Эхинодон
| Эхинодон Временной диапазон: ранний мел ,
| |
|---|---|

| |
| Стоматология паратипического экземпляра NHMUK 48215b | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Динозаврия |
| Клэйд : | † Орнитишия |
| Семья: | † Гетеродонтозавриды |
| Род: | † Эхинодон Оуэн , 1861 год. [ 1 ] |
| Разновидность: | † Е. becklesii
|
| Биномиальное имя | |
| † Эхинодон becklesii Оуэн, 1861 г. [ 1 ]
| |
Echinodon — род динозавров гетеродонтозаврид - , живших в раннем меловом периоде на юге Англии и, возможно, на западе Франции в берриасскую эпоху. Первыми экземплярами были кости челюсти, названные Echinodon becklesii сэром Ричардом Оуэном в 1861 году, и с момента их первоначального описания были обнаружены только дополнительные зубы. Видовое название дано в честь коллекционера Сэмюэля Беклза , который обнаружил материал Echinodon и многих других таксонов со всей Англии, а название рода переводится как «колючий зуб» в связи с анатомией зубов этого таксона.
Первоначально Echinodon считался разновидностью травоядных ящериц, но быстро превратился в промежуточного птицетазового вида. Его отнесли к кладе Stegosauria на основании общей анатомии зубов и ошибочно отнесенного панциря, который позже был идентифицирован как черепаховый . Затем эхинодон был отнесен к раннему птицетазовому семейству Fabrosauridae , которое позднее было идентифицировано как искусственная группа, в которой эхинодоны были отнесены к Heterodontosauridae . Первоначально это семейство считалось наиболее близким к более развитым орнитоподам , но в конечном итоге оно было повторно идентифицировано как самая базальная группа птицетазовых, что сделало Echinodon таксоном, произошедшим от многих родов ранней юры , с призрачной линией 50 миллионов лет несохранившейся эволюции. .
Все экземпляры Echinodon были найдены в группе Пурбек в Дорсете , которая по-разному считается происходящей из позднего юрского или самого раннего мелового периода. Текущие исследования допускают раннемеловой берриазский возраст, что делает Echinodon одновременно самым молодым и самым маленьким гетеродонтозавридом. Другие динозавры, которые жили рядом с ним, включают орнитопода Owenodon и теропода Nuthetes , которые также являются фрагментарными. также обитало множество мелких млекопитающих Рядом с ехинодонами , и отложения показывают, что группа Пурбек представляла собой лагунную среду с различной лагуной, первоначально похожую на современное Средиземноморье , но со временем она стала более влажной.
История открытия
[ редактировать ]
Множественные образцы костей челюсти были обнаружены Сэмюэлем Беклзом высоко на скале в заливе Дердлстоун на острове Пурбек на юге Англии . Эти окаменелости, в том числе множество зубов , а также кости верхней и нижней челюстей , были найдены рядом с раковинами и окаменелостями растений в слоях Пурбек . Впервые они были описаны в монографии, опубликованной в 1861 году сэром Ричардом Оуэном , британским палеонтологом , который также описал окаменелости игуанодона и мегалозавра . Оуэн дал окаменелостям название Echinodon becklesii , которые он считал частью клады ящериц Lacertilia . [ 1 ] В то время как видовое название было дано в честь Беклза за его открытие окаменелостей и позволило Оуэну изучить свою коллекцию окаменелостей Пурбека, общее название произошло от древнегреческого εχινος , «еж», и ὀδών , «зуб», которые Оуэн объединил как « «колючий зуб», чтобы описать анатомию зубцов по бокам зубов. Оуэн переписывался с британским палеонтологом Хью Фалконером , который предложил название «Sauraechinodon», но поскольку сокращенная форма Echinodon не была озабочена , Оуэн решил использовать сокращенную форму в качестве названия для своего нового животного. [ 1 ] В 1861 году Фальконер внес исправление, указав, что он предложил название «Саураехмодон» вместо «Саураехинодон». [ 2 ]
Хотя первоначально Оуэн описал его как lacertilian, в 1874 году он пересмотрел свою классификацию , сгруппировав Echinodon со Scelidosaurus и Iguanodon в кладу, которую он назвал Prionodontia , которая входила в более крупную кладу Dinosauria . [ 3 ] В 1888 году британский палеонтолог Ричард Лидеккер последовал предыдущей классификации Echinodon как динозавра, основываясь на анатомии его зубов, описав их как сходные со Scelidosaurus, но не отнеся их к более специфической кладе, чем Dinosauria indeterminate. Серия экземпляров, обозначенных Оуэном как типы, была куплена Британским музеем естественной истории (BMNH, ныне NHMUK) в 1876 году и является частью коллекции Беклза с номерами образцов от NHMUK от 48209 до 48215. [ 4 ] Лидеккер также уточнил, что Echinodon был обнаружен в слоях Среднего Пурбека, [ 4 ] неофициальное подразделение современной группы известняков Пурбек , официальное название исторических пластов Пурбек. [ 5 ] британский палеонтолог Питер Гальтон сузил место отложения Echinodon В 1978 году до пресноводного «грязного пласта». [ 6 ] также известная как «Яма млекопитающих», раскопанная Беклсом в 1857 году . [ 5 ] Однако не существует никаких доказательств связи Echinodon с каким-либо конкретным пластом формации Лулворт группы Пурбек. [ 7 ] Гальтон также отнес экземпляр NHMUK 48229 к Echinodon , фрагментарной зубной кости с зубами. [ 6 ] и единственное дальнейшее отнесение к этому роду включает отдельные зубы также из слоев Пурбека. [ 8 ]

