Медуза
| Медуза | |
|---|---|

| |
| Тихоокеанская морская крапива ( Chrysaora Fuscescens ) | |
| Научная классификация | |
| Домен: | Эукариота |
| Королевство: | Животное |
| Филум: | Cnidaria |
| Субфилум: | Медусозои |
| Groups included | |
| Cladistically included but traditionally excluded taxa | |
Медзой , также известная как морские желе , являются медузафазой некоторых желатиновых членов субфилума медузозоида , которая является основной частью филовой Cnidaria .
Медзой-это в основном свободно плавающие морские животные с зонтичными колокольчиками и запекающими щупальцами , хотя некоторые из них привязаны к морским дне стебли, а не подвижными. Колокол может пульсировать, чтобы обеспечить движение для эффективного передвижения . Щупальца вооружены жгучими клетками и могут использоваться для захвата добычи и защиты от хищников. У медузы сложный жизненный цикл . Медуза, как правило, является сексуальной фазой, которая производит Planula личинки . Затем они широко рассеиваются и входят в сидячую полип -фазу, которая может включать бесполое почкование до достижения сексуальной зрелости.
Jellyfish are found all over the world, from surface waters to the deep sea. Scyphozoans (the "true jellyfish") are exclusively marine, but some hydrozoans with a similar appearance live in freshwater. Large, often colorful, jellyfish are common in coastal zones worldwide. The medusae of most species are fast-growing, and mature within a few months then die soon after breeding, but the polyp stage, attached to the seabed, may be much more long-lived. Jellyfish have been in existence for at least 500 million years,[1] and possibly 700 million years or more, making them the oldest multi-organ animal group.[2]
Jellyfish are eaten by humans in certain cultures. They are considered a delicacy in some Asian countries, where species in the Rhizostomeae order are pressed and salted to remove excess water. Australian researchers have described them as a "perfect food": sustainable and protein-rich but relatively low in food energy.[3]
They are also used in cell and molecular biology research, especially the green fluorescent protein used by some species for bioluminescence. This protein has been adapted as a fluorescent reporter for inserted genes and has had a large impact on fluorescence microscopy.
The stinging cells used by jellyfish to subdue their prey can injure humans. Thousands of swimmers worldwide are stung every year, with effects ranging from mild discomfort to serious injury or even death. When conditions are favourable, jellyfish can form vast swarms, which can be responsible for damage to fishing gear by filling fishing nets, and sometimes clog the cooling systems of power and desalination plants which draw their water from the sea.
Names
The name jellyfish, in use since 1796,[4] has traditionally been applied to medusae and all similar animals including the comb jellies (ctenophores, another phylum).[5][6] The term jellies or sea jellies is more recent, having been introduced by public aquaria in an effort to avoid use of the word "fish" with its modern connotation of an animal with a backbone, though shellfish, cuttlefish and starfish are not vertebrates either.[7][8] In scientific literature, "jelly" and "jellyfish" have been used interchangeably.[9][10] Many sources refer to only scyphozoans as "true jellyfish".[11]
A group of jellyfish is called a "smack"[12] or a "smuck".[13]
Mapping to taxonomic groups

Phylogeny
Definition
The term jellyfish broadly corresponds to medusae,[4] that is, a life-cycle stage in the Medusozoa. The American evolutionary biologist Paulyn Cartwright gives the following general definition:
Typically, medusozoan cnidarians have a pelagic, predatory jellyfish stage in their life cycle; staurozoans are the exceptions [as they are stalked].[14]
The Merriam-Webster dictionary defines jellyfish as follows:
A free-swimming marine coelenterate that is the sexually reproducing form of a hydrozoan or scyphozoan and has a nearly transparent saucer-shaped body and extensible marginal tentacles studded with stinging cells.[15]
Given that jellyfish is a common name, its mapping to biological groups is inexact. Some authorities have called the comb jellies[16] and certain salps[16] jellyfish, though other authorities state that neither of these are jellyfish, which they consider should be limited to certain groups within the medusozoa.[17][18]
The non-medusozoan clades called jellyfish by some but not all authorities (both agreeing and disagreeing citations are given in each case) are indicated with "???" on the following cladogram of the animal kingdom:
| Animalia |
| ||||||||||||||||||



Medusozoan jellyfish
Jellyfish are not a clade, as they include most of the Medusozoa, barring some of the Hydrozoa.[19][20] The medusozoan groups included by authorities are indicated on the following phylogenetic tree by the presence of citations. Names of included jellyfish, in English where possible, are shown in boldface; the presence of a named and cited example indicates that at least that species within its group has been called a jellyfish.
| Cnidaria |
| ||||||||||||||||||||||||||||||







Taxonomy
The subphylum Medusozoa includes all cnidarians with a medusa stage in their life cycle. The basic cycle is egg, planula larva, polyp, medusa, with the medusa being the sexual stage. The polyp stage is sometimes secondarily lost. The subphylum include the major taxa, Scyphozoa (large jellyfish), Cubozoa (box jellyfish) and Hydrozoa (small jellyfish), and excludes Anthozoa (corals and sea anemones).[25] This suggests that the medusa form evolved after the polyps.[26] Medusozoans have tetramerous symmetry, with parts in fours or multiples of four.[25]
The four major classes of medusozoan Cnidaria are:
- Scyphozoa are sometimes called true jellyfish, though they are no more truly jellyfish than the others listed here. They have tetra-radial symmetry. Most have tentacles around the outer margin of the bowl-shaped bell, and long, oral arms around the mouth in the center of the subumbrella.[25]
- Cubozoa (box jellyfish) have a (rounded) box-shaped bell, and their velarium assists them to swim more quickly. Box jellyfish may be related more closely to scyphozoan jellyfish than either are to the Hydrozoa.[26]
- Hydrozoa medusae also have tetra-radial symmetry, nearly always have a velum (diaphragm used in swimming) attached just inside the bell margin, do not have oral arms, but a much smaller central stalk-like structure, the manubrium, with terminal mouth opening, and are distinguished by the absence of cells in the mesoglea. Hydrozoa show great diversity of lifestyle; some species maintain the polyp form for their entire life and do not form medusae at all (such as Hydra, which is hence not considered a jellyfish), and a few are entirely medusal and have no polyp form.[25]
- Staurozoa (stalked jellyfish) are characterized by a medusa form that is generally sessile, oriented upside down and with a stalk emerging from the apex of the "calyx" (bell), which attaches to the substrate. At least some Staurozoa also have a polyp form that alternates with the medusoid portion of the life cycle. Until recently, Staurozoa were classified within the Scyphozoa.[25]
There are over 200 species of Scyphozoa, about 50 species of Staurozoa, about 50 species of Cubozoa, and the Hydrozoa includes about 1000–1500 species that produce medusae, but many more species that do not.[27][28]
Fossil history



Since jellyfish have no hard parts, fossils are rare. The oldest unambiguous fossil of a free-swimming medusa is Burgessomedusa from the mid-Cambrian Burgess Shale of Canada, which is likely either a stem group of box jellyfish (Cubozoa) or Acraspeda (the clade including Staurozoa, Cubozoa, and Scyphozoa). Other claimed records from the Cambrian of China and Utah in the United States are uncertain, and possibly represent ctenophores instead.[29]
Anatomy

The main feature of a true jellyfish is the umbrella-shaped bell. This is a hollow structure consisting of a mass of transparent jelly-like matter known as mesoglea, which forms the hydrostatic skeleton of the animal.[25] The mesoglea is 95% or more composed of water,[30] and also contains collagen and other fibrous proteins, as well as wandering amebocytes that can engulf debris and bacteria. The mesogloea is bordered by the epidermis on the outside and the gastrodermis on the inside. The edge of the bell is often divided into rounded lobes known as lappets, which allow the bell to flex. In the gaps or niches between the lappets are dangling rudimentary sense organs known as rhopalia, and the margin of the bell often bears tentacles.[25]

