Зрительная кора
| Зрительная кора | |
|---|---|
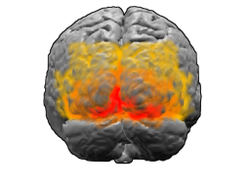 Вид на мозг сзади. Красный = зона Бродмана 17 (первичная зрительная кора); оранжевый = область 18; желтый = область 19 | |
 Мозг показан сбоку, обращенным влево. Вверху: вид снаружи, внизу: разрез посередине. Оранжевый = зона Бродмана 17 (первичная зрительная кора). | |
| Подробности | |
| Идентификаторы | |
| латинский | зрительная кора |
| МеШ | D014793 |
| НейроЛекс ID | nlx_143552 |
| ФМА | 242644 |
| Анатомические термины нейроанатомии | |
головного Зрительная кора мозга — это область коры головного мозга , которая обрабатывает визуальную информацию . Он расположен в затылочной доле . Сенсорная информация, исходящая от глаз, через латеральное коленчатое ядро таламуса проходит и затем достигает зрительной коры. Область зрительной коры, которая получает сенсорную информацию от латерального коленчатого ядра, представляет собой первичную зрительную кору, также известную как зрительная зона 1 ( V1 ), зона Бродмана 17 или полосатая кора. Экстрастриатные области состоят из зрительных областей 2, 3, 4 и 5 (также известных как V2, V3, V4 и V5 или зона Бродмана 18 и вся зона Бродмана 19 ). [1]
Оба полушария мозга включают зрительную кору; зрительная кора левого полушария получает сигналы из правого поля зрения , а зрительная кора правого полушария получает сигналы из левого поля зрения.
Введение
[ редактировать ]Первичная зрительная кора (V1) расположена внутри и вокруг шпорной щели доли затылочной . V1 каждого полушария получает информацию непосредственно от своего ипсилатерального латерального коленчатого ядра , которое получает сигналы от контралатерального зрительного полуполя.
Нейроны зрительной коры активируют потенциалы действия , когда зрительные стимулы появляются в их рецептивном поле . По определению, рецептивное поле — это область внутри всего поля зрения, которая вызывает потенциал действия. Но любой нейрон может лучше всего реагировать на подмножество стимулов в его рецептивном поле. Это свойство называется нейронной настройкой . В более ранних зрительных областях нейроны настраиваются проще. Например, нейрон в V1 может срабатывать на любой вертикальный стимул в своем рецептивном поле. В высших зрительных областях нейроны имеют сложную настройку. Например, в нижней височной коре (ИТ) нейрон может сработать только тогда, когда в его рецептивном поле появляется определенное лицо.
Более того, расположение рецептивных полей в V1 является ретинотопным, то есть соседние клетки в V1 имеют рецептивные поля, соответствующие соседним частям зрительного поля. Эта пространственная организация позволяет систематически представлять визуальный мир в V1. Кроме того, недавние исследования углубились в роль контекстной модуляции в V1, где на восприятие стимула влияет не только сам стимул, но и окружающий контекст, подчеркивая сложные возможности обработки V1 в формировании нашего визуального опыта. [2]
Зрительная кора получает кровоснабжение главным образом из шпорной ветви задней мозговой артерии .
Размер V1, V2 и V3 может различаться в три раза, и эта разница частично наследуется. [3]
Психологическая модель нейронной обработки зрительной информации
[ редактировать ]
Вентрально-дорсальная модель
[ редактировать ]V1 передает информацию по двум основным путям, называемым вентральным потоком и дорсальным потоком. [4]
- Вентральный поток начинается с V1, проходит через зрительную область V2, затем через зрительную область V4 и в нижнюю височную кору (IT-кора). Вентральный поток, иногда называемый «Какой путь», связан с распознаванием формы и представлением объектов. Это также связано с хранением долговременной памяти .
