Устойчивость к растительным заболеваниям


Устойчивость к заболеваниям растений защищает растения от патогенов двумя способами: предварительно сформированными структурами и химическими веществами, а также от реакции иммунной системы, вызванных инфекцией. По сравнению с восприимчивым растением, устойчивость к заболеваниям является снижение роста патогенов на растении или в растении (и, следовательно, снижение заболевания), в то время как термин термин, толерантность к заболеваниям описывает растения, у которых мало повреждения заболевания, несмотря на существенные уровни патогена. Результат заболевания определяется трехсторонним взаимодействием патогена, растения и условий окружающей среды (взаимодействие, известное как треугольник заболевания ).
Активирующие обороны соединения могут двигаться к клеткам в клетку и систематически через сосудистую систему завода. Тем не менее, растения не имеют циркулирующих иммунных клеток , поэтому у большинства типов клеток широкий набор антимикробной защиты. Хотя явные качественные различия в устойчивости к заболеваниям могут наблюдаться при сравнении нескольких образцов (позволяя классификации как «устойчивую» или «восприимчивую» после инфекции одним и тем же штаммом патогена при аналогичных уровнях инокулята в аналогичных средах), градация количественных различий в устойчивости к заболеваниям более обычно наблюдается между штаммами растений или генотипами . Растения последовательно сопротивляются определенным патогенам, но поддаются другим; Устойчивость обычно специфична для определенных видов патогенов или штаммов патогенов.
Фон
[ редактировать ]Устойчивость к заболеваниям растений имеет решающее значение для надежного производства продуктов питания и обеспечивает значительное снижение использования земли, воды, топлива и других ресурсов. Растения как в естественном, так и в культивируемом населении несут в себе устойчивость к болезням, но это не всегда защищало их.
Поздний Blight Великий голод Ирландии 1840 -х годов был вызван oomycete phytophthora infestans . Первый в мире массовый культивированный банановый сорта Грос Мишель был потерян в 1920-х годах из-за панамской болезни, вызванной грибком Fusarium Oxysporum . Нынешняя пшеничная ржавчина , листовая ржавчина и эпидемии ржавчины с желтой полосой, распространяющиеся из Восточной Африки на индийский субконтинент, вызваны ржавчинами грибов Puccinia graminis и P. strififormis . Другие эпидемии включают каштановый гниль , а также рецидивирующие тяжелые заболевания растений, такие как рисовый взрыв , нематода кисты сои и цитрусовый Canker . [ 1 ] [ 2 ]
Патогены растений могут быстро распространяться на больших расстояниях, подвергшихся воздействию воды, ветра, насекомых и людей. По оценкам, в больших регионах и многих видах сельскохозяйственных культур заболевания обычно снижаются урожайность на 10% каждый год в более развитых странах или сельскохозяйственных системах, но потери доходности заболеваниям часто превышают 20% в менее развитых условиях. [ 1 ]
Тем не менее, контроль заболеваний является достаточно успешным для большинства сельскохозяйственных культур. Контроль заболеваний достигается путем использования растений, которые были выведены для хорошей устойчивости к многим заболеваниям, и подходов к выращиванию растений, таких как вращение сельскохозяйственных культур , семена без патогенов, подходящая дата посадки и плотность растений, контроль влажности поля и использование пестицидов .
Общие механизмы устойчивости к заболеваниям
[ редактировать ]Предварительно сформированные структуры и соединения
[ редактировать ]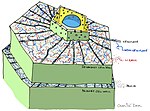
- Растительная кутикула /поверхность
- Растительные клеточные стенки
- Антимикробные химические вещества (например: полифенолы , сесквитерпеновые лактоны , сапонины )
- Антимикробные пептиды
- Ингибиторы фермента
- Детоксифицирующие ферменты, которые разрушают токсины , полученные из патогенов
- Рецепторы, которые воспринимают присутствие патогена и активируют индуцируемую защиту растений [ 3 ]
Индуцируемая постфекционная защита растений
[ редактировать ]- клеточной стенки Усиление ( целлюлоза , лигнин , суберин , каллоза , белки клеточной стенки) [ 4 ]
- Антимикробные химические вещества, включая активные формы кислорода, такие как перекись водорода или пероксинитрит , или более сложные фитоалексины, такие как генистеин или камалексин
- Антимикробные белки, такие как дефенсины , тионины или PR-1
- Антимикробные ферменты, такие как хитиназы , бета- глюканазы или пероксидазы [ 4 ]
- Гиперчувствительный ответ - быстрый ответ на гибель клеток хозяина, связанный с индукцией защиты.
Иммунная система
[ редактировать ]Иммунная система растений несет два взаимосвязанных уровня рецепторов, один наиболее часто определяющие молекулы вне клетки, а другой наиболее часто воспринимает молекулы внутри клетки. Обе системы ощущают злоумышленника и реагируют, активируя антимикробную защиту в инфицированных клетках и соседних клетках. В некоторых случаях, активирующие защиту сигналы, распространяются на остальную часть растения или даже на соседние растения. Две системы обнаруживают различные типы молекул патогенов и классы белков рецептора растений. [ 5 ] [ 6 ]
Первый уровень в первую очередь определяется рецепторами распознавания паттернов , которые активируются путем распознавания эволюционно консервативного патогена или микробного ассоциированного молекулярных паттернов (PAMPS или MAMP). Активация PRR приводит к внутриклеточной передаче сигналов, перепрограммированию транскрипции и биосинтезу сложного выходного отклика, который ограничивает колонизацию. Система известна как иммунитет, вызванный PAMP, или как иммунитет, вызванный рисунком (PTI). [ 7 ] [ 6 ] [ 8 ]
Второй уровень, в первую очередь, управляемый продуктами генов R , часто называют эффекторным иммунитетом (ETI). ETI обычно активируется наличием специфических патогенных «эффекторов», а затем вызывает сильные антимикробные ответы (см. Раздел гена R ниже).
В дополнение к PTI и ETI, защита растений может быть активирована путем определения соединений, связанных с повреждением (DAMP), таких как части стенки клеток растений, высвобождаемых во время патогенной инфекции. [ 9 ]
Ответы, активируемые рецепторами PTI и ETI, включают стробирование ионных каналов , окислительный взрыв , клеточные окислительно -восстановительные изменения или каскады протеинкиназы , которые непосредственно активируют клеточные изменения (такие как усиление клеточной стенки или антимикробные продукты) или активируют изменения в экспрессии генов , которые затем повышают другие защиты. ответы.
Иммунные системы завода показывают некоторые механистические сходства с иммунной системой насекомых и млекопитающих, но также демонстрируют много характеристик растений. [ 10 ] Два вышеупомянутых уровня являются центральными для иммунитета растений, но не полностью описывают растительные иммунные системы. Кроме того, многие конкретные примеры кажущихся PTI или ETI нарушают общие определения PTI/ETI, что свидетельствует о необходимости расширенных определений и/или парадигм. [ 11 ]
Термин количественная устойчивость (обсуждается ниже) относится к устойчивости заболевания растений, которая контролируется множественными генами и множественными молекулярными механизмами, которые каждый оказывает небольшое влияние на общую черту сопротивления. Количественная резистентность часто контрастирует с резистентностью ETI, опосредованной R-генами с одним основным эффектом.
Иммунитет, вызванный шаблоном
[ редактировать ]PAMPS патогена , консервативные молекулы, которые обитают в нескольких родах , называются многими исследователями как MAMPS. Защита, вызванная восприятием MAMP, достаточна для отталкивания большинства патогенов. Однако эффекторные белки патогена (см. Ниже) адаптированы к подавлению базальной защиты, такой как PTI. Было обнаружено много рецепторов для MAMP (и DAMPS). MAMP и DAMP часто обнаруживаются трансмембранными рецептор-киназами, которые несут LRR или внеклеточные домены LYSM . [ 5 ]
Эффектор вызвал иммунитет
[ редактировать ]Эффектор, запускаемый иммунитет (ETI), активируется присутствием эффекторов патогенов. Реакция ETI зависит от генов R и активируется специфическими штаммами патогена. Растение ETI часто вызывает апоптотический гиперчувствительный ответ .