On the underside of the bell is the manubrium, a stalk-like structure hanging down from the centre, with the mouth, which also functions as the anus, at its tip. There are often four oral arms connected to the manubrium, streaming away into the water below.[31] The mouth opens into the gastrovascular cavity, where digestion takes place and nutrients are absorbed. This is subdivided by four thick septa into a central stomach and four gastric pockets. The four pairs of gonads are attached to the septa, and close to them four septal funnels open to the exterior, perhaps supplying good oxygenation to the gonads. Near the free edges of the septa, gastric filaments extend into the gastric cavity; these are armed with nematocysts and enzyme-producing cells and play a role in subduing and digesting the prey. In some scyphozoans, the gastric cavity is joined to radial canals which branch extensively and may join a marginal ring canal. Cilia in these canals circulate the fluid in a regular direction.[25]
The box jellyfish is largely similar in structure. It has a squarish, box-like bell. A short pedalium or stalk hangs from each of the four lower corners. One or more long, slender tentacles are attached to each pedalium.[32] The rim of the bell is folded inwards to form a shelf known as a velarium which restricts the bell's aperture and creates a powerful jet when the bell pulsates, allowing box jellyfish to swim faster than true jellyfish.[25] Hydrozoans are also similar, usually with just four tentacles at the edge of the bell, although many hydrozoans are colonial and may not have a free-living medusal stage. In some species, a non-detachable bud known as a gonophore is formed that contains a gonad but is missing many other medusal features such as tentacles and rhopalia.[25] Stalked jellyfish are attached to a solid surface by a basal disk, and resemble a polyp, the oral end of which has partially developed into a medusa with tentacle-bearing lobes and a central manubrium with four-sided mouth.[25]
Most jellyfish do not have specialized systems for osmoregulation, respiration and circulation, and do not have a central nervous system. Nematocysts, which deliver the sting, are located mostly on the tentacles; true jellyfish also have them around the mouth and stomach.[33] Jellyfish do not need a respiratory system because sufficient oxygen diffuses through the epidermis. They have limited control over their movement, but can navigate with the pulsations of the bell-like body; some species are active swimmers most of the time, while others largely drift.[34] The rhopalia contain rudimentary sense organs which are able to detect light, water-borne vibrations, odour and orientation.[25] A loose network of nerves called a "nerve net" is located in the epidermis.[35][36] Although traditionally thought not to have a central nervous system, nerve net concentration and ganglion-like structures could be considered to constitute one in most species.[37] A jellyfish detects stimuli, and transmits impulses both throughout the nerve net and around a circular nerve ring, to other nerve cells. The rhopalial ganglia contain pacemaker neurones which control swimming rate and direction.[25]
In many species of jellyfish, the rhopalia include ocelli, light-sensitive organs able to tell light from dark. These are generally pigment spot ocelli, which have some of their cells pigmented. The rhopalia are suspended on stalks with heavy crystals at one end, acting like gyroscopes to orient the eyes skyward. Certain jellyfish look upward at the mangrove canopy while making a daily migration from mangrove swamps into the open lagoon, where they feed, and back again.[2]
Box jellyfish have more advanced vision than the other groups. Each individual has 24 eyes, two of which are capable of seeing colour, and four parallel information processing areas that act in competition,[38] supposedly making them one of the few kinds of animal to have a 360-degree view of its environment.[39]
Box jellyfish eye
The study of jellyfish eye evolution is an intermediary to a better understanding of how visual systems evolved on Earth.[40] Jellyfish exhibit immense variation in visual systems ranging from photoreceptive cell patches seen in simple photoreceptive systems to more derived complex eyes seen in box jellyfish.[40] Major topics of jellyfish visual system research (with an emphasis on box jellyfish) include: the evolution of jellyfish vision from simple to complex visual systems), the eye morphology and molecular structures of box jellyfish (including comparisons to vertebrate eyes), and various uses of vision including task-guided behaviors and niche specialization.
Evolution
Experimental evidence for photosensitivity and photoreception in cnidarians antecedes the mid 1900s, and a rich body of research has since covered evolution of visual systems in jellyfish.[41] Jellyfish visual systems range from simple photoreceptive cells to complex image-forming eyes. More ancestral visual systems incorporate extraocular vision (vision without eyes) that encompass numerous receptors dedicated to single-function behaviors. More derived visual systems comprise perception that is capable of multiple task-guided behaviors.
Although they lack a true brain, cnidarian jellyfish have a "ring" nervous system that plays a significant role in motor and sensory activity. This net of nerves is responsible for muscle contraction and movement and culminates the emergence of photosensitive structures.[40] Across Cnidaria, there is large variation in the systems that underlie photosensitivity. Photosensitive structures range from non-specialized groups of cells, to more "conventional" eyes similar to those of vertebrates.[41] The general evolutionary steps to develop complex vision include (from more ancestral to more derived states): non-directional photoreception, directional photoreception, low-resolution vision, and high-resolution vision.[40] Increased habitat and task complexity has favored the high-resolution visual systems common in derived cnidarians such as box jellyfish.[40]
Basal visual systems observed in various cnidarians exhibit photosensitivity representative of a single task or behavior. Extraocular photoreception (a form of non-directional photoreception), is the most basic form of light sensitivity and guides a variety of behaviors among cnidarians. It can function to regulate circadian rhythm (as seen in eyeless hydrozoans) and other light-guided behaviors responsive to the intensity and spectrum of light. Extraocular photoreception can function additionally in positive phototaxis (in planula larvae of hydrozoans),[41] as well as in avoiding harmful amounts of UV radiation via negative phototaxis. Directional photoreception (the ability to perceive direction of incoming light) allows for more complex phototactic responses to light, and likely evolved by means of membrane stacking.[40] The resulting behavioral responses can range from guided spawning events timed by moonlight to shadow responses for potential predator avoidance.[41][42] Light-guided behaviors are observed in numerous scyphozoans including the common moon jelly, Aurelia aurita, which migrates in response to changes in ambient light and solar position even though they lack proper eyes.[41]
The low-resolution visual system of box jellyfish is more derived than directional photoreception, and thus box jellyfish vision represents the most basic form of true vision in which multiple directional photoreceptors combine to create the first imaging and spatial resolution. This is different from the high-resolution vision that is observed in camera or compound eyes of vertebrates and cephalopods that rely on focusing optics.[41] Critically, the visual systems of box jellyfish are responsible for guiding multiple tasks or behaviors in contrast to less derived visual systems in other jellyfish that guide single behavioral functions. These behaviors include phototaxis based on sunlight (positive) or shadows (negative), obstacle avoidance, and control of swim-pulse rate.[43]
Box jellyfish possess "proper eyes" (similar to vertebrates) that allow them to inhabit environments that lesser derived medusae cannot. In fact, they are considered the only class in the clade Medusozoa that have behaviors necessitating spatial resolution and genuine vision.[41] However, the lens in their eyes are more functionally similar to cup-eyes exhibited in low-resolution organisms, and have very little to no focusing capability.[44][43] The lack of the ability to focus is due to the focal length exceeding the distance to the retina, thus generating unfocused images and limiting spatial resolution.[41] The visual system is still sufficient for box jellyfish to produce an image to help with tasks such as object avoidance.
Utility as a model organism
Box jellyfish eyes are a visual system that is sophisticated in numerous ways. These intricacies include the considerable variation within the morphology of box jellyfishes' eyes (including their task/behavior specification), and the molecular makeup of their eyes including: photoreceptors, opsins, lenses, and synapses.[41] The comparison of these attributes to more derived visual systems can allow for a further understanding of how the evolution of more derived visual systems may have occurred, and puts into perspective how box jellyfish can play the role as an evolutionary/developmental model for all visual systems.[45]
Characteristics
Box jellyfish visual systems are both diverse and complex, comprising multiple photosystems.[41] There is likely considerable variation in visual properties between species of box jellyfish given the significant inter-species morphological and physiological variation. Eyes tend to differ in size and shape, along with number of receptors (including opsins), and physiology across species of box jellyfish.[41]
Box jellyfish have a series of intricate lensed eyes that are similar to those of more derived multicellular organisms such as vertebrates. Their 24 eyes fit into four different morphological categories.[46] These categories consist of two large, morphologically different medial eyes (a lower and upper lensed eye) containing spherical lenses, a lateral pair of pigment slit eyes, and a lateral pair of pigment pit eyes.[43] The eyes are situated on rhopalia (small sensory structures) which serve sensory functions of the box jellyfish and arise from the cavities of the exumbrella (the surface of the body) on the side of the bells of the jellyfish.[41] The two large eyes are located on the mid-line of the club and are considered complex because they contain lenses. The four remaining eyes lie laterally on either side of each rhopalia and are considered simple. The simple eyes are observed as small invaginated cups of epithelium that have developed pigmentation.[47] The larger of the complex eyes contains a cellular cornea created by a mono ciliated epithelium, cellular lens, homogenous capsule to the lens, vitreous body with prismatic elements, and a retina of pigmented cells. The smaller of the complex eyes is said to be slightly less complex given that it lacks a capsule but otherwise contains the same structure as the larger eye.[47]
Box jellyfish have multiple photosystems that comprise different sets of eyes.[41] Evidence includes immunocytochemical and molecular data that show photopigment differences among the different morphological eye types, and physiological experiments done on box jellyfish to suggest behavioral differences among photosystems. Each individual eye type constitutes photosystems that work collectively to control visually guided behaviors.[41]
Box jellyfish eyes primarily use c-PRCs (ciliary photoreceptor cells) similar to that of vertebrate eyes. These cells undergo phototransduction cascades (process of light absorption by photoreceptors) that are triggered by c-opsins.[48] Available opsin sequences suggest that there are two types of opsins possessed by all cnidarians including an ancient phylogenetic opsin, and a sister ciliary opsin to the c-opsins group. Box jellyfish could have both ciliary and cnidops (cnidarian opsins), which is something not previously believed to appear in the same retina.[41] Nevertheless, it is not entirely evident whether cnidarians possess multiple opsins that are capable of having distinctive spectral sensitivities.[41]
Comparison with other organisms
Comparative research on genetic and molecular makeup of box jellyfishes' eyes versus more derived eyes seen in vertebrates and cephalopods focuses on: lenses and crystallin composition, synapses, and Pax genes and their implied evidence for shared primordial (ancestral) genes in eye evolution.[49]
Box jellyfish eyes are said to be an evolutionary/developmental model of all eyes based on their evolutionary recruitment of crystallins and Pax genes.[45] Research done on box jellyfish including Tripedalia cystophora has suggested that they possess a single Pax gene, PaxB. PaxB functions by binding to crystallin promoters and activating them. PaxB in situ hybridization resulted in PaxB expression in the lens, retina, and statocysts.[45] These results and the rejection of the prior hypothesis that Pax6 was an ancestral Pax gene in eyes has led to the conclusion that PaxB was a primordial gene in eye evolution, and that the eyes of all organisms likely share a common ancestor.[45]
The lens structure of box jellyfish appears very similar to those of other organisms, but the crystallins are distinct in both function and appearance.[49] Weak reactions were seen within the sera and there were very weak sequence similarities within the crystallins among vertebrate and invertebrate lenses.[49] This is likely due to differences in lower molecular weight proteins and the subsequent lack of immunological reactions with antisera that other organisms' lenses exhibit.[49]
All four of the visual systems of box jellyfish species investigated with detail (Carybdea marsupialis, Chiropsalmus quadrumanus, Tamoya haplonema and Tripedalia cystophora) have invaginated synapses, but only in the upper and lower lensed eyes. Different densities were found between the upper and lower lenses, and between species.[46] Four types of chemical synapses have been discovered within the rhopalia which could help in understanding neural organization including: clear unidirectional, dense-core unidirectional, clear bidirectional, and clear and dense-core bidirectional. The synapses of the lensed eyes could be useful as markers to learn more about the neural circuit in box jellyfish retinal areas.[46]
Evolution as a response to natural stimuli
The primary adaptive responses to environmental variation observed in box jellyfish eyes include pupillary constriction speeds in response to light environments, as well as photoreceptor tuning and lens adaptations to better respond to shifts between light environments and darkness. Interestingly, some box jellyfish species' eyes appear to have evolved more focused vision in response to their habitat.[50]
Pupillary contraction appears to have evolved in response to variation in the light environment across ecological niches across three species of box jellyfish (Chironex fleckeri, Chiropsella bronzie, and Carukia barnesi). Behavioral studies suggest that faster pupil contraction rates allow for greater object avoidance,[50] and in fact, species with more complex habitats exhibit faster rates. Ch. bronzie inhabit shallow beach fronts that have low visibility and very few obstacles, thus, faster pupil contraction in response to objects in their environment is not important. Ca. barnesi and Ch. fleckeri are found in more three-dimensionally complex environments like mangroves with an abundance of natural obstacles, where faster pupil contraction is more adaptive.[50] Behavioral studies support the idea that faster pupillary contraction rates assist with obstacle avoidance as well as depth adjustments in response to differing light intensities.
Light/dark adaptation via pupillary light reflexes is an additional form of an evolutionary response to the light environment. This relates to the pupil's response to shifts between light intensity (generally from sunlight to darkness). In the process of light/dark adaptation, the upper and lower lens eyes of different box jellyfish species vary in specific function.[43] The lower lens-eyes contain pigmented photoreceptors and long pigment cells with dark pigments that migrate on light/dark adaptation, while the upper-lens eyes play a concentrated role in light direction and phototaxis given that they face upward towards the water surface (towards the sun or moon).[43] The upper lens of Ch. bronzie does not exhibit any considerable optical power while Tr. cystophora (a box jellyfish species that tends to live in mangroves) does. The ability to use light to visually guide behavior is not of as much importance to Ch. bronzie as it is to species in more obstacle-filled environments.[43] Differences in visually guided behavior serve as evidence that species that share the same number and structure of eyes can exhibit differences in how they control behavior.
Largest and smallest
Jellyfish range from about one millimeter in bell height and diameter,[51] to nearly 2 metres (6+1⁄2 ft) in bell height and diameter; the tentacles and mouth parts usually extend beyond this bell dimension.[25]
The smallest jellyfish are the peculiar creeping jellyfish in the genera Staurocladia and Eleutheria, which have bell disks from 0.5 millimetres (1⁄32 in) to a few millimeters in diameter, with short tentacles that extend out beyond this, which these jellyfish use to move across the surface of seaweed or the bottoms of rocky pools;[51] many of these tiny creeping jellyfish cannot be seen in the field without a hand lens or microscope. They can reproduce asexually by fission (splitting in half). Other very small jellyfish, which have bells about one millimeter, are the hydromedusae of many species that have just been released from their parent polyps;[52] some of these live only a few minutes before shedding their gametes in the plankton and then dying, while others will grow in the plankton for weeks or months. The hydromedusae Cladonema radiatum and Cladonema californicum are also very small, living for months, yet never growing beyond a few mm in bell height and diameter.[53]

