Гаплогруппа I-M253
| Гаплогруппа I1 (M253) | |
|---|---|
| Возможное время происхождения | 3,170–4,600 [ 1 ] –5,070 п.н. (сегодняшняя диверсификация) [ 2 ] [ 3 ] (Ранее 11 000 п.н. [ 4 ] до 33 000 п.н. [ 5 ] ) 27 500 (диверсификация с I2-FGC77992) [ 1 ] |
| Возможное место происхождения | Северная Европа [ 6 ] |
| Предок | I* (M170) |
| Потомки | I1A (DF29/S438); I1b (S249/Z131); I1C (Y18119/Z17925) |
| Определение мутаций | M253, M307.2/P203.2, M450/S109, P30, P40, L64, L75, L80, L81, L118, L121/S62, L123, L124/S64, L125/S65, L157.1, L186, L187 |
Гаплогруппа I-M253 , также известная как i1 , является Y. гаплогруппой хромосом Генетические маркеры, подтвержденные в качестве идентификации I-M253, являются SNPS M253, M307.2/P203.2, M450/S109, P30, P40, L64, L75, L80, L81, L118, L121/S62, L123, L124/S64, L125/S65, L157.1, L186 и L187. [ 7 ] Это основная ветвь гаплогруппы I-M170 (i*).
Считается, что гаплогруппа I1 присутствовал среди верхних палелитических европейских охотников-собирателей в качестве незначительной линии, но из-за его почти тотального отсутствия в образцах до- неолитической ДНК она не может быть очень распространенной. Образцы неолита I1 также очень редки, что указывает на быстрой дисперсию, связанную с эффектом основателя в северном бронзовом веке . Сегодня он достигает своих пиковых частот в Швеции (52 процента мужчин в округе Вестра Гёталанд ) и Западной Финляндии (более 50 процентов в Сатакунта ). провинции [ 8 ] С точки зрения средних значений национального уровня, I-M253 встречается у 38-39% шведских мужчин, [ 9 ] [ 10 ] [ 8 ] 37% норвежских мужчин, [ 11 ] [ 12 ] [ 13 ] 34,8% датских мужчин, [ 14 ] [ 15 ] [ 16 ] 34,5% исландских мужчин, [ 17 ] [ 18 ] [ 19 ] и около 28% финских мужчин. [ 20 ] Брайан Сайкс в своей книге «Кровь острова » 2006 года дает членам - и условно -основательному патриарху I1 название « Водан ». [ 21 ]
Все известные живые члены спускаются от общего предка в 6 раз моложе общего предка с i2. [ 1 ]
Перед реклассификацией в 2008 году, [ 22 ] Группа была известна как I1A , имя, которое с тех пор было переназначено в первичную ветвь, Haplogroup I-DF29. Другими основными ветвями I1 (M253) являются I1B (S249/Z131) и I1C (Y18119/Z17925).
Более 99% живых мужчин с I1 принадлежат к филиалу DF29, которая, по оценкам, появилась в 2400 г. до н.э. [ 23 ] [ 24 ] Все мужчины DF29 имеют общего предка, родившегося между 2500 и 2400 г. до н.э. [ 25 ] Самым старым древним человеком с I1-DF29 является OLL009, человек из раннего бронзового века Швеции . [ 26 ] [ 27 ]
Происхождение
[ редактировать ]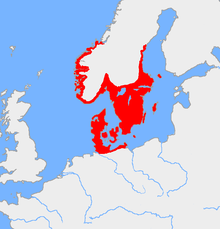
В то время как гаплогруппа I1, скорее всего, расходилась от I* уже 27 000 лет назад на гравитте , до сих пор исследования ДНК могли найти его только у трех палеолитических и мезолитических охотников. По состоянию на ноябрь 2022 года только 6 древних образцов ДНК от человеческих останков, датируемых раньше, чем скандинавский бронзовый век, были назначены в гаплогруппу I1:
- Охотник-собиратель из азильского в Испании в 11 466 г. до н.э., классифицированные как имеющая ныне вымершую ветвь I-Z2699. [ 24 ] [ 28 ]
- Похороны SF11 Дата: 7500 п.н.-Первый-это образец ДНК от скандинавского охотника-собирателя с лейблом SF11, найденным на Stora Karlsö на Готленде . Было обнаружено, что SF11 несет 9 из 312 SNP, которые определяют гаплогруппу I1. SF11 был классифицирован как i1-Z2699*. [ 29 ] [ 30 ] [ 31 ] [ 32 ] SF11 не был назначен конкретной археологической культуре из -за того, что скелет был найден в пещере Стора Фёрвар на Стора Карлсё.
- Похороны BAB5 Дата: 7300-5900 п.н.-второй-это отдельный образец из балатонзема-багодомб с надписью BAB5, из неолитической Венгрии . [ 33 ] Было обнаружено, что BAB5 несет 1 из 312 SNP, которые определяют гаплогруппу I1. BAB5 также может быть классифицирован как i1-Z2699*. [ 34 ] У BAB5 была генетическая близость к другим современным неолитическим фермерам Центральной Европы .
- Похороны179 Дата: 4010-3776 п.н.-Кроме того, третий древний образец I1-от человека, найденного в Кургане , датируемом до позднего неолитического периода кинжала в Скандинавии , помеченном RISE179. [ 35 ] Могила расположена недалеко от АббеКоса на южном побережье Скане Рейз179, имела генетическое сродство к популяциям культуры шестерновой посуды и необычной культуры . [ 35 ]
- Похороны OLL009 Дата: 3930-3750 п.н. - Четвертая древняя образец I1, предшествующая нордическому бронзовому веку (1700–500 гг. До н. более широкий горизонт посуды ». [ 36 ] OLL009 датируется скандинавским поздним неолитом и был обнаружен в похорон в Швеции недалеко от Эллсьё на восточном побережье Сконе. Подобно RISE179, он нес высокий процент происхождения западного степи-хердера и имел генетическое сродство к популяции культуры боевого топора и других популяций горизонта шестерни. [ 37 ] OLL009 имеет Y11204, но, похоже, не имеет Y164553 или Y11205. [ 38 ]
Несмотря на высокую частоту гаплогруппы I1 у современных скандинавов, I1 полностью отсутствует среди ранних сельскохозяйственных образцов ДНК из неолитической скандинавии [ 39 ] [ 40 ] [ 31 ] (Что также имеет место с другими гаплогруппами по всей Европе). За исключением одного образца ДНК (SF11), он также отсутствует у мезолитических охотников-собирателей в Скандинавии. I1 впервые начинает появляться в Скандинавии в заметной частоте во время поздней неолита в сочетании с входом групп, несущих родословные западных трасс в Скандинавию, но не увеличивается по частоте до начала северного бронзового века. [ 35 ] [ 41 ] [ 42 ]
которая датируется раньше, чем скандивный бронзовый век, в настоящее время неизвестно, присутствует ли I1 в качестве редкой гаплогруппы среди скандинавских культур фуражиров Из -за очень низкого числа древних образцов ДНК, которые были назначены I1 , Культура боевого топора , или, если она была приведена в Скандинавию входящими группами, такими как Battle Axe, которые могли ассимилировать ее из таких культур, как Furynelbeaker культура в Центральной Европе; Или сама степи. Будущие исследования, скорее всего, смогут определить, какое из этих двух возможных источников оказывается.
