Аллель
Аллель [1] , или аллеломорф , представляет собой вариант последовательности нуклеотидов в определенном месте или локусе молекулы ДНК . [2]
Аллели могут различаться в одном положении посредством однонуклеотидного полиморфизма (SNP). [3] но они также могут иметь вставки и делеции до нескольких тысяч пар оснований . [4]
Большинство наблюдаемых аллелей приводят к незначительным изменениям или вообще к их отсутствию в функции генного продукта , который они кодируют. Однако иногда разные аллели могут приводить к разным наблюдаемым фенотипическим признакам , таким как разная пигментация . Ярким примером этого является открытие Грегора Менделя о том, что белый и фиолетовый цвета цветов у растений гороха являются результатом одного гена с двумя аллелями.
Почти все многоклеточные организмы имеют два набора хромосом в какой-то момент своего биологического жизненного цикла ; то есть они диплоидны . В этом случае хромосомы могут быть парными . Каждая хромосома в паре содержит одни и те же гены в одном и том же порядке и в одном и том же месте по длине хромосомы. Для данного гена , если две хромосомы содержат один и тот же аллель, они и организм гомозиготны по этому гену. Если аллели разные, они и организм гетерозиготны по этому гену.
Популярные определения «аллеля» обычно относятся только к различным аллелям внутри генов. Например, группа крови АВО контролируется геном АВО , имеющим шесть общих аллелей (вариантов). В популяционной генетике почти каждого живого человека фенотип по гену АВО представляет собой комбинацию именно этих шести аллелей. [5] [6]
Этимология
[ редактировать ]Слово «аллель» — это краткая форма слова «аллеломорф» («другая форма», слово, придуманное британскими генетиками Уильямом Бейтсоном и Эдит Ребеккой Сондерс ) в 1900-х годах. [7] [8] который использовался на заре генетики для описания вариантов форм гена , обнаруженных как разные фенотипы . Оно происходит от греческого префикса ἀλληλο-, аллело- , означающего «взаимный», «взаимный» или «друг друга», который сам по себе связан с греческим прилагательным ἄλλος, allos (родственным латинскому alius ), означающим «другой».
Аллели, которые приводят к доминантным или рецессивным фенотипам
[ редактировать ]Во многих случаях генотипические взаимодействия между двумя аллелями в локусе можно описать как доминантные или рецессивные , в зависимости от того, на какой из двух гомозиготных фенотипов больше всего похожа гетерозигота . Если гетерозигота неотличима от одной из гомозигот, экспрессируется тот аллель, который приводит к «доминантному» фенотипу. [9] [10] а другой аллель считается «рецессивным». Степень и характер доминирования варьируются в зависимости от локуса. Этот тип взаимодействия был впервые формально описан Грегором Менделем . Однако многие признаки не поддаются этой простой классификации, и фенотипы моделируются кодоминированием и полигенным наследованием . [11]
Термин « аллель дикого типа » иногда используется для описания аллели, которая, как считается, вносит вклад в типичный фенотипический признак, наблюдаемый в «диких» популяциях организмов, таких как плодовые мухи ( Drosophila melanogaster ). Исторически считалось, что такой аллель «дикого типа» приводит к доминантному (подавляющему – всегда выраженному), общему и нормальному фенотипу, в отличие от « мутантных » аллелей, которые приводят к рецессивным, редким и часто вредным фенотипам. Раньше считалось, что большинство людей были гомозиготны по аллели «дикого типа» в большинстве генных локусов и что любой альтернативный «мутантный» аллель обнаруживался в гомозиготной форме у небольшого меньшинства «пораженных» людей, часто в виде генетических заболеваний . чаще в гетерозиготной форме у « носителей » мутантного аллеля. Сейчас понятно, что большинство или все локусы генов высоко полиморфны, имеют множество аллелей, частоты которых варьируются от популяции к популяции, и что значительная часть генетических вариаций скрыта в форме аллелей, которые не вызывают очевидных фенотипических различий. Аллели дикого типа часто обозначаются надстрочным знаком плюс ( то есть , п + для аллели p ). [12]
Множественные аллели
[ редактировать ]



Популяция или вид организмов обычно включает в себя несколько аллелей в каждом локусе у разных особей. Аллельную изменчивость в локусе можно измерить как количество присутствующих аллелей ( полиморфизм ) или долю гетерозигот в популяции. Нулевой аллель — это вариант гена, в котором отсутствует нормальная функция гена, поскольку он либо не экспрессируется, либо экспрессируемый белок неактивен.