- Дорсальный поток начинается с V1, проходит через зрительную область V2, затем в дорсомедиальную область (DM/V6) и средневисочную область (MT/V5) и в заднюю теменную кору . Спинной поток, иногда называемый «путь куда» или «путь как», связан с движением, представлением местоположения объектов и контролем глаз и рук, особенно когда визуальная информация используется для управления саккадами или тянуться.
Счет «что против где» в вентральных/дорсальных путях был впервые описан Унгерлейдером и Мишкиным . [5]
Совсем недавно Гудейл и Милнер расширили эти идеи и предположили, что вентральный поток имеет решающее значение для зрительного восприятия, тогда как дорсальный поток обеспечивает зрительный контроль квалифицированных действий. [6] Было показано, что зрительные иллюзии, такие как иллюзия Эббингауза, искажают суждения перцептивного характера, но когда субъект отвечает действием, например хватанием, никаких искажений не происходит. [7]
Работа, подобная работе Franz et al. [8] предполагает, что и система действия, и система восприятия в равной степени обманываются такими иллюзиями. Другие исследования, однако, убедительно подтверждают идею о том, что визуальные иллюзии не влияют на такие навыки, как хватание. [9] [10] и предполагают, что диссоциация действия/восприятия является полезным способом охарактеризовать функциональное разделение труда между дорсальными и вентральными зрительными путями в коре головного мозга. [11]
Первичная зрительная кора (V1)
[ редактировать ]Этот раздел нуждается в дополнительных цитатах для проверки . ( Ноябрь 2016 г. ) |


Первичная зрительная кора — наиболее изученная зрительная область мозга. У млекопитающих он расположен в заднем полюсе затылочной доли и представляет собой простейшую и раннюю кортикальную зрительную область. Он узкоспециализирован для обработки информации о статических и движущихся объектах и превосходно распознает образы .Более того, V1 характеризуется ламинарной организацией с шестью отдельными слоями, каждый из которых играет уникальную роль в визуальной обработке. Нейроны в поверхностных слоях (II и III) часто участвуют в локальной обработке и коммуникации внутри коры, тогда как нейроны в более глубоких слоях (V и VI) часто отправляют информацию в другие области мозга, участвующие в визуальной обработке и принятии решений более высокого порядка. изготовление.
Исследования V1 также выявили наличие ориентационно-селективных клеток, которые преимущественно реагируют на стимулы определенной ориентации, способствуя восприятию краев и контуров. Открытие этих ориентационно-селективных клеток сыграло фундаментальную роль в формировании нашего понимания того, как V1 обрабатывает визуальную информацию.
Кроме того, V1 демонстрирует пластичность, что позволяет ему претерпевать функциональные и структурные изменения в ответ на сенсорный опыт. Исследования показали, что сенсорная депривация или воздействие обогащенной среды могут привести к изменениям в организации и реагировании нейронов V1, подчеркивая динамическую природу этого важного центра визуальной обработки. [12] [ нужны разъяснения ]
Первичная зрительная кора, которая определяется ее функцией или стадией зрительной системы, примерно эквивалентна полосатой коре, также известной как зона Бродмана 17, которая определяется ее анатомическим расположением. Название «полосатая кора» происходит от линии Дженнари — характерной полосы, видимой невооруженным глазом.
Стоит отметить, что область Бродмана 17 — это лишь одно подразделение более широких областей Бродмана, которые представляют собой области коры головного мозга, определяемые на основе цитоархитектурных различий. В случае стриарной коры линия Дженнари соответствует полосе, богатой миелинизированными нервными волокнами, что обеспечивает четкий маркер области первичной зрительной обработки.