Образцы SF11 и BAB5 отличаются от других древних образцов ДНК, назначенных I1 в том смысле, что они оба, кажется, представляют сейчас, сейчас вытянуты ветви I1, которые еще не полностью превратились в I-M253. Поэтому вряд ли они были наследственными перевозчиками I1, которые все разделяют общего предка, который жил около 2600 г. до н.э.
Согласно исследованию, опубликованному в 2010 году, I-M253 произошел от 3170 и 5000 лет назад в Chalcolithic Europe . [ 2 ] Новое исследование в 2015 году оценило происхождение в размере от 3470 до 5 070 лет назад или от 3180 до 3760 лет назад, используя два различных метода. [ 3 ]
В 2007 году было высказано предположение, что он первоначально рассеялся из области, которая сейчас является Данией . [ 14 ] Тем не менее, профессор д -р Кеннет Нордтведт , штат Монтана , в отношении MRCA, в 2009 году написал в личном послании: «Мы не знаем, где существовал этот человек, но больший бассейн Нижнего Эльбе кажется как сердце I1».
Последние результаты (январь 2022 г.), опубликованные Y-Full Supply I1 (I-M253), были сформированы 27 500 YBP ( 95 CI : 29 800 YBP-25 200 YBP) с TMRCA 4600 YBP (95 CI: 5200 YBP-4000 YBP). Поскольку самая первая дата, рассчитанная оценка TMRCA I1, считается около 2600 г. до н. Филогения I1 показывает подпись быстрого экспансии, подобного звезду. [ 43 ] [ 44 ] Это говорит о том, что I1 превратился из редкого маркера к довольно распространенному в быстром взрыве. [ 3 ]
Структура
[ редактировать ]I-M253 ( M253, M307.2/P203.2, M450/S109, P30, P40, L64, L75, L80, L81, L118, L121/S62, L123, L124/S64, L125/S65, L157.1, L186 и L187 ) или i1 [ 7 ]
- I-DF29 (DF29/S438); I1a
- I-CTS6364 (CTS6364/Z2336); I1a1
- FGC20030; I1a1a ~
- S4767; I1a1a1 ~
- I-M227; I1a1a1a1a
- A394; I1a1a2 ~
- Y11221; I1a1a3 ~
- A5338; I1a1a4 ~
- S4767; I1a1a1 ~
- CTS10028; I1a1b
- I-L22 (L22/S142); I1a1b1
- CTS11651/Z2338; I1a1b1a ~
- I-P109; I1a1b1a1
- I-Y3662; I1a1b1a1e ~
- I-S14887; I1a1b1a1e2 ~
- I-Y11203; I1a1b1a1e2d ~
- I-Y29630; I1a1b1a1e2d2 ~
- I-Y11203; I1a1b1a1e2d ~
- I-S14887; I1a1b1a1e2 ~
- I-Y3662; I1a1b1a1e ~
- CTS6017; I1a1b1a2
- I-L205 (L205.1/L939.1/S239.1); I1a1b1a3
- CTS6868; I1a1b1a4
- I-Z74; I1a1b1a4a
- CTS2208; I1a1b1a4a1 ~
- I-L287; I1a1b1a4a1a
- I-L258 (L258/S335); I1A1B1A4A1A1
- I-L287; I1a1b1a4a1a
- I-L813; I1A1B1A4A2
- FGC12562; I1a1b1a4a3 ~
- CTS2208; I1a1b1a4a1 ~
- I-Z74; I1a1b1a4a
- I-P109; I1a1b1a1
- CTS11603/S4744; I1a1b1b ~
- I-FT40464
- I-Y19934
- I-L300 (L300/S241); I1a1b1b1a1
- I-Y31032
- I-Y32014
- I-Y22918
- I-Y21972
- I-Y24013
- I-Y24015
- I-Y31032
- I-Y19933
- I-Y19932
- I-Y22015
- I-FT57000
- I-Y22015
- I-Y19932
- I-L300 (L300/S241); I1a1b1b1a1
- I-Y19934
- I-FT40464
- CTS11651/Z2338; I1a1b1a ~
- FGC10477/Y13056; I1a1b2
- A8178, A8182, A8200, A8204; I1a1b3 ~
- F13534.2/Y17263.2; I1a1b4 ~
- I-L22 (L22/S142); I1a1b1
- FGC20030; I1a1a ~
- I-Z58 (S244/Z58); I1A2
- I-Z59 (S246/Z59); I1a2a
- I-Z60 (S337/Z60, S439/Z61, Z62); I1A2A1
- I-Z140 (Z140, Z141)
- I-L338
- I-F2642 (F2642)
- I-Z73
- I-L1302
- I-L573
- I-L803
- I-Z140 (Z140, Z141)
- I-Z382; I1A2A2
- I-Z60 (S337/Z60, S439/Z61, Z62); I1A2A1
- I-Z138 (S296/Z138, Z139); I1a2b
- I-Z2541
- I-Z59 (S246/Z59); I1a2a
- I-Z63 (S243/Z63); I1A3
- I -B151; I1a3a
- I-al849.2; I1A3A1
- I -B351; I1A3A2
- I-CTS10345
- I-Y10994
- I-Y7075
- I-CTS10345
- I-S2078
- I-S2077
- I-Y2245 (Y2245/PR683)
- I-L1237
- I-FGC9550
- I-S10360
- I-S15301
- I-Y7234
- I-L1237
- I-Y2245 (Y2245/PR683)
- I-S2077
- I -by62 (by62); I1A3A3
- I -B151; I1a3a
- I-CTS6364 (CTS6364/Z2336); I1a1
- I-Z131 (Z131/S249); I1b
- I-CTS6397 ; I1b1
- I-Z17943 (Y18119/Z17925, S2304/Z17937); I1c
Историческое расширение
[ редактировать ]
Гаплогруппа I1, а также подклады R1B, такие как R1B-U106 и подклады R1A, такие как R1A-Z284, тесно связаны с германскими народами и связаны с протогерманскими носителями скандичного бронзового века . [ 45 ] [ 46 ] Текущее исследование ДНК показывает, что I1 был близок к отсутствию в большинстве Европы за пределами Скандинавии и Северной Германии до периода миграции . Расширение I1 напрямую связано с расширением германских племен. Начиная с 900 г. до н.э., германские племена начали выходить из Южной Скандинавии и Северной Германии в близлежащие земли между Эльбой и Одер. Между 600 и 300 до н.э. Германская миграция в эту область привела к формированию культуры Wielbark , которая связана с готами . [ 47 ]
I1-Z63 был прослежен до места захоронения Ковалево в Польше, которое датируется римским железным веком . В 2017 году польские исследователи могли успешно назначить гаплогруппы иплогруппы YDNA 16 человек, которые были похоронены на месте. Из этих 16 человек 8 принадлежали I1. С точки зрения подкладов, три принадлежали I-Z63 и, в частности, подклад I-L1237. [ 48 ] Археологический сайт Kowalewko был связан с культурой Wielbark. Поэтому подклад I-L1237 I-Z63 можно рассматривать в некоторой степени как генетический индикатор готического племени поздней древности. I1-Z63 также был обнаружен в захоронениях, связанном с Готом и Ломбардом, остается в Колленьо, Италия. [ 49 ] [ 50 ] Кладбище датируется до конца 6-го века и далее предполагает, что I1-Z63 и нижестоящие подклады связаны с ранне средневековыми готическими миграциями.