The lion's mane jellyfish, Cyanea capillata, was long-cited as the largest jellyfish, and arguably the longest animal in the world, with fine, thread-like tentacles that may extend up to 36.5 m (119 ft 9 in) long (though most are nowhere near that large).[54][55] They have a moderately painful, but rarely fatal, sting.[56] The increasingly common giant Nomura's jellyfish, Nemopilema nomurai, found in some, but not all years in the waters of Japan, Korea and China in summer and autumn is another candidate for "largest jellyfish", in terms of diameter and weight, since the largest Nomura's jellyfish in late autumn can reach 2 m (6 ft 7 in) in bell (body) diameter and about 200 kg (440 lb) in weight, with average specimens frequently reaching 0.9 m (2 ft 11 in) in bell diameter and about 150 kg (330 lb) in weight.[57][58] The large bell mass of the giant Nomura's jellyfish[59] can dwarf a diver and is nearly always much greater than the Lion's Mane, whose bell diameter can reach 1 m (3 ft 3 in).[60]
The rarely encountered deep-sea jellyfish Stygiomedusa gigantea is another candidate for "largest jellyfish", with its thick, massive bell up to 100 cm (3 ft 3 in) wide, and four thick, "strap-like" oral arms extending up to 6 m (19+1⁄2 ft) in length, very different from the typical fine, threadlike tentacles that rim the umbrella of more-typical-looking jellyfish, including the Lion's Mane.[61]
Desmonema glaciale, which lives in the Antarctic region, can reach a very large size (several meters).[62][63] Purple-striped jelly (Chrysaora colorata) can also be extremely long (up to 15 feet).[64]
Life history and behavior

1–3 Larva searches for site
4–8 Polyp grows
9–11 Polyp strobilates
12–14 Medusa grows
Life cycle
Jellyfish have a complex life cycle which includes both sexual and asexual phases, with the medusa being the sexual stage in most instances. Sperm fertilize eggs, which develop into larval planulae, become polyps, bud into ephyrae and then transform into adult medusae. In some species certain stages may be skipped.[65]
Upon reaching adult size, jellyfish spawn regularly if there is a sufficient supply of food. In most species, spawning is controlled by light, with all individuals spawning at about the same time of day; in many instances this is at dawn or dusk.[66] Jellyfish are usually either male or female (with occasional hermaphrodites). In most cases, adults release sperm and eggs into the surrounding water, where the unprotected eggs are fertilized and develop into larvae. In a few species, the sperm swim into the female's mouth, fertilizing the eggs within her body, where they remain during early development stages. In moon jellies, the eggs lodge in pits on the oral arms, which form a temporary brood chamber for the developing planula larvae.[67]
The planula is a small larva covered with cilia. When sufficiently developed, it settles onto a firm surface and develops into a polyp. The polyp generally consists of a small stalk topped by a mouth that is ringed by upward-facing tentacles. The polyps resemble those of closely related anthozoans, such as sea anemones and corals. The jellyfish polyp may be sessile, living on the bottom of boat hulls or other substrates, or it may be free-floating or attached to tiny bits of free-living plankton[68] or rarely, fish[69][70] or other invertebrates. Polyps may be solitary or colonial.[71] Most polyps are only millimetres in diameter and feed continuously. The polyp stage may last for years.[25]
После интервала и стимулированного сезонными или гормональными изменениями, полип может начать воспроизводить асексально путем почки и, в Scyphozoa, называется сегментирующей полипов или пирозой. Начнулся больше Scyphistomae, а также Ephyrae. [ 25 ] Начинающие сайты варьируются в зависимости от видов; Из луковиц щупальца , манубрий (над ртом) или гонадов гидромедусов. [ 68 ] В процессе, известном как стробиляция , щупальца полипа реабсорбируются, и тело начинает сузиться, образуя поперечные сужения, в нескольких местах вблизи верхней конечности полипа. Они углубляются, когда сайты сужения мигрируют по телу, и отдельные сегменты, известные как отделение Ephyra. Это предшественники свободного плавания взрослой стадии Медузы, которая является стадией жизни, которая обычно идентифицируется как медуза. [ 25 ] [ 72 ] Ephyrae, обычно всего в миллиметре по всему первоначальному, уплывают от полипа и растут. Полипы Limnomedusae могут асексально производить ползучую личинку личинки , которая ползает перед тем, как превратиться в другой полип. [ 25 ] Несколько видов могут вырабатывать новые медузы путем почтения непосредственно со стадии Медусана. Некоторые гидромедусы размножаются путем деления. [ 68 ]
Продолжительность жизни
Мало что известно о истории жизни многих медуз как о местах на морском дне, где не были обнаружены бентические формы этих видов. Тем не менее, асексально воспроизводящая форма стробила может иногда жить в течение нескольких лет, производя новые медузы (личинки Ephyra) каждый год. [ 73 ]
Необычный вид, Turritopsis dohrnii , ранее классифицированный как Turritopsis nutricula , [ 74 ] может быть эффективно бессмертным из-за его способности при определенных обстоятельствах превращаться из Медузы обратно в стадию полипов, тем самым избегая смерти, которая обычно ожидает после репродукции Медузы, если в противном случае они не съели какой-либо другой организм. До сих пор это изменение наблюдалось только в лаборатории. [ 75 ]
Локомоция

Используя лунное желе aurelia aurita в качестве примера, было показано, что медузы являются наиболее энергоэффективными пловцами всех животных. [ 76 ] Они движутся через воду, радиально расширяя и сжимая свои колокольчики, чтобы толкнуть воду за ними. Они останавливаются между этапами сокращения и расширения, чтобы создать два вихревых кольца. Мышцы используются для сокращения тела, которое создает первый вихрь и продвигает животное вперед, но мезоглея настолько упругая, что расширение приводится в действие исключительно путем расслабления колокола, что высвобождает энергию, хранящуюся от сокращения. Тем временем второе вихревое кольцо начинает вращаться быстрее, втягивая воду в колокол и толкая центр тела, давая вторичное и «свободное» усиление вперед. Механизм, называемый пассивной энергией, работает только в относительно маленькой медузах, движущейся на низких скоростях, что позволяет животному двигаться на 30 процентов дальше в каждом цикле плавания. Медзой достигла на 48 процентов более низкую стоимость транспорта (потребление пищи и кислорода по сравнению с энергией, потраченной на движение), чем другие животные в аналогичных исследованиях. Одна из причин этого заключается в том, что большая часть желатиновой ткани колокола неактивна, не используя энергию во время плавания. [ 77 ]
Экология
Диета
Медузы, как и другие Cnidarians, как правило, плотоядные (или паразитные), [ 78 ] питание планктонными организмами, ракообразными, мелкими рыбами, рыбными яйцами и личинками и другими медузами, проглатывая пищу и мочеивая непереваренные отходы через рот. Они пассивно охотятся, используя свои щупальца в качестве дрейфовых линий, или погружаются через воду, когда их щупальца широко распространяются; щупальца, которые содержат нематоцисты, чтобы оглушить или убить добычу , могут затем сгибаться, чтобы помочь привести его ко рту. [ 25 ] Их техника плавания также помогает им запечатлеть добычу; Когда их колокол расширяется, он затягивает воду, которая приносит большую добычу в пределах досягаемости щупальца. [ 79 ]
Несколько видов, таких как гемистома Aglaura, являются всеядными, питающимися микроплангом, которая представляет собой смесь зоопланктона и фитопланктона (микроскопических растений), таких как динофлагелляты . [ 80 ] Другие питают взаимные водоросли ( зооксантеллы ) в своих тканях; [ 25 ] Пятнистая медуза ( Mastigias Papua ) типична для них, получая часть своей питания от продуктов фотосинтеза и части от захваченного зоопланктона. [ 81 ] [ 82 ] ( Вернутая медуза Cassiopea Andromeda ) также имеет симбиотическую связь с микроводоросли , но захватывает крошечных животных, чтобы дополнить свою диету. Это делается путем выпуска крошечных шариков живых клеток, состоящих из мезоглеи . Они используют реснички, чтобы провести их через воду и жаждущие клетки, которые оглушают добычу. Кажется, что капля также обладают пищеварительными возможностями. [ 83 ]
Хищничество
Другие виды медузы являются одними из наиболее распространенных и важных хищников медуз. Морские анемоны могут съесть медузу, которая дрейфовала в их ассортимент. Другие хищники включают тунцы , акулы, рыбу -меч , морские черепахи и пингвины. [ 84 ] [ 85 ] Медзой, вымытая на пляже, потребляются лисами, другими наземными млекопитающими и птицами. [ 86 ] В целом, однако, мало животных охотятся на медузы; Они могут широко считаться лучшими хищниками в пищевой цепи. Как только медуза станут доминирующими в экосистеме, например, из -за перелова, которая удаляет хищники личинок медузы, не может быть никакого очевидного способа восстановления предыдущего баланса: они едят рыбные яйца и ювенильную рыбу и конкурируют с рыбой, предотвращая рыбные запасы от восстановления. [ 87 ]
Симбиоз
Некоторые мелкие рыбы невосприимчивы к укусам медузы и живут среди щупальц, служат приманкой в рыбной ловушке; Они безопасны от потенциальных хищников и могут делиться рыбой, пойманной медузой. [ 88 ] Медзоя пушечного ядра имеет симбиотические отношения с десятью различными видами рыб, и с длинным пауком -крабом , который живет внутри колокола, разделяя пищу медузы и заклеивая его ткани. [ 89 ]
Цветут

Круги представляют записи данных; Большие круги обозначают более высокую уверенность в результатах.
Медзой образует большие массы или цветы в определенных условиях окружающей среды океанских токов , питательных веществ , солнечного света, температуры, сезона, доступности добычи, снижения хищничества и кислорода концентрации . Токи собирают медузу вместе, особенно в годы с необычайно высоким населением. Медзой может обнаружить морские течения и плавать против течения, чтобы собраться в цветах. [ 91 ] [ 92 ] Медзой лучше выжить в богатой питательной, бедной кислородами, чем конкурентами, и, таким образом, могут пировать на планктоне без конкуренции. Медзой также может извлечь выгоду из более соленой воды, так как более соляные воды содержат больше йода , что необходимо для того, чтобы полипы превратились в медузы. Повышение температуры моря, вызванные изменением климата, также может способствовать цветению медуз, потому что многие виды медуз могут выжить в более теплых водах. [ 93 ] Повышенные питательные вещества из сельскохозяйственного или городского стока с питательными веществами, включая азотные и фосфорные соединения, увеличивают рост фитопланктона, вызывая эвтрофикацию и цветение водорослей . Когда фитопланктон умирает, они могут создавать мертвые зоны , так называемые, потому что они гипоксичны (низкий уровень кислорода). Это, в свою очередь, убивает рыбу и других животных, но не медуза, [ 94 ] позволяя им расцвести. [ 95 ] [ 96 ] Популяции медуз могут расширяться во всем мире в результате стока земли и перелога их естественных хищников . [ 97 ] [ 98 ] Медзой хорошо подходит для получения выгоды от нарушения морских экосистем. Они быстро воспроизводят; Они охотятся на многих видов, в то время как немногие виды охотятся на них; И они питаются через штрих, а не визуально, поэтому они могут эффективно кормить ночью и в мутных водах. [ 99 ] [ 100 ] может быть трудно Рыбам восстановить себя в морских экосистемах, как только они доминируют в медузе, потому что медуза питается планктоном, который включает в себя рыбные яйца и личинки . [ 101 ] [ 102 ] [ 96 ]