Кроме того, функциональное значение стриарной коры выходит за рамки ее роли первичной зрительной коры. Он служит важнейшим центром первоначальной обработки визуальной информации, такой как анализ основных характеристик, таких как ориентация, пространственная частота и цвет. Интеграция этих функций в стриарной коре формирует основу для более сложной визуальной обработки, выполняемой в зрительных областях более высокого порядка. Недавние исследования нейровизуализации способствовали более глубокому пониманию динамических взаимодействий внутри полосатой коры и ее связей с другими зрительными и незрительными областями мозга, проливая свет на сложные нейронные цепи, лежащие в основе зрительного восприятия. [13] Он представляет собой миелинизированные аксоны латерального коленчатого тела, оканчивающиеся в четвертом слое серого вещества .
Первичная зрительная кора разделена на шесть функционально различных слоев, обозначенных от 1 до 6. Слой 4, который получает большую часть зрительной информации от латерального коленчатого ядра (LGN), далее делится на 4 слоя, обозначенных 4A, 4B, 4Cα и 4Cβ. . Субламина 4Cα получает в основном магноцеллюлярные входные данные от LGN, тогда как слой 4Cβ получает входные данные от парвоцеллюлярных путей. [14]
Среднее количество нейронов в первичной зрительной коре взрослого человека в каждом полушарии оценивается в 140 миллионов. [15]
Функция
[ редактировать ]Эта статья может быть слишком технической для понимания большинства читателей . ( сентябрь 2016 г. ) |
Начальная стадия обработки изображений в коре головного мозга, известная как V1, играет фундаментальную роль в формировании нашего восприятия визуального мира. V1 обладает тщательно определенной картой, называемой ретинотопической картой, которая сложно организует пространственную информацию из поля зрения. У людей верхний банк шпорной борозды в затылочной доле активно реагирует на нижнюю половину поля зрения, тогда как нижний банк реагирует на верхнюю половину. Это ретинотопическое картирование концептуально представляет собой проекцию зрительного изображения с сетчатки на V1.
Важность этой ретинотопической организации заключается в ее способности сохранять пространственные отношения, присутствующие во внешней среде. Соседние нейроны в V1 реагируют на соседние части поля зрения, создавая систематическое представление визуальной сцены. Это отображение распространяется как по вертикали, так и по горизонтали, обеспечивая сохранение как горизонтальных, так и вертикальных отношений внутри визуального ввода.
Более того, ретинотопическая карта демонстрирует поразительную степень пластичности, адаптируясь к изменениям зрительного опыта. Исследования показали, что изменения в сенсорной информации, например, вызванные зрительной тренировкой или депривацией, могут привести к сдвигам в ретинотопической карте. Эта адаптивность подчеркивает способность мозга реорганизоваться в ответ на изменяющиеся требования окружающей среды, подчеркивая динамичный характер визуальной обработки.
Помимо своей роли в пространственной обработке, ретинотопическая карта в V1 устанавливает сложные связи с другими зрительными областями, образуя сеть, имеющую решающее значение для интеграции разнообразных зрительных функций и построения связного зрительного восприятия. Этот механизм динамического отображения незаменим для нашей способности эффективно ориентироваться и интерпретировать визуальный мир.
Соответствие между конкретными участками V1 и субъективным зрительным полем исключительно точное, вплоть до картирования слепых пятен сетчатки. С эволюционной точки зрения это соответствие является фундаментальной особенностью, обнаруженной у большинства животных, обладающих V1. У людей и других видов животных, имеющих фовеа (колбочки в сетчатке), значительная часть V1 отображается в небольшой центральной части поля зрения — явление, называемое корковым увеличением. Это увеличение отражает повышенную способность представления и обработки данных, относящуюся к центральному полю зрения, необходимую для детальной остроты зрения и обработки изображений с высоким разрешением.