В конце 1970-х и начале 1980-х годов раскопки Музея естественной истории округа Лос-Анджелес обнаружили множество мелких окаменелостей птицетазовых в Фрута палеонтологической области в Колорадо . Возраст этих останков, собранных в песчаниках у основания пачки Браши-Бейсин , формации Моррисон составлял приблизительно 150,2-150,3 миллиона лет, и их коллекционер Джордж Каллистон в 1984 году первоначально описал их как промежуточные останки фабросавра . Три года спустя Каллистон пересмотрел свое описание и передал материал, включая кости челюсти, позвонки и большую часть задних конечностей, Echinodon sp., задание, поддержанное Гальтоном в 2002 году , хотя в 2006 году он пересмотрел материал и заметил различия в анатомии зубов от Собственно эхинодон . В 2009 году этим окаменелостям был присвоен собственный род Fruitadens haagarorum таксон , родственный, но отличный от Echinodon . [ 9 ]
два предчелюстных зуба, приписываемые ехинодону . были обнаружены В 2021 году из костного пласта Анжак-Шаранта во Франции, который является ровесником отложений Пурбек, [ 10 ]
Описание
[ редактировать ]
Известный материал Echinodon ограничен костями черепа, но многочисленные экземпляры включают по крайней мере некоторые предчелюстные , верхнечелюстные , слезные , скуловые , небные , эктоптеригоидные и зубные кости , а также большинство зубов как верхней, так и нижней челюстей. Судя по пропорциям родственного рода Heterodontosaurus , череп Echinodon имел длину 62 мм (2,4 дюйма), что сопоставимо с черепом Tianyulong с длиной 66 мм (2,6 дюйма), но меньше, чем у взрослых Fruitadens с длиной 75 мм (3,0 дюйма). , что делает Эхинодона самым маленьким предположительно взрослым гетеродонтозавром и одним из самых маленьких нептичьих динозавров . [ 7 ]
основная часть обеих предчелюстных костей сохранилась У лектотипического экземпляра Echinodon , хотя и сломана и раздавлена. Предчелюстное отверстие присутствует возле переднего края отдельной кости, а ямка , в которой оно расположено, по форме больше похожа на Heterodontosaurus , чем на более развитый Hypsilophodon . Поверхность передней части кости вдоль зубного ряда текстурированная и лишена зубов, что отличает кость от кости лесотозавра , у которой зубы есть по всей длине. На предчелюстной кости было всего три зуба без зубчиков (зазубрин), присутствующих у большинства базальных птицетазовых, хотя все зубы примерно одинакового размера, причем третий является самым крупным. [ 7 ]
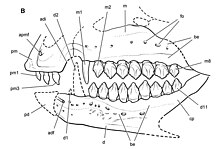
Среди материала Echinodon сохранились три верхнечелюстные кости , сохранившие почти всю кость, включая большинство участков соприкосновения с другими костями черепа. Верхняя челюсть слегка уплощена, что сводит к минимуму силу щечной выемки - диагностического признака птицетазовых, у которых верхнечелюстной зубной ряд и кость непосредственно над ним вставлены от наружного края кости. Наличие дугообразной диастемы у Echinodon является предметом разногласий. [ 7 ] В то время как Гальтон и американский палеонтолог Пол Серено интерпретируют диастему как присутствующую у гетеродонтозавра , [ 6 ] [ 11 ] Британские палеонтологи Дэвид Б. Норман и Пол М. Барретт в 2002 году пришли к выводу, что, основываясь на изображениях Оуэна до более позднего повреждения окаменелости NHMUK 48209, диастема отсутствовала. [ 8 ] Серено подтвердил В 2012 году , что имеется диастема, выгнутая на основании NHMUK 48211 верхней челюсти. [ 7 ] имеется девять зубов В верхней челюсти Echinodon , первый из которых представляет собой увеличенный и тонкий клык, похожий на те, что наблюдаются в предчелюстной кости Lycorhinus и Heterodontosaurus . [ 7 ] Оуэн также обнаружил частичный зуб перед клыковидным, но с тех пор он был утерян из-за повреждения материала. Хотя Норман и Барретт использовали это как доказательство существования второго собачьего существа, меньшего, чем предыдущее, [ 8 ] Гальтон в 1978 году и Серено в 2012 году идентифицировали только одну собаку. [ 6 ] [ 7 ] Первый пост-клыковидный зуб ехинодона является самым крупным, хотя и лишь немного выше следующих, которые имеют одинаковый размер. В середине коронок зубов имеется круглый выступ, но на коронке нет выступающих гребней. На каждой стороне коронок зубов имеется от восьми до десяти зубчиков. [ 7 ]