R гены и белки R
[ редактировать ]Растения разработали гены R (гены устойчивости), чьи продукты опосредуют устойчивость к специфическим вирусу, бактериям, оомицету, грибке, нематоде или штаммам насекомых. R -продукты генов - это белки, которые позволяют распознавать специфические эффекторы патогена, либо путем прямого связывания, либо путем распознавания изменения эффектора белка -хозяина. [ 6 ] Многие R-гены кодируют белки NB-LRR (белки с нуклеотид-связывающими и богатыми лейцином доменов, также известные как белки NLR или белки стенда, среди других имен). Большинство растительных иммунных систем несут репертуар из 100–600 различных гомологов генов. Было продемонстрировано отдельные гены R, опосредуют устойчивость к специфическим вирусу, бактериям, оомицету, грибке, нематоде или штаммам насекомых. R -продукты генов контролируют широкий набор реакций устойчивости к заболеваниям, индукция которых часто достаточна для остановки дальнейшего роста/распространения патогенов.
Изученные гены R обычно придают специфичность для определенных штаммов видов патогенов (те, которые экспрессируют признанный эффектор). Как впервые отметил Гарольд Флор в своей формулировке отношений генов с геном в середине 20-го века , ген растительного r обладает специфичностью для гена авирулентности патогена (ген AVR). В настоящее время известно, что гены авирулентности кодируют эффекторы. Ген патогена AVR должен соответствовать специфичности с геном R для этого гена R для устойчивости, что указывает на взаимодействие рецептора/ лиганда для генов AVR и R. [ 10 ] Альтернативно, эффектор может модифицировать свою клеточную мишень -хозяина (или молекулярную приманку этой цели), а продукт гена R (белок NLR) активирует защиту, когда он обнаруживает модифицированную форму цели или приманки. [ 6 ] [ 12 ]
Эффекторная биология
[ редактировать ]Эффекторы являются центральными в патогенном или симбиотическом потенциале микробов и микроскопических животных, колонизирующих растения, таких как нематоды. [ 13 ] [ 14 ] [ 15 ] Эффекторы, как правило, являются белками, которые доставляются вне микроба и в клетку -хозяина. Эти эффекторы, полученные из колониста, манипулируют физиологией и развитием клеток. Таким образом, эффекторы предлагают примеры совместной эволюции (пример: грибковой белок, который функционирует за пределами гриба, но внутри растительных клеток развивался для выполнения функций, специфичных для растений). Диапазон хозяев патогена определяется, среди прочего, наличием соответствующих эффекторов, которые позволяют колонизации конкретного хозяина. [ 5 ] Эффекторы, полученные из патогенов, являются мощным инструментом для идентификации функций растений, которые играют ключевую роль в заболеваниях и устойчивости к заболеваниям. По -видимому, большинство эффекторов функционируют, чтобы манипулировать физиологией хозяина, чтобы позволить заболевать. Хорошо изученные патогены бактериальных растений обычно экспрессируют несколько десятков эффекторов, часто доставляемых в хозяин с помощью секреции типа III . [ 13 ] Грибковые, оомиковые и нематодные патогены растений, по -видимому, экспрессируют несколько сотен эффекторов. [ 14 ] [ 15 ]
Так называемые «основные» эффекторы определяются оперативно путем их широкого распределения по популяции конкретного патогена и их существенного вклада в вирулентность патогена. Геномика может быть использована для идентификации основных эффекторов, которые затем можно использовать для обнаружения новых генных аллелей R , которые можно использовать при селекции растений для устойчивости к заболеваниям.
Небольшие РНК и РНК
[ редактировать ]Пути растений SRNA считаются важными компонентами патогенового иммунитета, ассоциированного с патогеном, и иммунитетом, вызванным эффектором (ETI). [ 16 ] [ 17 ] индуцированные бактериями микроРНК (miRNAs) у арабидопсиса влияют на гормональную передачу сигналов, включая ауксин, абсцизовую кислоту (ABA), жамоновую кислоту (JA) и салициловую кислоту (SA). Было показано, что [ 18 ] [ 19 ] Достижения в исследованиях генома, проведенных в исследованиях, выявили массивную адаптацию паттернов экспрессии miRNA -хозяина после заражения грибковыми патогенами Fusarium virguliforme , [ 20 ] трава Erysiphe [ 21 ] Verticillium dahliae , [ 22 ] И Cronartium Oak , [ 23 ] и oomycete phytophthora sojae . [ 24 ] Изменения в экспрессии SRNA в ответ на грибковые патогены показывают, что молчание генов может быть вовлечено в этот защитный путь. Тем не менее, есть также доказательства того, что противогрибковая защита ответа на Colletotrichum spp. Инфекция в кукурузе не полностью регулируется специфической индукцией miRNA, но вместо этого может действовать, чтобы точно наладить баланс между генетическими и метаболическими компонентами при инфекции. [ Цитация необходима ]
Транспортировка SRNAS во время инфекции, вероятно, облегчается внеклеточными везикулами (EV) и мультивессусными телами (MVBS). [ 25 ] Композиция РНК в растительных электромобилях не была полностью оценена, но вполне вероятно, что они частично ответственны за РНК переноса. Растения могут транспортировать вирусные РНК, мРНК , микроРНК и небольшие мешающие РНК (миРНК) системно через флоэму. [ 26 ] Считается, что этот процесс происходит через Plasmodesmata и включает РНК-связывающие белки, которые помогают локализации РНК в клетках мезофилла. Несмотря на то, что они были идентифицированы в флоэме с мРНК, нет никаких определенных доказательств того, что они опосредуют долгосрочный транспорт РНК. [ 27 ] Следовательно, электромобили могут способствовать альтернативному пути нагрузки РНК в флоэму или, возможно, могут переносить РНК через апопласт. [ 28 ] Существуют также доказательства того, что EV растительных электромобилей могут позволить межвидовую перенос SRNAS с помощью РНК-интерференции, таких как молчание генов, индуцированное хозяином (HIGS). [ 29 ] [ 30 ] Транспортировка РНК между растениями и грибами, по -видимому, является двунаправленной, поскольку было показано, что SRNAs из грибного патогена Botrytis cinerea нацелены на гены обороны хозяина у арабидопсиса и помидора. [ 31 ]
Сопротивление на уровне видов
[ редактировать ]В небольшом количестве случаев гены растений эффективны против целых видов патогенов, даже если этот вид патогенный для других генотипов этих видов хозяина. Примеры включают ячмень MLO против мучнистой роды , пшеница LR34 против ржавчины листьев и пшеницы YR36 против ржавчины пшеницы . Можно существовать множество механизмов для этого типа сопротивления в зависимости от конкретной комбинации генов и растительного патогена. Другие причины эффективного иммунитета растений могут включать в себя отсутствие коадоптации (у патогена и/или растения отсутствует множество механизмов, необходимых для колонизации и роста у этих видов-хозяев), или особенно эффективный набор заранее сформированной защиты. [ Цитация необходима ]
Сигнальные механизмы
[ редактировать ]Восприятие присутствия патогена
[ редактировать ]Сигнализация обороны растений активируется рецепторами, определяющими патоген, которые описаны в вышеупомянутом разделе. [ 5 ] Активированные рецепторы часто вызывают реактивную продукцию кислорода и оксида азота , кальция , калий и протон -ионные потоки , измененные уровни салициловой кислоты и других гормонов и активация киназ MAP и другие специфические протеинкиназы . [ 10 ] Эти события, в свою очередь, обычно приводят к модификации белков, которые контролируют транскрипцию генов , и активации экспрессии генов , связанной с защитой . [ 8 ]
Факторы транскрипции и гормональный ответ
[ редактировать ]Были идентифицированы многочисленные гены и/или белки, а также другие молекулы, которые опосредуют трансдукцию сигнала обороны растений. [ 32 ] [ 33 ] Динамика цитоскелета и транспорта пузырьков помогает ориентироваться на защиту растений в направлении патогенной атаки.