Примечательно, что нейроны в V1 имеют наименьший размер рецептивного поля, что означает самое высокое разрешение, среди микроскопических областей зрительной коры. Эта специализация дает V1 возможность улавливать мелкие детали и нюансы визуального ввода, подчеркивая его ключевую роль в качестве важнейшего узла ранней визуальной обработки и внося значительный вклад в наше сложное и нюансированное визуальное восприятие. [16]
Помимо своей роли в пространственной обработке, ретинотопическая карта в V1 неразрывно связана с другими зрительными областями, образуя сеть, которая способствует интеграции различных зрительных особенностей и построению связного зрительного восприятия. Этот механизм динамического отображения имеет основополагающее значение для нашей способности эффективно ориентироваться и интерпретировать визуальный мир. [17] Соответствие между данным местоположением в V1 и субъективным полем зрения очень точное: даже слепые пятна сетчатки отображаются в V1. С точки зрения эволюции это соответствие очень простое и обнаруживается у большинства животных, обладающих V1. У людей и других животных с ямкой ( колбочками в сетчатке) большая часть V1 отображается в маленькой центральной части поля зрения — явление, известное как корковое увеличение . [18] Возможно, с целью точного пространственного кодирования нейроны в V1 имеют наименьший размер рецептивного поля (то есть самое высокое разрешение) среди всех микроскопических областей зрительной коры.
Свойства настройки нейронов V1 (на что реагируют нейроны) сильно различаются со временем. На раннем этапе (40 мс и далее) отдельные нейроны V1 имеют сильную настройку на небольшой набор стимулов. То есть реакции нейронов могут различать небольшие изменения в зрительной ориентации , пространственных частотах и цветах (как в оптической системе камеры-обскуры , но проецируются на клетки сетчатки глаза, которые сгруппированы по плотности и тонкости). [17] Каждый нейрон V1 последовательно передает сигнал от клетки сетчатки. Более того, отдельные нейроны V1 у людей и других животных с бинокулярным зрением обладают глазным доминированием, а именно настройкой на один из двух глаз. В V1 и первичной сенсорной коре в целом нейроны со схожими настроечными свойствами имеют тенденцию группироваться в кортикальные столбцы . Дэвид Хьюбел и Торстен Визель предложили классическую модель организации кортикальных столбцов в виде кубика льда для двух свойств настройки: доминирования глаза и ориентации. Однако эта модель не может учесть цвет, пространственную частоту и многие другие особенности, на которые настроены нейроны. [ нужна ссылка ] . Точная организация всех этих корковых столбцов в V1 остается горячей темой текущих исследований. Математическое моделирование этой функции сравнивают с преобразованиями Габора . [ нужна ссылка ]
Позже (через 100 мс) нейроны в V1 также становятся чувствительными к более глобальной организации сцены (Lamme & Roelfsema, 2000). [19] Эти свойства ответа, вероятно, обусловлены повторяющейся обработкой обратной связи (влиянием областей коры более высокого уровня на области коры более низкого уровня) и латеральными связями пирамидных нейронов . [20] В то время как связи прямой связи в основном являются движущими, связи обратной связи имеют в основном модулирующий эффект. [21] [22] Фактические данные показывают, что обратная связь, возникающая в областях более высокого уровня, таких как V4, IT или MT, с более крупными и сложными рецептивными полями, может изменять и формировать ответы V1, учитывая контекстуальные или экстраклассические эффекты рецептивного поля. [23] [24] [25]
Визуальная информация, передаваемая в V1, не кодируется с точки зрения пространственного (или оптического) изображения. [ нужна ссылка ] а скорее лучше описать как обнаружение края . [26] Например, для изображения, состоящего из половины черной и половины белой, разделительная линия между черным и белым имеет самый сильный локальный контраст (то есть обнаружение краев) и кодируется, в то время как немногие нейроны кодируют информацию о яркости (черный или белый в зависимости от цвета). се). По мере того, как информация далее передается в последующие зрительные области, она кодируется как все более нелокальные частотно-фазовые сигналы. Обратите внимание, что на этих ранних стадиях корковой зрительной обработки пространственное расположение зрительной информации хорошо сохраняется за счет кодирования локального контраста (обнаружения границ).