Слезные, скуловые и небные кости сохранились незначительно, хотя край орбиты . на фрагменте слезной кости можно идентифицировать [ 7 ] Эктоптеригоидная кость неба частично цела и сохранилась в сочленении с верхней челюстью NHMUK 48210. Отсутствие сохранности ограничивает анатомические детали, которые можно идентифицировать, помимо ее сходства с костью, известной у других птицетазовых животных. [ 7 ]

Предзубная . не сохранилась кость у Echinodon , но ее наличие можно подтвердить по морфологии переднего конца известных зубных костей Как и у других гетеродонтозаврид, предзубная часть при жизни лишь слабо сочленялась с зубной, у нее отсутствовали боковые и вентральные отростки. Зубная кость для базального птицетазового животного была глубокой, ее высота в середине длины составляла 30% от общей длины. Кость сужается спереди по всей длине, хотя края субпараллельны под серединой зубного ряда. ряд отверстий По краю щечной выемки, как и на верхней челюсти, имеется . Венечный отросток выражен, в отличие от других базальных птицетазовых, хотя контакт зубных костей V-образный, как и у базальных птицетазовых. [ 7 ] В зубной кости было 11 зубов, первые два из которых были специализированными по сравнению с большинством птицетазовых. [ 7 ] Хотя специализированные передние зубы не обсуждались Гальтоном в 1978 г., [ 6 ] и были признаны отсутствующими Норманом и Барреттом в 2002 году, [ 8 ] Серено описал первые две альвеолы Echinodon , а второй был значительно увеличен , как отличающиеся по размеру от следующих зубов в 2012 году. Первая альвеола была чрезвычайно уменьшена, что указывает на небольшой колышковидный первый зубной зуб, как у Lycorhinus что указывает на клыковидную форму, большую, чем у Lycorhinus. один в верхней челюсти. [ 7 ] Обычные зубные зубы были немного выше, чем зубы верхней челюсти, и имели зубцы вдоль верхней половины коронки, а не верхних 25%. Как и на верхней челюсти, коронки имеют медиальную выпуклость, восемь-десять зубчиков по обе стороны от вершины и симметричную эмаль . [ 7 ]
Классификация
[ редактировать ]Оуэн первоначально отнес эхинодона к травоядным ящерицам. [ 1 ] он изменил его размещение на место в Динозаврии. [ 3 ] Эхинодон считался промежуточным, потенциально птицетазовым динозавром, пока Франц Нопца не отнес его к кладе Scelidosaurinae внутри Stegosauridae в 1928 году на основании высокого венечного отростка и сужающихся зубов. [ 12 ] оба из которых считаются широко распространенными среди птицетазовых. [ 13 ] Изолированный кожный панцирь, обнаруженный в пластах Пурбек, был отнесен к эхинодонам на основании классификации стегозавров Джастина Делара в 1959 году . [ 14 ] хотя с тех пор они были переданы -солемидидам черепахам . [ 13 ] [ 15 ]

Классификации многих базальных птицетазовых животных были рассмотрены Ричардом Талборном в 1971 году , где он поместил эхинодона в группу Hypsilophodontidae : полученных от триасовых форм без клыков (называемых «фабросаврами»), близких к юрским родам Laosaurus и Nanosaurus и более примитивных, чем меловые роды Hypsilophodon. , паркзозавр и Тецелозавр . [ 16 ] В следующем году Гальтон опубликовал опровержение, не согласившись с использованием Тулборном Hypsilophodontidae. Поскольку зубы предчелюстной кости, один из диагностических признаков этого семейства, предложенных Тулборном, были примитивным признаком, Гальтон утверждал, что их не следует использовать для классификации орнитопод. Вместо этого Fabrosaurus и Echinodon были объединены в новое семейство Fabrosauridae на основе латерально расположенных зубов и удалены из Hypsilophodontidae. [ 17 ] Гальтон продолжил эту классификацию, назвав род Lesothosaurus в 1978 году, и вместе с его описанием рассмотрел анатомию нескольких базальных родов птицетазовых, включая Echinodon , Nanosaurus и Fabrosaurus . Все, кроме Echinodon, были окончательно отнесены им к Fabrosauridae, хотя место Echinodon было сомнительным, поскольку он также имел сходство с кладой Heterodontosauridae , которая, как известно, также обладала увеличенными клыками. Однако Гальтон считал принадлежность к фабросавридам более вероятной, поскольку у зубов этого таксона отсутствуют плоские фасетки стирания. [ 6 ]
Серено был первым, кто реклассифицировал Echinodon как члена семейства Heterodontosauridae. В 1991 году он пересмотрел черепную анатомию лесотозавра и сравнил его с большим количеством других базальных птицетазовых животных. Зубы Echinodon , по сути, были интерпретированы как имеющие плоские фасетки износа на зубах, а также зубы, вставленные от края верхней челюсти, как у птицетазовых, более продвинутых, чем у Lesothosaurus . Поскольку именно это было предложено объединить род с «фабросаврами», Серено считал Echinodon гетеродонтозавридом на основании наличия клыков и дугообразных промежутков в передней части зубного ряда. [ 18 ] гетеродонтозавр , абриктозавр и неописанная форма из формации Кайента . К семейству также были отнесены [ 11 ] Норман и Барретт заново описали Echinodon в 2002 году и поддержали классификацию гетеродонтозаврид, но вместо этого отнесли его к кладе на основании отсутствия отверстий на костях, несущих зубы, и наличия зубцов, ограниченных верхней третью коронки. [ 13 ] [ 8 ]