Механизмы транскрипционных факторов и гормонов
[ редактировать ]Активность иммунной системы растений частично регулируется сигнальными гормонами, такими как: [ 34 ] [ 35 ]
Среди этих путей может быть существенный перекрестный разговор. [ 34 ]
Регулирование путем деградации
[ редактировать ]Как и во многих путях передачи сигнала, экспрессия генов растений во время иммунных ответов может регулироваться путем разложения. Это часто происходит, когда связывание гормонов с гормональными рецепторами стимулирует убиквитин , связанную с деградацией репрессорных белков, которые блокируют экспрессию определенных генов. Чистым результатом является активированная гормоном экспрессия генов. Примеры: [ 36 ]
- Ауксин : связывается с рецепторами, которые затем рекрутируют и разлагают репрессоры транскрипционных активаторов, которые стимулируют специфичную для ауксин экспрессии генов.
- Жасмоновая кислота: аналогично ауксину, за исключением того, что рецепторы джассоната, влияющие на сигнальные медиаторы джассоната-ответа, такие как белки JAZ.
- Гиббелловая кислота : гиббеллин вызывает конформационные изменения рецептора и связывание и деградацию белков Della .
- Этилен: ингибирующее фосфорилирование активатора этилена EIN2 блокируется этиленовым связыванием. Когда это фосфорилирование уменьшается, белок EIN2 расщепляется, а часть белка перемещается в ядро для активации экспрессии гена этилен-ответа.
Передача сигналов убиквитина и E3
[ редактировать ]Убиквитинирование играет центральную роль в передаче сигналов клеток, которая регулирует процессы, включая деградацию белка и иммунологический ответ. [ 37 ] Хотя одной из основных функций убиквитина является нацеливание белков для разрушения, он также полезен в сигнальных путях, высвобождении гормонов, апоптозе и транслокации материалов по всей клетке. Убиквитинирование является компонентом нескольких иммунных ответов. Без надлежащего функционирования убиквитина инвазия патогенов и других вредных молекул резко возрастет из -за ослабленной иммунной защиты. [ 37 ]

E3 передача сигналов
[ редактировать ]E3 Фермент убиквитиновой лигазы является основным компонентом, который обеспечивает специфичность в путях деградации белка, включая иммунные сигнальные пути. [ 36 ] Компоненты фермента E3 могут быть сгруппированы с помощью доменов, которые они содержат, и включают несколько типов. [ 38 ]
К ним относятся кольцо и единая субъединица U-Box, HECT и CRLS . [ 39 ] [ 40 ] Пути сигналов растений, включая иммунные ответы, контролируются несколькими путями обратной связи, которые часто включают отрицательную обратную связь; и они могут регулироваться ферментами деубиквитинирования, деградацией транскрипционных факторов и деградацией негативных регуляторов транскрипции. [ 36 ] [ 41 ]
Количественное сопротивление
[ редактировать ]Различия в устойчивости заболевания растений часто являются постепенными или количественными, а не качественными. Термин количественного устойчивости (QR) относится к устойчивости заболевания растений, которая контролируется множественными генами и множественными молекулярными механизмами, которые каждый оказывает небольшое или незначительное влияние на общую черту сопротивления. [ 42 ] QR важен при размножении растений, потому что результирующее сопротивление часто более долговечнее (эффективное в течение большего количества лет) и, скорее всего, будет эффективным против большинства или всех штаммов конкретного патогена. QR, как правило, эффективен против одного вида патогенов или группы близкородственных видов, а не в целом эффективен по отношению к нескольким патогенам. [ 42 ] QR часто получают путем разведения растений без знания причинно -следственных генетических локусов или молекулярных механизмов. QR, вероятно, будет зависеть от многих компонентов иммунной системы растений, обсуждаемых в этой статье, а также признаков, которые являются уникальными для определенных парами растительных патогенов (таких как чувствительность к определенным эффекторам патогена), а также общие признаки растений, такие как лист Характеристики поверхности или корневая система или архитектура навеса растения. Термин QR является синонимом незначительной резистентности к генам . [ 43 ]
Устойчивость к растениям взрослых и устойчивость к рассаду
[ редактировать ]Устойчивость растений для взрослых (APR) - это специализированный термин, относящийся к количественному сопротивлению, который не эффективен на стадии проростки, но эффективен на многих оставшихся стадиях роста растений. [ 43 ] [ 44 ] [ 42 ] Разница между сопротивлением растений и устойчивости к рассадам у взрослых особенно важна для годовых культур . [ 45 ] Устойчивость к рассадам - это сопротивление, которая начинается на стадии развития растений и продолжается на протяжении всей жизни. При использовании специалистов этот термин не относится к сопротивлению, которое активно только на стадии саженца. «Резистентность на рассадах» предназначена для синонимы основной устойчивости к генам или всей устойчивости к стадии (ASR) и используется в качестве контраста с «устойчивостью к растению для взрослых». [ 43 ] Резистентность на рассадах часто опосредуется отдельными генами R, но не все гены R кодируют устойчивость к рассадам.
Разведение растений для устойчивости к болезням
[ редактировать ]Заводчики растений подчеркивают отбор и развитие устойчивых к заболеванию растительных линий. Заболевания растений также могут быть частично контролироваться с использованием пестицидов и методами культивирования, такими как вращение сельскохозяйственных культур , обработка почвы, плотность посадки, некистные семена и очистка оборудования, но разновидности растений с присущей (генетически определенной) заболеванием устойчивости к заболеваниям, как правило, предпочтительнее. [ 2 ] Разведение для устойчивости к болезням началось, когда растения были впервые одомашнены. Усилия по размножению продолжаются, потому что популяции патогенов находятся под отбором давления и развивают повышенную вирулентность, патогены перемещаются (или перемещаются) в новые области, изменение методов выращивания или климата способствует некоторым патогенам и может снизить эффективность сопротивления, а размножение растений для других признаков может разрушить предварительное сопротивление Полем [ 46 ] Линия растений с приемлемой сопротивлением против одного патогена может не иметь сопротивления против других.
Разведение для сопротивления обычно включает в себя:
- Идентификация растений, которые могут быть менее желательными другими способами, но которые имеют полезную признаку устойчивости к заболеваниям, включая дикие растительные линии, которые часто выражают повышенную устойчивость.
- Пересечение желаемого, но воспринимаемого болезней разнообразия к растению, которое является источником сопротивления.
- Рост кандидатов на размножение в условиях, проводящих заболевание, возможно, включая прививку патогена. Внимание должно быть уделено конкретным изолятам патогена, чтобы учесть изменчивость в пределах одного вида патогенов.
- Отбор устойчивых к болезням людей, которые сохраняют другие желательные признаки, такие как урожайность, качество и включая другие черты устойчивости к заболеваниям. [ 46 ]
Сопротивление называется долговечностью , если она продолжает эффективно в течение нескольких лет широко распространенного использования по мере развития популяций патогенов. « Вертикальное сопротивление » специфична для определенных рас или штаммов видов патогенов, часто контролируется отдельными генами R и может быть менее долговечным. Горизонтальная или широкая спектра устойчивость к целым видам патогенов часто является лишь не полностью эффективной, но более долговечной и часто контролируется многими генами, которые разделяют в популяциях размножения. [ 2 ] Долговечность сопротивления важна, даже когда ожидается, что будущее улучшенные сорта будут на пути: среднее время от признания человека новой угрозы грибковых заболеваний для высвобождения устойчивой культуры для этого патогена составляет не менее двенадцати лет. [ 47 ] [ 48 ]
Такие культуры, как картофель, яблоко, банан и сахарный тростник, часто распространяются вегетативным воспроизведением , чтобы сохранить очень желательные разновидности растений, потому что для этих видов вызов серьезно нарушает предпочтительные признаки. См. Также бесполое распространение . Вегетативно распространенные культуры могут быть одними из лучших мишеней для улучшения устойчивости с помощью биотехнологического растений метода трансформации для управления генами, которые влияют на устойчивость к болезням. [ 1 ]
Научное размножение для устойчивости к заболеваниям возникло с сэром Роулендом Биффеном , который идентифицировал один рецессивный ген для устойчивости к желтой ржавчине пшеницы. Почти каждая культура была затем разведена, чтобы включить гены устойчивости к заболеваниям (R), многие из которых путем интрогрессии от совместимых диких родственников. [ 1 ]
GM или трансгенная устойчивость к заболеванию
[ редактировать ]Термин GM ( «генетически модифицированный» ) часто используется в качестве синонима трансгенного для обозначения растений, модифицированных с использованием технологий рекомбинантной ДНК. Растения с устойчивостью к трансгенным/ГМ против насекомых -вредителей были чрезвычайно успешными в качестве коммерческих продуктов, особенно в кукурузе и хлопке, и ежегодно сажают на более чем 20 миллионов гектаров в более чем 20 странах по всему миру [ 49 ] (См. Также генетически модифицированные культуры ). Устойчивость к трансгенным заболеваниям растений к микробным патогенам была впервые продемонстрирована в 1986 году. Экспрессия вирусного покрытия последовательностей генов генов обеспечивала устойчивость к вирусу через небольшие РНК . Это оказалось широко применимым механизмом для подавления репликации вируса. [ 50 ] Комбинируя гены белка покрытия из трех различных вирусов, ученые разработали гибриды сквоша с полевой, многовиральной устойчивостью. Подобные уровни устойчивости к этому разнообразию вирусов не были достигнуты с помощью обычного размножения.
