнистые птицы из раннего третичного заведения северного полушария». Публикации орнитологического клуба Nuttall (Кембридж Массачусетс, США: Орнитологический клуб Nuttall) 22
- ^ Навалон, Уильям; Маруган-Лобон, Иисус; Chiappe, Luis M.; Луис Санз, Хосе; Buscalioni, Angel D. (6 октября 2015 г.). Я Научные отчеты 5 1 ) : ( два : 10 1038/ srep1 4594305PMC 26440221PMID
- ^ Брусатт, Стивен Л.; O'Connor, Jingmai K.; Джарвис, Эрих Д. (5 октября 2015 г.). «Происхождение и диверсификация птиц» . Текущая биология . 25 (19): R888–898. BIBCODE : 2015CBIO ... 25.R888B . doi : 10.1016/j.cub.2015.08.003 . HDL : 10161/11144 . ISSN 1879-0445 . PMID 26439352 . S2CID 3099017 .
- ^ Аттерхолт, Джесси; Хатчисон, Дж. Говард; О'Коннор, Цзингмай К. (2018). «Наиболее полный энантиорнитин из Северной Америки и филогенетический анализ Avisauridae» . ПЕРЕЙ . 6 : E5910. doi : 10.7717/peerj.5910 . ISSN 2167-8359 . PMC 6238772 . PMID 30479894 .
- ^ Ван, Ся; McGowan, Alistair J.; Дайк, Гарет Дж.; Терви, Сэмюэль Т. (7 декабря 2011 г.). «Пропорции крыла и стили полета: первый шаг к прогнозированию полетов мезозойских птиц» . Plos один . 6 (12): E28672. Bibcode : 2011ploso ... 628672W . doi : 10.1371/journal.pone.0028672 . PMC 3233598 . PMID 22163324 .
- ^ Chiappe, Luis M.; Suzuki, Shigeru; Дайк, Гарет Дж.; Ватабе, Махито; Tsogtbaatar, K.; Барсболд, Ринчен (январь 2007 г.). «Новая писательница энантирнитина из покойного мелового из пустыни Гоби». Журнал систематической палеонтологии . 5 (2): 193–208. Bibcode : 2007Jspal ... 5..193c . doi : 10.1017/s1477201906001969 . S2CID 85391743 .
- ^ Чжэн, Сяотинг; Ван, Сяоли; О'Коннор, Цзингмай; Чжоу, Чжонге (9 октября 2012 г.). «Понимание ранней эволюции птичьей грудины от ювенильных энантиорнитинов» . Природная связь . 3 (1): 1116. Bibcode : 2012natco ... 3.1116z . doi : 10.1038/ncomms2104 . PMID 23047674 .
- ^ Jump up to: а беременный О'Коннор, Цзингмай; Дайк, Грет (2010). Аннтанториты and Cathayornis yandica (Aves: Enantiornithes)". Записи австралийского музея 62 : 7–2 doi : 10.3853/ j.0067-1975.62.2010.1540
- ^ Кларк, Джулия А.; Норелл, Марк А. (2002). «Морфология и филогенетическое положение Апсаравис -Укхааны из позднего мела Монголии». Американский музей новичок (3387): 1–46. Citeseerx 10.1.1.693.8475 . doi : 10.1206/0003-0082 (2002) 387 <0001: tmappo> 2,0.co; 2 . S2CID 52971055 .
- ^ Jump up to: а беременный Sereno, PC (2005) Taxonsearch: Archosauria Archosauria Archosauria 2007-02-19 на машине Wayback . Версия 1.0, 2005-NOV- 7. Получено 2006-OCT-02.
- ^ Ван, Сюри; Кау, Андреа; Ло, Сяоалинг; Кундрат, Мартин; Wu, Wensheng; Джу, Шубин; Го, Чжэнь; Лю, Йичуань; Джи, Цянь (2022-02-11). «Новая птица, похожая на Бохайорнитид из нижнего мела Китая, заполняет разрыв в неравенстве энантиорнитина» . Журнал палеонтологии . 96 (4): 961–976. Bibcode : 2022jpal ... 96..961W . doi : 10.1017/jpa.2022.12 . ISSN 0022-3360 . S2CID 247432530 .