к гетеродонтозавридам Хотя отнесение Echinodon было подтверждено в дальнейших анализах, включая некоторые из первых филогенетических анализов птицетазовых, само положение семейства со временем изменилось. [ 19 ] Семейство Echinodon, либо исключая Echinodon , как в исследовании Тулборна 1971 года, либо включая этот род, как в работе Гальтона и Серено, первоначально считалось группой базальных орнитопод, более продвинутых, чем анкилозавры и стегозавры . [ 16 ] [ 17 ] [ 6 ] [ 18 ] [ 11 ] Дальнейшие исследования, в том числе обширный филогенетический анализ, проведенный британским палеонтологом Ричардом Дж. Батлером и его коллегами в 2008 году, подтвердили, что гетеродонтозавриды являются наиболее базальными птицетазами. [ 19 ] Пересмотренный анализ Батлера и др. в 2011 году также было решено базальное размещение гетеродонтозаврид и решены внутренние отношения в семье, при этом Echinodon является базальным родом в семье наряду с Abrictosaurus . [ 20 ] После дополнительного анализа, проведенного южноамериканским палеонтологом Диего Полом и его коллегами в 2011 году, Echinodon был признан базальным птицетазовым животным, но не принадлежащим к семейству Heterodontosauridae. Пол и др. считали, что размещение за пределами Heterodontosauridae произошло не потому, что он не принадлежал к семейству, а из-за неполноты его остатков. [ 21 ]
Кладограмма . ниже соответствует анализу Серено в 2012 году, включая правильно названные гетеродонтозавриды, описанные в то время Клада, включающая позднеюрских и раннемеловых Echinodon , Fruitadens и Tianyulong, плохо поддерживалась из-за некоторых особенностей зубов. [ 7 ]

Согласно решению анализа 2012 года, проведенного Серено, Echinodon и другие лавразийские гетеродонтозавриды были исключены из клады гондванских гетеродонтозаврид. Echinodon , Fruitadens , Tianyulong и неописанный гетеродонтозаврид Kayenta имеют зубы с низкой коронкой, в отличие от гондванских форм, а также дольчатую поясную кость и выступающую переднюю бороздку, ведущую к отверстию на боковой поверхности зубной кости. Все гетеродонтозавриды, кроме Kayenta, также значительно моложе остальных гетеродонтозаврид, и присутствие трех, объединенных в кладу, исключающую другие таксоны, предполагает неизвестную линию гетеродонтозаврид, которая существует 50 миллионов лет на протяжении юрского периода. Отсутствие данных об этой призрачной линии может быть связано с исключительно небольшим размером группы. [ 7 ]
Следуя более ранним результатам китайского палеонтолога Сюй Сина и его коллег в 2006 году , когда клада Heterodontosauriformes была создана для объединения гетеродонтозавров, цератопсов и пахицефалозавров , французский палеонтолог Поль-Эмиль Дьедонне и его коллеги предположили, что Echinodon и другие гетеродонтозавры были ранними формами пахицефалозавров. Их результаты поместили Echinodon в качестве родственного таксона таксонам, которые обычно классифицируются как Pachycephalosauria, за ним следует Tianyulong , в то время как все остальные изученные гетеродонтозавры образовали группу у истоков клады. Эта гипотеза позволит уменьшить призрачную линию пахицефалозавров и вернуть происхождение церапод к ранней юре. [ 22 ]
Палеобиология
[ редактировать ]Анатомия эхинодона и других гетеродонтозаврид плохо изучена; большинство палеобиологических аспектов основано на почти полном роде Heterodontosaurus . Фасеты износа на коронках Heterodontosaurus указывают на окклюзию верхней части зуба, в отличие от подразумеваемого движения челюсти у других форм, таких как Echinodon , где фасеты износа находятся только по бокам коронок. Несмотря на уникальное движение челюстей гетеродонтозавров по сравнению с более примитивными гетеродонтозавридами, функция увеличенных клыков, вероятно, была одинаковой во всей кладе. [ 7 ] По мнению Серено в 2012 году, основной функцией предчелюстного зубного ряда, исходя из наличия износа кончиков предчелюстной коронки и движения клыковидных зубов относительно других зубов при закрытии челюсти, является обрезка растительности. [ 7 ] Это контрастирует с более ранними гипотезами Батлера и соавт. в 2008 году выяснилось, что увеличенные клыки клады предназначены для всеядной диеты. [ 19 ] Гипотеза всеядности была также поддержана Норманом и его коллегами в 2011 году, утверждая, что клыки и предчелюстные зубы не изнашиваются в результате обрезки растительности. [ 23 ] участки Беззубые предчелюстных и предзубных костей, вероятно, были основой ороговатых клювов, как и у других птицетазовых. [ 7 ]