В 2015 году исследование ДНК проверило гаплогруппы Y-ДНК из 12 образцов, датированных 300–400 г. н.э. от саксонии-анхальта в Германии. 8 из них принадлежали к гаплогруппе I1. Это доказательство ДНК согласуется с историческими миграциями германских племен из Скандинавии в Центральную Европу. [ 51 ]
Кроме того, I1-Z63 был обнаружен в поздней античности Crypta Balbi в Риме, на этот раз с нижним подкладом I-Y-Y7234. [ 52 ] Материальные результаты, связанные с ломбардами, были раскопаны в Cryppa Balbi.
Вилла Pla de l'horta возле Жироны в Испании расположена в непосредственной близости от некрополя с серией гробниц, связанных с Visigoths . Могильные товары и типология гробниц указывают на беспорядочное происхождение людей. Небольшое количество людей, похороненных на месте, было отобрано для анализа ДНК в исследовании 2019 года. Один из образцов принадлежал гаплогруппе I1. [ 53 ] Этот вывод согласуется с общим наследственным происхождением Всиготов и Остроготов .
Англосаксонское . поселение Британии ввело I1 на Британских островах [ 54 ]
В возрасте викингов I-M253 увидел еще одно расширение. Margaryan et al. 2020 проанализировал 442 человека викингов из различных археологических мест в Европе. I-M253 был наиболее распространенной Y-HAPLogroup, обнаруженной в исследовании. Норвежские и датские викинги принесли больше I1 в Великобританию и Ирландию, в то время как шведские викинги представили его России и Украине и принесли больше этого в Финляндию и Эстонию. [ 55 ]
Географическое распределение
[ редактировать ]I-M253 встречается с самой высокой плотностью в северной Европе и других странах, которые пережили обширную миграцию из Северной Европы, либо в период миграции , эпохи викингов или современного времени. Это найдено во всех местах, внесенных норвежством.
В современную эпоху значительное население I-M253 также укоренилось в странах иммигрантов и бывших европейских колониях, таких как Соединенные Штаты , Австралия , Новая Зеландия и Канада .
| Население | Размер выборки | Я (всего) | I1 (I-M253) | I1a1a (i-m227) | Источник |
|---|---|---|---|---|---|
| Албанцы ( Тирана ) | 55 | 21.82%=(12/55) | 3.6%=(2/55) | 0.0 | Battaglia et al. 2008 |
| Албанцы ( Северная Македония ) | 64 | 17.2%=(11/64) | 4.7%=(3/64) | 0.0 | Battaglia et al. 2008 |
| Албанцы ( Тирана ) Албанцы ( Северная Македония ) |
55+64=119 | 19.33%=(23/119) | 4.2%=(5/119) | 0.0 | Battaglia et al. 2008 |
| Косово Албанцы ( Пристина ) | 114 | 7.96%=(9/114) | 5.31%=(6/114) | 0.0 | Перич и соавт. 2005 |
| Албанцы ( Тирана ) Албанцы ( Северная Македония ) Косово Албанцы ( Пристина ) |
55+64+114=233 | 13.73%=(32/233) | 4.72%=(11/233) | 0.0 | Перич и соавт. 2005 Battaglia et al. 2008 |
| Австрия | 43 | 9.3 | 2.3 | 0.0 | Anderhill et al. 2007 |
| Беларусь: Vitbsk | 100 | 15 | 1.0 | 0.0 | Anderhill et al. 2007 |
| Беларусь: Брест | 97 | 20.6 | 1.0 | 0.0 | Anderhill et al. 2007 |
| Босния | 100 | 42 | 2.0 | 0.0 | Швеция и соавт. 2004 |
| Болгария | 808 | 26.6 | 4.3 | 0.0 | Karachanak et al. 2013 |
| Чешская Республика | 47 | 31.9 | 8.5 | 0.0 | Anderhill et al. 2007 |
| Чешская Республика | 53 | 17.0 | 1.9 | 0.0 | Швеция и соавт. 2004 |
| Дания | 122 | 39.3% (48/122) | 34.8% (40/122) | 0.0 | Anderhill et al. 2007 |
| Англия | 104 | 19.2 | 15.4 | 0.0 | Anderhill et al. 2007 |
| Эстония | 210 | 18.6 | 14.8 | 0.5 | Швеция и соавт. 2004 |
| Эстония | 118 | 11.9 | Lappalainen et al. 2008 | ||
| Финляндия (национальная) | 28.0 | Lappalainen et al. 2006 | |||
| Финляндия: Запад | 230 | 40% (92/230) | Lappalainen et al. 2008 | ||
| Финляндия: Восток | 306 | 19% (58/306) | Lappalainen et al. 2008 | ||
| Финляндия: регион Сатакунта | 50+ | Lappalainen et al. 20089 | |||
| Франция | 58 | 17.2 | 8.6 | 1.7 | Anderhill et al. 2007 |
| Франция | 12 | 16.7 | 16.7 | 0.0 | Cann et al. 2002 |
| Франция (низкая нормандия) | 42 | 21.4 | 11.9 | 0.0 | Швеция и соавт. 2004 |
| Германия | 125 | 24 | 15.2 | 0.0 | Anderhill et al. 2007 |
| Греция | 171 | 15.8 | 2.3 | 0.0 | Anderhill et al. 2007 |
| Венгрия | 113 | 25.7 | 13.3 | 0.0 | Швеция и соавт. 2004 |
| Ирландия | 100 | 11 | 6.0 | 0.0 | Anderhill et al. 2007 |
| Volga Tatars | 53 | 13.2 | 11.3 | 0.0 | Трофимова 2015 |
| Латвия | 113 | 3.5 | Lappalainen et al. 2008 | ||
| Литва | 164 | 4.9 | Lappalainen et al. 2008 | ||
| Нидерланды | 93 | 20.4 | 14 | 0.0 | Anderhill et al. 2007 |
| Норвегия | 1766 | 37% (653/1766) | Stenersen et al. 2006 | ||
| Россия (национальная) | 16 | 25 | 12.5 | 0.0 | Cann et al. 2002 |
| Россия: ПСКОВ | 130 | 16.9 | 5.4 | 0.0 | Anderhill et al. 2007 |
| Россия: Кострома | 53 | 26.4 | 11.3 | 0.0 | Anderhill et al. 2007 |
| Россия: Смоленск | 103 | 12.6 | 1.9 | 0.0 | Anderhill et al. 2007 |
| Россия: Воронез | 96 | 19.8 | 3.1 | 0.0 | Anderhill et al. 2007 |
| Russia: Arkhangelsk | 145 | 15.8 | 7.6 | 0.0 | Anderhill et al. 2007 |
| Россия: казаки | 89 | 24.7 | 4.5 | 0.0 | Anderhill et al. 2007 |
| Россия: Карелианцы | 140 | 10 | 8.6 | 0.0 | Anderhill et al. 2007 |
| Россия: Карелианцы | 132 | 15.2 | Lappalainen et al. 2008 | ||
| Россия: Вепс | 39 | 5.1 | 2.6 | 0.0 | Anderhill et al. 2007 |
| Словакия | 70 | 14.3 | 4.3 | 0.0 | Швеция и соавт. 2004 |