Теоретическое объяснение вычислительной функции простых клеток первичной зрительной коры представлено в работе. [27] [28] [29] Описано, как форма рецептивного поля аналогична той, которая была обнаружена при измерениях биологического рецептивного поля, выполненных ДеАнджелисом и соавт. [30] [31] могут быть получены как следствие структурных свойств окружающей среды в сочетании с требованиями внутренней согласованности, чтобы гарантировать согласованное представление изображений в нескольких пространственных и временных масштабах. Также описано, как характерные формы рецептивных полей, настроенные на разные масштабы, ориентации и направления в пространстве изображений, позволяют зрительной системе вычислять инвариантные реакции при естественных преобразованиях изображений на более высоких уровнях зрительной иерархии. [32] [28] [29]
У приматов одной из функций V1 может быть создание карты значимости (выделение важного) на основе визуальных входных данных, чтобы направлять сдвиги внимания, известные как сдвиги взгляда . [33]
Согласно гипотезе значимости V1 , V1 делает это путем преобразования визуальных сигналов в частоту срабатывания нейронов от миллионов нейронов, так что визуальное местоположение, о котором сигнализирует самый активный нейрон, является наиболее заметным местом, привлекающим сдвиг взгляда. Выходные сигналы V1 принимаются верхними холмиками (в среднем мозге), среди других мест, которые считывают действия V1, чтобы направлять сдвиги взгляда.
Различия в размерах V1, по-видимому, также влияют на восприятие иллюзий . [34]
V2
[ редактировать ]| Цветовой центр | |
|---|---|
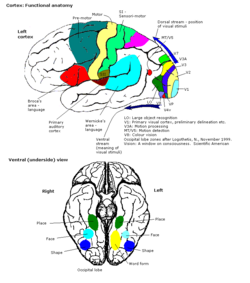 Разница V-регионов. Больше изображений в центре цвета | |
| Идентификаторы | |
| МеШ | D014793 |
| НейроЛекс ID | nlx_143552 |
| ФМА | 242644 |
| Анатомическая терминология | |
Зрительная область V2 , или вторичная зрительная кора , также называемая престриарной корой , [35] получает сильные соединения прямой связи от V1 (прямо и через пульвинар) и отправляет надежные соединения к V3, V4 и V5. Кроме того, он играет решающую роль в интеграции и обработке визуальной информации.
Прямые связи от V1 к V2 способствуют иерархической обработке зрительных стимулов. Нейроны V2 основываются на базовых функциях, обнаруженных в V1, извлекая более сложные визуальные атрибуты, такие как текстура, глубина и цвет. Эта иерархическая обработка важна для построения более тонкого и детального представления визуальной сцены.
Более того, реципрокные обратные связи от V2 к V1 играют значительную роль в модуляции активности нейронов V1. Считается, что эта петля обратной связи участвует в таких процессах, как внимание, группировка восприятия и разделение фигуры и фона. Динамическое взаимодействие между V1 и V2 подчеркивает сложную природу обработки информации в зрительной системе.
Более того, связи V2 с последующими зрительными областями, включая V3, V4 и V5, способствуют формированию распределенной сети визуальной обработки. Эти связи позволяют интегрировать различные визуальные особенности, такие как движение и форма, на нескольких этапах визуальной иерархии. [36]
С точки зрения анатомии V2 разделен на четыре квадранта: дорсальное и вентральное представительство в левом и правом полушариях . Вместе эти четыре региона составляют полную карту визуального мира. V2 имеет много общих свойств с V1: клетки настроены на простые свойства, такие как ориентация, пространственная частота и цвет. Реакции многих нейронов V2 также модулируются более сложными свойствами, такими как ориентация иллюзорных контуров , [37] [38] бинокулярное неравенство , [39] и является ли стимул частью фигуры или фона. [40] [41] Недавние исследования показали, что клетки V2 демонстрируют небольшую модуляцию внимания (больше, чем V1, меньше, чем V4), настроены на умеренно сложные паттерны и могут управляться множественными ориентациями в разных субрегионах в пределах одного рецептивного поля.