Экология
[ редактировать ]
Группа Пурбек представляет собой своеобразную последовательность эвапоритов , тонких песчаников и ракушечных известняков с прослоями мергеля и сланцев . Седиментология показывает, что они отлагались в различных пресноводных , солоноватых , гиперсоленых и квазиморских средах. Флора и фауна характеризуют изменчивые наземные , озерные , солончаковые и лагунные ассоциации. Климат ранней группы Пурбека, вероятно, был похож на современный средиземноморский и стал более влажным к концу берриаса. [ 24 ] Хотя группа Пурбек первоначально была известна как неофициальные пласты Пурбек, теперь ее можно разделить на верхнюю формацию Дарлстон и нижнюю формацию Лулворт . «Слои Верхнего Пурбека» и большая часть «Слоев Среднего Пурбека» содержатся в формации Дарлстон, старейшим отложением которой являются шлаковые пласты пачки лестничных проемов. [ 5 ] Слои шлака иногда считались границей юрского и мелового периодов, в результате чего вся формация Лулворт относилась к поздней юре, титонскому возрасту. [ 6 ] Однако, несмотря на неопределенность в отношении возраста пластов из-за отсутствия корреляции с фауной или датировкой, общепринято считать, что группа Пурбек полностью относится к раннемеловому периоду, а формация Лулворт относится к раннему берриасу . [ 6 ] [ 24 ] [ 25 ] Группа Пурбек явно подстилается позднеюрской Портлендской группой в заливе Дарлстон и имеет переходную, но местами неясную границу с вышележащей группой Уэлден в районе Певерил-Пойнт . [ 24 ]

Существует большая неопределенность относительно местонахождения образцов, собранных в формации Лулворт; единственный окончательный способ тестирования — это проанализировать матрицу каждого образца, чтобы определить его соленость. [ 5 ] Группа Пурбек имеет самую разнообразную птицетазовую фауну среди всех отложений Дорсета и является одной из немногих отложений берриаса в мире, но почти полностью ограничивается черепным или зубным материалом и следами. Owenodon hoggii — единственный другой названный птицетаз из слоев и известен только по зубной кости с зубами, первоначально описанными как вид Iguanodon . бедренная кость и спинная часть промежуточной гадрозавриформы Также известны , а также промежуточные орнитоподы и анкилозавры, известные как по окаменелостям тела, так и по отпечаткам ног. [ 13 ] Помимо птицетазовых, в формации Лулворт также обитают тероподы Nuthetes , амфибии , черепахи , ящерицы , змеи , млекопитающие и крокодилы , а также различные беспозвоночные. [ 5 ] [ 26 ] [ 27 ] К амфибиям формации Лулворт относятся саламандры Apricosiren и промежуточные батрахозавроидиды , альбанерпетонтиды Celtedens и лягушки Sunnybatrachus . [ 26 ] Известны четыре таксона черепах: криптодиры Dorsetochelys , Helochelydra , Hylaeochelys и Pleurosternon . [ 28 ] [ 29 ] Пурбек — одно из самых разнообразных раннемеловых отложений лепидозавров в мире . [ 30 ] Роды Becklesius , Dorsetisaurus , Durotrigia , Paramacellodus , Pseudosaurillus , Parasaurillus , Purbicella , Saurillus , Parviraptor и три неназванные морфологии зубов представляют известные чешуйчатые , [ 31 ] окаменелости, относящиеся к ринхоцефалам Homoeosaurus и Opisthias . также были найдены [ 32 ]
Разнообразный комплекс млекопитающих включает мелких плауновидных Durlstodon и Durlstotherium ; [ 24 ] неевтерийские перамурцы Peramus , Peramuroides , Magnimus и Kouriogenys ; [ 33 ] [ 34 ] неэвтерийные симметродонты Spalacotherium , Tinodon и Thereuodon ; [ 35 ] [ 36 ] неэвтериевые дриолестоиды Achyrodon , Amblotherium , Dorsetodon , Chunnelodon и Phascolestes ; [ 37 ] [ 38 ] [ 39 ] неэутерийные , мультитуберкулезные Albionbaatar , Bolodon , Gerhardodon ; Plagiaulax и Sunnyodon виды [ 40 ] [ 41 ] [ 42 ] неэвтериевые эвтриконодонты Trioracodon и Triconodon , , не относящиеся к млекопитающим морганукодонты Purbeckodon ; [ 43 ] и немлекопитающих докодонтов Peraiocynodon . [ 44 ] Крокодилы из отложений Лулворта включают Goniopholis gracilidens , Theriosuchus pusillus , Pholidosaurus purbeckensis , сомнительные останки, ранее известные как Goniopholis tenuidens , [ 5 ] и сомнительный таксон Macellodus brodiei . [ 1 ] [ 31 ] В определенных участках внутри формации также сохранились примитивные бекасы Simulidium и Pseudosimulium . [ 27 ] и нематоцеровые мухи Eoptychoptera , Brodilka и Eucorethrina . [ 25 ]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д и ж Оуэн, Р. (1861). Монография об ископаемых рептилиях формаций Уэлден и Пурбек. Часть V. Отряд Lacertilia . Палеонтографическое общество . стр. 31–39.