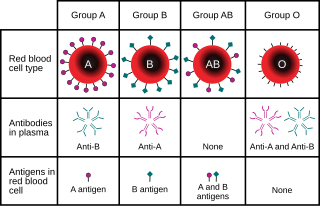
Например, в локусе гена АВО группы крови углеводных антигенов у человека: [13] классическая генетика признает три аллеля, I А , я Б , и i, определяющие совместимость переливания крови . Любая особь имеет один из шести возможных генотипов (I А я А , я А я, я Б я Б , я Б я, я А я Б и ii) которые производят один из четырех возможных фенотипов : «Тип А» (произведенный I А я А гомозиготный и я А i гетерозиготные генотипы), «Тип Б» (произведен I Б я Б гомозиготный и я Б i гетерозиготные генотипы), «Тип АВ» производства I А я Б гетерозиготный генотип, и «Тип О», продуцируемый ii гомозиготным генотипом. (Сейчас известно, что каждый из аллелей A, B и O на самом деле представляет собой класс множества аллелей с разными последовательностями ДНК, которые продуцируют белки с идентичными свойствами: в локусе ABO известно более 70 аллелей. [14] Следовательно, человек с кровью «типа А» может быть гетерозиготой АО, гомозиготой АА или гетерозиготой АА с двумя разными аллелями «А».)
Частоты генотипов
[ редактировать ]Частоту аллелей в диплоидной популяции можно использовать для прогнозирования частот соответствующих генотипов (см. принцип Харди-Вайнберга ). Для простой модели с двумя аллелями;


где p — частота одного аллеля, а q — частота альтернативного аллеля, сумма которых обязательно равна единице. Тогда п 2 – доля популяции, гомозиготная по первому аллелю, 2 pq – доля гетерозигот, q 2 — фракция, гомозиготная по альтернативному аллелю. Если первый аллель доминирует над вторым, то доля популяции, которая будет демонстрировать доминантный фенотип, равна p. 2 + 2 pq , а фракция с рецессивным фенотипом равна q 2 .
С тремя аллелями:
- и


В случае наличия нескольких аллелей в диплоидном локусе количество возможных генотипов (G) с количеством аллелей (а) определяется выражением:

Аллельное доминирование при генетических нарушениях
[ редактировать ]Ряд генетических нарушений возникает, когда человек наследует два рецессивных аллеля одного генного признака. Рецессивные генетические нарушения включают альбинизм , муковисцидоз , галактоземию , фенилкетонурию (ФКУ) и болезнь Тея-Сакса . Другие нарушения также обусловлены рецессивными аллелями, но поскольку локус гена расположен на Х-хромосоме, так что у мужчин имеется только одна копия (то есть они гемизиготны ), они чаще встречаются у мужчин, чем у женщин. Примеры включают красно-зеленую дальтонизм и синдром хрупкой Х-хромосомы .
Другие заболевания, такие как болезнь Хантингтона , возникают, когда человек наследует только один доминантный аллель.
Эпиаллели
[ редактировать ]Хотя наследственные признаки обычно изучаются с точки зрения генетических аллелей, эпигенетические метки, такие как метилирование ДНК, могут быть унаследованы в определенных областях генома у определенных видов - процесс, называемый трансгенерационным эпигенетическим наследованием . Термин эпиаллель используется, чтобы отличить эти наследственные метки от традиционных аллелей, которые определяются нуклеотидной последовательностью . [15] особый класс эпиаллелей, метастабильные эпиаллели , который характеризуется стохастическим (вероятностным) установлением эпигенетического состояния, которое может наследоваться митотически. У мышей и людей был обнаружен [16] [17]
Идиоморф
[ редактировать ]Термин «идиоморф» от греческого «morphos» (форма) и «idio» (единственное, уникальное) был введен в 1990 году вместо «аллеля» для обозначения последовательностей в одном и том же локусе у разных штаммов, которые не имеют сходства последовательностей и не имеют сходства. вероятно, не имеют общих филогенетических отношений. Его используют главным образом в генетических исследованиях микологии . [18] [19]
См. также
[ редактировать ]Ссылки и примечания
[ редактировать ]- ^ Великобритания : / ˈ æ l iː l / , / ə ˈ l iː l / ; США : / ə ˈ l iː l / ; современное образование от греческого ἄλλος állos , «другой»
- ^ Граур, Д. (2016). Молекулярная и геномная эволюция . Сандерленд, Массачусетс (США): Sinauer Associates, Inc.
- ^ Смигельски, Элизабет М.; Сироткин, Карл; Уорд, Минхонг; Шерри, Стивен Т. (1 января 2000 г.). «dbSNP: база данных однонуклеотидных полиморфизмов» . Исследования нуклеиновых кислот . 28 (1): 352–355. дои : 10.1093/нар/28.1.352 . ISSN 0305-1048 . ПМК 102496 . ПМИД 10592272 .
- ^ Элстон, Роберт; Сатагопан, Джая; Сунь, Шуйин (2012). «Генетическая терминология». Статистическая генетика человека . Методы молекулярной биологии. Том. 850. стр. 1–9. дои : 10.1007/978-1-61779-555-8_1 . ISBN 978-1-61779-554-1 . ISSN 1064-3745 . ПМЦ 4450815 . ПМИД 22307690 .
