Первичное производство
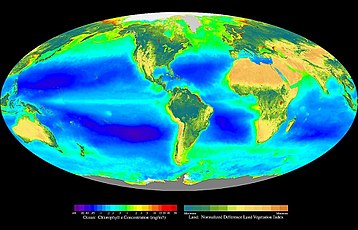
В экологии . основное производство синтез органических соединений из атмосферной или водной углекислоты — В основном это происходит в процессе фотосинтеза , который использует свет в качестве источника энергии, но также происходит в результате хемосинтеза , который использует окисление или восстановление неорганических химических соединений в качестве источника энергии. Почти вся жизнь на Земле напрямую или косвенно зависит от первичного производства. Организмы, ответственные за первичное производство, известны как первичные продуценты или автотрофы и составляют основу пищевой цепи . В наземных экорегионах это в основном растения , тогда как в водных экорегионах в этой роли преобладают водоросли . Экологи различают первичную продукцию как чистую , так и валовую : первая учитывает потери в таких процессах, как клеточное дыхание , а вторая – нет.
Обзор
[ редактировать ]Первичное производство — производство химической энергии в органических соединениях живыми организмами . Основным источником этой энергии является солнечный свет , но незначительная часть первичного производства обеспечивается литотрофными организмами, использующими химическую энергию неорганических молекул.
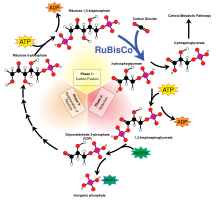
Независимо от источника, эта энергия используется для синтеза сложных органических молекул из более простых неорганических соединений, таких как углекислый газ (CO 2 ) и вода (H 2 O). Следующие два уравнения представляют собой упрощенное представление фотосинтеза (вверху) и (одной из форм) хемосинтеза (внизу):
- CO 2 + H 2 O + свет → CH 2 O + O 2
- CO 2 + O 2 + 4 H 2 S → CH 2 O + 4 S + 3 H 2 O
В обоих случаях конечной точкой является полимер восстановленных ( углеводов CH 2 O) n , обычно таких молекул, как глюкоза или другие сахара . Эти относительно простые молекулы могут затем использоваться для дальнейшего синтеза более сложных молекул, включая белки , сложные углеводы , липиды и нуклеиновые кислоты , или вдыхаться для выполнения работы . Потребление первичных продуцентов гетеротрофными организмами, такими как животные , затем переносит эти органические молекулы (и энергию, запасенную в них) вверх по пищевой сети , питая все . живые системы Земли [ нужна ссылка ]
Валовое первичное производство и чистое первичное производство
[ редактировать ]Валовая первичная продукция (GPP) — это количество химической энергии, обычно выражаемое в виде углеродной биомассы , которую первичные производители создают за определенный промежуток времени. Некоторая часть этой фиксированной энергии используется первичными продуцентами для клеточного дыхания и поддержания существующих тканей (т.е. «дыхание роста» и « дыхание поддержания »). [1] [2] Оставшаяся фиксированная энергия (т. е. масса фотосинтата) называется чистой первичной продукцией (ЧПП).
- НПП = ГПП – дыхание [растениями]
Чистая первичная продукция — это скорость, с которой все автотрофы в экосистеме производят чистую полезную химическую энергию. Чистая первичная продукция может быть направлена на рост и воспроизводство первичных производителей. Таким образом, он доступен для потребления травоядными животными. [ нужна ссылка ]
Как валовая, так и чистая первичная продукция обычно выражается в единицах массы на единицу площади за единицу времени. В наземных экосистемах масса углерода на единицу площади в год (г C м −2 тот −1 ) чаще всего используется в качестве единицы измерения. Обратите внимание, что иногда проводят различие между «производством» и «производительностью», причем первое - это количество произведенного материала (г C м −2 ), последний - скорость его производства (г C м −2 тот −1 ), но эти термины чаще используются как взаимозаменяемые. [ нужна ссылка ]
Наземное производство
[ редактировать ]На суше почти вся первичная продукция в настоящее время осуществляется сосудистыми растениями , при этом небольшая часть приходится на водоросли и несосудистые растения, такие как мхи и печеночники . До эволюции сосудистых растений несосудистые растения, вероятно, играли более значительную роль. Первичное производство на суше является функцией многих факторов, но главным образом местной гидрологии и температуры (последняя в некоторой степени зависит от света, особенно от фотосинтетически активной радиации (ФАР), источника энергии для фотосинтеза). Хотя растения покрывают большую часть поверхности Земли, их численность сильно сокращается там, где температуры слишком экстремальны или там, где необходимые растительные ресурсы (в основном вода и PAR) ограничены, например, в пустынях или полярных регионах . [ нужна ссылка ]