- ^ Фальконер, Х. (1861). «Заметка о синонимии ископаемого рода Echinodon профессора Оуэна» . Анналы и журнал естественной истории . 8 (46): 341. дои : 10.1080/00222936108697425 .
- ^ Перейти обратно: а б Оуэн, Р. (1874). Монография об ископаемых рептилиях формаций Уэлден и Пурбек. Дополнение № В. Динозаврия ( Игуанодон ) . Том. Доп. 5. Палеонтографическое общество. стр. 1–18.
- ^ Перейти обратно: а б Лидеккер, Р. (1888). Каталог ископаемых рептилий и амфибий в Британском музее (естественная история). Часть I. Содержащая отряды Ornithosauria, Crocodilia, Dinosauria, Squamata, Rhynchocephalia и Pterosauria . Тейлор и Фрэнсис . стр. 247–248.
- ^ Перейти обратно: а б с д и ж Солсбери, Юго-Запад (2002 г.). «Крокодилы из группы известняков Пурбека нижнего мела (берриаса) в Дорсете, южная Англия». Специальные статьи по палеонтологии . 68 (Жизнь и окружающая среда во времена Пурбека): 121–144.
- ^ Перейти обратно: а б с д и ж г час я Гальтон, П.М. (1978). «Fabrosauridae, базальное семейство птицетазовых динозавров (Reptilia: Ornithopoda)». Палеонтологический журнал . 52 (1–2): 138–159. Бибкод : 1978PalZ...52..138G . дои : 10.1007/BF03006735 . S2CID 84613826 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т Серено, Пол К. (2012). «Таксономия, морфология, жевательная функция и филогения гетеродонтозавридных динозавров» . ZooKeys (226): 1–225. дои : 10.3897/zookeys.226.2840 . ПМК 3491919 . ПМИД 23166462 .
- ^ Перейти обратно: а б с д и Норман, Д.Б.; Барретт, премьер-министр (2002). «Порнитишские динозавры нижнего мела (берриаса) Англии». Специальные статьи по палеонтологии . 68 (Жизнь и окружающая среда во времена Пурбека): 161–189.
- ^ Батлер, Р.Дж.; Гальтон, премьер-министр; Порро, Л.Б.; Кьяппе, LM; Хендерсон, DM; Эриксон, генеральный директор (2009). «Нижние пределы размеров тела птицетазовых динозавров, выведенные на основе нового верхнеюрского гетеродонтозаврид из Северной Америки» . Труды Королевского общества Б. 277 (1680): 375–381. дои : 10.1098/rspb.2009.1494 . ПМЦ 2842649 . ПМИД 19846460 .
- ^ Ронан Аллен, Ромен Вулло, Ли Розада, Жереми Анкетен, Рено Бурже и др.. Палеобиоразнообразие позвоночных раннего мела (берриаса) Анжак-Шаранта Лагерштетт (юго-запад Франции): последствия для круговорота континентальной фауны на границе J/K . Geodiversitas, Национальный музей естественной истории Парижа, В печати. ffhal-03264773f
- ^ Перейти обратно: а б с Серено, ПК (1997). «Происхождение и эволюция динозавров» (PDF) . Ежегодный обзор наук о Земле и планетах . 25 : 435–489. Бибкод : 1997AREPS..25..435S . дои : 10.1146/annurev.earth.25.1.435 .
- ^ Нопца, Ф. (1928). «Роды пресмыкающихся» . Палеобиология . 1 : 163–188.
- ^ Перейти обратно: а б с д Барретт, премьер-министр; Мейдмент, ЮКР (2011). «Динозавры Дорсета: Часть III, птицетазовые динозавры (Dinosauria, Ornithischia) с дополнительными комментариями о зауроподах». Труды Дорсетского общества естественной истории и археологии . 132 : 145–163.
- ^ Делар, Дж. Б. (1959). «Мезозойские рептилии Дорсета. Часть 2». Труды Дорсетского общества естественной истории и археологии . 80 : 52–90.