Как подозревалось на рубеже этого столетия, [ 107 ] [ 108 ] Цветения медуз увеличиваются по частоте. В период с 2013 по 2020 год Средиземноморская научная комиссия еженедельно контролировала частоту таких вспышек в прибрежных водах от Марокко до Черного моря, что обнаруживает относительно высокую частоту этих цветов почти круглый год, с пиками, наблюдаемыми с марта и часто и часто. Снова осенью. Цветы вызваны различными видами медуз, в зависимости от их локализации в бассейне: наблюдается явное доминирование Велелла Велелла в и западном вспышек Ноцтилука средиземноморском Пелагии и вспышки Mnemiopsis Leidyi в Черном море. [ 109 ]
Некоторые популяции медуз, которые продемонстрировали явное увеличение за последние несколько десятилетий, являются инвазивными видами , вновь прибывшие из других мест обитания: примеры включают Черное море , Каспийское море , Балтийское море , Центральное и Восточное Средиземное море , Гавайи, а также тропические и субтропические части Запада. Атлантика (включая Карибский бассейн , Мексиканский залив и Бразилия). [ 105 ] [ 106 ]
Цветение медуз может оказать существенное влияние на структуру сообщества. Некоторые плотоядные виды медуз добывают на зоопланктоне, в то время как другие сталкиваются с первичными производителями. [ 110 ] Снижение зоопланктона и ichthyoplankton из -за цветения медузы может пролистывать трофические уровни. Популяции медуз высокой плотности могут переоценить других хищников и уменьшить набор рыб. [ 111 ] Увеличение выпаса первичных производителей с помощью медузы также может прервать перенос энергии до более высоких трофических уровней. [ 112 ]
Во время цветения медузы значительно изменяют доступность питательных веществ в окружающей среде. Блум требует большого количества доступных органических питательных веществ в толще воды для роста, что ограничивает доступность для других организмов. [ 113 ] Некоторые медузы имеют симбиотическую связь с одноклеточными динофлагеллятами, что позволяет им ассимилировать неорганический углерод, фосфор и азот, создавая конкуренцию за фитопланктон. [ 113 ] Их большая биомасса делает их важным источником растворенного и частиц органического вещества для микробных сообществ посредством экскреции, производства слизи и разложения. [ 90 ] [ 114 ] Микробы разбивают органическое вещество на неорганический аммоний и фосфат. Тем не менее, низкая доступность углерода меняет процесс с производства к дыханию, создавая низкие области кислорода, что делает растворенный неорганический азот и фосфор в значительной степени недоступен для первичной продукции.
Эти цветы оказывают очень реальное влияние на отрасли промышленности. Медзой может переоценить рыбу, используя открытые ниши в проличном рыболовстве. [ 115 ] Уловка медузы может напрягать рыболовное снаряжение и привести к расходам, связанным с поврежденным снаряжением. Электростанции были закрыты из -за медузы, блокирующей поток охлаждающей воды. [ 116 ] Цветы также были вредны для туризма, вызывая рост укусов, а иногда и закрытие пляжей. [ 117 ]
Медзой образует компонент желе-палочек , события, где желатиновый зоопланктон падает в морское дно, обеспечивая еду для бентических организмов там. [ 118 ] В умеренных и субполярных областях желе, которые обычно следуют сразу после цветения. [ 119 ]
Среда обитания

Большинство медуз - морские животные, хотя несколько гидромедусов обитают в пресной воде . Самым известным пресноводным примером является космополитическая гидрозой -медуза, Craspedacusta sowerbii . Это меньше, чем дюйм (2,5 см) в диаметре, бесцветный и не укун. [ 120 ] Некоторые популяции медуз ограничились прибрежными соленственными озерами, такими как озеро медуз в Палау. [ 121 ] Озеро медузы - это морское озеро , где миллионы золотых медуз ( мастигис spp.) Ежедневно мигрируют по озеру. [ 82 ]
Хотя большинство медуз живут хорошо от пола океана и являются частью планктона, некоторые виды тесно связаны с дном на протяжении большей части своей жизни и могут считаться бентическими . Вернутая медуза в роде Cassiopea обычно лежит на дне мелких лагун, где они иногда мягко пульсируют с помощью зонтичной вершины. Даже некоторые глубоководные виды гидромедусов и scyphomedusae обычно собираются на дне или рядом с ними. Все stauroMedusae найдены прикрепленными к морским водоромам или каменистым или другим твердому материалу на дне. [ 122 ]
Некоторые виды явно адаптируются к приливному потоку. В бухте Роско медуза ездит на току в приливе, пока они не попадут в гравийный бар , а затем не спустится под током. Они остаются в неподвижном воде, пока прилив не поднимется, поднимается и позволяя ему подметать их обратно в залив. Они также активно избегают пресной воды из горного таяния снега, ныряют, пока не найдут достаточно соли. [ 2 ]
Паразиты
Медзой - хозяева широкого спектра паразитических организмов. Они действуют как промежуточные хозяева эндопаразитовых гельминтов , причем инфекция передается окончательной рыбе -хозяине после хищничества . Некоторые дигенеанские трематоды , особенно виды в семействе Lepocreadiidae , используют медузу в качестве их вторых промежуточных хозяев. Рыба заражается трематодами, когда питаются зараженной медузой. [ 123 ] [ 124 ]
Отношение к людям

Рыболовство
Медзой давно едят в некоторых частях света. [ 3 ] Рыболовство начало собирать американскую пушечную медузу, Stomolophus meleagris , вдоль южного атлантического побережья Соединенных Штатов и в Мексиканском заливе для экспорта в Азию. [ 126 ]
Медзой также собирают для их коллагена , который исследуется для использования в различных приложениях, включая лечение ревматоидного артрита . [ 127 ]
Аквакультура и рыболовство других видов часто страдают от серьезных потерь - и, следовательно, потери продукции - из -за медузы. [ 128 ] [ 129 ]
Продукция

Аристотель заявил в тех частях животных IV, что медузы (морские отрывы) ели зимой в тушеной рыбной туфли. [ 130 ]
В некоторых странах, включая Китай, Японию и Корею, медузы являются деликатесом. Медзой сушат, чтобы предотвратить порчу. Только около 12 видов Scyphozoan Jellyзу, принадлежащих к заказом Rhizostomeae , собирают для пищи, в основном в Юго -Восточной Азии. [ 131 ] Рукостомы, особенно Rhopilema Esculentum в Китае ( 海蜇 hǎizhe , «морские скаты») и Stomolophus meleagris (медуза пушечного явления) в Соединенных Штатах, предпочитаются из -за их более крупных и более жестких тел и потому, что их токсины безвредны для людей. [ 126 ]
Традиционные методы обработки, выполненные мастером медузы, включают многофазную процедуру от 20 до 40 дней, в которой после удаления гонад и слизистых оболочек зонтик и оральные руки обрабатываются смесью столовой соли и квасцы ,, и сжатый. Обработка делает медузу более суше и кислыми, создавая четкую текстуру. Медзой, подготовленная таким образом, сохраняет 7–10% от их первоначального веса, а обработанный продукт состоит из примерно 94% воды и 6% белка. Свежепрофильная медуза имеет белый, сливочный цвет и становится желтым или коричневым во время длительного хранения. [ 126 ]
В Китае обработанные медузы избавили путем замачивания в воде в течение ночи и едят приготовленную или сырую. Блюдо часто подается измельченным с заправкой масла, соевого соуса, уксуса и сахара или в виде салата с овощами. В Японии вылеченные медузы промывают, разрезают на полоски и подают с уксусом в качестве закуски. [ 126 ] [ 132 ] Опытные, готовые к употреблению продукты также доступны. [ 126 ]
Биотехнология

Плиний Старший сообщил в своей естественной истории , что слизь медузы " Pulmo Marinus " вызывает свет, когда натирал ходьбу. [ 133 ]
В 1961 году Осаму Симомура экстрагировал зеленый флуоресцентный белок (GFP) и другой биолюминесцентный белок, называемый аэкрорин , из большого и обильного гидромедуса aequorea victoria , изучая фотопротеины , которые вызывают биолюминесценцию у этого вида. [ 134 ] Три десятилетия спустя Дуглас Прашер секвенировал и клонировал ген для GFP. [ 135 ] Мартин Чалфи выяснил, как использовать GFP в качестве флуоресцентного маркера генов, вставленных в другие клетки или организмы. [ 136 ] Роджер Циен позже химически манипулировал GFP, чтобы произвести другие флуоресцентные цвета для использования в качестве маркеров. В 2008 году Shimomura, Chalfie и Tsien получили Нобелевскую премию по химии за их работу с GFP. [ 134 ] Чудолетный GFP стал широко используемым в качестве флуоресцентной метки, чтобы показать, какие клетки или ткани экспрессируют специфические гены. Техника генетической инженерии объединяет ген , представляющий интерес для гена GFP. Затем плавленую ДНК помещают в клетку, чтобы генерировать либо клеточную линию, либо (с помощью методов ЭКО ) целое животное, несущее ген. В клетке или животном искусственный ген включается в одних и тех же тканях и в то же время, что и нормальный ген, создавая слияние нормального белка с GFP, прикрепленным к концу, освещение животного или клетки показывает, что ткани экспрессируют этот белок - или на каком этапе развития. Флуоресценция показывает, где экспрессируется ген. [ 137 ]
Аквариум -дисплей