Вода «потребляется» растениями в процессах фотосинтеза (см. выше) и транспирации . Последний процесс (на который приходится около 90% использования воды) обусловлен испарением воды из листьев растений. Транспирация позволяет растениям переносить воду и минеральные питательные вещества из почвы в регионы роста, а также охлаждает растение. Диффузия водяного пара из листа, сила, которая управляет транспирацией, регулируется структурами, известными как устьица . Эти структуры также регулируют диффузию углекислого газа из атмосферы в лист, так что уменьшение потери воды (за счет частичного закрытия устьиц) также снижает поступление углекислого газа. Некоторые растения используют альтернативные формы фотосинтеза, называемые метаболизмом крассуловой кислоты (CAM) и C4 . Они используют физиологические и анатомические адаптации для повышения эффективности использования воды и позволяют увеличить первичное производство в условиях, которые обычно ограничивают фиксацию углерода растениями C3 (большинство видов растений). [ нужна ссылка ]
Как показано на анимации, в бореальных лесах Канады и России наблюдается высокая продуктивность в июне и июле, а затем ее медленное снижение осенью и зимой. Круглый год тропические леса в Южной Америке, Африке, Юго-Восточной Азии и Индонезии имеют высокую продуктивность, что неудивительно, учитывая обилие солнечного света, тепла и осадков. Однако даже в тропиках продуктивность в течение года меняется. Например, бассейн Амазонки демонстрирует особенно высокую продуктивность примерно с августа по октябрь — период засушливого сезона в этом районе. Поскольку деревья имеют доступ к обильному запасу грунтовых вод, которые накапливаются в сезон дождей, они растут лучше, когда дождливое небо проясняется и позволяет большему количеству солнечного света достигать леса. [3]
Океаническая продукция
[ редактировать ]
На суше, в океанах, наоборот, почти весь фотосинтез осуществляется водорослями, с небольшой долей вклада сосудистых растений и других групп. Водоросли охватывают широкий спектр организмов: от одиночных плавающих клеток до прикрепленных морских водорослей . В их число входят фотоавтотрофы из самых разных групп. Эубактерии так и в наземных экосистемах, и хотя некоторые археи фототрофны являются важными фотосинтезирующими устройствами как в океанических , , ни одна из них, как известно, не использует фотосинтез с выделением кислорода. [4] Ряд эукариот вносят значительный вклад в первичную продукцию океана, включая зеленые , бурые и красные водоросли , а также разнообразную группу одноклеточных групп. Сосудистые растения также представлены в океане такими группами, как морские травы .
В отличие от наземных экосистем, большая часть первичной продукции в океане осуществляется свободноживущими микроскопическими организмами, называемыми фитопланктоном . Более крупные автотрофы, такие как морские травы и макроводоросли ( водоросли ), обычно обитают в прибрежной зоне и прилегающих мелководьях, где они могут прикрепляться к нижележащему субстрату, но при этом оставаться в пределах фотической зоны . Есть исключения, такие как Sargassum , но подавляющее большинство свободно плавающих производств происходит внутри микроскопических организмов.

Факторы, ограничивающие первичное производство в океане, также сильно отличаются от факторов на суше. Доступность воды, очевидно, не является проблемой (хотя ее соленость может быть проблемой). Точно так же температура, хотя и влияет на скорость метаболизма (см. вопрос 10 ), в океане варьируется не так широко, как на суше, поскольку теплоемкость морской воды смягчает изменения температуры, а образование морского льда изолирует ее при более низких температурах. Однако доступность света, источника энергии для фотосинтеза, и минеральных питательных веществ , строительных блоков для нового роста, играют решающую роль в регулировании первичной продукции в океане. [5] Доступные модели системы Земли предполагают, что продолжающиеся биогеохимические изменения океана могут вызвать сокращение NPP океана на 3–10% от текущих значений в зависимости от сценария выбросов. [6]
Свет
[ редактировать ]
Освещенная солнцем зона океана называется фотической зоной (или эвфотической зоной). Это относительно тонкий слой (10–100 м) у поверхности океана, где достаточно света для фотосинтеза. Для практических целей толщина фотозоны обычно определяется глубиной, на которой свет достигает 1% от его поверхностного значения. Свет ослабляется в толще воды за счет его поглощения или рассеяния самой водой, а также растворенными в ней материалами или твердыми частицами (включая фитопланктон).
Чистый фотосинтез в толще воды определяется взаимодействием между фотозоной и перемешанным слоем . Турбулентное перемешивание за счет энергии ветра на поверхности океана гомогенизирует толщу воды по вертикали до тех пор, пока турбулентность не рассеется (создавая вышеупомянутый смешанный слой). Чем глубже перемешанный слой, тем меньше среднее количество света, улавливаемого в нем фитопланктоном. Смешанный слой может варьироваться от более мелкого, чем фотическая зона, до более глубокого, чем фотическая зона. Когда он находится намного глубже, чем фотическая зона, это приводит к тому, что фитопланктон проводит слишком много времени в темноте, чтобы произошел чистый рост. Максимальная глубина смешанного слоя, на которой может произойти чистый рост, называется критической глубиной . Пока имеется достаточное количество питательных веществ, чистая первичная продукция происходит всякий раз, когда смешанный слой оказывается меньше критической глубины.