Утверждается, что весь вентральный зрительно-гиппокампальный поток важен для зрительной памяти. [42] Эта теория, в отличие от доминирующей, предсказывает, что изменения памяти распознавания объектов (ORM) могут быть результатом манипуляций в V2, области, которая тесно взаимосвязана внутри вентрального потока зрительной коры. В мозгу обезьяны эта область получает сильные прямые связи от первичной зрительной коры (V1) и посылает сильные проекции на другие вторичные зрительные коры (V3, V4 и V5). [43] [44] Большинство нейронов этой области у приматов настроены на простые зрительные характеристики, такие как ориентация, пространственная частота, размер, цвет и форма. [38] [45] [46] Анатомические исследования показывают, что слой 3 области V2 участвует в обработке визуальной информации. В отличие от слоя 3, слой 6 зрительной коры состоит из многих типов нейронов, и их реакция на зрительные стимулы более сложна.
В одном исследовании было обнаружено, что клетки слоя 6 коры V2 играют очень важную роль в хранении памяти распознавания объектов, а также в преобразовании кратковременных воспоминаний об объектах в долговременные воспоминания. [47]
Третья зрительная кора, включая область V3.
[ редактировать ]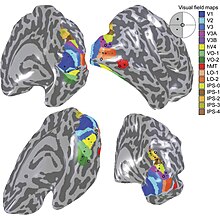
Термин «третий зрительный комплекс» относится к области коры головного мозга, расположенной непосредственно перед V2, которая включает в себя область, называемую у человека зрительной областью V3 . «Сложная» номенклатура оправдана тем фактом, что до сих пор существуют некоторые разногласия относительно точной протяженности области V3: некоторые исследователи предполагают, что кора, расположенная перед V2, может включать два или три функциональных подразделения. Например, Дэвид Ван Эссен и другие (1986) предположили существование «дорсального V3» в верхней части полушария головного мозга, который отличается от «вентрального V3» (или вентральной задней области, VP), расположенного в нижняя часть мозга. Дорсальный и вентральный V3 имеют четкие связи с другими частями мозга, выглядят по-разному на срезах, окрашенных разными методами, и содержат нейроны, реагирующие на разные комбинации зрительных стимулов (например, цветоселективные нейроны чаще встречаются в вентральной части мозга). В3). У людей также сообщалось о дополнительных подразделениях, включая V3A и V3B. Эти подразделения расположены вблизи дорсального V3, но не примыкают к V2.
Дорсальный V3 обычно считается частью дорсального потока, получающего входные сигналы от V2 и первичной зрительной области и проецирующегося на заднюю теменную кору . Анатомически он может располагаться в области Бродмана 19 . Брэддик с помощью фМРТ предположил, что область V3/V3A может играть роль в обработке глобального движения. [48] Другие исследования предпочитают рассматривать дорсальный V3 как часть более крупной области, называемой дорсомедиальной областью (DM), которая содержит представление всего поля зрения. Нейроны в области DM реагируют на когерентное движение крупных паттернов, охватывающих обширные участки зрительного поля (Lui et al., 2006).