Медузы выставлены во многих общественных аквариумах . Часто фон танка синий, а животные освещаются бок о бок, что увеличивает контраст между животным и фоном. В естественных условиях многие желе настолько прозрачны, что они почти невидимы. [ 138 ] Медузы не адаптированы к закрытым пространствам. Они зависят от течений, чтобы перевозить их с места на место. Профессиональные экспонаты, как и в аквариуме Monterey Bay, имеют точные потоки воды, как правило, в круглых резервуарах, чтобы избежать захвата образцов в углах. Отток распространяется на большую площадь поверхности, и приток попадает как лист воды перед оттоком, поэтому медузы не втягивают в него. [ 139 ] По состоянию на 2009 год медузы стала популярной в домашних аквариумах, где им требуется аналогичное оборудование. [ 140 ]
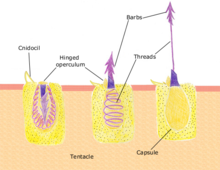
Укусы
Медзой вооружена нематоцистами, типом специализированной жгучей ячейки. [ 141 ] Контакт с щупальцом медузы может вызвать миллионы нематоцист, чтобы проткнуть кожу и вводить яд , [ 142 ] Но только яд какой -то вида вызывает неблагоприятную реакцию у людей. [ 143 ] В исследовании, опубликованном в биологии связи , исследователи обнаружили, что виды медуз, называемые Cassiopea xamachana , который, когда запускается, выпустит крошечные шарики клеток, которые плавают вокруг медузы, укоренивших все на своем пути. Исследователи описали их как «самопрокатирующие микроскопические гранаты» и назвали их Cassiosomes. [ 144 ]
Эффекты укусов варьируются от легкого дискомфорта до сильной боли и смерти. [ 145 ] [ 146 ] Большинство укусов медузы не смертельны, но укусы какую -то Jellyfish ( Irukandji Jellyfish ), такие как морская оса , могут быть смертельными. Укусы могут вызывать анафилаксию (форму шока), которая может быть смертельной. Медзой убивает от 20 до 40 человек в год только на Филиппинах. В 2006 году Испанский Красной Крест обработал 19 000 забитых пловцов вдоль Коста -Бравы . [ 146 ] [ 147 ]
Уксус (3–10% водная уксусная кислота ) может помочь с медуз коробки укусами [ 148 ] [ 149 ] Но не укусы португальского человека войны . [ 148 ] Очистка площади желе и щупальца уменьшает стрельбу из нематоцисты. [ 150 ] Соскабливание пораженной кожи, например, с краем кредитной карты, может удалить оставшиеся нематоцисты. [ 151 ] После того, как кожа была очищена от нематоцист, крем для гидрокортизона, нанесенный локально, уменьшает боль и воспаление. [ 152 ] Антигистаминные препараты могут помочь контролировать зуд . [ 151 ] Антиенины с иммуно -базой используются для серьезных укусов медуз. [ 153 ] [ 154 ]
На острове Эльба и Corsica Dittrichia Viscosa в настоящее время используется жителями и туристами для лечения укусов от медузы, пчел и ошей, нажимая свежие листья на коже с быстрыми результатами.
-
 Box Jellyfish маленькая и ядовитая.
Box Jellyfish маленькая и ядовитая. -
 Медуза дерматит
Медуза дерматит -
 Флаг на пляже Зарауц, предупреждающий о присутствии медузы в воде
Флаг на пляже Зарауц, предупреждающий о присутствии медузы в воде