Как на величину смешивания ветров, так и на доступность света на поверхности океана влияют в различных пространственных и временных масштабах. Наиболее характерным из них является сезонный цикл (вызванный последствиями наклона Земли оси ), хотя величина ветра дополнительно имеет сильные пространственные компоненты . Следовательно, первичная продукция в регионах с умеренным климатом , таких как Северная Атлантика , носит сильно сезонный характер и зависит как от падающего света на поверхность воды (снижается зимой), так и от степени смешивания (увеличивается зимой). В тропических регионах, таких как круговороты в центре крупных бассейнов , свет может незначительно меняться в течение года, а смешивание может происходить лишь эпизодически, например, во время сильных штормов или ураганов .
Питательные вещества
[ редактировать ]
Смешивание также играет важную роль в ограничении первичного производства питательных веществ. Неорганические питательные вещества, такие как нитраты , фосфаты и кремниевая кислота , необходимы фитопланктону для синтеза клеток и клеточных механизмов. Из-за гравитационного опускания твердых частиц (например, планктона , мертвых или фекальных материалов) питательные вещества постоянно теряются из фотической зоны и пополняются только за счет смешивания или подъема более глубоких вод. Ситуация усугубляется там, где солнечное нагревание в летнее время и уменьшение ветров увеличивают вертикальную стратификацию и приводят к сильному термоклину , поскольку из-за этого смешиванию ветров становится труднее уносить более глубокие воды. Следовательно, между событиями смешивания первичная продукция (и возникающие в результате процессы, ведущие к опусканию сыпучего материала) постоянно потребляют питательные вещества в перемешанном слое, и во многих регионах это приводит к истощению питательных веществ и снижению производства смешанного слоя летом (даже в наличие обильного света). Однако, пока фотическая зона достаточно глубока, первичное производство может продолжаться ниже смешанного слоя, где скорость роста, ограниченная светом, означает, что питательных веществ часто бывает больше.
Железо
[ редактировать ]Еще одним фактором, относительно недавно обнаруженным и играющим значительную роль в первичном производстве океана, является микроэлемент железо . [7] Он используется в качестве кофактора в ферментах, участвующих в таких процессах, как восстановление нитратов и фиксация азота . Земли Основным источником железа в океанах является пыль из пустынь , поднимаемая и переносимая ветром в виде эоловой пыли .
В регионах океана, удаленных от пустынь или не добираемых пыльными ветрами (например, в южной и северной части Тихого океана), недостаток железа может серьезно ограничить объем первичной продукции, которая может иметь место. Эти области иногда называют регионами HNLC (с высоким содержанием питательных веществ и низким содержанием хлорофилла), поскольку нехватка железа одновременно ограничивает рост фитопланктона и оставляет избыток других питательных веществ. Некоторые ученые предложили внести в эти районы железо как средство повышения первичной продуктивности и улавливания углекислого газа из атмосферы. [8]
Измерение
[ редактировать ]Методы измерения первичной продукции различаются в зависимости от того, является ли валовая или чистая продукция желаемой мерой, и от того, находятся ли в центре внимания наземные или водные системы. Валовое производство почти всегда труднее измерить, чем чистое, из-за дыхания, которое представляет собой непрерывный и непрерывный процесс, в ходе которого некоторые продукты первичного производства (например, сахара) потребляются до того, как их можно будет точно измерить. Кроме того, наземные экосистемы, как правило, более сложны, поскольку значительная часть общей продуктивности перемещается в подземные органы и ткани, где ее трудно измерить с точки зрения логистики. Мелководные водные системы также могут столкнуться с этой проблемой.
Масштаб также сильно влияет на методы измерения. Скорость ассимиляции углерода в растительных тканях, органах, целых растениях или образцах планктона можно определить количественно с помощью биохимических методов , но эти методы явно не подходят для крупномасштабных наземных полевых ситуаций. Там чистая первичная продукция почти всегда является желательной переменной, а методы оценки включают в себя различные методы оценки изменений биомассы в сухом весе с течением времени. Оценки биомассы часто преобразуются в энергетические меры, такие как килокалории, с помощью эмпирически определенного коэффициента преобразования.