- ^ Барретт, премьер-министр; Кларк, Дж. Б.; Бринкман, Д.Б.; Чепмен, SD; Энсом, ПК (2002). «Морфология, гистология и идентификация« граниконов »известняковой формации Пурбек (нижний мел: берриас) в Дорсете, южная Англия» (PDF) . Меловые исследования . 23 (2): 279–295. Бибкод : 2002CrRes..23..279B . дои : 10.1006/cres.2002.1002 .
- ^ Перейти обратно: а б Талборн, РА (1971). «Происхождение и эволюция птицетазовых динозавров». Природа . 234 (5324): 75–78. Бибкод : 1971Natur.234...75T . дои : 10.1038/234075a0 . S2CID 4193262 .
- ^ Перейти обратно: а б Гальтон, премьер-министр (1972). «Классификация и эволюция динозавров-орнитопод». Природа . 239 (5373): 464–466. Бибкод : 1972Natur.239..464G . дои : 10.1038/239464a0 . S2CID 4196759 .
- ^ Перейти обратно: а б Серено, ПК (1991). « Лесотозавр» , «Фаброзавриды» и ранняя эволюция орнитисхий». Журнал палеонтологии позвоночных . 11 (2): 168–197. Бибкод : 1991JVPal..11..168S . дои : 10.1080/02724634.1991.10011386 .
- ^ Перейти обратно: а б с Батлер, Р.Дж.; Апчерч, П.; Норман, Д.Б. (2008). «Филогения птицетазовых динозавров». Журнал систематической палеонтологии . 6 (1): 1–40. Бибкод : 2008JSPal...6....1B . дои : 10.1017/S1477201907002271 . S2CID 86728076 .
- ^ Батлер, Р.Дж.; Лиён, Дж.; Джун, К.; Годфруа, П. (2011). «Посткраниальная остеология и филогенетическое положение небольшого птицетазового динозавра Changchunsaurus parvus из формации Квантоу (мел: апт-сеноман) провинции Цзилинь, северо-восточный Китай». Палеонтология . 54 (3): 667–683. Бибкод : 2011Palgy..54..667B . дои : 10.1111/j.1475-4983.2011.01046.x .
- ^ Пол, Д.; Раухут, OWM; Бесерра, М. (2011). «Среднеюрский гетеродонтозавридный динозавр из Патагонии и эволюция гетеродонтозаврид». Naturwissenschaften . 98 (5): 369–379. Бибкод : 2011NW.....98..369P . дои : 10.1007/s00114-011-0780-5 . ПМИД 21452054 . S2CID 22636871 .
- ^ Дьедонне, П.-Э.; Крузадо-Кабальеро, П.; Годфруа, П.; Тортоса, Т. (2020). «Новая филогения цераподских динозавров». Историческая биология . 33 (10): 1–21. Бибкод : 2021HBio...33.2335D . дои : 10.1080/08912963.2020.1793979 .
- ^ Норман, Д.Б.; Кромптон, штат Аризона; Батлер, Р.Дж.; Порро, Л.Б.; Чариг, AJ (2011). «Нижнеюрский птицетазовый динозавр Heterodontosaurus tucki Crompton and Charig 1962: анатомия черепа, функциональная морфология, таксономия и взаимоотношения» . Зоологический журнал Линнеевского общества . 162 : 182–279. дои : 10.1111/j.1096-3642.2011.00697.x .
- ^ Перейти обратно: а б с д Свитман, Южная Каролина; Смит, Г.; Мартилль, DM (2017). «Высокопроизводные эвтерийные млекопитающие раннего мелового периода на юге Британии» . Acta Palaeontologica Polonica . 62 (4): 657–665. дои : 10.4202/app.00408.2017 .
- ^ Перейти обратно: а б Лукашевич, Э.Д.; Корам, РА; Яржембовский, Э.А. (2001). «Новые настоящие мухи (Insecta: Diptera) из нижнего мела южной Англии». Меловые исследования . 22 (4): 451–460. Бибкод : 2001CrRes..22..451L . дои : 10.1006/cres.2001.0265 .
- ^ Перейти обратно: а б Эванс, SE; Макгоуэн, Дж.Дж. (2002). «Останки лисамфибий из группы известняков Пурбек, южная Англия». Специальные статьи по палеонтологии . 68 (Жизнь и окружающая среда во времена Пурбека): 103–119.
- ^ Перейти обратно: а б Мостовский, М.Б.; Росс, Эй Джей; Шадзевский Р.; Кшеминский, В. (2003). «Переописание Simulidium priscum Westwood и Pseudosimulium humidum (Brodie) (Insecta: Diptera: Rhagionidae) из группы известняков Пурбека (нижний мел) Англии». Журнал систематической палеонтологии . 1 (1): 59–64. Бибкод : 2003JSPal...1...59M . дои : 10.1017/S1477201903001020 . S2CID 55161519 .
- ^ Милнер, Арканзас (2004). «Черепахи из известняковой группы Пурбек в Дорсете, южная Англия» (PDF) . Палеонтология . 47 (6): 1441–1467. Бибкод : 2004Palgy..47.1441M . дои : 10.1111/j.0031-0239.2004.00418.x . S2CID 86775565 .