Аналогичная стратегия была развернута для борьбы с вирусом Ringspot Papaya , который к 1994 году угрожал уничтожить Гавайях на индустрию папайи . Полевые испытания продемонстрировали отличную эффективность и высокое качество фруктов. К 1998 году была одобрена первая трансгенная вирусная папайя. Устойчивость к болезням была долговечностью в течение более 15 лет. Трансгенная папайя составляет ~ 85% гавайского производства. Фрукты утверждаются для продажи в США, Канаде и Японии.
Линии картофеля, экспрессирующие последовательности вирусной репликазы, которые обеспечивают устойчивость к вирусу картофеля Leafroll, были проданы под торговыми названиями NewLeaf Y и NewLeaf Plus, и были широко приняты в коммерческом производстве в 1999–2001 гг. Чтобы закрыть свой натуральный картофельный бизнес. [ 51 ] NewLeaf Y и NewLeaf Plus картофель несли две черты GM, так как они также выразили BT-опосредованную сопротивление корадовому картофельному жуку.
Никакая другая культура с инженерной устойчивостью к заболеванию к микробным патогенам не достигла рынка к 2013 году, хотя более десятка были в некотором состоянии разработки и тестирования.
| Год публикации | Обрезать | Устойчивость к болезням | Механизм | Статус развития |
|---|---|---|---|---|
| 2012 | Помидор | Бактериальное место | R ген из перца | 8 лет полевых испытаний |
| 2012 | Рис | Бактериальная гниль и бактериальная полоса | Инженер E Gene | Лаборатория |
| 2012 | Пшеница | Порошкообразная плита | Сверхэкспрессированный r ген из пшеницы | 2 года полевых испытаний во время публикации |
| 2011 | Яблоко | Apple Scab Fungus | тионин Ген из ячменя | 4 года полевых испытаний во время публикации |
| 2011 | Картофель | Картофельный вирус y | Полученная из патогена сопротивление | 1 год полевых испытаний на момент публикации |
| 2010 | Яблоко | Огненная упадок | Антибактериальный белок от моли | 12 лет полевых испытаний во время публикации |
| 2010 | Помидор | Многобактериальное сопротивление | PRR от Arabidopsis | Лабораторная масштаба |
| 2010 | Банан | Ксантомонас Увядает | Новый ген из перца | Теперь в полевых испытаниях |
| 2009 | Картофель | Поздний упадок | Г гены от диких родственников | 3 года полевых испытаний |
| 2009 | Картофель | Поздний упадок | R ген из дикого родственника | 2 года полевых испытаний во время публикации |
| 2008 | Картофель | Поздний упадок | R ген из дикого родственника | 2 года полевых испытаний во время публикации |
| 2008 | слива | Вирус Pox Pox сливы | Полученная из патогена сопротивление | Одобрения регулирующих органов, без коммерческих продаж |
| 2005 | Рис | Бактериальная полоса | R ген из кукурузы | Лаборатория |
| 2002 | Ячмень | Стебель ржавчина | Ген покоящейся лимфоцит -киназы (RLK) из устойчивого ячменного сорта | Лаборатория |
| 1997 | Папайя | Кольцо вируса | Полученная из патогена сопротивление | Утверждено и коммерчески продано с 1998 года, продается в Японию с 2012 года |
| 1995 | Давить | Три вируса мозаики | Полученная из патогена сопротивление | Утверждено и коммерчески продано с 1994 года |
| 1993 | Картофель | Картофельный вирус x | Индуцированный интерферон фермент млекопитающий | 3 года полевых испытаний во время публикации |
ПРР -передача
[ редактировать ]Исследования, направленные на инженерное сопротивление, следует за несколькими стратегиями. Одним из них является перенос полезных PRR в виды, которым им не хватает. Идентификация функциональных PRR и их переноса к видам -получателям, у которых отсутствует ортологичный рецептор, может обеспечить общий путь к дополнительным расширенным репертуарам PRR. Например, рецептор Arabidopsis PRR EF-TU (EFR) распознает фактор удлинения бактериального трансляции EF-TU . Исследования, проведенные в лаборатории Сэйнсбери, показали, что развертывание EFR в Никотиану Бентамиана или Solanum Lycopersicum (томат), который не может распознать EF-TU , обеспечил устойчивость к широкому диапазону бактериальных патогенов. Экспрессия EFR в томате была особенно эффективной против широко распространенной и разрушительной почвенной бактерии Ralstonia solanacearum . [ 52 ] И наоборот, ген томатного PRR Verticillium 1 ( VE1 ) может быть перенесен от томата в арабидопсис , где он обеспечивает устойчивость к расу 1 Verticillium изолятов. [ 1 ]
Укладка
[ редактировать ]Вторая стратегия пытается одновременно развернуть несколько генов NLR, стратегию размножения, известную как укладка. с помощью ДНК Сортировки, полученные либо молекулярной размножением , либо переносом генов, вероятно, будут демонстрировать более прочную устойчивость, потому что патогены должны будут мутировать множественные эффекторные гены. Секвенирование ДНК позволяет исследователям функционально «шахтные» гены NLR из нескольких видов/штаммов. [ 1 ]
Эффекторный ген AVRBS2 от Xanthomona Perforans является причинным агентом бактериального точечного заболевания перца и томата. Первый «эффектор-рациональный» поиск потенциально прочного гена R последовал за обнаружением того, что AVRBS2 обнаруживается у большинства вызывающих болезней видов Xanthomonas и необходим для подготовки патогенов. Ген NLR BS2 из дикого перца, Capsicum chacoense , был перемещен в помидор, где он ингибировал рост патогена. Полевые испытания продемонстрировали надежную устойчивость без бактерицидных химических веществ. Тем не менее, редкие штаммы Xanthomonas преодолели BS2 -опосредованную устойчивость в перцем путем приобретения мутаций AVRBS2 , которые избегают распознавания, но сохраняют вирулентность. Укладка генов R, которые каждый распознает различный эффектор, может задержать или предотвратить адаптацию. [ 1 ]
Более 50 локусов в штаммах пшеницы придают устойчивости к болезням с стеблем пшеницы, патогены листьев и желтой полосы ржавчины. Ген NLR STEM Rust 35 ( SR35 ), клонированный из диплоидного относительного культивируемой пшеницы, Triticum Monococcum , обеспечивает устойчивость к изоляту ржавчины пшеницы UG99 . Аналогичным образом, SR33 , от относительного эгилопса Tauschii , кодирует ортолог пшеницы в гены . -резистентности генов пшеницы Оба гена необычны в пшенице и ее родственниках. В сочетании с геном SR2 , который действует аддитивно с SR33, они могут обеспечить прочную устойчивость к болезням UG99 и его производные. [ 1 ]
Гены исполнителя
[ редактировать ]Другой класс генов устойчивости к болезням растений открывает «дверь ловушки», которая быстро убивает вторгнутые клетки, останавливая пролиферацию патогена. xanthomonas и ralstonia Ксантомонас и активатор транскрипции -подобные (TAL) эффекторам-это ДНК-связывающие белки, которые активируют экспрессию генов хозяина для усиления вирулентности патогена. Как линии риса, так и перца независимо эволюционировали сайты связывания тала-эффекта, которые вместо этого действуют как палач, который вызывает гиперчувствительную гибель клеток хозяина при повышении. XA27 из риса и BS3 и BS4C из перца, являются такими генами «исполнителя» (или «казней»), которые кодируют не гомологичные растительные белки неизвестной функции. Гены исполнителя экспрессируются только в присутствии конкретного эффектора TAL. [ 1 ]
Инженерные гены исполнителей были продемонстрированы путем успешного переработки промотора перца BS3 , чтобы содержать два дополнительных сайта связывания для эффекторов TAL от разрозненных штаммов патогенов. Впоследствии, инженерный ген исполнителя был развернут в рисе, добавив пять сайтов связывания эффектора TAL к промотору XA27 . Синтетическая конструкция XA27 обеспечивает устойчивость к ксантомонаскому бактериальному гнили и видам бактериальных полос листьев. [ 1 ]