Земной
[ редактировать ]
В наземных экосистемах исследователи обычно измеряют чистую первичную продукцию (ЧПП). Хотя его определение простое, полевые измерения, используемые для оценки продуктивности, различаются в зависимости от исследователя и биома. Полевые оценки редко учитывают подземную продуктивность, травоядность, оборот, опад , летучие органические соединения , корневые выделения и распределение по симбиотическим микроорганизмам. Оценки АЭС на основе биомассы приводят к занижению АЭС из-за неполного учета этих компонентов. [9] [10] Однако многие полевые измерения хорошо коррелируют с АЭС. Существует ряд всесторонних обзоров полевых методов, используемых для оценки АЭС. [9] [10] [11] Оценки дыхания экосистемы , общего количества углекислого газа, производимого экосистемой, также можно сделать с помощью измерений газовых потоков .
Основным неучтенным резервом является подземная продуктивность, особенно производство и оборот корней. Подземные компоненты АЭС трудно измерить. ПАЭС (подземная АЭС) часто оценивается на основе соотношения АНЭС: ПАЭС (надземная АЭС:подземная АЭС), а не на основе прямых измерений.
Валовую первичную продукцию можно оценить на основе измерений чистого экосистемного обмена (NEE) углекислого газа, выполненных методом вихревой ковариации . Ночью этот метод измеряет все компоненты дыхания экосистемы. Это дыхание масштабируется до дневных значений и далее вычитается из NEE. [12]
Луга
[ редактировать ]
Чаще всего предполагается, что для измерения NPP используется пиковая стоячая биомасса. В системах с постоянной подстилкой обычно сообщается о живой биомассе. Измерения пиковой биомассы более надежны, если система преимущественно однолетняя. Однако многолетние измерения могли бы быть надежными, если бы существовала синхронная фенология, обусловленная сильным сезонным климатом. Эти методы могут занижать оценку ANPP на лугах в 2 ( умеренный климат ) или 4 ( тропический климат ) раза. [10] Повторные измерения стоячей живой и мертвой биомассы позволяют более точно оценить все пастбища, особенно те, которые имеют большой оборот, быстрое разложение и межвидовые различия во времени достижения пика биомассы. Продуктивность водно-болотных угодий (болот и болот) измеряется аналогичным образом. В Европе ежегодное скашивание делает очевидным ежегодный прирост биомассы водно-болотных угодий.
Леса
[ редактировать ]
Методы, используемые для измерения продуктивности лесов , более разнообразны, чем методы измерения продуктивности лугов. конкретного древостоя Прирост биомассы, основанный на аллометрии плюс опаде, считается подходящим, хотя и неполным, расчетом надземной чистой первичной продукции (ANPP). [9] Полевые измерения, используемые в качестве показателя для ANPP, включают годовое количество опада, прирост диаметра или площади основания ( DBH или BAI), а также прирост объема.
Водный
[ редактировать ]| Часть серии о |
| Углеродный цикл |
|---|
 |
В водных системах первичная продукция обычно измеряется с использованием одного из шести основных методов: [13]
- изменения концентрации кислорода в запечатанной бутылке (разработаны Гардером и Граном в 1927 году)
- включение неорганического углерода-14 ( 14 С в виде гидрокарбоната натрия ) в органическое вещество [14] [15]
- Стабильные изотопы кислорода ( 16 ТЕМ, 18 О и 17 ТЕМ) [16] [17]
- кинетика флуоресценции (метод все еще остается темой исследования)
- Стабильные изотопы углерода ( 12 С и 13 С) [18]
- Соотношение кислород/аргон [19]
Методика, разработанная Гардером и Граном, использует изменения концентрации кислорода в различных экспериментальных условиях для определения валового первичного производства. Обычно три одинаковых прозрачных сосуда заполняются пробой воды и закрываются пробками . Первый анализируется сразу и используется для определения начальной концентрации кислорода; обычно это делается путем титрования по Винклеру . Два других сосуда инкубируют, по одному на свету и затемнено. По истечении установленного периода времени эксперимент заканчивают и измеряют концентрацию кислорода в обоих сосудах. Поскольку фотосинтез не происходит в темном сосуде, он обеспечивает определенное дыхание экосистемы . Легкий сосуд обеспечивает как фотосинтез, так и дыхание, поэтому обеспечивает меру чистого фотосинтеза (т.е. производство кислорода посредством фотосинтеза вычитает потребление кислорода при дыхании). Затем валовую первичную продукцию получают путем прибавления потребления кислорода в темном сосуде к чистому производству кислорода в светлом сосуде.
Техника использования 14 Включение C (добавленного как Na 2 CO 3 ) для вывода о первичном производстве сегодня наиболее часто используется, поскольку оно чувствительно и может использоваться во всех океанских средах. Как 14 C радиоактивен (в результате бета-распада ), его содержание в органическом материале относительно легко измерить с помощью таких устройств, как сцинтилляционные счетчики .