- ^ Перес-Гарсия, А. (2014). «Ревизия малоизвестной Dorsetochelys typocardium , относительно многочисленной плевростернидной черепахи (Paracryptodira) в раннем мелу Европы». Меловые исследования . 49 : 152–162. Бибкод : 2014CrRes..49..152P . дои : 10.1016/j.cretres.2014.02.015 .
- ^ Свитман, Южная Каролина; Эванс, SE (2011). «Лепидозавры (Ящерицы)». В Баттене, Д. (ред.). Окаменелости английского Уэлдена . Палеонтологическая ассоциация. стр. 264–284. ISBN 978-1-444-36711-9 .
- ^ Перейти обратно: а б Эванс, SE; Джонс, MEH; Мацумото, Р. (2012). «Новый череп ящерицы из группы известняков Пурбек (нижний мел) в Англии». Бюллетень геологического общества Франции . 183 (6): 517–524. дои : 10.2113/gssgfbull.183.6.517 .
- ^ Эванс, SE (1998). «Фауна лепидозавров раннего мела: переходная клада». В Лукасе, СГ; Киркланд, Дж.И.; Эстеп, JW (ред.). Нижне- и среднемеловые наземные экосистемы . Бюллетень Музея естественной истории Нью-Мексико , 14. стр. 195–200.
- ^ Дэвис, Б.М. (2012). «Микрокомпьютерная томография выявляет разнообразие перамуранских млекопитающих из группы Пурбека (берриаса) Англии» . Палеонтология . 55 (4): 789–817. Бибкод : 2012Palgy..55..789D . дои : 10.1111/j.1475-4983.2012.01161.x .
- ^ Сигоньо-Рассел, Д. (1999). «Повторная оценка перамуры (млекопитающих, кладотерий) на основе новых экземпляров нижнего мела из Англии и Марокко». Геодиверситас . 21 (1): 93–127.
- ^ Энсом, ПК; Сигоньо-Рассел, Д. (2000). «Новые симметродонты (Mammalia, Theria) из группы известняков Пурбек, нижний мел, южная Англия» (PDF) . Меловые исследования . 21 (6): 767–779. Бибкод : 2000CrRes..21..767E . дои : 10.1006/cres.2000.0227 .
- ^ Сигоньо-Рассел, Д.; Энсом, ПК (1998). « Тереуодон (Theria, Symmetrodonta) из нижнего мела Северной Африки и Европы и краткий обзор симметродонтов». Меловые исследования . 19 (3–4): 445–470. Бибкод : 1998CrRes..19..445S . дои : 10.1006/cres.1998.0115 .
- ^ Энсом, ПК; Сигоньо-Рассел, Д. (1998). «Новые дриолестоидные млекопитающие из группы известняков Пурбек базального мела на юге Англии». Палеонтология . 41 (1): 35–55.
- ^ Оуэн, Р. (1871). «Монография ископаемых млекопитающих мезозойских формаций» . Палеонтографическое общество . 24 (110): 1–115. Бибкод : 1871МПалС..24Д...5О . дои : 10.1080/02693445.1871.12113244 .
- ^ Аверьянов, АО; Мартин, Т.; Лопатин, А.В. (2013). «Новая филогения базальных Trechnotheria и Cladotheria и сходство с южноамериканскими эндемичными млекопитающими позднего мела». Naturwissenschaften . 100 (4): 311–326. Бибкод : 2013NW....100..311A . дои : 10.1007/s00114-013-1028-3 . ПМИД 23494201 . S2CID 18504005 .
- ^ Килан-Яворовска, З.; Энсом, ПК (1994). «Крошечные плагиаулакоидные мультитуберкулезные млекопитающие из известняковой формации Пурбек в Дорсете, Англия». Палеонтология . 37 (1): 17–31.
- ^ Килан-Яворовска, З.; Энсом, ПК (1994). «Многотуберкулезные млекопитающие из известняковой формации Пурбек верхней юры на юге Англии». Палеонтология . 35 (1): 95–126.
- ^ Фальконер, Х. (1857). «Описание двух видов ископаемых млекопитающих рода Plagiaulax из Пурбека». Ежеквартальный журнал Геологического общества . 13 (1–2): 261–282. дои : 10.1144/GSL.JGS.1857.013.01-02.39 . S2CID 128645686 .
- ^ Батлер, премьер-министр; Сигоньо-Рассел, Д.; Энсом, ПК (2012). «Возможное сохранение морганукодонтанов в группе известняков Пурбек нижнего мела (Дорсет, Англия)». Меловые исследования . 33 (1): 135–145. Бибкод : 2012CrRes..33..135B . дои : 10.1016/j.cretres.2011.09.007 .
- ^ Сигоньо-Рассел, Д. (2003). «Докодонты Британского мезозоя». Acta Palaeontologica Polonica . 48 (3): 357–374.