- ^ Карвалью; Новас; Агнолин; Исаси; Фрейтас; Андраде (2015). «Новый род и виды энантиорнитинской птицы из раннего мела Бразилии» . Бразильский журнал геологии . 45 (2): 161–171. doi : 10.1590/23174889201500020001 .
- ^ Ван, Ли; О'Коннор, Чжоу; Вы (2015). «Второй вид энантиорнитинской птицы из нижнего мела -бассейна Чанга, северо -западный Китай с последствиями для таксономического разнообразия Changma avifauna». Мерашные исследования . 55 : 56–65. Bibcode : 2015crres..55 ... 56w . doi : 10.1016/j.cretres.2015.01.008 .
- ^ Ван, М.; О'Коннор, JK; Zhou, Z. (2014). «Новая надежная птица энантиорнитина из нижнего мела Китая со сканирующими адаптациями». Журнал палеонтологии позвоночных . 34 (3): 657–671. Bibcode : 2014jvpal..34..657W . doi : 10.1080/02724634.2013.812101 . S2CID 85313872 .
- ^ Dalsätt, J.; Ericson, pg; Чжоу, З. (2015). «Новые Enantiornithes (Aves) из раннего мела Китая». Acta Geologica Sinica . 86 (2): 801–807. doi : 10.1111/1755-6724.12270 . S2CID 131170912 .
- ^ Белл, Алисса К.; Chiappe, Luis M.; Эриксон, Грегори М.; Suzuki, Shigeru; Ватабе, Махито; Барсболд, Ринчен; Tsogtbaatar, K. (февраль 2010 г.). «Описание и экологический анализ Голланды Лучерии, покойной меловой птицы из пустыни Гоби (Монголия)». Мерашные исследования . 31 (1): 16–26. Bibcode : 2010crres..31 ... 16b . doi : 10.1016/j.cretres.2009.09.001 .
- ^ Хартман, Скотт; Мортимер, Микки; Wahl, William R.; Lomax, Dean R.; Липпинкотт, Джессика; Лавелис, Дэвид М. (10 июля 2019 г.). «Новый паравийский динозавр из позднего юрского периода Северной Америки поддерживает позднее приобретение полета птиц» . ПЕРЕЙ . 7 : E7247. doi : 10.7717/peerj.7247 . PMC 6626525 . PMID 31333906 .
- ^ Зеленков, Никита В.; Averianov, Александр О. (13 июня 2015 г.). «Исторический образец энантиорнитинской птицы из раннего мела Монголии, представляющий новый таксон со специализированной морфологией шеи». Журнал систематической палеонтологии . 14 (4): 319–338. doi : 10.1080/14772019.2015.1051146 . S2CID 85784633 .
- ^ Ван, М.; Лю Д. (2015). «Таксономическая переоценка Cathayornithidae (Aves: Enantiornithes)». Журнал систематической палеонтологии . 14 : 1–19. doi : 10.1080/14772019.2014.994087 . S2CID 86665059 .
- ^ Ван, Сяоли; Кларк, Александр Д.; O'Connor, Jingmai K.; Чжан, Сяньгу; Ван, Син; Чжэн, Сяотинг; Чжоу, Чжонге (2024-02-27). «Первый беззубые энантиорнитин (Aves: Ornithothoraces) из нижнего мела Джехола Авифауна» . Мерашные исследования . 159 (в печати): 105867. DOI : 10.1016/j.cretres.2024.105867 . ISSN 0195-6671 .
- ^ Лю, ди; Chiappe, Luis M.; Серрано, Франциско; Хабиб, Майкл; Чжан, Югуан; Мэн, Циндзин; Шоуки, Мэтью (11 октября 2017 г.). «Аэродинамика полета в энантиорнитинах: информация от новой китайской ранней меловой птицы» . Plos один . 12 (10): E0184637. Bibcode : 2017ploso..1284637L . doi : 10.1371/journal.pone.0184637 . PMC 5636078 . PMID 29020077 .