Аллели восприимчивости проводятся
[ редактировать ]Большинство растительных патогенов перепрограммируют паттерны экспрессии генов хозяина, чтобы непосредственно принести пользу патогену. Репрограммируемые гены, необходимые для выживания патогена и пролиферации, можно рассматривать как «гены восприимчивости к болезням». Рецессивные гены устойчивости являются кандидатами на заболевание. Например, мутация отключила ген арабидопсиса, кодирующий пектат лиязу (вовлеченную в деградацию клеточной стенки), обеспечивая устойчивость к патогеном порошкообразного патогена Mildew Golovinomyces Cichoracearum . Точно так же ген ячменя MLO и самопроизвольно мутированные гороховые и томатные ортологи MLO также обеспечивают резистентность к мучнике порошкообразной. [ 1 ]
LR34 - это ген, который обеспечивает частичную устойчивость к листьям и желтой ржавчине и порошкообразной пшеничной ростовщике в пшенице. LR34 кодирует транспортер аденозин трифосфат (АТФ) -связывающую кассету (ABC). Доминирующий аллель, который обеспечивает устойчивость к заболеваниям, был недавно обнаружен в культивируемой пшенице (не в диких штаммах) и, как и MLO , обеспечивает устойчивость к широкому спектру ячменя. [ 1 ]
Естественные аллели трансляции трансляции трансляции хозяина eIF4E и eIF4G также являются рецессивными генами вирусной устойчивости. Некоторые были развернуты для контроля потивирусов в ячменя, рисе, помидоров, перца, гороха, салата и дыни. Открытие вызвало успешный экран мутанта для химически индуцированных аллелей eIF4E в томате. [ 1 ]
Природная промоторная вариация может привести к эволюции рецессивных аллелей устойчивости к заболеваниям. Например, ген резистентного устойчивости XA13 в рисе является аллелем OS-8N3 . OS-8N3 транскрипционно активируется Xanthomonas oryzae Pv. штаммы Oryzae , которые выражают эффектор Pthxo1 . Ген XA13 имеет мутированный эффектор-связывающий элемент в своем промоторе, который устраняет связывание PthxO1 и делает эти линии устойчивыми к штаммам, которые полагаются на PthxO1 . Этот вывод также продемонстрировал, что OS-8N3 требуется для восприимчивости. [ 1 ]
XA13/OS-8N3 необходим для развития пыльцы, показывая, что такие мутантные аллели могут быть проблематичными, если фенотип восприятия заболевания изменяет функцию в других процессах. Тем не менее, мутации в OS11N3 элементе эффектора -связанного эффектора (OSSWEET14) были сделаны путем объединения эффекторов TAL к нуклеазам ( TALEN ). Растения риса с энтузиазмом генома с измененными сайтами связывания OS11N3 оставались устойчивыми к Xanthomonas oryzae pv. Oryzae , но все еще обеспечивает нормальную функцию развития. [ 1 ]
Ген молчания
[ редактировать ]Резистентность на основе РНК является мощным инструментом для технических культур. Преимущество РНКи как новой генной терапии против грибковой, вирусной и бактериальной инфекции у растений заключается в том, что она регулирует экспрессию генов посредством деградации РНК мессенджера , репрессии трансляции и ремоделирования хроматина с помощью мелких некодирующих РНК. Механистически процессы молчания руководствуются продуктами обработки двухцепочечного триггера РНК ( DSRNA ), которые известны как небольшие мешающие РНК и микроРНК . [ 53 ]
Хост диапазон
[ редактировать ]Среди тысяч видов растительных патогенных микроорганизмов только небольшое меньшинство способны заразить широкий спектр видов растений. Большинство патогенов вместо этого демонстрируют высокую степень специфичности хозяина. Часто говорят, что виды растений, не относящиеся к хозяйству, выражают устойчивость к не-боста . Термин резистентность хозяина используется, когда виды патогенов могут быть патогенными для видов хозяина, но некоторые штаммы этого вида растений противостоят определенным штаммам видов патогенов. Причины сопротивления хозяина и устойчивости к не-боста могут перекрываться. Диапазон хозяев патогена определяется, среди прочего, наличием соответствующих эффекторов, которые позволяют колонизации конкретного хозяина. [ 5 ] Диапазон хозяев патогена может измениться довольно внезапно, если, например, способность патогена синтезировать специфический для хозяина токсин или эффектор получен путем переноса/переноса генов горизонтали . [ 54 ] [ 55 ]
Эпидемии и биология населения
[ редактировать ]Нативные популяции часто характеризуются существенным разнообразием генотипа и диспергированными популяциями (рост в смеси со многими другими видами растений). Они также подвергались коэволюции растений-патогена . Следовательно, до тех пор, пока новые патогенные микроорганизмы не вводятся/не развиваются, такие популяции обычно демонстрируют лишь низкую частоту эпидемий тяжелых заболеваний . [ 56 ]
Монотропные сельскохозяйственные системы обеспечивают идеальную среду для эволюции патогенов, поскольку они предлагают высокую плотность образцов целевых с аналогичными/идентичными генотипами. [ 56 ] Рост мобильности, вытекающий из современных транспортных систем, обеспечивает патогены с доступом к более потенциальным целям. [ 56 ] Изменение климата может изменить жизнеспособный географический диапазон видов патогенов и привести к тому, что некоторые заболевания станут проблемой в областях, где заболевание ранее было менее важным. [ 56 ]
Эти факторы делают современное сельское хозяйство более подверженным эпидемиям болезней. Общие растворы включают постоянное размножение для устойчивости к заболеваниям, использование пестицидов, использование пограничных проверок и ограничения на импорт растений, поддержание значительного генетического разнообразия в бассейне генов сельскохозяйственных культур (см. Разнообразие сельскохозяйственных культур ) и постоянное наблюдение для ускорения инициации соответствующих ответов. Некоторые виды патогенов обладают гораздо большей способностью преодолеть устойчивость к болезням растений, чем другие, часто из -за их способности быстро развиваться и широко рассеиваться. [ 56 ]
Тематическое исследование американского каштана
[ редактировать ]Каштановый гниль был впервые замечен на американских каштановых деревьях, которые росли в том, что теперь известно как зоопарк Бронкса в 1904 году. В течение многих лет после этого инцидента утверждалось относительно того, какова была личность патогена, а также подходящим подход к его контролю. Самые ранние попытки решить проблему на каштане включали химические растворы или физические. Они пытались использовать фунгициды, отрезали конечности от деревьев, чтобы остановить инфекцию, и полностью удалить зараженные деревья из жилья, чтобы не позволить им заражать других. Все эти стратегии оказались безуспешными. Были представлены даже карантинные меры, которые помогли принятию Закона о карантине растений. Каштановый гниль по -прежнему оказался огромной проблемой, поскольку он быстро перемещался через густонаселенные леса каштановых деревьев. В 1914 году эта идея считала устойчивостью к разрушению деревьев с помощью различных средств и механизмов размножения. [ 57 ]
Смотрите также
[ редактировать ]- Ген-гена отношения
- Индуцированное системное сопротивление
- Защита растений от травоядных
- Патология растений
- Использование растений эндофитных грибов в обороне
- Системное приобретенное сопротивление
Ссылки
[ редактировать ]- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л м не а п Дангл, JL; Хорват, DM; Staskawicz, BJ (2013). «Поворота иммунной системы завода от рассечения до развертывания» . Наука . 341 (6147): 746–751. Bibcode : 2013sci ... 341..746d . doi : 10.1126/science.1236011 . PMC 3869199 . PMID 23950531 .