Механические проблемы
Медзой в больших количествах может заполнять и разделить рыбалку и раздавить захваченную рыбу. [ 155 ] Они могут засорять охлаждающее оборудование, имея инвалидные электростанции в нескольких странах; медуза вызвала каскадную отключение на Филиппинах в 1999 году, [ 146 ] а также повреждение электростанции Diablo Canyon в Калифорнии в 2008 году. [ 156 ] Они также могут остановить опреснительные растения и двигатели кораблей. [ 155 ] [ 157 ]
Смотрите также
- Медуза дерматит
- Список доисторических медузозоанов
- Ocean Sunfish , значительный хищник медузы
- Ctenophora
Примечания
Ссылки
- ^ «Окаменечные записи показывают неуловимую медузу более 500 миллионов лет» . Scienceday . Получено 10 февраля 2023 года .
- ^ Jump up to: а беременный в Энджер, Натали (6 июня 2011 г.). «Гораздо больше, чем плазма и яд» . New York Times . Архивировано из оригинала 18 мая 2013 года . Получено 2 декабря 2011 года .
- ^ Jump up to: а беременный Изабель Родд (20 октября 2020 г.). «Почему медуза может быть« идеальной едой » . BBC News . Получено 7 мая 2023 года .
- ^ Jump up to: а беременный "медуза" . Онлайн этимологический словарь . Получено 9 июня 2018 года .
- ^ Келман, Джанет Харви; Преподобный Теодор Вуд (1910). Морской берег, показан детям . Лондон: TC & EC Джек. п. 146. OL 7043926M .
- ^ Каплан, Юджин Х.; Каплан, Сьюзен Л.; Петерсон, Роджер Тори (август 1999 г.). Полевой гид по коралловым рифам: Карибский бассейн и Флорида . Бостон: Хоутон Миффлин. п. 55. ISBN 978-0-618-00211-5 .
- ^ «Желело для цветов» . 6 апреля 2009 года. Архивировано с оригинала 6 апреля 2009 года . Получено 10 февраля 2023 года .
- ^ "Что такое рыба?" Полем Энциклопедия жизни. Архивировано с оригинала 24 марта 2018 года . Получено 13 октября 2018 года .
И большинство людей знают, что миноги, акулы, лучи, угри, морские коньки и другие странно выглядящие водные существа-рыбы, в то время как моллюски, каракуляра, морские звезды, раки и медуза (несмотря на их имена) не рыбы.
- ^ Бротц, Лукас. Изменение популяций медуз: тенденции в крупных морских экосистемах архивировали 16 апреля 2013 года на машине Wayback . 2011. С.1.
- ^ Coulombe, Дебора А. (14 февраля 1990 г.). Приморский натуралист: руководство по обучению на побережье . Саймон и Шустер . п. 60. ISBN 978-0-671-76503-3 Полем Архивировано из оригинала 31 декабря 2013 года . Получено 20 марта 2013 года .
- ^ Клаппенбах, Лора. «Десять фактов о медузе» . Архивировано из оригинала 26 февраля 2009 года . Получено 24 января 2010 года .
- ^ Липтон, Джеймс (1991). Возвышение жаворонков . Викинг. ISBN 978-0-670-30044-0 .
- ^ Maciver, Angus (2004). Первая помощь на английском языке . Ходдер Гибсон (Хахетте). ISBN 978-1-444-19376-3 .
- ^ Картрайт, Полин; Halgedahl, Susan L .; Хендрикс, Джонатан Р.; и др. (2007). Хамфрис, Стюарт (ред.). «Исключительно сохранившие медузы из среднего кембрия» . Plos один . 2 (10): E1121. Bibcode : 2007ploso ... 2.1121c . doi : 10.1371/journal.pone.0001121 . ISSN 1932-6203 . PMC 2040521 . PMID 17971881 .
- ^ "Медуза" . Мерриам-Уэбстер . 1 сентября 2018 года . Получено 11 сентября 2018 года .
- ^ Jump up to: а беременный в дюймовый и фон глин час я «Пятницы медуз | виды медузы» . Ориентированные на политику морские экологические исследования в южной европейской морях (Персей) . Получено 28 августа 2018 года .
- ^ Jump up to: а беременный Миллс, CE (8 ноября 2010 г.). "Ctenophores" . Университет Вашингтона . Получено 28 августа 2018 года .
- ^ Jump up to: а беременный «Наши железные родственники, похожие на желе: общие заблуждения о Salps» . Программа Nereus . Получено 28 августа 2018 года .
- ^ Jump up to: а беременный Запата, Фелипе; Гетц, Фрейя Э.; Смит, Стивен А.; и др. (2015). «Филогеномный анализ подтверждает традиционные отношения в Cnidaria» . Plos один . 10 (10): E0139068. BIBCODE : 2015PLOSO..1039068Z . doi : 10.1371/journal.pone.0139068 . PMC 4605497 . PMID 26465609 .
- ^ Каял, Эсан; Бентлаж, Бастиан; Сабрина Панки, М.; и др. (2018). «Филогеномика обеспечивает надежную топологию основных линий Cnidarian и понимания происхождения ключевых признаков организма» . BMC Эволюционная биология . 18 (1): 68. Bibcode : 2018bmcee..18 ... 68K . doi : 10.1186/s12862-018-1142-0 . PMC 5932825 .
- ^ «StauroMedusae UK, онлайн -гид по преследуемым медузам (Stauromedusae), найденную в прибрежных водах Соединенного Королевства и Ирландии. Включает в себя заметки об их идентификации, и где и как их найти. Back UK CHECTLIST для преследуемых медузы (Stauromedusae)» . StauroMedusae UK . Получено 28 августа 2018 года .
- ^ Ширвотер, Бернд; Хелм, Ребекка Р.; Данн, Кейси В. (2017). «Индолы вызывают метаморфозу в широком разнообразии медузы, но не в железной желе (коронаты)» . Plos один . 12 (12): E0188601. BIBCODE : 2017PLOSO..1288601H . doi : 10.1371/journal.pone.0188601 . PMC 5744923 . PMID 29281657 .
- ^ Осборн, К.Дж. (2014). "Красная бумажная медуза" . Смитсоновский институт . Получено 13 октября 2018 года .
- ^ Дейли, Джейсон (1 марта 2017 г.). «Посмотрите на завораживающую« космическую медузу » . Смитсоновский институт . Получено 28 августа 2018 года .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л м не а п Q. ведущий с Т Рупперт, Эдвард Э.; Fox, Richard, S.; Барнс, Роберт Д. (2004). Зоология беспозвоночных, 7 -е издание . Cengage Learning. С. 148–174. ISBN 978-81-315-0104-7 .
{{cite book}}: Cs1 maint: несколько имен: список авторов ( ссылка ) - ^ Jump up to: а беременный Cnidaria Archived 2012-09-21 в The Wayback Machine , Tree of Life.
- ^ Маркес, AC; Ag Collins (2004). «Кладистический анализ Медусозоиа и эволюции Cnidarian». БИОЛОГИЯ БЕЗОПАСНОСТИ . 123 : 23–42. doi : 10.1111/j.1744-7410.2004.tb00139.x . S2CID 28342963 .
- ^ Kramp, PL (1961). «Синопсис медалей мира». Журнал Морской биологической ассоциации Соединенного Королевства . 40 : 1–469. Bibcode : 1961jmbuk..40 .... 7k . doi : 10.1017/s0025315400007347 (неактивный 27 августа 2024 г.).
{{cite journal}}: CS1 Maint: doi неактивен с августа 2024 года ( ссылка ) - ^ Луна, Джастин; Карон, Жан-Бернард; Мойсик, Джозеф (9 августа 2023 г.). «Макроскопическая Медуза, свободно плавающая от Средней Камбрийской сланцы Берджесс» . Труды Королевского общества B: Биологические науки . 290 (2004). doi : 10.1098/rspb.2022.2490 . ISSN 0962-8452 . PMC 103944413 . PMID 37528711 .
- ^ Ши, Юн-Хва; Рудло, Джек (1994). «Потенциал использования медузы в качестве пищи в западных странах». Тенденции в области пищевых наук и техники . 5 (7): 225–229. doi : 10.1016/0924-2244 (94) 90253-4 .
- ^ «Медзой - визуальный словарь» . Infovisual.info . Получено 10 февраля 2023 года .
- ^ Вагонер, Бен; Коллинз, Аллен Г. "Кубозоа: подробнее о морфологии" . Калифорнийский музей палеонтологии . Получено 6 января 2019 года .
- ^ «Нематоцисты» . Jellieszone . 2 апреля 2015 года. Архивировано с оригинала 2 апреля 2015 года . Получено 29 марта 2014 года .
- ^ Kier, William (2012). «Разнообразие гидростатических скелетов» . Журнал экспериментальной биологии . 215 (Pt 8): 1247–1257. doi : 10.1242/jeb.056549 . PMID 22442361 .
- ^ Саттерли, Р.А. (2002). «Нейрональный контроль плавания в медузе: сравнительная история» . Канадский журнал зоологии . 80 (10): 1654–1669. doi : 10.1139/Z02-138 . Архивировано из оригинала (PDF) 12 июля 2013 года.
- ^ Кацуки, Такео; Greenspan, Ralph J. (2013). «Медзой нервные системы» . Текущая биология . 23 (14): R592 - R594. Bibcode : 2013cbio ... 23.r592k . doi : 10.1016/j.cub.2013.03.057 . PMID 23885868 .
- ^ Саттерли, Ричард А. (2011). "У медузы есть центральные нервные системы?" Полем Журнал экспериментальной биологии . 214 (8): 1215–1223. doi : 10.1242/jeb.043687 . PMID 21430196 .
- ^ Wehner, R. (2005). «Сенсорная физиология: безмозглые глаза» (PDF) . Природа . 435 (7039): 157–159. Bibcode : 2005natur.435..157W . doi : 10.1038/435157a . PMID 15889076 . S2CID 4408533 . Архивировано (PDF) из оригинала 29 июля 2013 года.
- ^ «Мультигластная медуза помогает с загадкой Дарвина» . Новый ученый . Получено 10 февраля 2023 года .
- ^ Jump up to: а беременный в дюймовый и фон Нильссон, DE (2013). «Эволюция глаз и ее функциональная основа» . Визуальная нейробиология . 30 (1–2): 5–20. doi : 10.1017/s0952523813000035 . PMC 3632888 . PMID 23578808 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л м не а п Гарм, Андерс; Ekström, Peter (2010). «Доказательства для нескольких фотосистем в медузе». Глава 2 - Доказательства для нескольких фотосистем в медузе . Тол. 280. Академическая пресса. С. 41–78. doi : 10.1016/s1937-6448 (10) 80002-4 . ISBN 9780123812605 Полем ISSN 1937-6448 . PMID 20797681 .
{{cite book}}:|journal=игнорируется ( помощь ) - ^ Suga H, Tschopp P, Graziussi DF, et al. (10 августа 2010 г.). «Гибко развернутые гены PAX в развитии глаз при ранней эволюции животных, продемонстрированных исследованиями гидрозой -медузы» . ПНА . 107 (32): 14263–14268. Bibcode : 2010pnas..10714263S . doi : 10.1073/pnas.1008389107 . PMC 2922549 . PMID 20660753 .
- ^ Jump up to: а беременный в дюймовый и фон О'Коннор М., Гарм А., Нильссон Д.Е. (2009). «Структура и оптика глаз коробки медузы Chiropsella Bronzie». Журнал сравнительной физиологии а . 195 (6): 557–569. doi : 10.1007/s00359-009-0431-x . PMID 19347342 . S2CID 9563849 .
- ^ Nilsson DE, Gislén L, Coates M, et al. (2005). «Расширенная оптика в глазном медузе». Природа . 435 (7039): 201–205. Bibcode : 2005natur.435..201n . doi : 10.1038/nature03484 . PMID 15889091 . S2CID 4418085 .
- ^ Jump up to: а беременный в дюймовый Piatigorsky J, Kozmik Z (2004). «Cubozoan Jellyfish: модель Evo/Devo для глаз и других сенсорных систем» . Инт. J. Dev. Биол . 48 (8–9): 719–729. doi : 10.1387/ijdb.041851jp . PMID 15558464 .
- ^ Jump up to: а беременный в Грей Г.С., Мартин В.Дж., Саттерли Р.А. (август 2009 г.). «Ультраструктура синапсов сетчатки у кубозоан» (PDF) . Биол Булл . 217 (1): 35–49. doi : 10.1086/bblv217n1p35 . PMID 19679721 . S2CID 24400231 .
- ^ Jump up to: а беременный Бергер, Эдвард В. (1898). «Гистологическая структура глаз Cubomedusae» . Журнал сравнительной неврологии . 8 (3): 223–230. doi : 10.1002/cne.910080317 . S2CID 85422599 .
- ^ Суга, Хироши; Шмид, Волкер; Gehring, Walter J. (2008). «Эволюция и функциональное разнообразие медуз опсинов» . Текущая биология . 18 (1): 51–55. Bibcode : 2008cbio ... 18 ... 51 с . doi : 10.1016/j.cub.2007.11.059 . ISSN 0960-9822 . PMID 18160295 . S2CID 13344739 .
- ^ Jump up to: а беременный в дюймовый Piatigorsky J, Horwitz J, Kuwabara T, Cutress C (1989). «Клеточная глазная линза и кристаллины кубомедусанской медузы». Журнал сравнительной физиологии а . 164 (5): 577–587. doi : 10.1007/bf00614500 . PMID 2565398 . S2CID 19797109 .
- ^ Jump up to: а беременный в Сеймур, Джейми Э.; О'Хара, Эмили П. (2020). «Ответ зрачка на свет у трех видов Cubozoa (ящик -медуза)» . Планктон и бентос исследования . 15 (2): 73–77. doi : 10.3800/pbr.15.73 . S2CID 219759193 .
- ^ Jump up to: а беременный Миллс, CE; Хирано, YM (2007). Энциклопедия приливов и скалистых берегов: Hydromedusae . Калифорнийский университет. С. 286–288. ISBN 978-0-520-25118-2 .
- ^ Mills, CE (1976). « Podocoryne Selena , новый вид Hydroid из Мексиканского залива, и сравнение с гидрактинией Echinata » . Биологический бюллетень . 151 (1): 214–224. doi : 10.2307/1540715 . JSTOR 1540715 .