- ^ Зельцам А., Халленслебен М., Коллманн А., Блащик Р. (октябрь 2003 г.). «Природа разнообразия и диверсификации в локусе АВО» . Кровь . 102 (8): 3035–42. дои : 10.1182/кровь-2003-03-0955 . ПМИД 12829588 .
- ^ Огасавара К., Баннаи М., Сайто Н., Ябэ Р., Наката К., Такенака М., Фудзисава К., Утикава М., Исикава Ю., Джуджи Т., Токунага К. (июнь 1996 г.). «Обширный полиморфизм гена группы крови АВО: три основные линии аллелей общих фенотипов АВО». Генетика человека . 97 (6): 777–83. дои : 10.1007/BF02346189 . ПМИД 8641696 . S2CID 12076999 .
- ^ Крафт, Джуд (2013). «Гены и генетика: язык научных открытий» . Гены и генетика . Оксфордский словарь английского языка . Архивировано из оригинала 29 января 2018 года . Проверено 14 января 2016 г.
- ^ Бейтсон, В. и Сондерс, Э.Р. (1902) «Факты наследственности в свете открытия Менделя». Отчеты Комитету по эволюции Королевского общества, I. стр. 125–160.
- ^ Хартл, Дэниел Л.; Элизабет В. Джонс (2005). Основная генетика: взгляд на геномику (4-е изд.). Издательство Джонс и Бартлетт. п. 600. ИСБН 978-0-7637-3527-2 .
- ^ Монга, Иша; Куреши, Абид; Тхакур, Нишант; Гупта, Амит Кумар; Кумар, Манодж (сентябрь 2017 г.). «ASPsiRNA: ресурс ASP-siRNA, обладающий терапевтическим потенциалом в отношении генетических нарушений человека, и алгоритм прогнозирования их ингибирующей эффективности» . Г3 . 7 (9): 2931–2943. дои : 10.1534/g3.117.044024 . ПМК 5592921 . ПМИД 28696921 .
- ^ «Алель» . Genome.gov . Архивировано из оригинала 28 июня 2021 года . Проверено 3 июля 2021 г.
- ^ Б.А. Пирс (2020). Генетика. Концептуальный подход (7-е изд.). Макмиллан. п. 60. ИСБН 978-1-319-21680-1 .
- ^ Виктор А. МакКьюсик; Кассандра Л. Книффин; Пол Дж. Конверс; Ада Хамош (10 ноября 2009 г.). «АВО-гликозилтрансфераза; АВО» . Интернет-менделевское наследование у человека . Национальная медицинская библиотека. Архивировано из оригинала 24 сентября 2008 года . Проверено 24 марта 2010 г.
- ^ Йип СП (январь 2002 г.). «Вариация последовательности локуса АВО человека» . Анналы генетики человека . 66 (1): 1–27. дои : 10.1017/S0003480001008995 . ПМИД 12014997 .
- ^ Даксингер, Люсия; Уайтлоу, Эмма (31 января 2012 г.). «Понимание трансгенерационного эпигенетического наследования через гаметы у млекопитающих». Обзоры природы Генетика . 13 (3): 153–62. дои : 10.1038/nrg3188 . ПМИД 22290458 . S2CID 8654616 .
- ^ Ракян, Вардман К; Блюитт, Марни Э; Друкер, Рики; Прейс, Йост I; Уайтлоу, Эмма (июль 2002 г.). «Метастабильные эпиаллели у млекопитающих». Тенденции в генетике . 18 (7): 348–351. дои : 10.1016/S0168-9525(02)02709-9 . ПМИД 12127774 .
- ^ Уотерленд, РА; Долиной, округ Колумбия; Лин, младший; Смит, Калифорния; Ши, Х; Тахилиани, КГ (сентябрь 2006 г.). «Материнские метиловые добавки увеличивают метилирование ДНК потомства в Axin Fused». Бытие . 44 (9): 401–6. дои : 10.1002/dvg.20230 . ПМИД 16868943 . S2CID 36938621 .
- ^ Гласс, Нидерланды; Ли, Л. (1992). «Выделение мутантов Neurospora crassa a типа спаривания путем повторной индуцированной точечной мутации (RIP)» . Генетика . 132 : 125–133. дои : 10.1093/генетика/132.1.125 . ПМК 1205111 . ПМИД 1398049 .
- ^ Метценберг, Роберт Л.; Гласс, Н. Луиза (1990). «Тип спаривания и стратегии спаривания Neurospora ». Биоэссе . 12 (2): 53–59. дои : 10.1002/bies.950120202 . ПМИД 2140508 . S2CID 10818930 .
Внешние ссылки
[ редактировать ]| Базы данных органов управления : Национальные |
|---|