- ^ Jump up to: а беременный в Агриос, Джордж Н. (2005). Патология растений, пятое издание . Академическая пресса. ISBN 978-0120445653 .
- ^ Lutz, Diana (2012). Ключевая часть системы быстрого реагирования растений выявила архив 2023-04-18 на машине Wayback . Вашингтонский университет в Сент -Луисе.
- ^ Jump up to: а беременный Дадакова, К.; Хавелькова, М.; Куркова, Б.; Тлолкова, я.; Каспаровский, Т.; Zdrahal, Z.; Lochman, J. (2015-04-24). «Протеом и транскриптный анализ клеточных культур Vitis Vinifera, подвергшихся инфекции Botrytis cinerea». Журнал протеомики . 119 : 143–153. doi : 10.1016/j.jprot.2015.02.001 . PMID 25688916 .
- ^ Jump up to: а беременный в дюймовый и Доддс, Пн; Rathjen, JP (2010). «Иммунитет растений: к интегрированному обзору взаимодействия растений с патогеном». Nature Reviews Genetics . 11 (8): 539–548. doi : 10.1038/nrg2812 . HDL : 1885/29324 . PMID 20585331 . S2CID 8989912 .
- ^ Jump up to: а беременный в дюймовый Джонс, JD; Dangl, JL (2006). «Иммунная система растений» . Природа . 444 (7117): 323–329. Bibcode : 2006natur.444..323J . doi : 10.1038/nature05286 . PMID 17108957 .
- ^ Couto, D.; Zipfel, C. (2016). «Регуляция передачи сигналов рецептора распознавания паттернов у растений» . Природа обзоры иммунологии . 16 (9): 537–552. doi : 10.1038/nri.2016.77 . PMID 27477127 . S2CID 21610374 .
- ^ Jump up to: а беременный Ли, Бо; Мэн, Сянзонг; Шан, Либо; Он, пинг (май 2016 г.). «Транскрипционная регуляция инициируемого рисунком иммунитета у растений» . Ячейка и микроб . 19 (5): 641–650. doi : 10.1016/j.chom.2016.04.011 . PMC 5049704 . PMID 27173932 .
- ^ Молина, Антонио; Мидес, Ева; Бекте, Лора; Родригес, Тинуаро; Мелида, Хьюго; Денанс, Николас; Санчес-Валлет, Андреа; Rivière, Marie-Pierre; Лопес, Джемма; Фрейер, Амандин; Барлет, Ксавье; Патхил, Сивакумар; Хан, Майкл; Гоффнер, Дебора (2021-02-02). «Состав клеточной стенки Arabidopsis определяет специфичность и приспособленность устойчивости к заболеваниям » Труды Национальной академии наук 118 (5): E2010243118. Bibcode : 2021pnas..11810243M Doi : 10.1073/ pnas.201024311 ISSN 0027-8 PMC 7865177 PMID 33509925
- ^ Jump up to: а беременный в Нюрнбергер, Торстен; Бруннер, Frédéric; Кеммерлинг, Биргит; Пиатер, Лизель (апрель 2004 г.). «Врожденный иммунитет у растений и животных: поразительные сходства и очевидные различия». Иммунологические обзоры . 198 (1): 249–266. doi : 10.1111/j.0105-2896.2004.0119.x . PMID 15199967 . S2CID 32547933 .
- ^ Thomma, B.; Nurnberger, T.; Joosten, M. (2011). «Обоблок и эффекторов: размытая дихотомия pti-eti» . Растительная ячейка . 23 (4): 4–15. doi : 10.1105/tpc.110.082602 . PMC 3051239 . PMID 21278123 .
- ^ Даксбери, Зейн; Ма, Ян; Furzer, Oliver J.; Да, Sell Un; Cevik, Volkan; Джонс, Джонатан Д.Г.; Саррис, Панагиотис Ф. (2016-06-24). «Восприятие патогена NLRS у растений и животных: параллельные миры». Биологии . 38 (8). Wiley : 769–781. doi : 10.1002/bies.201600046 . ISSN 0265-9247 . PMID 27339076 . S2CID 3810233 .
- ^ Jump up to: а беременный Линдеберг, Магдален; Cunnac, Sébastien; Колмер, Алан (апрель 2012 г.). «Pseudomonas syringae тип III репертуи эффекторных действий: последние слова в бесконечных аргументах». Тенденции в микробиологии . 20 (4): 199–208. doi : 10.1016/j.tim.2012.01.003 . PMID 22341410 .
- ^ Jump up to: а беременный Рафики, Марьям; Эллис, Джеффри Дж; Людовичи, Виктория А.; Хардхем, Адриенн Р.; Доддс, Питер Н. (2012). «Проблемы и прогресс в понимании роли эффекторов во взаимодействиях растений и фунгевых». Современное мнение о биологии растений . 15 (4): 477–482. Bibcode : 2012copb ... 15..477r . doi : 10.1016/j.pbi.2012.05.003 . PMID 22658704 .
- ^ Jump up to: а беременный Хевези, Тарек; Баум, Томас Дж. (Январь 2013 г.). «Манипулирование растительными клетками с помощью эффекторов нематод кисты и корневого узла» . Молекулярные взаимодействия растений-микробов . 26 (1): 9–16. doi : 10.1094/mpmi-05-12-0106-fi . PMID 22809272 .
- ^ Джин, Хэлинг (2008-07-11). «Эндогенные мелкие РНК и антибактериальный иммунитет у растений» . Письма Febs . 582 (18): 2679–2684. BIBCODE : 2008 FEBSL.582.2679J . doi : 10.1016/j.febslet.2008.06.053 . ISSN 0014-5793 . PMC 5912937 . PMID 18619960 .
- ^ Падманабхан, Челлапп; Чжан, Сяоминг; Джин, Hading (август 2009 г.). «Малые РНК -хозяины являются большими вкладчиками в врожденном иммунитете». Современное мнение о биологии растений . 12 (4): 465–472. Bibcode : 2009copb ... 12..465p . doi : 10.1016/j.pbi.2009.06.005 . ISSN 1369-5266 . PMID 19608454 .
- ^ Чжан, Вейксионг; Гао, Шан; Чжоу, Сян; Челлаппан, Падманабхан; Чен, Чжэн; Чжоу, Сюэфенг; Чжан, Сяоминг; Отварит, NYSSA; Coutino, Gabriela (2010-12-12). «Бактерий-чувствительные микроРНК регулируют врожденный иммунитет растений путем модуляции гормонов растений» . Растительная молекулярная биология . 75 (1–2): 93–105. doi : 10.1007/s11103-010-9710-8 . ISSN 0167-4412 . PMC 3005105 . PMID 21153682 .
- ^ Боккара, Мартин; Саразин, Алексис; ТИЕБЕАЛД, Одон; Джей, Флоренция; Воиннет, Оливье; Наварро, Лайонел; Колот, Винсент (2014-01-16). «Путь молчания Arabidopsis miR472-Rdr6 модулирует иммунитет, вызванный PAMP и эффектор, посредством посттранскрипционного контроля генов устойчивости к заболеваниям» . PLO -патогены . 10 (1): E1003883. doi : 10.1371/journal.ppat.1003883 . ISSN 1553-7374 . PMC 3894208 . PMID 24453975 . (Ошибка: Doi : 10.1371/journal.ppat.1004814 , PMID 25859662 , реверкция Watch )
- ^ Радван, Осман; Лю, Ю; КЛОГ, Стивен Дж. (Август 2011 г.). «Транскрипционный анализ ответа корня сои на фусарий Virguliforme, причинный агент синдрома внезапной смерти» . Молекулярные взаимодействия растений-микробов . 24 (8): 958–972. doi : 10.1094/mpmi-11-10-0271 . ISSN 0894-0282 . PMID 21751852 .