| Словения | 95 | 26.3 | 7.4 | 0.0 | Anderhill et al. 2007 |
| Швеция (национальная) | 160 | 35.6% (57/160) | Lappalainen et al. 2008 | ||
| Швеция (национальная) | 38.0 | Lappalainen et al. 2009 | |||
| Швеция: Вестра Гёталанд | 52 | Lappalainen et al. 2009 | |||
| Швейцария | 144 | 7.6 | 5.6 | 0.0 | Швеция и соавт. 2004 |
| Турция | 523 | 5.4 | 1.1 | 0.0 | Anderhill et al. 2007 |
| Украина: ЛВИВ | 101 | 23.8 | 4.9 | 0.0 | Anderhill et al. 2007 |
| Украина: Иванов-Франкив | 56 | 21.4 | 1.8 | 0.0 | Anderhill et al. 2007 |
| Украина: Хмельниц | 176 | 26.2 | 6.1 | 0.0 | Anderhill et al. 2007 |
| Ukraine: Cherkasy | 114 | 28.1 | 4.3 | 0.0 | Anderhill et al. 2007 |
| Украина: Билхород | 56 | 26.8 | 5.3 | 0.0 | Anderhill et al. 2007 |
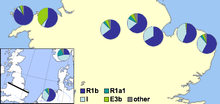
В 2002 году Майкл Э. Уил и его коллеги опубликовали статью, демонстрирующие генетические доказательства различий в популяции между английским и валлийским населением, в том числе заметно более высокий уровень гаплогруппы Y-ДНК в Англии, чем в Уэльсе. Они считали это убедительным доказательством англосаксонского массового вторжения в восточную Великобританию из Северной Германии и Дании в течение периода миграции . [ 56 ] Авторы предположили, что популяции с большими пропорциями гаплогруппы I1 происходили из северной Германии или южной скандинавской, особенно Дании, и что их предки мигрировали через Северное море с англосаксонскими миграциями и датскими викингами . Основным требованием исследователей было
Требуется то, что англосаксонское иммиграционное событие, затрагивающее 50–100% центрального английского генового пула мужского пола. Мы отмечаем, однако, что наши данные не позволяют нам различать событие, которое просто добавлено в генный пул коренного английского языка, от одного, где коренные мужчины были перемещены в другом месте или тот, где коренные мужчины были уменьшены в количестве ... Это исследование показывает То, что валлийская граница была скорее генетическим барьером для англосаксонского гена y хромосомы, чем Северное море ... Эти результаты указывают на то, что политическая граница может быть более важной, чем геофизическая Один из популяций генетического структурирования.

В 2003 году Кристиан Капелли и его коллеги опубликовали статью, которые поддерживали, но модифицировали выводы Уила и коллег. [ 57 ] В этой статье, в которой выбрали Великобритания и Ирландия на сетке, обнаружила меньшую разницу между валлийскими и английскими образцами, с постепенным снижением частоты гаплогруппы I1, движущейся на запад на юге Великобритании. Результаты предположили авторам, что норвежские захватчики викингов оказали сильное влияние на северную область Британских островов, но что как английские, так и шотландские выборки на английском, так и на материковом уровне оказывают влияние немецкого/датского.
Видимые члены I-M253
[ редактировать ]Благодаря прямым тестированию или тестированию их потомков и генеалогических данных, были показано, что следующие известные люди были I-M253:
- Александр Гамильтон , отец -основатель Соединенных Штатов . [ 58 ]
- Варангианский ) , Шмон , который, как говорят, является основателем российской семьи Воронтсова Нобла (включая принца Михаила Семонович Воронтсова и Иллариона Иванович Воронтов-Дашков принадлежавшие к гаплогруппе I1-Y15024. [ 59 ] [ 60 ] [ 61 ] [ 62 ]
- Принц Рурикид ( Свиатопольк Акционированный сын Владимира Великого ) принадлежал I1-S2077. [ 63 ] [ 64 ] [ 65 ]
- Биргер Ярл , герцог Швеции Восточного Гейтса дома Бьялбо , основатель Стокгольма ; Его останки были эксгумированы и протестированы в 2002 году и признаны I-M253. [ 66 ] Дом Бьялбо также предоставил три царя Норвегии и одного короля Дании в 14 веке.
- Британский музыкант Гордон Самнер, известный как Стинг [ 67 ]
- Уильям Брэдфорд , губернатор Plymouth Colony [ 68 ]
- Уильям Брюстер , ранний колонист, который эмигрировал в Америку на Mayflower [ 68 ]
- Генерал Конфедерации Роберт Э. Ли . Другими видными членами семьи Ли Вирджинии и Мэриленда были Ричард Ли I и Ричард Генри Ли . [ 69 ]
- Дом Гримальди принадлежит к скандинавскому подкладу I1, ниже по течению от I1-Y3549. [ 70 ]
- Президент Соединенных Штатов Эндрю Джексон . [ 71 ] [ 72 ]
- Русский писатель Лео Толстой . [ 73 ] [ 74 ]
- Исландский историк и поэт Снорри Стурлусон [ 73 ]
- Шведский ученый и богослов Эмануэль Сведенборг [ 75 ] [ 76 ]
- Сиер Ван Ренсбург , Фермер -Патриотическая фигура и Мистик. [ 77 ] [ 78 ]
- Бьорн Уолруос , финский бизнесмен и миллионер. [ 73 ]
- Финский математик Рольф Неванлинна . [ 79 ] [ 80 ] [ 81 ]
- Американский изобретатель Сэмюэль Морс . [ 82 ] [ 83 ] [ 84 ]
- Шведские футболисты Себастьян Ларссон и его отец Сванте Ларссон принадлежат к I1-Y24470. [ 85 ] [ 86 ] [ 87 ] [ 88 ]
- Шведский YouTuber Felix Kjellberg (Pewdiepie) принадлежит I1-L22. [ 89 ]
- Шведский актер Бьорн Андресен принадлежит к гаплогруппе I1-L22. [ 90 ] [ 91 ] [ 92 ] [ 93 ] Его предок Йохан Андресен жил по обе стороны шведской норвежской границы. [ 94 ] [ 95 ]
- Американский актер Крис Пайн принадлежит к гаплогруппе I1-A13819. [ 96 ] [ 97 ]
- Шведский саами -хоккеист Бёрем Салминг . [ 98 ]
- Американский колонист Эдмунд Райс .
