Аллель
Аллель [ 1 ] , или аллеломорф , является вариантом последовательности нуклеотидов в определенном месте или в локусе на молекуле ДНК . [ 2 ]
Аллели могут отличаться в одном положении с помощью отдельных нуклеотидных полиморфизмов (SNP), [ 3 ] Но они также могут иметь вставки и удаление до нескольких тысяч пар оснований . [ 4 ]
Большинство наблюдаемых аллелей приводят к незначительному изменению функции продукта гена, для которого он кодирует. Однако иногда разные аллели могут привести к различным наблюдаемым фенотипическим признакам , таким как различная пигментация . Примечательным примером этого является открытие Грегора Менделя о том, что белые и фиолетовые цветы в растениях гороха были результатом одного гена с двумя аллелями.
Почти все многоклеточные организмы имеют два набора хромосом в какой -то момент в их биологическом жизненном цикле ; То есть они диплоидны . В этом случае хромосомы могут быть в паре . Каждая хромосома в паре содержит одни и те же гены в одном и том же порядке и поместите, по всей длине хромосомы. Для данного гена , если две хромосомы содержат один и тот же аллель, они и организм гомозиготны по этому гену. Если аллели разные, они и организм гетерозиготных по отношению к этому геном.
Популярные определения «аллеля» обычно относятся только к разным аллелям в генах. Например, группировка крови ABO контролируется геном ABO , который имеет шесть общих аллелей (варианты). В генетике популяции живого человека почти каждый фенотип для гена ABO является некоторой комбинацией только этих шести аллелей. [ 5 ] [ 6 ]
Этимология
[ редактировать ]Слово «аллель» - это короткая форма «аллеломорф» («Другая форма», слово, придуманное британскими генетиками Уильямом Бейтсоном и Эдит Ребекка Сондерс ) в 1900 -х годах, [ 7 ] [ 8 ] который использовался в первые дни генетики для описания вариантов форм гена , обнаруженного как различные фенотипы . Он вытекает из греческого префикса ἀλληλο-, аллело- , что означает «взаимный», «взаимный» или «друг друга», что само по себе связано с греческим прилагательным ἄλλος, аллос (родственные с латинскими амиусом ), что означает «другой».
Аллели, которые приводят к доминирующим или рецессивным фенотипам
[ редактировать ]Во многих случаях генотипические взаимодействия между двумя аллелями в локусе могут быть описаны как доминирующие или рецессивные , согласно которому из двух гомозиготных фенотипов гетерозигота больше всего напоминает. Там, где гетерозигота неотличима от одного из гомозигот, экспрессированный аллель - это тот, который приводит к «доминантному» фенотипу, [ 9 ] [ 10 ] А другой аллель, как говорят, «рецессивный». Степень и схема доминирования варьируются среди локусов. Этот тип взаимодействия был сначала формально описан Грегором Менделем . Тем не менее, многие черты бросают вызов этой простой категоризации, и фенотипы смоделированы по доминированию и полигенному наследованию . [ 11 ]
Термин « дикий тип » иногда используется для описания аллеля, который, как считается, способствует типичному фенотипическому характеру, как видно из «диких» популяций организмов, таких как фруктовые мухи ( Drosophila melanogaster ). Такой аллель «дикого типа» исторически рассматривался как приводящий к доминирующему (подавляющему - всегда экспрессирующему), общему и нормальному фенотипу, в отличие от « мутантных » аллелей, которые приводят к рецессивным, редким и часто вредным фенотипам. Ранее считалось, что большинство людей были гомозиготными по аллелю «дикого типа» в большинстве локусов генов, и что любой альтернативный «мутантный» аллель был обнаружен в гомозиготной форме в небольшом меньшинстве «пораженных» людей, часто в качестве генетических заболеваний , и и и чаще в гетерозиготной форме в « носителях » для мутантного аллеля. В настоящее время ценится, что большинство или все локусы генов очень полиморфны, с множественными аллелями, частоты которых варьируются от популяции к популяции, и что большая часть генетических изменений скрыта в форме аллелей, которые не вызывают очевидных фенотипических различий. Аллели дикого типа часто обозначаются со знаком SuperScript Plus ( т.е. с + для аллеля P ). [ 12 ]
Несколько аллелей
[ редактировать ]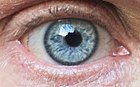
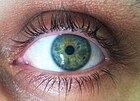



Популяция или виды организмов обычно включают несколько аллелей в каждом локусе среди различных людей. Аллельное изменение в локусе измеримо как количество присутствующих аллелей ( полиморфизм ), или доля гетерозигот в популяции. Нулевый аллель - это вариант генов, в котором отсутствует нормальная функция гена, потому что он либо не экспрессируется, либо экспрессированный белок неактивен.