В зависимости от выбранного времени инкубации можно оценить чистую или валовую первичную продукцию. Валовую первичную продукцию лучше всего оценивать, используя относительно короткое время инкубации (1 час или менее), поскольку потеря инкорпорированной 14 C (при дыхании и выделении/экссудации органических веществ) будет более ограниченным. Чистая первичная продукция — это доля валового производства, оставшаяся после того, как в результате этих процессов потерь была израсходована часть фиксированного углерода.
Процессы потерь могут составлять от 10 до 60 % от суммы включенных 14 C в зависимости от инкубационного периода, условий окружающей среды (особенно температуры) и используемых экспериментальных видов . Помимо тех, которые вызваны физиологией самого подопытного, необходимо учитывать и потенциальные потери, связанные с активностью потребителей. Это особенно верно в экспериментах с использованием природных сообществ микроскопических автотрофов, когда невозможно изолировать их от потребителей.
Преимущество методов, основанных на стабильных изотопах и отношениях O 2 /Ar, заключается в том, что они позволяют оценить скорость дыхания на свету без необходимости инкубации в темноте. Среди них метод тройных изотопов кислорода и O 2 /Ar имеет дополнительное преимущество, заключающееся в том, что он не требует инкубации в закрытых контейнерах, а O 2 /Ar можно даже непрерывно измерять в море с использованием масс-спектрометрии на входе в эквилибратор (EIMS). [20] или масс-спектрометрию с мембранным входом (MIMS). [21] Однако, если желательны результаты, относящиеся к углеродному циклу, вероятно, лучше полагаться на методы, основанные на изотопах углерода (а не кислорода). Важно отметить, что метод, основанный на стабильных изотопах углерода, не является просто адаптацией классического метода. 14 Метод С, но совершенно другой подход, не страдающий проблемой отсутствия учета рециркуляции углерода при фотосинтезе.
Глобальный
[ редактировать ]Поскольку первичное производство в биосфере является важной частью углеродного цикла , его оценка в глобальном масштабе важна для науки о системе Земли . Однако количественная оценка первичной продукции в этом масштабе затруднена из-за разнообразия сред обитания на Земле, а также из-за влияния погодных явлений (наличия солнечного света, воды) на ее изменчивость. Используя спутниковые оценки индекса нормализованной разницы растительности морской поверхности (NDVI) для наземных сред обитания и хлорофилла для океанов, было подсчитано, что общая (фотоавтотрофная) первичная продукция на Земле составила 104,9 петаграмм углерода в год (Pg C год −1 ; эквивалент не- SI Gt C yr −1 ). [22] Из них 56,4 Пг С/год. −1 (53,8%), являлся продуктом деятельности наземных организмов, а остальные 48,5 Пг С/год −1 , приходилось на океаническую продукцию.
Масштабирование оценок GPP на уровне экосистемы, основанных на измерениях вихревой ковариации чистого экосистемного обмена (см. выше) до региональных и глобальных значений с использованием пространственных деталей различных переменных-предсказателей, таких как климатические переменные и данные дистанционного зондирования fAPAR или LAI, привело к валовой первичной продукции на суше 123±8 Гт углерода (НЕ углекислого газа) в год в течение 1998-2005 гг. [23]
В площадном выражении было подсчитано, что продуктивность земли составила примерно 426 г См. −2 тот −1 (без учета районов с постоянным ледяным покровом), а для океанов — 140 г См. −2 тот −1 . [22] Еще одно существенное различие между сушей и океаном заключается в их постоянных запасах: на долю океанических автотрофов приходится почти половина общей продукции, но на их долю приходится лишь около 0,2% от общей биомассы.
Текущие и прошлые оценки
[ редактировать ]Современную первичную продуктивность можно оценить с помощью различных методологий, включая измерения с борта корабля, спутников и наземных обсерваторий. Исторические оценки основывались на биогеохимических моделях и геохимических показателях. Одним из примеров является использование бария , где концентрация барита в морских отложениях увеличивается вместе с экспортом углерода на поверхность. [24] [25] [26] Другой пример — использование кислорода сульфата тройных изотопов . [27] [28] [29] В совокупности эти записи предполагают большие сдвиги в первичном производстве на протяжении всего прошлого Земли, причем заметный рост связан с Великим событием окисления Земли (приблизительно 2,4–2,0 миллиарда лет назад) и неопротерозоем (приблизительно 1,0–0,54 миллиарда лет назад). [29]
Человеческое воздействие и присвоение
[ редактировать ]Человеческие общества являются частью земного АЭС-цикла, но оказывают в нем непропорционально большое влияние. [30] В 1996 году Хосеп Гари разработал новый индикатор устойчивого развития, основанный именно на оценке присвоения человеком АЭС: он придумал его «HANPP» (человеческое присвоение чистой первичной продукции) и представил его на первой конференции Европейского общества Экологическая экономика. [31] С тех пор HANPP получил дальнейшее развитие и широко применяется в исследованиях в области экологической экономики, а также в анализе политики устойчивого развития. HANPP представляет собой показатель воздействия человека на природу и может применяться в различных географических масштабах, а также во всем мире.