- Немецкий композитор Людвиг Ван Бетховен . [ 99 ]
Маркеры
[ редактировать ]
Ниже приведены технические характеристики для известных мутаций гаплогруппной SNP и STR I-M253.
Имя: M253 [ 100 ]
- Тип: SNP
- Источник: М (Питер Андерхилл из Стэнфордского университета )
- Положение: Chry: 13532101..13532101 (+ Strand)
- Положение (пара оснований): 283
- Общий размер (пары оснований): 400
- Длина: 1
- ISGG HG: I1
- Праймер F (вперед 5 ′ → 3 ′): gcaacaatgaggttttttttg
- Праймер R (обратный 5 ′ → 3 '): cagctcccctctatgcagttt
- YCC HG: I1
- Нуклеотидные аллели изменяются (мутация): C на t
Имя: M307 [ 101 ]
- Тип: SNP
- Источник: M (Питер Андерхилл)
- Позиция: Chry: 21160339..21160339 (+ strand)
- Длина: 1
- ISGG HG: I1
- Праймер F: ttattggcatttcaggaagtg
- Primer R: Gggtgaggcaggaaaaatagc
- YCC HG: I1
- Нуклеотидные аллели изменяются (мутация) g на :
Имя: P30 [ 102 ]
- Тип: SNP
- Источник: PS ( Майкл Хаммер из Университета Аризоны и Джеймс Ф. Уилсон , в Эдинбургском университете)
- Позиция: Chry: 13006761..13006761 (+ strand)
- Длина: 1
- ISGG HG: I1
- Праймер F: GGTGGGCTTTTTTTTAAAAAAAAAAA
- Праймер R: Agccaaataccagtcgtcac
- YCC HG: I1
- Нуклеотидные аллели изменяются (мутация): g на
- Регион: ARSDP
Имя: P40 [ 103 ]
- Тип: SNP
- Источник: PS (Майкл Хаммер и Джеймс Ф. Уилсон)
- Позиция: Chry: 12994402..129944402 (+ Strand)
- Длина: 1
- ISGG HG: I1
- Праймер F: ggagaaaaggtgagaaacc
- Праймер R: Ggacaagggggcagatt
- YCC HG: I1
- Нуклеотидные аллели изменяются (мутация): C на t
- Регион: ARSDP
Смотрите также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а беременный в "I1 ytree" . Yfull.com. 2022-04-06 . Получено 2022-04-19 .
- ^ Jump up to: а беременный Педро Соарес, Алессандро Ахилли, Орнелла Семино, Уильям Дэвис, Винсент Маколей, Ханс-Юрген Бандельт, Антонио Торрони и Мартин Б. Ричардс, Археогенетика Европы, Current Biology , vol. 20 (23 февраля 2010 г.), R174 - R183. Гаплогруппа ИДНК I: Подклад I1 , ДНК семейного дерева,
- ^ Jump up to: а беременный в Батини С., Халласт П., Задик Д., Делсер П.М., Бенаццо А., Гиротто С. и др. (Май 2015). «Масштабное недавнее расширение европейских патрилиниз, показанных в результате повторного воздействия населения» . Природная связь . 6 : 7152. Bibcode : 2015natco ... 6.7152b . doi : 10.1038/ncomms8152 . PMC 4441248 . PMID 25988751 .
- ^ Rootsi S, Magri C, Kivisild T, Benuzzi G, Help H, Bermisheva M, et al. (Июль 2004 г.). «Филогеография гаплогруппы Y-хромосомы I выявляет различные домены доисторического потока генов в Европе» (PDF) . Американский журнал человеческой генетики . 75 (1): 128–37. doi : 10.1086/422196 . PMC 1181996 . PMID 15162323 . Архивировано из оригинала (PDF) на 2009-06-24 . Получено 2008-03-20 .
- ^ Anderhill PA, Myres NM, Rootsi S, Chow CT, Lin AA, Otillar RP, et al. (2007). «Новые филогенетические отношения для гаплогруппы Y-хромосомы I: переосмысление его филогеографии и предыстории». В Mellars P, Boyle K, Bar-Yosef O, Stringe C (Eds.). Переосмысление человеческой революции . Кембридж, Великобритания: Монографии Института Макдональда. С. 33–42. ISBN 978-1-902937-46-5 .
- ^ Lappalainen, T.; Laitinen, v.; Салмела, Е.; Андерсен, П.; Huoponen, K.; Savontus, M.-L.; Lahermo, P. (2008). «Миграционные волны в регион Балтийского моря» . Анналы человеческой генетики . 72 (3): 337–348. Doi : 10.1111/j.1469-1809.2007.00429.x . PMID 18294359 . S2CID 32079904 .
- ^ Jump up to: а беременный ISOGG, гаплогруппа Y-ДНК I и ее подклады-2017 (31 января 2017 г.).
- ^ Jump up to: а беременный Lappalainen T, Laitinen V, Salmela E, Andersen P, Huoponen K, Savontaus ML, Lahermo P (май 2008). «Миграционные волны в регион Балтийского моря» . Анналы человеческой генетики . 72 (Pt 3): 337–348. doi : 10.1111/j.1469-1809.2007.00429.x . PMID 18294359 . S2CID 32079904 .
- ^ Lappalainen T, Hannelius U, Salmela E, Von Döbeln U, Lindgren CM, Huoponen K, et al. (Январь 2009 г.). «Структура популяции в современной Швеции-анализ Y-хромосомальной и митохондриальной ДНК». Анналы человеческой генетики . 73 (1): 61–73. doi : 10.1111/j.1469-1809.2008.00487.x . PMID 19040656 . S2CID 205598345 .
- ^ «FamilyStriedna - Швеция ДНК -проект - Sverige Projects» .
- ^ BM, Central M, Норвегия" Forensic Science International 164 (1): 10–1 doi : 10.1016/j.forciint . PMID 16333760 .
- ^ «FamilyStriedna - проект ДНК Норвегии - NorgeProsektet» . FamilyStreedna.com . Получено 2020-11-26 .
- ^ "Y-DNA Happlagrpper" . Норвегии Норвеж Проект Показан 2020-12-27 .
- ^ Jump up to: а беременный Петр А. Андерхилл и др., Новые филогенетические отношения для гаплогруппы Y-хромосомы I: переосмысление ее филогеографии и предыстории, в переосмыслении человеческой революции (2007), с. 33–42. P. Mellars, K. Boyle, O. Bar-Yosef, C. Stringer (Eds.) Институт археологических исследований Макдональда, Кембридж, Великобритания.
- ^ Санчес JJ (2004). «Y -хромосома SNP Гаплогруппы в датчане, гренландцев и сомалийцах» (PDF) . Международная серия Конгресса . 1261 : 347–49. doi : 10.1016/s0531-5131 (03) 01635-2 -через isfg.org.
- ^ «FamilyTriedna - DENMARN DNA Project» . FamilyStreedna.com . Получено 2020-12-10 .