- ^ Костелло Дж. (1988). «Лабораторная культура и кормление гидромедуса Cladonema californicum Hyman (Anthemedusa: Cladonemidae)». Журнал экспериментальной морской биологии и экологии . 123 (2): 177–188. Bibcode : 1988jembe.123..177c . doi : 10.1016/0022-0981 (88) 90168-2 .
- ^ «Редкое наблюдение за медузой гривы льва в заливе Трамор» . Уотерфорд сегодня. 1 августа 2007 года. Архивировано из оригинала 30 мая 2010 года . Получено 18 октября 2010 года .
- ^ «Медзока гривы льва - справочная библиотека» . Redorbit. 12 июня 2003 года. Архивировано с оригинала 30 июля 2010 года . Получено 18 октября 2010 года .
- ^ «150 Убил медузы на Ржи -Бич» . Wmur.com. 21 июля 2010 г. Архивировано с оригинала 14 октября 2011 года . Получено 11 июня 2018 года .
- ^ Omori, Makoto; Китамура, Минору (2004). «Таксономический обзор трех японских видов съедобной медузы (Scyphozoa: Rhizostomeae)» (PDF) . Биология и экология планктона . 51 (1): 36–51. Архивировано (PDF) из оригинала 23 марта 2012 года.
- ^ Uye, Shin-Ichi (2008). «Цветы гигантской медузы Nemopilema nomurai : угроза для устойчивости рыболовства восточноазиатских маргинальных морей» (PDF) . Plankton & Benthos Research . 3 (Дополнение): 125–131. doi : 10.3800/pbr.3.125 . Архивировано (PDF) из оригинала 16 мая 2013 года.
- ^ «Гигантская эчизен медуза у побережья Японии» . Би -би -си. 30 ноября 2009 г. Архивировано с оригинала 1 января 2011 года.
- ^ Kramp, PL (1961). «Синопсис медалей мира». Журнал Морской биологической ассоциации Соединенного Королевства . 40 : 1–469. Bibcode : 1961jmbuk..40 .... 7k . doi : 10.1017/s0025315400007347 (неактивный 27 августа 2024 г.).
{{cite journal}}: CS1 Maint: doi неактивен с августа 2024 года ( ссылка ) - ^ Буртон, Джоди (23 апреля 2010 г.). «Гигантская глубокая море медуза снималась в Мексиканском заливе» . BBC Earth News . Архивировано из оригинала 5 июля 2010 года.
- ^ «Фотографии антарктического гигантского желе (Desmonema glaciale) • Inaturalist» .
- ^ Лига, Майкл (11 октября 2011 г.). «Путь покончить с погружением» . Polartrec . Станция МакМердо , Антарктида. Архивировано из оригинала 13 июня 2023 года . Получено 13 июня 2023 года .
- ^ «Дайвинг под водой с гигантской медузой» . 26 апреля 2010 года.
- ^ «Как воспроизводят медузы? Какое влияние оказывает их укус на людей? В чем разница между красной и полупрозрачной медузой?» Полем Scientific American . 15 октября 2013 года. Архивировано с оригинала 23 октября 2013 года . Получено 22 октября 2013 года .
- ^ Миллс, Клаудия (1983). «Паттерны вертикальной миграции и активности DIL Hydromedusae: исследования в большом аквариуме». Журнал Plankton Research . 5 (5): 619–635. doi : 10.1093/plankt/5.5.619 . S2CID 13914997 .
- ^ Епископ, Эндрю. «Лунное желе ( Аурелия Аурита )» . Морские беспозвоночные Бермудских островов . Получено 11 июня 2018 года .
- ^ Jump up to: а беременный в Mills, CE (1987). J. Bouillon; Ф. Буэро; Ф. Цикогна; PFS Cornelius (ред.). На месте и корабельные исследования живых гидромедусов и гидроидов: предварительные наблюдения за адаптацией жизненного цикла к открытому океану . Кларендон Пресс. ISBN 978-0-19-857190-2 .
{{cite book}}:|work=игнорируется ( помощь ) - ^ Featkes, J. Walter (1887). «Гидоид паразита на рыбе» . Природа . 36 (939): 604–605. Bibcode : 1887natur..36..604f . doi : 10.1038/036604b0 . S2CID 4078889 .
- ^ Шухерт, Питер. «Гидрозоа» . Архивировано из оригинала 4 февраля 2010 года . Получено 24 января 2010 года .
- ^ «Как растут медузы, от яиц до полипов до медуз» . Мыслить . Получено 10 февраля 2023 года .
- ^ Хьюз, Клэр. «Жизненный цикл коробки медузы» . Artforlibraries.org . Архивировано с оригинала 4 марта 2016 года . Получено 2 января 2016 года .
- ^ Бруска, Ричард (2016). Беспозвоночные . Sinauer Associates. п. 310. ISBN 978-1-60535-375-3 .
- ^ Миглиетта, депутат; Piraino, S.; Kubota, S.; Schuchert, P. (2007). «Виды в роде Turritopsis (Cnidaria, Hydrozoa): молекулярная оценка» . Журнал зоологической систематики и эволюционных исследований . 45 (1): 11–19. doi : 10.1111/j.1439-0469.2006.00379.x .
- ^ Piraino, S.; Boero, F.; Aeschbach, B.; Шмид, В. (1996). «Изменение жизненного цикла: преобразование медуз в полипы и трансдифференцировку клеток в турритопсисе nutriculaula (Cnidaria, гидрозоа)». Биологический бюллетень . 190 (3): 302–312. doi : 10.2307/1543022 . JSTOR 1543022 . PMID 29227703 . S2CID 3956265 .
- ^ Рати, Акшат (15 мая 2014 г.). «Медзой-наиболее энергоэффективные пловцы, подтверждает новая метрика» . Ars Technica . Архивировано с оригинала 3 ноября 2014 года . Получено 3 декабря 2014 года .
- ^ Gemmell, BJ; Костелло, JH; Колин, SP; и др. (2013). «Пассивная энергия в медузах способствует двигательному преимуществу по сравнению с другими метазоанами» . Труды Национальной академии наук . 110 (44): 17904–17909. BIBCODE : 2013PNAS..11017904G . doi : 10.1073/pnas.1306983110 . PMC 3816424 . PMID 24101461 .
- ^ Бруска, Ричард (2016). Беспозвоночные . Sinauer Associates. п. 296. ISBN 978-1-60535-375-3 Полем
Все Cnidarians - плотоядные (или паразиты). Как правило, насыщенные нематоцистами, кормившие щупальца, захватывают добычу животных и несут ее в область рта, где он проглатывается целым.
- ^ «Большая медуза унаследовала океан, находит исследование» . msnbc.com . 15 сентября 2011 года. Архивировано с оригинала 14 июля 2014 года . Получено 3 декабря 2014 года .
- ^ Дэвис, Ch; Slotwinski, как «Австралийская морская зоопланктона-мука, кладоцеранцы» (PDF) . Институт морских и антарктических исследований, Университет Тасмании . Получено 12 октября 2018 года .
- ^ Доусон, Майкл Н. (2000). «Измененные мезокосмы в качестве альтернативы для береговых планктонкрезелей: заметки о землепольни из медуз из морских озер» . Журнал Plankton Research . 22 (9): 1673–1682. doi : 10.1093/plankt/22.9.1673 .
- ^ Jump up to: а беременный Доусон, Майк Н.; Мартин, Лора Э.; Лолита К, Лолита К.; Пенланд (май 2001 г.). Медзой роя, туристы и Христос-Кильд . Тол. 451. Springer. С. 131–144. doi : 10.1023/a: 1011868925383 . ISBN 978-0-7923-6964-6 Полем S2CID 1679552 .
{{cite book}}:|journal=игнорируется ( помощь ) - ^ Немо, Лесли (13 февраля 2020 года). «Ядовитый сопли помогает этим медузам без щупальца» . Обнаружить . Получено 11 июля 2021 года .
- ^ Инь, Стеф (29 сентября 2017 г.). «Кто ест медузу? Пингвины, вот кто» . New York Times . Архивировано с оригинала 1 октября 2017 года . Получено 4 октября 2017 года .
- ^ Тибот, Жан-Батист; Арноулд, Джон Пи; Гомес-Лайх, Агустина; и др. (2017). «Медзовика и другие желания в качестве пищи для четырех видов пингвинов-понимание видео, передаваемых хищниками». Границы в экологии и окружающей среде . 15 (8): 437–441. Bibcode : 2017free ... 15..437t . doi : 10.1002/плата. 1529 . S2CID 90152409 .
- ^ Гершвин, Лиза-Энн (2016). Медуза: естественная история . Университет Чикагской Прессы. п. 140. ISBN 978-0-226-28767-6 .
- ^ Гершвин, Лиза-Энн (2013). Удар ! Университет Чикагской Прессы. С. 274 -. ISBN 978-0-226-02010-5 Полем Архивировано с оригинала 6 августа 2016 года.
- ^ Colugo7 (2006). "Медзой" . Веб -проект Tree of Life . Архивировано с оригинала 12 июня 2018 года . Получено 7 июня 2018 года .
{{cite web}}: CS1 Maint: числовые имена: список авторов ( ссылка ) - ^ Гриффин, Дубос Б.; Мерфи, Томас М. "Медзока пушкина" (PDF) . Департамент природных ресурсов Южной Каролины. Архивировано из оригинала (PDF) 19 января 2008 года . Получено 7 июня 2018 года .
- ^ Jump up to: а беременный Броц, Лукас; Чеунг, Уильям В.Л.; Клейснер, Кристин; и др. (2012). «Увеличение популяций медуз: тенденции в крупных морских экосистемах» . Гидробиология . 688 : 3–20. doi : 10.1007/s10750-012-1039-7 .
- ^ Джилл, Виктория. «Медуза» может почувствовать океанские течения » . BBC News. Архивировано с оригинала 26 января 2015 года . Получено 26 января 2015 года .
- ^ Хейс, Грэм С. (2017). «Океанские течения и морская жизнь» . Текущая биология . 27 (11): R470 - R473. Bibcode : 2017cbio ... 27.r470h . doi : 10.1016/j.cub.2017.01.044 . HDL : 10536/dro/du: 30100274 . PMID 28586681 .
- ^ Шубин, Кристи (10 декабря 2008 г.). «Антропогенные факторы, связанные с цветами медуз - окончательный проект II» . Тропические полевые курсы: Западная Программа: Университет Майами. Архивировано из оригинала 14 июня 2010 года . Получено 19 ноября 2009 года .
- ^ "Что такое мертвая зона?" Полем Национальная служба океана . Получено 3 октября 2018 года .
- ^ Йонг, изд (6 июня 2011 г.). «Медзой сдвигает океанскую пищевую сеть, кормив бактерии слизью и экскрементами» . Откройте для себя журнал. Архивировано с оригинала 6 ноября 2018 года . Получено 3 октября 2018 года .
- ^ Jump up to: а беременный «Цветение медуз может быть признаком больных морей» . www.eurocbc.org . Получено 10 февраля 2023 года .
- ^ Hays, GC; Bastian, T.; Дойл, Тк; и др. (2011). «Высокая активность и поиски Леви: медуза может искать в толще воды, как рыба» (PDF) . Труды Королевского общества б . 279 (1728): 465–473. doi : 10.1098/rspb.2011.0978 . PMC 3234559 . PMID 21752825 . Архивировано (PDF) из оригинала 25 декабря 2012 года.
- ^ Pauly, D.; Кристенсен, В.; Dalsgaard, J.; и др. (1998). «Рыбалка по морской пищевой сети» (PDF) . Наука . 279 (5352): 860–863. Bibcode : 1998sci ... 279..860p . doi : 10.1126/science.279.5352.860 . PMID 9452385 . Архивировано (PDF) из оригинала 10 июля 2012 года.
- ^ Ричардсон, AJ; Бакун, А.; Hays, GC; Gibbons, MJ (2009). «Jellyfish Joyride: причины, последствия и ответы управления на более желатиновое будущее» (PDF) . Тенденции в экологии и эволюции . 24 (6): 312–322. Bibcode : 2009tecoe..24..312r . doi : 10.1016/j.tree.2009.01.010 . PMID 19324452 . [ Постоянная мертвая ссылка ]
- ^ Aksnes, DL; Nejstgaard, J.; Sædberg, E.; Sørnes, T. (2004). «Оптический контроль населения рыбы и зоопланктона» . Лимнология и океанография . 49 (1): 233–238. Bibcode : 2004limoc..49..233a . doi : 10.4319/lo.2004.49.1.0233 .
- ^ Линам, CP; Гиббонс, MJ; Аксельсен, будь; и др. (2006). «Медзой обогнала рыбу в тяжелой ловковой экосистеме» (PDF) . Текущая биология . 16 (13): 492–493. Bibcode : 2006cbio ... 16.r492l . doi : 10.1016/j.cub.2006.06.018 . PMID 16824906 . S2CID 62793057 . Архивировано (PDF) из оригинала 23 марта 2011 года.
- ^ Pauly, D.; Грэм, W.; Libralato, S.; и др. (2009). «Медзой в экосистемах, онлайн -базах данных и экосистемных моделях» . Гидробиология . 616 : 67–85. doi : 10.1007/s10750-008-9583-x . S2CID 12415790 . Архивировано из оригинала (PDF) 12 июля 2013 года.
- ^ Доусон, Миннесота; Sen Gupta, A.; Англия, MH (2005). «Связанная биофизическая мировая модель океана и молекулярные генетические анализы идентифицируют множественные введения криптогенных видов» . Прокурор Нат. Академический Наука США . 102 (34): 11968–73. Bibcode : 2005pnas..10211968d . doi : 10.1073/pnas.0503811102 . PMC 1189321 . PMID 16103373 .
- ^ Доусон, М.Н. (2003). «Макроморфологические различия среди загадочных видов лунной медузы, Aurelia (Cnidaria: Scyphozoa)». Морская биология . 143 (2): 369–79. Bibcode : 2003marbi.143..369d . doi : 10.1007/s00227-003-1070-3 . S2CID 189820003 .
- ^ Jump up to: а беременный Ринат, Зафрир (15 июня 2009 г.). «Самая инвазивная медуза в мире, распространяющаяся вдоль Израильского побережья» . Хаарец . Получено 13 октября 2018 года .