Широкая степень использования человеком ресурсов планеты, в основном посредством землепользования , приводит к различным уровням воздействия на реальную АЭС ( Закон об АЭС ). Хотя в некоторых регионах, таких как долина Нила , орошение привело к значительному увеличению первичной продукции, на большей части планеты наблюдается заметная тенденция сокращения NPP из-за изменений земель (ΔNPP LC ) на 9,6% по всей мировой территории. масса. [32] В дополнение к этому, конечное потребление людьми увеличивает общий объем HANPP. [30] до 23,8% потенциальной растительности (ЧПП 0 ). [32] Подсчитано, что в 2000 году 34% свободной ото льда территории Земли (12% пахотных земель ; 22% пастбищ ) было отведено под сельское хозяйство. [33] Это непропорциональное количество уменьшает количество энергии, доступной другим видам, оказывая заметное воздействие на биоразнообразие , потоки углерода, воды и энергии, а также экосистемные услуги . [32] и ученые задаются вопросом, насколько велика эта доля, прежде чем эти услуги начнут разрушаться. [34] Ожидается также сокращение NPP в океане в результате продолжающегося изменения климата, что потенциально может повлиять на морские экосистемы (~ 10% глобального биоразнообразия), а также на товары и услуги (1-5% от общего объема мирового биоразнообразия), которые обеспечивают океаны. [6]
См. также
[ редактировать ]- Биологический насос
- Биомасса (экология)
- f-коэффициент
- Производительность
- Сероводород (H 2 S)
- Списки организмов по численности населения
Ссылки
[ редактировать ]- ^ Амтор, Дж. С. и Бальдокки, Д. Д. (2001). Дыхание наземных высших растений и чистая первичная продукция. В земной глобальной продуктивности , Academic Press, 33-59.
- ^ Сигман, DM; Хейн, член парламента (2012). «Биологическая продуктивность океана» (PDF) . Знания о природном образовании . 3 (6): 1–16 . Проверено 1 июня 2015 г.
«Валовая первичная продукция» (GPP) относится к общей скорости производства органического углерода автотрофами, а «дыхание» относится к энергетическому окислению органического углерода обратно в углекислый газ. «Чистая первичная продукция» (NPP) равна GPP минус собственная скорость дыхания автотрофов; таким образом, это скорость, с которой полный метаболизм фитопланктона производит биомассу. «Вторичная продукция» (СП) обычно относится к скорости роста гетеротрофной биомассы.
- ^ Jump up to: а б «Чистая первичная производительность: глобальные карты» . Earthobservatory.nasa.gov . 26 марта 2018 года . Проверено 26 марта 2018 г.
- ^ Шефер Г., Энгельхард М., Мюллер В. (1 сентября 1999 г.). «Биоэнергетика архей» . Микробиол. Мол. Биол. Преподобный . 63 (3): 570–620. дои : 10.1128/MMBR.63.3.570-620.1999 . ПМЦ 103747 . ПМИД 10477309 .
- ^ Сигман, DM; Хейн, член парламента (2012). «Биологическая продуктивность океана» (PDF) . Знания о природном образовании . 3 (6): 1–16 . Проверено 1 июня 2015 г.
Глубокий максимум хлорофилла (DCM) возникает там, где достаточно света для фотосинтеза и при этом имеется значительный запас питательных веществ снизу.
- ^ Jump up to: а б Мора, К.; и др. (2013). «Биотическая и человеческая уязвимость к прогнозируемым изменениям в биогеохимии океана в 21 веке» . ПЛОС Биология . 11 (10): e1001682. дои : 10.1371/journal.pbio.1001682 . ПМК 3797030 . ПМИД 24143135 .
- ^ Мартин, Дж. Х.; Фицуотер, ЮВ (1988). «Дефицит железа ограничивает рост фитопланктона в субарктике северо-восточной части Тихого океана». Природа . 331 (6154): 341–343. Бибкод : 1988Natur.331..341M . дои : 10.1038/331341a0 . S2CID 4325562 .
- ^ Купер, диджей; Уотсон, Эй Джей; Найтингейл, П.Д. (1996). на поверхности океана «Значительное снижение летучести CO 2 в ответ на внесение железных удобрений на месте ». Природа . 383 (6600): 511–513. Бибкод : 1996Natur.383..511C . дои : 10.1038/383511a0 . S2CID 4325717 .
- ^ Jump up to: а б с Кларк, округ Колумбия; Браун, С.; Киклайтер, Д.В.; Чемберс, JQ; Томлинсон-младший; Ни, Дж. (2001). «Измерение чистой первичной продукции в лесах: концепции и полевые методы». Экологические приложения . 11 (2): 356–370. doi : 10.1890/1051-0761(2001)011[0356:MNPPIF]2.0.CO;2 . ISSN 1051-0761 . S2CID 37718097 .