- ^ Helgason A, Sigureth Ardóttir S, Nicholson J, Sykes B, Hill EW, Bradley DG, et al. (Сентябрь 2000). «Оценка скандинавского и гэльского происхождения у мужских поселенцев Исландии» . Американский журнал человеческой генетики . 67 (3): 697–717. doi : 10.1086/303046 . PMC 1287529 . PMID 10931763 .
- ^ «Большой северный региональный проект Y-DNA» . Семейство . Апрель 2021 года.
- ^ Ebenesdotir SS, Sandoval-Valasco M, Gunnarsdótir ED, Jagadesan A, Guddarsdótir VB, Thordardót EL, et al. (Июнь 2018 г.). «Древние геном из Исландии показывают создание человеческой популяции » Наука . 360 (6392): 1028–1 Bibcode : 2018sci ... 360.1028e Doi : 10.1126/ science.aar2 HDL : 10852/7 29853688PMID
- ^ Lappalainen T, Koivumäki S, Salmela E, Huoponen K, Sistonen P, Savontaus ML, Lahermo P (июль 2006 г.). «Региональные различия между финнами: y-хромосомная перспектива». Ген . 376 (2): 207–15. Doi : 10.1016/j.gene.2006.03.004 . PMID 16644145 .
- ^ «Кровь островов: изучение генетических корней нашей истории племени» . История Ирландия . 2013-02-22 . Получено 2020-12-10 .
- ^ Карафет Т.М., Мендес Ф.Л., Мейлерман М.Б., Андерхилл П.А., Зегура С.Л., Хаммер Мф (май 2008 г.). «Новые бинарные полиморфизмы изменяют и увеличивают разрешение человеческого хромосомного дерева гаплогруппы человека» . Исследование генома . 18 (5): 830–38. doi : 10.1101/gr.7172008 . PMC 2336805 . PMID 18385274 .
- ^ Батини С., Халласт П., Задик Д., Делсер П.М., Бенаццо А., Гиротто С. и др. (Май 2015). «Масштабное недавнее расширение европейских патрилиниз, показанных в результате повторного воздействия населения» . Природная связь . 6 (1): 7152. Bibcode : 2015natco ... 6.7152b . doi : 10.1038/ncomms8152 . PMC 4441248 . PMID 25988751 .
- ^ Jump up to: а беременный «Добро пожаловать в FamilyTreedna Discover (бета)» . FamilyStreedna Discover (бета) . Получено 2022-12-25 .
- ^ "I-DF29 Ytree" . yfull.com . Получено 2022-12-25 .
- ^ Malmström H, Günther T, Svensson EM, Juras A, Fraser M, Munters AR, et al. (Октябрь 2019). скандинавского боевого топора «Геномное происхождение Разбирательство. Биологические науки . 286 (1912): 20191528. DOI : 10.1098/rspb.2019.1528 . PMC 6790770 . PMID 31594508 .
- ^ "I-Y1204 Ytree" . yfull.com . Получено 2022-12-25 .
- ^ "HAPLOTREE.INFO - ANCIONTDNA.INFO. Карта на основе всей древней ДНК против 2.07.26" .
- ^ Гюнтер Т. (2017). «Геномика мезолитической скандинавии выявляет маршруты колонизации и высокошитую адаптацию» (PDF) . Природа . 23 : 6 - через Biorxiv.
- ^ Репозиторий SF11-Large, Stora Karlsö I-Z2699*. «HAPLOTREE.INFO Образец: SF11» . haplotree.info .
{{cite web}}: CS1 Maint: числовые имена: список авторов ( ссылка ) - ^ Jump up to: а беременный Skoglund P, Malmström H, Omrak A, Raghavan M, Valdiosera C, Günther T, et al. (Май 2014). «Геномное разнообразие и примеси отличаются от скандинавских фуражиров и фермеров каменного века» . Наука . 344 (6185): 747–50. Bibcode : 2014sci ... 344..747S . doi : 10.1126/science.1253448 . PMID 24762536 . S2CID 206556994 .
- ^ "FamilyStreedna.com I-Z2699 Дерево" . Семейство . Апрель 2021 года.
- ^ Szécsényi-Nagy A, Brandt G, Haak W, Keerl V, Jakucs J, Möller-Rieker S, et al. (Апрель 2015). «Отслеживание генетического происхождения первых фермеров Европы раскрывает понимание их социальной организации» . Разбирательство. Биологические науки . 282 (1805). doi : 10.1098/rspb.2015.0339 . PMC 4389623 . PMID 25808890 .
- ^ "BAB5 I-Z2699*" . haplotree.info .
- ^ Jump up to: а беременный в Allentoft Me, Sikora M, Sjögren KG, Rasmussen S, Rasmussen M, Stendup J, et al. (Июнь 2015 г.). «Геномика населения евразии бронзового века» . Природа . 522 (7555): 167–72. Bibcode : 2015natur.522..167a . doi : 10.1038/nature14507 . PMID 26062507 . S2CID 4399103 .
- ^ скандинавского боевого топора «Yfull | Геномное происхождение yfull.com . Получено 2021-01-24 .
- ^ Malmstrom H (июль 2019). происхождение скандинавского боевого топора « Геномное Uppsala Genomics . 1 : 3 - через jakobssonlab.iob.uu.se/.
- ^ "I-Y1204 Ytree" .
- ^ Sánchez-Quinto F, Malmström H, Fraser M, Girdland-Flink L, Svensson EM, Simões LG, et al. (Май 2019). «Мегалитические гробницы в западной и северной неолитической Европе были связаны с родственным обществом» . Труды Национальной академии наук Соединенных Штатов Америки . 116 (19): 9469–74. Bibcode : 2019pnas..116.9469S . doi : 10.1073/pnas.1818037116 . PMC 6511028 . PMID 30988179 .
- ^ Malmsström H, Linderholm A, Skoglund P, Storå J, Sjödin P, Gilbert MT, et al. (Январь 2015). «Древняя митохондриальная ДНК от северной полосы неолитического выращивания сельского хозяйства в Европе проливает свет на процесс дисперсии» . Философские транзакции Королевского общества Лондона. Серия B, биологические науки . 370 (1660): 20130373. DOI : 10.1098/rstb.2013.0373 . PMC 4275881 . PMID 25487325 .
- ^ Карлссон А.О., Уоллерстрем Т., Гётерстрем А., Холмлунд Г. (август 2006 г.). «Разнообразие Y-хромосомы в Швеции-давняя перспектива» . Европейский журнал человеческой генетики . 14 (8): 963–70. doi : 10.1038/sj.ejhg.5201651 . PMID 16724001 . S2CID 23227271 .
- ^ Malmström H, Günther T, Svensson EM, Juras A, Fraser M, Munters AR, et al. (Октябрь 2019). скандинавского боевого топора «Геномное происхождение Разбирательство. Биологические науки . 286 (1912): 20191528. DOI : 10.1098/rspb.2019.1528 . PMC 6790770 . PMID 31594508 .