- ^ Синь, Минминг; Ван, Ю; Яо, Йингин; Се, Чаоджи; Пэн, Хуру; Ни, Zhongfu; Солнце, Qixin (2010). «Разнообразие микроРНК реагирует на инфекцию мучнистой росы и тепловое напряжение в пшенице ( Triticum aestivum L.)» . BMC Биология растений . 10 (1): 123. doi : 10.1186/1471-2229-10-123 . ISSN 1471-2229 . PMC 3095282 . PMID 20573268 .
- ^ Инь, Зуджун; Ли, Ян; Хан, Xiulan; Shen, Fafu (2012-04-25). «Обще геном профилирование miRNAs и других мелких некодирующих РНК в валовых корнях хлопка, анокулированных Verticillium dahliae » . Plos один . 7 (4): E35765. Bibcode : 2012ploso ... 735765y . doi : 10.1371/journal.pone.0035765 . ISSN 1932-6203 . PMC 3338460 . PMID 22558219 .
- ^ Лу, Шанфа; Солнце, ин-хсуан; Амерсон, Генри; Чианг, Винсент Л. (2007-08-07). «МикроРНК в сосне Лонколли ( Pinus Taeda L.) и их связь с веретеном развитием ржавчиной галльи» . Заводский журнал . 51 (6): 1077–1098. doi : 10.1111/j.1365-313x.2007.03208.x . ISSN 0960-7412 . PMID 17635765 .
- ^ Го, на; Ты, Вэнь-Ву; Ву, Ся-Лин; Шен, Дэн-Ю; Ван, Юань-Чао; Син, Хан; Dou, Dao-Long (ноябрь 2011 г.). «Профилирование микрочипов выявляет микроРНК, включающие устойчивость сои к Phytophthora sojae ». Геном . 54 (11): 954–958. doi : 10.1139/g11-050 . ISSN 0831-2796 . PMID 21995769 .
- ^ Микали, Кристина О.; Нейман, Улла; Груневальд, Дорит; Панструга, Ральф; О'Коннелл, Ричард (2010-10-28). «Биогенез специализированного границы раздела растений во время интернализации клеток-хозяина Golovinomyces orontii Haustoria» . Клеточная микробиология . 13 (2): 210–226. doi : 10.1111/j.1462-5822.2010.01530.x . ISSN 1462-5814 . PMID 20880355 . S2CID 39002035 .
- ^ Кер, Дж.; Buhtz, A. (2007-12-18). «Долголетний транспорт и перемещение РНК через флоэму» . Журнал экспериментальной ботаники . 59 (1): 85–92. doi : 10.1093/jxb/erm176 . ISSN 0022-0957 . PMID 17905731 .
- ^ Gómez, G.; Torres, H.; Паллас, В. (2004-11-29). «Идентификация трансатируемых РНК-связывающих белков флоэмы из дыни, потенциальные компоненты системы транспортировки РНК на длинные дистанции» . Заводский журнал . 41 (1): 107–116. doi : 10.1111/j.1365-313x.2004.02278.x . ISSN 0960-7412 . PMID 15610353 .
- ^ Раттер, Брайан Д; Иннес, Роджер В. (август 2018 г.). «Внеклеточные пузырьки как ключевые медиаторы взаимодействия растений -микробов» . Современное мнение о биологии растений . 44 : 16–22. Bibcode : 2018copb ... 44 ... 16r . doi : 10.1016/j.pbi.2018.01.008 . ISSN 1369-5266 . PMID 29452903 . S2CID 3377305 .
- ^ Гаг, Сиддхеш Б. (декабрь 2017 г.). «Индуцированное хозяева генов, появляющаяся наука, чтобы разработать сопротивление культурам против вредных патогенов растений». Физиологическая и молекулярная патология растений . 100 : 242–254. Bibcode : 2017pmpp..100..242G . doi : 10.1016/j.pmpp.2017.10.003 . ISSN 0885-5765 .
- ^ Новара, Даниэла; Гей, Александра; Лаком, Кристоф; Шоу, Джейн; Рассылка, Кристофер; Douchkov, Dimitar; Хенсель, Гётц; Kumlehn, Jochen; Швейзер, Патрик (сентябрь 2010 г.). «HIGS: индуцированное хозяином, молчание генов в облигационном биотрофическом грибном патоген-блудке Graminis» . Растительная ячейка . 22 (9): 3130–3141. doi : 10.1105/tpc.110.077040 . ISSN 1040-4651 . PMC 2965548 . PMID 20884801 .
- ^ Вейберг, Арне; Ван, Мин; Лин, Фенг-Мао; Чжао, Хонгвей; Чжан, Чжихон; Калошиан, Ишгухи; Huang, Hsien-da; Джин, Хэлинг (2013-10-03). «Грибковые малые РНК подавляют иммунитет растений путем захвата вмешательства РНК -интерференции хозяина» . Наука . 342 (6154): 118–123. Bibcode : 2013sci ... 342..118W . doi : 10.1126/science.1239705 . ISSN 0036-8075 . PMC 4096153 . PMID 24092744 .
- ^ Хаммонд-Косак, Ким Е; Паркер, Джейн Е (апрель 2003 г.). «Расшифрирование растительного общения: свежие перспективы для размножения молекулярной резистентности». Текущее мнение о биотехнологии . 14 (2): 177–193. doi : 10.1016/s0958-1669 (03) 00035-1 . PMID 12732319 .
- ^ Дадакова, Катерина; Клемппова, Джитка; Джендрисакова, Тереза; Лохман, Ян; Каспаровский, Томас (2013-12-01). «Выяснение сигнальных молекул, участвующих в восприятии эргостерола в табаке». Физиология растений и биохимия . 73 : 121–127. Bibcode : 2013plpb ... 73..121d . doi : 10.1016/j.plaphy.2013.09.009 . PMID 24095918 .
- ^ Jump up to: а беременный Мур, Джон У.; Loake, Gary J.; Споэл, Стивен Х. (август 2011 г.). «Динамика транскрипции при иммунитете растений» . Растительная ячейка . 23 (8): 2809–2820. doi : 10.1105/tpc.111.087346 . PMC 3180793 . PMID 21841124 .
- ^ Бюргер, Марко; Шори, Джоанн (2019-08-14). «Подчеркивается о гормонах: как растения организуют иммунитет» . Ячейка и микроб . 26 (2): 163–172. doi : 10.1016/j.chom.2019.07.006 . ISSN 1934-6069 . PMC 7228804 . PMID 31415749 .
- ^ Jump up to: а беременный в Саданандом, Ари; Бейли, Марк; Эван, Ричард; Ли, Джек; Нелис, Стюарт (октябрь 2012 г.). «Система убиквитин -протеасом: центральный модификатор передачи сигналов растений» . Новый фитолог . 196 (1): 13–28. doi : 10.1111/j.1469-8137.2012.04266.x . PMID 22897362 .
- ^ Jump up to: а беременный Trujillo, M.; Ширасу К. (август 2010 г.). «Убиквитинирование в иммунитете растений». Современное мнение о биологии растений . 13 (4): 402–408. Bibcode : 2010copb ... 13..402t . doi : 10.1016/j.pbi.2010.04.002 . PMID 20471305 .
- ^ Крейг, А.; Ewan, R.; Mesmar, J.; Гудис, В.; Это распространено, А. (1 март 2009 г.). " Интоне Журнал экспериментальной ботаники . 60 (4): 1123–1 doi : 1093/ jxb/ erp059 PMID 19276192 .
- ^ Moon, J. (1 декабря 2004 г.). «Путь убиквитин-протеемного пути и развитие растений» . Растительная ячейка онлайн . 16 (12): 3181–3195. doi : 10.1105/tpc.104.161220 . PMC 535867 . PMID 15579807 .
- ^ Трухильо, Марко; Ширасу, Кен (1 августа 2010 г.). «Убиквитинирование в иммунитете растений». Современное мнение о биологии растений . 13 (4): 402–408. Bibcode : 2010copb ... 13..402t . doi : 10.1016/j.pbi.2010.04.002 . PMID 20471305 .