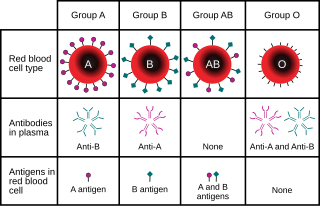
Например, в генном локусе для ABO типа крови углеводных антигенов у людей, [ 13 ] Классическая генетика распознает три аллеля, я А , Я Беременный , и я, которые определяют совместимость переливания крови . У любого человека есть один из шести возможных генотипов (я А я А , Я А я, я Беременный я Беременный , Я Беременный я, я А я Беременный и ii), которые производят один из четырех возможных фенотипов : «тип A» (произведен I А я А гомозиготный и я А я гетерозиготные генотипы), «Тип B» (произведен I Беременный я Беременный гомозиготный и я Беременный я гетерозиготные генотипы), «тип AB», произведенный I А я Беременный Гетерозиготный генотип и «тип O», продуцируемый II гомозиготным генотипом II. (В настоящее время известно, что каждый из аллелей A, B и O на самом деле представляет собой класс множественных аллелей с различными последовательностями ДНК, которые производят белки с идентичными свойствами: более 70 аллелей известно в локусе ABO. [ 14 ] Следовательно, человек с кровью «типа А» может быть гетерозигота АО, гомозигота АА или гетерозигота АА с двумя разными «А» аллелями).
Частоты генотипа
[ редактировать ]Частота аллелей в диплоидной популяции может быть использована для прогнозирования частот соответствующих генотипов (см. Принцип Харди -Вайнберг ). Для простой модели, с двумя аллелями;


где p - частота одного аллеля, а Q - частота альтернативного аллеля, который обязательно суммирует единство. Затем с 2 Является ли доля населения гомозиготной для первого аллеля, 2 pq - это часть гетерозигот и q 2 является фракцией гомозиготной для альтернативного аллеля. Если первый аллель доминирует ко второму, то доля популяции, которая покажет доминирующий фенотип, является P 2 + 2 pq , а фракция с рецессивным фенотипом является q 2 .
С тремя аллелями:
- и


В случае нескольких аллелей в диплоидном локусе количество возможных генотипов (G) с рядом аллелей (а) определяется выражением:

Аллельное доминирование при генетических расстройствах
[ редактировать ]Ряд генетических нарушений вызваны, когда индивидуум наследует два рецессивных аллеля на черту одного гена. Рецессивные генетические расстройства включают альбинизм , муковисцидоз , галактосемию , фенилкетонурию (ПКУ) и болезнь Тей -сах . Другие расстройства также связаны с рецессивными аллелями, но из -за того, что генный локус расположен на Х -хромосоме, так что у мужчин есть только одна копия (то есть они гемизиготные ), они чаще встречаются у мужчин, чем у женщин. Примеры включают красный -зеленый цвет слепоты и хрупкий синдром X.
Другие расстройства, такие как болезнь Хантингтона , возникают, когда человек наследует только один доминирующий аллель.
Эпиаллелы
[ редактировать ]В то время как наследуемые признаки обычно изучаются с точки зрения генетических аллелей, эпигенетические знаки, такие как метилирование ДНК, могут быть унаследованы в специфических геномных областях у определенных видов, процесс, называемый трансгенерационным эпигенетическим наследством . Термин Epiallele используется для отличия этих наследственных марок от традиционных аллелей, которые определяются нуклеотидной последовательности . [ 15 ] Специфический класс Epiallele, метастабильные эпиаллелы , был обнаружен у мышей и у людей, который характеризуется стохастическим (вероятностным) установлением эпигенетического состояния, которое может быть митотически унаследовать. [ 16 ] [ 17 ]
Идиоморф
[ редактировать ]Термин «идиоморф» от греческого «morphos» (форма) и «Idio» (единственный, уникальный), был введен в 1990 году вместо «аллеля» для обозначения последовательностей в одном и том же локусе в разных штаммах, которые не имеют сходства последовательности и Вероятно, не разделяют общие филогенетические отношения. Он используется в основном в генетических исследованиях микологии . [ 18 ] [ 19 ]
Смотрите также
[ редактировать ]Ссылки и примечания
[ редактировать ]- ^ Великобритания : / ˈ æ l ː l / , / ə ˈ l ː l / ; США : / ə ˈ l iː l / ; Современное образование от греческого ἄλλος állos , "Другое"
- ^ Graur, D (2016). Молекулярная и геномная эволюция . Sunderland MA (США): Sinauer Associates, Inc.
- ^ Смигильски, Элизабет М.; Сироткин, Карл; Уорд, Мингхонг; Шерри, Стивен Т. (1 января 2000 г.). «DBSNP: база данных отдельных нуклеотидных полиморфизмов» . Исследование нуклеиновых кислот . 28 (1): 352–355. doi : 10.1093/nar/28.1.352 . ISSN 0305-1048 . PMC 102496 . PMID 10592272 .