- ^ Poznik GD, Xue Y, Mendez FL, Willems TF, Massaia A, Wilson Sayres MA, et al. (Июнь 2016 г.). «Подчеркиваемые всплески в демографии мужчины человека, выведенные из 1244 мировых последовательностей Y-хромосомы» . Природа генетика . 48 (6): 593–99. doi : 10.1038/ng.3559 . HDL : 11858/00-001M-0000-002A-F024-C . PMC 4884158 . PMID 27111036 .
- ^ Вудли М (февраль 2017 г.). «Выбор голоцена для вариантов, связанных со когнитивными способностями: сравнение древних и современных геномов» (PDF) . biorxiv.org/ . doi : 10.1101/109678 . S2CID 196631764 . Получено 27 января 2021 года .
- ^ Шмидт К.Х. (1991). «Кельты и этногенез германских людей» . Historische Sprachforschung / Историческая лингвистика . 104 (1): 129–52. ISSN 0935-3518 . JSTOR 40849016 .
- ^ Bergerbrant S (май 2007 г.). «Личность бронзового века: костюм, конфликт и контакт в Северной Европе 1600–1300 гг. До н.э.» (PDF) . Стокгольмские исследования по археологии . 43 : 7–201-через Diva-propal.org.
- ^ Teska M, Michalowski A (2014). «Связь между Вилькопольской и регионом Балтийского моря в римском железе» . Археология Литуана . 14 : 63–77. doi : 10.15388/archlit.2013.0.2641 . S2CID 56295624 .
- ^ Zenczak M, Handschuh L, Juras A, Marcinkowska-Swojak M, Philips A, Piontek J, Stolarek I, Figlerowicz M (2017). Выступление гаплогруппы Y-хромосомы через секвенирование следующего поколения обогащенных древних библиотек ДНК . Антропологическая генетика. п. Номер презентации: AG 16.
- ^ Amorim CE, Vai S, Poth C, Modi A, Koncz I, Hakenbeck S, et al. (Сентябрь 2018 г.). «Понимание варварской социальной организации 6-го века и миграции посредством палеогеномики» . Природная связь . 9 (1): 3547. Bibcode : 2018natco ... 9.3547a . doi : 10.1038/s41467-018-06024-4 . PMC 6134036 . PMID 30206220 .
- ^ Estes R (2020-10-16). «Лонгбардс древняя ДНК из Паннонии и Италии - что говорит нам их ДНК? Вы связаны?» Полем ДНКэксель - генетическая генеалогия . Получено 2020-12-11 .
- ^ Лабудде Д (июль 2015 г.). «Гендерное распределение раскопок находит из римского имперского и миграционного периода» . Researchgate . 1 : 2 - через Researchgate.net.
- ^ Antonio ML, Gao Z, Moots HM, Lucci M, Candilio F, Sawyer S, et al. (Ноябрь 2019). «Древний Рим: генетический перекресток Европы и Средиземного моря» . Наука . 366 (6466): 708–14. Bibcode : 2019sci ... 366..708a . doi : 10.1126/science.aay6826 . PMC 7093155 . PMID 31699931 .
- ^ Olalde I, Mallick S, Patterson N, Rohland N, Villalba-Mouco V, Silva M, et al. (Март 2019). «Геномная история иберийского полуострова за последние 8000 лет» . Наука . 363 (6432): 1230–34. Bibcode : 2019sci ... 363.1230o . doi : 10.1126/science.aav4040 . PMC 6436108 . PMID 30872528 .
- ^ Martiniano R, Caffell A, Holst M, Hunter-Mann K, Montgomery J, Müldner G, et al. (Январь 2016). «Геномные сигналы миграции и непрерывности в Британии перед англосаксами» . Природная связь . 7 (1): 10326. Bibcode : 2016natco ... 710326M . doi : 10.1038/ncomms10326 . PMC 4735653 . PMID 26783717 .
- ^ Margaryan A, Lawson DJ, Sikora M, Racimo F, Rasmussen S, Moltke I, et al. (Сентябрь 2020 г.). «Геномика популяции мира викингов». Природа . 585 (7825): 390–96. Bibcode : 2020nater.585..390M . Biorxiv 10.1101/703405 . doi : 10.1038/s41586-020-2688-8 . HDL : 10852/83989 . PMID 32939067 . S2CID 201195157 .
- ^ Weale Me, Weiss DA, Jager RF, Bradman N, Thomas Mg (июль 2002 г.). «Y-хромосома свидетельствует о англосаксонской массовой миграции» . Молекулярная биология и эволюция . 19 (7): 1008–21. doi : 10.1093/oxfordjournals.molbev.a004160 . PMID 12082121 .
- ^ Capelli C, Redhead N, Abernethy JK, Gratrix F, Wilson JF, Moen T, et al. (Май 2003 г.). «Ай -хромосомная перепись британских островов» . Текущая биология . 13 (11): 979–84. doi : 10.1016/s0960-9822 (03) 00373-7 . HDL : 20.500.11820/8ACB01F3-A7C1-45F5-89DE-B796266D651E . PMID 12781138 . S2CID 526263 .
- ^ «Отец -основатель ДНК» . isogg.org .
- ^ «FamilyStreedna - генетическое тестирование на происхождение, семейная история и генеалогия» . FamilyStreedna.com . Получено 2020-12-10 .
- ^ "Faderslinjen, ДНК" . sikaby.se . Получено 2020-12-10 .
- ^ «FamilyTriedna - Россиадна Проект» . FamilyStreedna.com . Получено 2020-12-10 .
- ^ «Наш вождь викингов на востоке» . Sikaby.se . Получено 2020-12-10 .
- ^ «Образец от Homo Sapiens - Biosample - NCBI» . ncbi.nlm.nih.gov . Получено 2020-12-10 .
- ^ Margaryan A, Lawson DJ, Sikora M, Racimo F, Rasmussen S, Moltke I, et al. (Сентябрь 2020 г.). «Геномика популяции мира викингов» . Природа . 585 (7825): 390–96. Bibcode : 2020nater.585..390M . doi : 10.1038/s41586-020-2688-8 . HDL : 10852/83989 . PMID 32939067 . S2CID 221769227 .
- ^ Duczko W (2004). Викинг Рус: Исследования о присутствии скандинавцев в Восточной Европе . Брилль ISBN 978-90-04-13874-2 .
- ^ Malmström H, Vretemark M, Tillmar A, Durling MB, Skoglund P, Gilbert MT, et al. (Январь 2012 г.). «Поиск основателя Стокгольма-родственного исследования, основанного на Y-хромосомной, аутосомной и митохондриальной ДНК». Анналы анатомии - Анатомиссер Анзегер . Специальный выпуск: древняя ДНК. 194 (1): 138–45. doi : 10.1016/j.aanat.2011.03.014 . PMID 21596538 .
- ^ Британское вторжение, найдя ваши корни
- ^ Jump up to: а беременный "Mayflower DNA Project" . Mayflowerdna.org . Получено 2020-11-23 .
- ^ «FamilyTriedna - исследовательский проект ДНК Ли Фамилии (и Ли, Леа и т. Д.)» . FamilyStreedna.com . Получено 2020-12-10 .
- ^ «FamilyTriedna - Генеалогический проект Гримальди в фтДНК» . FamilyStreedna.com . Получено 2020-12-11 .