- ^ Бриан, Фредерик; Boero, Ferdinando (2001). «Гелатиновые вспышки зоопланктона - обзор цветов медуз» . Ciesm Monographs . 14 : 5-17.
- ^ Mills, CE (2001). «Цветение медуз: увеличивается ли популяции во всем мире в ответ на изменение условий океана?» (PDF) . Гидробиология . 451 : 55–68. doi : 10.1023/a: 1011888006302 . S2CID 10927442 . Архивировано (PDF) из оригинала 3 марта 2016 года.
- ^ "Ciesm GIS" . www.ciesm.org . Получено 10 февраля 2023 года .
- ^ Purcell, J.; Арай, М. (2001). Purcell, J. E; Грэм, В. М; Dumont, H. J (Eds.). «Взаимодействие пелагических Cnidarians и Ctenophores с рыбой: обзор». Гидробиология . 541 : 27–44. doi : 10.1007/978-94-010-0722-1 . ISBN 978-94-010-3835-5 Полем S2CID 27615539 .
- ^ Brodeur, Richard D.; Ссылка, Джейсон С.; Смит, будь; и др. (2016). «Экологические и экономические последствия игнорирования медузы: просьба о повышении мониторинга экосистем». Рыболовство . 41 (11): 630–637. Bibcode : 2016 Рыбка ... 41..630B . doi : 10.1080/03632415.2016.1232964 .
- ^ Рузицка, JJ; Brodeur, Rd; Emmett, RL; и др. (2012). «Межгодовая изменчивость в текущей структуре пищевой сети в Северной Калифорнии: изменения путей потока энергии и роль кормовой рыбы, эуфаузиидов и медуз». Прогресс в океанографии . 102 : 19–41. Bibcode : 2012proce.102 ... 19r . doi : 10.1016/j.pocean.2012.02.002 .
- ^ Jump up to: а беременный Питт, Кайли; Уэльс, Дэвид; Кондон, Роберт (январь 2009 г.). «Влияние цветов медузы на углерод, азот и фосфор езда на велосипеде и производство планктона». Гидробиология . 616 : 133–149. doi : 10.1007/S10750-008-9584-9 . S2CID 22838905 .
- ^ Кондон, Роберт Х.; Дуарте, Карлос М.; Питт, Кайли А.; и др. (15 января 2013 г.). «Рецидивирующие цветы медузы являются следствием глобальных колебаний» . Труды Национальной академии наук . 110 (3): 1000–1005. Bibcode : 2013pnas..110.1000c . doi : 10.1073/pnas.1210920110 . PMC 3549082 . PMID 23277544 .
- ^ Линам, Кристофер П.; Гиббонс, Марк Дж.; Axelsen, Bjørn E.; и др. (11 июля 2006 г.). «Медзой обогнала рыбу в тяжелой рыбаленной экосистеме» . Текущая биология . 16 (13): R492–493. Bibcode : 2006cbio ... 16.r492l . doi : 10.1016/j.cub.2006.06.018 . PMID 16824906 . S2CID 62793057 .
- ^ Масиламани, J; Десудосс, K; Канавиллл, Нандакумар; и др. (10 сентября 2000 г.). «Вход в медуз: угроза гладкой эксплуатации прибрежных электростанций» . Текущая наука . 79 : 567–569.
- ^ Purcell, Jennifer E.; Uye, Shin-ichi; Ло, Вен-Ценг (22 ноября 2007 г.). «Антропогенные причины цветения медуз и их прямые последствия для людей: обзор» . Серия прогресса в морской экологии . 350 : 153–174. Bibcode : 2007meps..350..153p . doi : 10.3354/meps07093 .
- ^ Свитман, Эндрю К.; Смит, Крейг Р.; Дейл, Трин; Джонс, Даниэль ОБ (2014). «Быстрое поглощение туши медузы показывает важность желатинового материала для глубоководных пищевых сетей» . Труды Королевского общества B: Биологические науки . 281 (1796): 20142210. DOI : 10.1098/rspb.2014.2210 . PMC 4213659 . PMID 25320167 .
- ^ Лебрато, Марио; Пахлоу, Маркус; Ослис, Андреас; и др. (2011). «Долганирование глубины экспорта органического вещества, связанного с желе -водопадом» (PDF) . Лимнология и океанография . 56 (5): 1917–1928. Bibcode : 2011limoc..56.1917L . doi : 10.4319/lo.2011.56.5.1917 . HDL : 10072/43275 . S2CID 3693276 .
- ^ Didžiulis, Viktoras. «Инвазивные виды инопланетянина: Craspedacusta Sowerbyi » (PDF) . Нобанис. Архивировано (PDF) из оригинала 17 мая 2014 года . Получено 16 июня 2016 года .
- ^ Доусон, Майк Н.; Мартин, Лора Э.; Пенланд, Лолита К. (2001). Медзой роя, туристы и Христос-Кильд . Тол. 451. С. 131–144. doi : 10.1023/a: 1011868925383 . ISBN 978-0-7923-6964-6 Полем S2CID 1679552 .
{{cite book}}:|journal=игнорируется ( помощь ) - ^ Миллс, CE; Land, YM (2007). "Стюрумдудус" 541–543 .
- ^ Кондо, Юсуке; Оцука, Сусуму; Хирабаяши, Такеши; и др. (2016). «Сезонные изменения в инфекции с видами трематодов, использующие медузы в качестве хозяев: свидетельство передачи на точную рыбу -хозяина посредством медалистика» . Паразит . 23 : 16. doi : 10.1051/parasite/2016016 . PMC 4824873 . PMID 27055563 . Архивировано с оригинала 4 ноября 2017 года.
- ^ Леунг, Томми (26 мая 2016 г.). " Opechona olssoni " . Блог: паразит дня. Архивировано с оригинала 30 июня 2016 года . Получено 1 июня 2016 года .
- ^ "FAOSTAT" . www.fao.org . Получено 10 февраля 2023 года .
- ^ Jump up to: а беременный в дюймовый и Ши, да. Пегги; Леонг, Фуи-Мин; Рудло, Джек (2001). "Медуза как еда". Гидробиология . 451 (1–3): 11–17. doi : 10.1023/a: 1011875720415 . S2CID 20719121 .
- ^ Джордж, Алета (1 ноября 2012 г.). «Челеты в центре внимания» . Эндокринные новости . Эндокринное общество . Получено 16 июня 2018 года .
- ^ Клинтон, Мораг; Ферье, Дэвид Эк; Мартин, Сэмюэль А.М.; Брирли, Эндрю С. (2 апреля 2021 года). Байрон, Кэрри (ред.). «Воздействие медузы на аквакультуру морской клетки: обзор существующих знаний и проблемы для здоровья рыбы». ICES Journal of Marine Science . 78 (5). Международный совет по разведке моря ( OUP ): 1557–1573. doi : 10.1093/icesjms/fsaa254 . HDL : 10023/25136 . ISSN 1054-3139 .
- ^ Bosch-Belmar, Mar; Милисенда, Джакомо; Бассо, Лорена; и др. (3 сентября 2020 г.). «Медуриза влияет на морскую аквакультуру и рыболовство» . Отзывы в области рыболовства Science & Aquaculture . 29 (2). Тейлор и Фрэнсис : 242–259. doi : 10.1080/23308249.2020.1806201 . HDL : 10447/531140 . ISSN 2330-8249 . S2CID 225330785 .
- ^ Аристотель ; Уильям Огл (транс.) (2018). Части животных . Тол. IV п. 6. ISBN 9782378989842 . [ Постоянная мертвая ссылка ]
- ^ Omori, M.; Нет, E. (2001). "Рыба -медуз Гидробиология 451 : 19–2 doi : 10.1023/a: 101189821323 . S2CID 6518460 .
- ^ FIRTH, FE (1969). Энциклопедия морских ресурсов . Ван Ностренд Рейнхольд. ISBN 978-0-442-22399-1 .
- ^ «Как желе получило свое сияние» . Американский музей естественной истории . Архивировано с оригинала 12 июня 2018 года . Получено 11 июня 2018 года .
- ^ Jump up to: а беременный Shimomura, O.; Джонсон, FH; Saiga, Y. (1962). «Экстракция, очистка и свойства аэворина, биолюминесцентный белок из светящегося гидромедусана, aequorea ». Журнал клеточной и сравнительной физиологии . 59 (3): 223–39. doi : 10.1002/jcp.1030590302 . PMID 13911999 .
- ^ Прашер, округ Колумбия; Eckenrode, VK; Уорд, WW; и др. (1992). «Основная структура зеленого флуоресцентного белка Aequorea Victoria». Ген . 111 (2): 229–33. doi : 10.1016/0378-1119 (92) 90691-H . PMID 1347277 .
- ^ Chalfie, M.; Tu, y.; Euskirchen, G.; и др. (Февраль 1994 г.). «Зеленый флуоресцентный белок как маркер для экспрессии генов». Наука . 263 (5148): 802–5. Bibcode : 1994sci ... 263..802c . doi : 10.1126/science.8303295 . PMID 8303295 . S2CID 9043327 .
- ^ Pieribone, v.; Грубер, Д.Ф. (2006). Аглоу в темноте: революционная наука о биофлуоресценции . Гарвардский университет издательство. ISBN 978-0-674-02413-7 .
- ^ Херринг, Питер (2002). Биология издательства глубокого океана Оксфордского университета . Издательство Оксфордского университета. С. 190–191 . ISBN 978-0-19-854956-7 .
- ^ «Патент США на танк медуз» . Архивировано из оригинала 20 февраля 2015 года.
- ^ Ричтел, Мэтт (14 марта 2009 г.). «Как избежать сжигания твоей медузы» . New York Times . Архивировано из оригинала 26 марта 2010 года . Получено 6 мая 2010 года .
- ^ МакГи, Ричард Дж.; Вебстер, Анжела С .; Льюис, Шарон Р.; Уэлсфорд, Мишель (5 июня 2023 г.). «Вмешательства для симптомов и признаков, возникающих в результате укусов медуз» . Кокрановская база данных систематических обзоров . 2023 (6): CD009688. doi : 10.1002/14651858.cd009688.pub3 . ISSN 1469-493X . PMC 10240560 . PMID 37272501 .
- ^ Purves, WK; Садава, Д.; Orians, GH; Хеллер, HC 1998. Жизнь. Наука биологии. Часть 4: Эволюция разнообразия. Глава 31
- ^ «Танки с медузами и живая медуза домашних животных для продажи в Jellyfish Art - купить медузы и танки с медуза» . jellyfishart.com . Архивировано с оригинала 2 марта 2012 года . Получено 3 декабря 2014 года .
- ^ Джаймо, Кара (13 февраля 2020 года). «Вы не трогали эту медузу, но они могут укусить вас крошечными гранатами» . New York Times . ISSN 0362-4331 . Получено 27 февраля 2020 года .
- ^ Махон, Эндрю; Маллинсон, Том Э (2020). "Медзовый жалки льва". Международная практика фельдшера . 10 (2): 46–48. doi : 10.12968/ippr.2020.10.2.46 . ISSN 2052-4889 . S2CID 225837493 .
- ^ Jump up to: а беременный в Такер, Эбигейл (июль 2010 г.). «Новый король моря». Смитсоновский институт . 54 (4): 540–561. doi : 10.1177/1363461517722869 . PMID 28752797 . S2CID 12532183 .
- ^ Адамс, Джули (13 августа 2016 г.). "Box Jellyfish: Почему они такие смертоносные?" Полем Наша прекрасная планета. Архивировано с оригинала 10 сентября 2016 года . Получено 17 сентября 2016 года .
- ^ Jump up to: а беременный Fenner, P.; Уильямсон, Дж.; Бернетт, Дж.; Рифкин Дж. (1993). «Первая помощь лечения укусов медуз в Австралии. Реакция на недавно дифференцированные виды». Медицинский журнал Австралии . 158 (7): 498–501. doi : 10.5694/j.1326-5377.1993.tb137588.x . PMID 8469205 . S2CID 42453046 .
- ^ Currie, B.; Хо, с.; Alderslade, P. (1993). «Ящик-мученика, кока-кола и старое вино». Медицинский журнал Австралии . 158 (12): 868. doi : 10.5694/j.1326-5377.1993.tb137688.x . PMID 8100984 . S2CID 19857333 .
- ^ Harwick, R.; Калланан, В.; Уильямсон, Дж. (1980). Netarming the Box- : медуза Медицинский журнал Австралии 1 (1): 15–2 doi : 10.5694/j.1326-537.1980 . PMID 6102347 . S2CID 204054168 .
- ^ Jump up to: а беременный Перкинс, Р.; Морган С. (2004). «Отравление, энвеномация и травма от морских существ». Американский семейный врач . 69 (4): 885–90. PMID 14989575 .
- ^ Симмонс, Брайан Дж.; Гриффит, Роберт Д.; Falto-Aizpurua, Leyre A.; Nouri, Keyvan (2015). "Лунная медуза ужавает " Джама Дерматология 151 (4): 454–6 Doi : 10.1001/jamadermatol.2014.4644 . PMID 25517656 .
- ^ Бакстер, эх; Марр, AGM (май 1974). «Морская оса ( Chironex fleckeri ) Антифицировать: нейтрализующая потенция против яда трех других видов медуз». Токсикона . 12 (3): 223–225. Bibcode : 1974txcn ... 12..223b . doi : 10.1016/0041-0101 (74) 90062-2 . PMID 4156430 .
- ^ «Укусы медуз: лечение и лекарства» . Клиника Майо . Майо Фонд медицинского образования и исследований. 1 сентября 2011 года. Архивировано с оригинала 20 мая 2013 года . Получено 15 апреля 2013 года .
- ^ Jump up to: а беременный «Медзой стала дикой-только текст» . Nsf.gov. Архивировано из оригинала 12 июля 2010 года . Получено 18 октября 2010 года .
- ^ «Текущий отчет об уведомлении о событиях» . Нр. 22 октября 2008 года. Архивировано с оригинала 5 июня 2011 года . Получено 14 июля 2010 года .
- ^ Рялл, Джулиан (2 ноября 2009 г.). «Японский рыбацкий траулер, утопленный гигантской медузой» . Лондон: Telegraph.co.uk. Архивировано с оригинала 4 ноября 2009 года.
Дальнейшее чтение
- Джули Бервальд (2017). Беспостол: наука о медузе и искусство выращивания основы . Риверхед книги. ISBN 978-0-7352-1126-1 .
Внешние ссылки
- Медуза и расческа желе - Смитсоновский океанский портал
- Факты медузы - информация о безопасности медузы и медуз
- « Там нет такой вещи, как медуза » из канала Mbari YouTube
- «Порочные красоты - медуза» - документальный фильм о медузе
- Они вступают во владение! nybooks.com 26 сентября 2013 г. Тим Фланнери
- Фото
Экстутные Cnidaria классы |
|---|