- ^ Ширсекар, Гаутам; Дай, Liangying; Ху, Яджун; Ван, Сююджун; Зенг, Лиронг; Ван, Го-Лиан (февраль 2010 г.). «Роль убиквитинирования в врожденном иммунитете растений и вирулентности патогена». Журнал биологии растений . 53 (1): 10–18. Bibcode : 2010jpbio..53 ... 10s . doi : 10.1007/s12374-009-9087-x . S2CID 36185464 .
- ^ Jump up to: а беременный в Каугер, Кристина; Браун, Джеймс К.М. (2019-08-25). «Долговечность количественного сопротивления в культурах: больше, чем мы знаем?». Ежегодный обзор фитопатологии . 57 (1). Ежегодные обзоры : 253–277. doi : 10.1146/annurev-phyto-082718-100016 . ISSN 0066-4286 . PMID 31206351 . S2CID 190533925 .
- ^ Jump up to: а беременный в Парк, Роберт Ф.; Голегаонкар, Прашант Г.; Деревнина, Лида; Сандху, Каранджит с.; Караоглу, Гайдар; Elmansour, Huda M.; Dracatos, Peter M.; Singh, Davinder (2015-08-04). «Листовая ржавчина культивируемого ячменя: патология и контроль». Ежегодный обзор фитопатологии . 53 (1). Ежегодные обзоры : 565–589. doi : 10.1146/annurev-phyto-080614-120324 . ISSN 0066-4286 . PMID 26047566 .
- ^ Bhavani, Sridhar (2021-06-18). «Сопротивление растений для взрослых (APR): стратегия преодолеть постоянные патогены» . Cimmyt (Международный центр улучшения кукурузы и пшеницы) . Получено 2021-07-06 .
- ^ Сингх, Рави П.; Сингх, Паван К.; Руткоски, Джессика; Ходсон, Дэвид П.; Он, Синьяо; Jørgensen, Lise N.; Hovmøller, Mogens S.; Уэрта-Эспино, Хулио (2016-08-04). «Влияние на заболевание на потенциал урожайности пшеницы и перспективы генетического контроля». Ежегодный обзор фитопатологии . 54 (1). Ежегодные обзоры : 303–322. doi : 10.1146/annurev-phyto-080615-095835 . ISSN 0066-4286 . PMID 27296137 . S2CID 4603818 .
- ^ Jump up to: а беременный Stuthman, DD; Леонард, KJ; Миллер-Гарвин, Дж. (2007). «Разведение урожая для долговечной устойчивости к болезням». Достижения в области агрономии . 95 : 319–367. doi : 10.1016/s0065-2113 (07) 95004-x . ISBN 9780123741653 .
- ^ Shimelis, H.; Laing, M. «Сроки в обычном улучшении урожая: процедуры предварительного воспитания и размножения». Австралийский журнал науки о урожаях : 1542–1549. EISSN 1835-2707 . ISSN 1835-2693 . S2CID 55486617 .
- ^ Mahlein, A.-K.; Куска, MT; Behmann, J.; Polder, G.; Уолтер А. (2018-08-25). «Гиперспектральные датчики и технологии визуализации в фитопатологии: состояние искусства». Ежегодный обзор фитопатологии . 56 (1). Ежегодные обзоры : 535–558. doi : 10.1146/annurev-phyto-080417-050100 . ISSN 0066-4286 . PMID 30149790 . S2CID 52096158 .
- ^ Табашник, Брюс Э.; Брево, Тьерри; Карриер, Ив (2013). «Сопротивление насекомых к культурам BT: уроки первых миллиардов акров». Nature Biotechnology . 31 (6): 510–521. doi : 10.1038/nbt.2597 . PMID 23752438 . S2CID 205278530 .
- ^ Кавана, та; Spillane, C. (1995-02-01). «Стратегии устойчивости к инженерным вирусам у трансгенных растений». Euphytica . 85 (1–3): 149–158. doi : 10.1007/bf00023943 . ISSN 0014-2336 . S2CID 20940279 .
- ^ Kaniewski, Wojciech K.; Томас, Питер Э. (2004). «Картофельная история». Agbioforum . 7 (1 и 2): 41–46.
- ^ Lacombe, Séverine; Ругон-Кардосо, Алехандра; Шервуд, Эмма; Питерс, Немо; Дальбек, Дуглас; Ван Эссе, Х. Питер; Курильщик, Мэтью; Раллапалли, Ганасьям; Thomma, Bart PHJ; Стаскавич, Брайан; Джонс, Джонатан Д.Г.; Зипфель, Кирилл (17 апреля 2010 г.). «Межсемейство переноса рецептора распознавания растителей рецептор обеспечивает устойчивость к широкому спектру бактерий» . Nature Biotechnology . 28 (4): 365–369. doi : 10.1038/nbt.1613 . PMID 20231819 . S2CID 7260214 - через www.nature.com.
- ^ Картикеян, А.; Deivamani, M.; Shobhana, VG; Sudha, M.; Anandhan, T. (2013). «Вмешательство РНК: эволюции и применение в лечении заболеваний растений». Архив фитопатологии и защиты растений . 46 (12): 1430–1441. BIBCODE : 2013ARPPP..46.1430K . doi : 10.1080/03235408.2013.769315 . S2CID 85060938 .
- ^ Bettgenhaeuser, Jan; Гилберт, Брайан; Айлифф, Майкл; Моск, Мэтью Дж. (11 декабря 2014 г.). «Неухостое сопротивление патогенам ржавчины - продолжение континуа» . Границы в науке о растениях . 5 : 664. DOI : 10.3389/fpls.2014.00664 . PMC 4263244 . PMID 25566270 .
- ^ Restrepo, Сильвия; Табима, Хавьер Ф.; Mideros, Maria F.; Грюнвальд, Никлаус Дж.; Матут, Даниэль Р. (4 августа 2014 г.). «Видообразование в грибковых патогенах растений» . Ежегодный обзор фитопатологии . 52 (1). Ежегодные обзоры: 289–316. doi : 10.1146/annurev-phyto-102313-050056 . ISSN 0066-4286 . PMID 24906125 .
- ^ Jump up to: а беременный в дюймовый и Макдональд, Б.А.; Линде, С. (2002). «Генетика популяции патогенов, эволюционный потенциал и долговечная устойчивость». Ежегодный обзор фитопатологии . 40 : 349–379. doi : 10.1146/annurev.phyto.40.120501.101443 . PMID 12147764 . S2CID 23726106 .
- ^ Барнс, Джессика С; Дельборн, Джейсон А (сентябрь 2022 г.). «Политика генетического технонаука для сохранения: случай устойчивого к гниле американского каштана» . Окружающая среда и планирование E: природа и пространство . 5 (3): 1518–1540. Bibcode : 2022enple ... 5.1518b . doi : 10.1177/25148486211024910 .
Дальнейшее чтение
[ редактировать ]- Лукас, JA, «Защита растений». Глава 9 в патологии растений и патогенах растений, 3 -е изд. 1998 Blackwell Science. ISBN 0-632-03046-1
- Хаммонд-Косак, К. и Джонс, JDG «Ответы на патогены растений». В кн.: Бьюкенен, Грурис и Джонс, ред. Биохимия и молекулярная биология растений, второе издание. 2015. Wiley-Blackwell, Хобокен, Нью-Джерси. ISBN 9780470714218
- Доддс, П.; Rathjen, J. (2010). «Иммунитет растений: к интегрированному обзору взаимодействия растений с патогеном». Nature Reviews Genetics . 11 (8): 539–548. doi : 10.1038/nrg2812 . HDL : 1885/29324 . PMID 20585331 . S2CID 8989912 .
- Мишельмор, Ричард В.; Кристопулу, Марилена; Колдуэлл, Кэтрин С. (2013-08-04). «Воздействие генетики, функции и эволюции устойчивости на долговечное будущее». Ежегодный обзор фитопатологии . 51 (1). Ежегодные обзоры : 291–319. doi : 10.1146/annurev-phyto-082712-102334 . ISSN 0066-4286 . PMID 23682913 . S2CID 22234708 .
- Шуман, Г. Заболевания растений: их биология и социальное воздействие. 1991 APS Press, St. Paul, Mn. ISBN 0890541167