- ^ Jump up to: а б с Скарлок, JMO; Джонсон, К.; Олсон, Р.Дж. (2002). «Оценка чистой первичной продуктивности на основе измерений динамики биомассы пастбищ» . Биология глобальных изменений . 8 (8): 736–753. Бибкод : 2002GCBio...8..736S . дои : 10.1046/j.1365-2486.2002.00512.x . S2CID 84344715 .
- ^ Лейт, Х.; Уиттакер, Р.Х. (1975). Первичная продуктивность биосферы . Нью-Йорк: Springer-Verlag . ISBN 978-0-387-07083-4 .
- ^ Райхштейн, Маркус; Фальге, Ева; Бальдокки, Деннис; Папале, Дарио; Обине, Марк; Бербижье, Поль; и др. (2005). «О разделении чистого экосистемного обмена на ассимиляцию и дыхание экосистемы: обзор и усовершенствованный алгоритм». Биология глобальных изменений . 11 (9): 1424–1439. Бибкод : 2005GCBio..11.1424R . дои : 10.1111/j.1365-2486.2005.001002.x . ISSN 1354-1013 . S2CID 55932276 .
- ^ Марра, Дж. (2002), стр. 78-108. В: Уильямс, П. Дж. ЛеБ., Томас, Д. Н., Рейнольдс, К. С. (ред.), Продуктивность фитопланктона: ассимиляция углерода в морских и пресноводных экосистемах. Блэквелл, Оксфорд, Великобритания
- ^ Стиман-Нильсен, Э. (1951). «Измерение продукции органического вещества в море с помощью углерода-14». Природа . 167 (4252): 684–685. Бибкод : 1951Natur.167..684N . дои : 10.1038/167684b0 . ПМИД 14826912 . S2CID 4299572 .
- ^ Стиман-Нильсен, Э. (1952). «Использование радиоактивного углерода (C14) для измерения органической продукции в море». Дж. Минусы. Межд. Исследовать. Мер . 18 (2): 117–140. дои : 10.1093/icesjms/18.2.117 .
- ^ Бендер, Майкл; и др. (1987). «Сравнение 4 методов определения производства планктонного сообщества». Лимнология и океанография . 32 (5): 1085–1098. Бибкод : 1987LimOc..32.1085B . дои : 10.4319/lo.1987.32.5.1085 .
- ^ Луз и Баркан, Б; Баркан, Э (2000). «Оценка продуктивности океана по трехизотопному составу растворенного кислорода». Наука . 288 (5473): 2028–2031. Бибкод : 2000Sci...288.2028L . дои : 10.1126/science.288.5473.2028 . ПМИД 10856212 .
- ^ Карвальо и Эйр (2012). «Измерение планктонного дыхания СО2 на свету». Лимнология и океанография: Методы . 10 (3): 167–178. дои : 10.4319/лом.2012.10.167 . S2CID 93847401 .
- ^ Крейг и Хейворд (1987). «Перенасыщение кислородом в океане: биологический и физический вклад». Наука . 235 (4785): 199–202. Бибкод : 1987Sci...235..199C . дои : 10.1126/science.235.4785.199 . ПМИД 17778634 . S2CID 40425548 .
- ^ Кассар, Н.; Б. А. Барнетт; М.Л. Бендер; Дж. Кайзер; РЦ Хамме; Б. Тилбрук (2009). «Непрерывные высокочастотные измерения растворенного O 2 /Ar с помощью масс-спектрометрии на входе эквилибратора». Анальный. Хим . 81 (5): 1855–1864. дои : 10.1021/ac802300u . ПМИД 19193192 .
- ^ Кайзер, Дж.; МК Ройер; Б. Барнетт; М.Л. Бендер (2005). «Оценки продуктивности морских судов на основе непрерывных измерений соотношения O-2/Ar с помощью масс-спектрометрии с мембранным входом». Геофиз. Рез. Летт . 32 (19): н/д. Бибкод : 2005GeoRL..3219605K . CiteSeerX 10.1.1.584.1430 . дои : 10.1029/2005GL023459 . S2CID 53465452 .
- ^ Jump up to: а б Поле, CB; Беренфельд, МЮ; Рандерсон, Джей Ти; Фальковски, П. (1998). «Первичная продукция биосферы: интеграция наземных и океанических компонентов» . Наука . 281 (5374): 237–240. Бибкод : 1998Sci...281..237F . дои : 10.1126/science.281.5374.237 . ПМИД 9657713 .