- ^ "Jackson DNA Project" . Семейство . 11 декабря 2020 года.
- ^ Hay M (июль 2020 г.). «Происхождение и история гаплогруппы I1 (Y-ДНК)» . Researchgate . 1 : 9.
- ^ Jump up to: а беременный в "Гаплогруппа I1 (Y-DNA) " Епипедия . 2020-12-1
- ^ Петр Толстой. Моя родословная. Выпуск от 18.04.2010 (in Russian) , retrieved 2020-12-15
- ^ "I-BY229 Ytree" . yfull.com . Получено 2020-12-10 .
- ^ «Сведенборг» . Höijen (на шведском языке). Архивировано из оригинала 2020-10-26 . Получено 2020-12-10 .
- ^ «Claas Jansz Van Rensburg, SV/Prog» . geni_family_tree . Получено 2021-01-03 .
- ^ «Янсе /Янсен Ван Ренсбург I-M253 Обсуждение генеалогии» . geni_family_tree . Получено 2021-01-03 .
- ^ «Рольф Х. Неванлинна» . geni_family_tree . Получено 2020-12-26 .
- ^ Оленс (2018-03-30). «I1: Рольф Неванлинна (Né Neovius)» . Потомки гаплогруппы IJ-M429 . Получено 2020-12-26 .
- ^ "Арне Эдвард Неванлинна" . geni_family_tree . Получено 2020-12-26 .
- ^ «Тестирование ДНК Морса/Мосса» . morsesociety.org . Получено 2020-12-10 .
- ^ «FamilyStriedna - проект Морс / Мосс ДНК» . FamilyStreedna.com . Получено 2020-12-10 .
- ^ «Семейное древо Питера Морса» . geni.com . Получено 2020-12-10 .
- ^ «FamilyTriedna - Швеция ДНК -проект - Sverigeprojektet» . FamilyStreedna.com . Получено 2021-02-14 .
- ^ «Муниципалитет Eskilstuna · em GN398 - семья Ларссона, Торшалла около 1900» . Эскилстунский муниципалитет (на шведском языке) . Получено 2021-02-14 .
- ^ "I-Y24470 Ytree" . yfull.com . Получено 2021-02-14 .
- ^ «Предки семьи Ларссона» . hoserudkullen.se . Получено 2021-02-14 .
- ^ Заархивировано в Ghostarchive и на машине Wayback : «Я провел тест ДНК ... (я думаю, теперь я отменен)» - через YouTube .
- ^ «FamilyStriedna - проект ДНК Норвегии - NorgeProsektet» . FamilyStreedna.com . Получено 2021-02-02 .
- ^ Tovseth A (июнь 2018 г.). «Андрезен, Färnskog & Hansen Family Research» . Kjellivar.tripod.com . Получено 2 февраля 2021 года .
- ^ «Люди по имени Андресен | Семейство. Се» . Slaktings.se . Получено 2021-02-02 .
- ^ «Бьорн Андресен: Моя страсть к маме никогда не отвечала - Катарина Хер встречается» . Радио Швеция (на шведском языке) . Получено 2021-02-02 .
- ^ «Йохан Питер Андресен - происхождение» . Ancestry.se . Получено 2021-02-02 .
- ^ «Семейное древо Даниэля Андресена» . Генонет . Получено 2021-02-02 .
- ^ «FamilyStriedna - Pine/Pyne Genealogy DNA Project» . FamilyStreedna.com . Получено 2020-12-10 .
- ^ "Джеймс Пайн, ср" . geni_family_tree . Получено 2020-12-10 .
- ^ Janlind F (20 февраля 2021 года). «Бьянка Сальми о отношениях с Борье:« Чувство ужасного » [ Бьянка Салминг в отношении своих отношений с Борье:« Чувствует себя ужасно »]. Goteborgs-posten (на шведском языке).
- ^ Mercedes (2023-03-26). «Открытие ДНК Бетховена - узнайте, связаны ли вы» . Из кого вы сделаны? Полем Получено 2023-10-07 .
- ^ snpdev. «Справочный отчет о кластере SNP (RefSNP): RS9341296» . nih.gov .
- ^ snpdev. «Справочный отчет SNP (RefSNP): RS13447354» . nih.gov .
- ^ P30 [ Постоянная мертвая ссылка ]
- ^ P40 [ Постоянная мертвая ссылка ]
Дальнейшее чтение
[ редактировать ]- Allentoft Me, Sikora M, Sjögren KG, Rasmussen S, Rasmussen M, Stendup J, et al. (Июнь 2015 г.). «Геномика населения евразии бронзового века» . Природа . 522 (7555): 167–72. Bibcode : 2015natur.522..167a . doi : 10.1038/nature14507 . PMID 26062507 . S2CID 4399103 .
- Brunel S, Bennett EA, Cardin L, Garraud D, Barrand Emam H, Beylier A, et al. (Июнь 2020 г.). «Древние геномы современной Франции раскрывают 7000 лет своей демографической истории» . Труды Национальной академии наук Соединенных Штатов Америки . 117 (23): 12791–98. Bibcode : 2020pnas..11712791B . doi : 10.1073/pnas.1918034117 . PMC 7293694 . PMID 32457149 .
- Malmström H, Gilbert MT, Thomas MG, Brandström M, Storå J, Molnar P, et al. (Ноябрь 2009 г.). «Древняя ДНК раскрывает отсутствие преемственности между неолитическими охотниками-собирателями и современными скандинавтами» . Текущая биология . 19 (20): 1758–62. doi : 10.1016/j.cub.2009.09.017 . PMID 19781941 . S2CID 9487217 .
- Malmström H, Günther T, Svensson EM, Juras A, Fraser M, Munters AR, Pospieszny L, Tõrv M, Lindström J, Götherström A, Storå J, Jakobsson M (октябрь 2019). скандинавского боевого топора «Геномное происхождение Разбирательство. Биологические науки . 286 (1912): 20191528. DOI : 10.1098/rspb.2019.1528 . PMC 6790770 . PMID 31594508 .
- Villalba-Mouco V, Van de Loosdrecht MS, Poth C, Mora R, Martínez-Moreno J, Rojo-Guerra M, et al. (Апрель 2019). «Выживание покойного плейстоценового охотника-собирателя на иберийском полуострове» . Текущая биология . 29 (7). Cell Press : 1169–77.e7. doi : 10.1016/j.cub.2019.02.006 . HDL : 10261/208851 . PMID 30880015 . S2CID 76663708 .
Внешние ссылки
[ редактировать ]- Гаплогруппа I Базы данных
- Проект Aplogroup I1 на ftdna
- Датские демесские региональные ДНК проект в FTDNA
- Гаплогруппа I-P109 Проект
- Проект ДНК Британских островов
- Общие базы данных Y-DNA
Есть несколько баз данных общего доступа с I-M253, в том числе:
- [1] Архивированный 2011-01-04 на машине Wayback
- YHRD: Y-хромосома STR HAPLOTYPE REPORTIPE BATABASE
- I1 ytree