- ^ Элстон, Роберт; Сатагопан, Джая; Sun, Shuying (2012). «Генетическая терминология». Статистическая человеческая генетика . Методы в молекулярной биологии. Тол. 850. С. 1–9. doi : 10.1007/978-1-61779-555-8_1 . ISBN 978-1-61779-554-1 Полем ISSN 1064-3745 . PMC 4450815 . PMID 22307690 .
- ^ Seltsam A, Hallensleben M, Kollmann A, Blasczyk R (октябрь 2003 г.). «Природа разнообразия и диверсификации в локусе ABO» . Кровь . 102 (8): 3035–42. doi : 10.1182/blood-2003-03-0955 . PMID 12829588 .
- ^ Огасавара К., Баннай М., Сайту Н., Ябе Р., Наката К., Такека М., Фудзисава К., Учикава М., Ишикава Ю., Джуджи Т, Токунага К (июнь 1996 г.). «Обширный полиморфизм гена группы крови ABO: три основные линии аллелей для общих фенотипов ABO». Человеческая генетика . 97 (6): 777–83. doi : 10.1007/bf02346189 . PMID 8641696 . S2CID 12076999 .
- ^ Craft, Jude (2013). «Гены и генетика: язык научного открытия» . Гены и генетика . Оксфордский английский словарь . Архивировано с оригинала 29 января 2018 года . Получено 14 января 2016 года .
- ^ Бейтсон, У. и Сондерс, Эр (1902) «Факты наследственности в свете открытия Менделя». Отчеты Комитету эволюции Королевского общества, I. pp. 125–160
- ^ Хартл, Даниэль Л.; Элизабет В. Джонс (2005). Основная генетика: перспектива геномики (4 -е изд.). Jones & Bartlett Publishers. п. 600. ISBN 978-0-7637-3527-2 .
- ^ MGA, Иша; Куреши, Абид; Танакур, Nistant; Гупта, Акт Карар; Карар, Манодж (сентябрь 2017 г.). "ASPSPSI: ASP-SYA . G3 . 7 (9): 2931–2 doi : 10.1534/ g3.117.04 PMC 5592921 . PMID 28696921 .
- ^ "Аллель" . Genome.gov . Архивировано из оригинала 28 июня 2021 года . Получено 3 июля 2021 года .
- ^ Ба Пирс (2020). Генетика концептуальный подход (7 изд.). Макмиллан. п. 60. ISBN 978-1-319-21680-1 .
- ^ Виктор А. МакКусик; Кассандра Л. Книффин; Пол Дж. Конверс; Ада Хамош (10 ноября 2009 г.). «Або гликозилтрансфераза; abo» . Онлайн Мендельанский наследство в человеке . Национальная библиотека медицины. Архивировано из оригинала 24 сентября 2008 года . Получено 24 марта 2010 года .
- ^ Yip SP (январь 2002 г.). «Изменение последовательности в локусе человека» . Анналы человеческой генетики . 66 (1): 1–27. doi : 10.1017/s0003480001008995 . PMID 12014997 .
- ^ Дейсингер, Люсия; Уайтлау, Эмма (31 января 2012 г.). «Понимание трансгенерационного эпигенетического наследования через гаметы у млекопитающих». Nature Reviews Genetics . 13 (3): 153–62. doi : 10.1038/nrg3188 . PMID 22290458 . S2CID 8654616 .
- ^ Ракян, Вардхман К; Блевитт, Марни Э; Друкер, Рики; Preis, jost i; Уайтлау, Эмма (июль 2002 г.). «Метастабильные эпиаллелы у млекопитающих». Тенденции в генетике . 18 (7): 348–351. doi : 10.1016/s0168-9525 (02) 02709-9 . PMID 12127774 .
- ^ Waterland, RA; Долиной, округ Колумбия; Лин, младший; Смит, Калифорния; Ши, х; Тахилиани, кг (сентябрь 2006 г.). «Материнские метильные добавки увеличивают метилирование ДНК потомства при слитых аксин». Бытие . 44 (9): 401–6. doi : 10.1002/dvg.20230 . PMID 16868943 . S2CID 36938621 .
- ^ Стекло, NL; Ли Л. (1992). «Выделение мутантов Neurospora Crassa мутант типа спаривания с помощью повторной индуцированной точечной (RIP) мутации» . Генетика . 132 : 125–133. doi : 10.1093/Genetics/132.1.125 . PMC 1205111 . PMID 1398049 .
- ^ Метценберг, Роберт Л.; Гласс, Н. Луиза (1990). «Тип спаривания и стратегии спаривания в нейроспоре ». Биологии . 12 (2): 53–59. doi : 10.1002/bies.950120202 . PMID 2140508 . S2CID 10818930 .
Внешние ссылки
[ редактировать ]| Базы данных управления авторитетом : национальный |
|---|