- ^ Бир, К.; Райхштейн, М.; Томеллери, Э.; Сиас, П.; Юнг, М.; Карвалье, Н.; и др. (2010). «Валовое поглощение углекислого газа на Земле: глобальное распределение и зависимость от климата» (PDF) . Наука . 329 (5993): 834–838. Бибкод : 2010Sci...329..834B . дои : 10.1126/science.1184984 . ISSN 0036-8075 . ПМИД 20603496 . S2CID 206524740 .
- ^ Пракаш Бабуа, К.; Брамсак, Х.-Дж.; Шнетгер, Б.; Бетчер, Мэн (2002). «Барий как показатель продуктивности отложений континентальных окраин: исследование восточной части Аравийского моря». Морская геология . 184 (3–4): 189–206. Бибкод : 2002МГеол.184..189П . дои : 10.1016/S0025-3227(01)00286-9 .
- ^ Э.Д. Гольдберг, ГОС Аррениус Химия пелагических отложений Geochim. Космохим. Acta , 13 (1958), стр. 153–212.
- ^ Даймонд, Джек; Зюсс, Эрвин; Лайл, Митч (1992). «Барий в глубоководных отложениях: геохимический показатель палеопродуктивности» (PDF) . Палеоокеанография . 7 (2): 163–181. Бибкод : 1992PalOc...7..163D . дои : 10.1029/92PA00181 . S2CID 130057872 .
- ^ Крокфорд, Питер В.; Хейлс, Джастин А.; Бао, Хуэймин; Планавский, Ной Дж.; Беккер, Андрей; Фралик, Филип В.; Халверсон, Гален П.; Буй, Тхи Хао; Пэн, Юнбо; Винг, Босуэлл А. (июль 2018 г.). «Тройной изотоп кислорода свидетельствует об ограниченной первичной продуктивности в середине протерозоя» . Природа . 559 (7715): 613–616. дои : 10.1038/s41586-018-0349-y . ISSN 1476-4687 .
- ^ Крокфорд, Питер В.; Кунцманн, Маркус; Беккер, Андрей; Хейлс, Джастин; Бао, Хуэймин; Халверсон, Гален П.; Пэн, Юнбо; Буй, Тхи Х.; Кокс, Грант М.; Гибсон, Тимоти М.; Верндле, Сара; Рейнберд, Роберт; Лепланд, Айво; Суонсон-Хайсел, Николас Л.; Мастер, Шарад (20 мая 2019 г.). «Клейпул продолжил: Расширение изотопной записи осадочных сульфатов» . Химическая геология . 513 : 200–225. doi : 10.1016/j.chemgeo.2019.02.030 . ISSN 0009-2541 .
- ^ Jump up to: а б Крокфорд, Питер В.; Бар Он, Инон М.; Уорд, Люс М.; Майло, Рон; Халеви, Италия (ноябрь 2023 г.). «Геологическая история первичной продуктивности» . Современная биология . 33 (21): 4741–4750.e5. дои : 10.1016/j.cub.2023.09.040 . ISSN 0960-9822 .
- ^ Jump up to: а б Витоусек, премьер-министр; Эрлих, PR; Эрлих, А.Х.; Мэтсон, Пенсильвания (1986). «Присвоение человеком продуктов фотосинтеза» (PDF) . Бионаука . 36 (6): 368–373. дои : 10.2307/1310258 . JSTOR 1310258 . Архивировано из оригинала (PDF) 5 марта 2011 г.
- ^ Гари, JA (1996). «ГАЭС рассчитана на основе земельного покрова как индикатор экологической устойчивости». Материалы международной конференции: Экология, общество, экономика : 1–18. hdl : 10419/183215 .
- ^ Jump up to: а б с Хаберл, Х.; Эрб, К.Х.; Краусманн, Ф.; Гаубе, В.; Бондо, А.; Плуцар, К.; Гингрич, С.; Лухт, В.; Фишер-Ковальски, М. (2007). «Количественная оценка и картирование присвоения человеком чистой первичной продукции в наземных экосистемах» . Учеб. Натл. акад. наук. США 104 (31): 12942–12947. Бибкод : 2007PNAS..10412942H . дои : 10.1073/pnas.0704243104 . ЧВК 1911196 . ПМИД 17616580 .
- ^ Раманкутти, Н.; Эван, AT; Монфреда, К.; Фоли, Дж. А. (2008). «Земледелие на планете: 1. Географическое распределение мировых сельскохозяйственных земель в 2000 году» . Глобальные биогеохимические циклы . 22 (1): GB1003. Бибкод : 2008GBioC..22.1003R . дои : 10.1029/2007GB002952 . S2CID 128460031 .
- ^ Фоли, Дж.А.; Монфреда, К.; Раманкутти, Н.; Закс, Д. (2007). «Наша доля планетарного пирога» . Учеб. Натл. акад. наук. США 104 (31): 12585–12586. Бибкод : 2007PNAS..10412585F . дои : 10.1073/pnas.0705190104 . ЧВК 1937509 . ПМИД 17646656 .