- ^ Ху, Хан; О'Коннор, Цзингмай К. (14 ноября 2016 г.). «Первые виды энантиорнитов из Сихеданга выясняют развитие скелета в ранних мелах энантиорнитины». Журнал систематической палеонтологии . 15 (11): 909–926. doi : 10.1080/14772019.2016.1246111 . S2CID 89551799 .
- ^ Ван, мин; Чжоу, Чжонге; Сюй, Гуанхуй (7 января 2014 г.). «Первая писательница энантиорнитина из верхнего мела Китая». Журнал палеонтологии позвоночных . 34 (1): 135–145. Bibcode : 2014jvpal..34..135W . doi : 10.1080/02724634.2013.794814 . S2CID 85255564 .
- ^ Ван, мин; Чжоу, Чжонге (12 апреля 2017 г.). «Морфологическое исследование первой известной писци иродной энантиорнитиной птицы из раннего мела Китая». Журнал палеонтологии позвоночных . 37 (2): E1278702. Bibcode : 2017jvpal..37e8702w . doi : 10.1080/02724634.2017.1278702 . S2CID 89858642 .
- ^ Ху, Донгу; Лю, Инг; Ли, Джинхуа; Сюй, Син; Hou, Lianhai (июль 2015 г.). «Yuanjiawaornis viriosus, Gen. Et sp. Nov., Большая питательная птица энантиорнитина из нижнего мела Западного Ляонинг, Китай». Мерашные исследования . 55 : 210–219. Bibcode : 2015crres..55..210H . doi : 10.1016/j.cretres.2015.02.013 .
- ^ О'Коннор, Патрик М.; Тернер, Алан Х.; Гроенке, Джозеф Р.; Феличе, Райан Н.; Роджерс, Рэймонд Р.; Краузе, Дэвид В.; Рахантарисоа, Лидия Дж. (10 декабря 2020 г.). «Поздняя меловая птица из Мадагаскара раскрывает уникальное развитие клюков». Природа . 588 (7837): 272–276. Bibcode : 2020nater.588..272O . doi : 10.1038/s41586-020-2945-x . PMID 33239782 . S2CID 227174405 .
- ^ Ху, Хан; O'Connor, Jingmai K.; Чжоу, Чжонге; Фарке, Эндрю А. (3 июня 2015 г.). «Новые виды Pengornithidae (Aves: Enantiornithes) из нижнего мела Китая предполагают специализированную сканирующую среду обитания, ранее неизвестную у ранних птиц» . Plos один . 10 (6): E0126791. BIBCODE : 2015PLOSO..1026791H . doi : 10.1371/journal.pone.0126791 . PMC 4454694 . PMID 26039693 .
- ^ Chiappe, Luis M.; Циндзин, Мэн; Серрано, Франциско; Сигурдсен, Тронд; Мин, Ван; Белл, Алисса; Ди, Лю (25 октября 2019 г.). «Новая Bohaiornis -Like Bird из раннего мела Китая: взаимосвязь Enantiornithine и выступления полета» . ПЕРЕЙ . 7 : E7846. doi : 10.7717/peerj.7846 . PMC 6816414 . PMID 31667014 .
- ^ Ван, Ян; Ван, мин; O'Connor, Jingmai K.; Ван, Сяоли; Чжэн, Сяотинг; Чжан, Xiaomei (11 января 2016 г.). «Новая птица Ихола Энантиорнитин с трехмерным сохранением и фолликулами яичников». Журнал палеонтологии позвоночных . 36 (2): E1054496. BIBCODE : 2016JVPAL..36E4496W . doi : 10.1080/02724634.2015.1054496 . S2CID 85807045 .
- ^ Курочкин, en; Chatterjee, S.; Михайлов, К.Е. (19 декабря 2013 г.). «Эмбриональная птица энантиорнитина и связанные яйца из мела Монголии». Палеонтологический журнал . 47 (11): 1252–1269. Bibcode : 2013palj ... 47.1252K . doi : 10.1134/s0031030113110087 . S2CID 86747842 .
Внешние ссылки
[ редактировать ]![]() СМИ, связанные с Enantiornithes в Wikimedia Commons
СМИ, связанные с Enantiornithes в Wikimedia Commons