Вентральный V3 (VP) имеет гораздо более слабые связи с первичной зрительной областью и более сильные связи с нижней височной корой . В то время как более ранние исследования предполагали, что ВП содержит представительство только верхней части зрительного поля (выше точки фиксации), более поздние работы показывают, что эта область более обширна, чем предполагалось ранее, и, как и другие зрительные области, она может содержать полную визуальное представление. Пересмотренная, более обширная VP Роза и Твидейл называют вентролатеральную заднюю область (VLP). [49]
V4
[ редактировать ]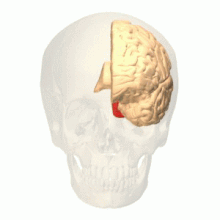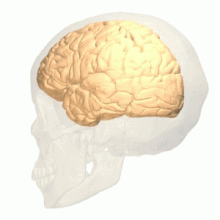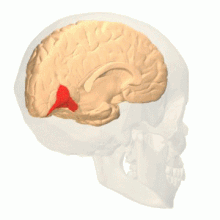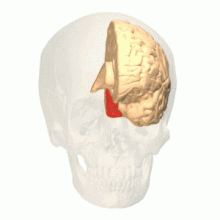

Зрительная область V4 — одна из зрительных областей экстрастриарной зрительной коры. У макак он расположен кпереди от V2 и кзади от задней нижневисочной области (PIT) . Он включает как минимум четыре области (левый и правый V4d, левый и правый V4v), а некоторые группы сообщают, что он также содержит ростральные и каудальные подразделения. Неизвестно, является ли человеческий V4 таким же обширным, как у гомолога макаки , который является предметом споров. [50]
V4 является третьей кортикальной областью вентрального потока , получающей сильные сигналы прямой связи от V2 и посылающей сильные связи с PIT . Он также получает прямой ввод от V1, особенно для центрального пространства. Кроме того, он имеет более слабые связи с V5 и дорсальной предлунной извилиной (DP).
V4 — первая область вентрального потока, демонстрирующая сильную модуляцию внимания. Большинство исследований показывают, что избирательное внимание может изменить скорость стрельбы в V4 примерно на 20%. Фундаментальная статья Морана и Дезимоне, описывающая эти эффекты, стала первой статьей, обнаружившей эффекты внимания в любом месте зрительной коры. [51]
Как и V2, V4 настроен на ориентацию, пространственную частоту и цвет. В отличие от V2, V4 настроен на особенности объектов средней сложности, такие как простые геометрические фигуры, хотя полного параметрического описания пространства настройки для V4 никто не разработал. Зрительная область V4 не настроена на сложные объекты, такие как лица, в отличие от областей нижневисочной коры .
Огневые свойства Фау-4 впервые описал Семир Зеки в конце 1970-х годов, который также дал название местности. До этого V4 был известен по анатомическому описанию — предлунная извилина . Первоначально Зеки утверждал, что целью V4 была обработка информации о цвете. Работы начала 1980-х годов доказали, что V4 так же непосредственно участвует в распознавании формы, как и более ранние области коры. [ нужна ссылка ] Это исследование подтвердило гипотезу двух потоков , впервые выдвинутую Унгерлейдером и Мишкиным в 1982 году.
Недавние исследования показали, что V4 демонстрирует долговременную пластичность. [52] кодирует значимость стимула, контролируется сигналами, поступающими из лобных полей глаза , [53] и с вниманием демонстрирует изменения пространственного профиля своих рецептивных полей. [ нужна ссылка ] Кроме того, недавно было показано, что активация области V4 у человека (область V4h) наблюдается при восприятии и удержании цвета предметов, но не их формы. [54] [55]
Средневисочная зрительная зона (V5)
[ редактировать ]Средневисочная зрительная область ( MT или V5 ) представляет собой область экстрастриарной зрительной коры. У некоторых видов обезьян как Нового Света , так и обезьян Старого Света область МТ содержит высокую концентрацию нейронов, избирательных по направлению. [56] Считается, что МТ у приматов играет важную роль в восприятии движения , интеграции локальных сигналов движения в глобальное восприятие и управлении некоторыми движениями глаз . [56]
Соединения
[ редактировать ]МТ связан с широким спектром корковых и подкорковых областей мозга. Его вход поступает из зрительных областей коры V1, V2 и дорсальной области V3 ( дорсомедиальная область ). [57] [58] кониоцеллюлярные области LGN , [59] и нижний пульвинар . [60] Характер проекций на МТ несколько меняется при представлении фовеальных и периферических полей зрения, при этом последние получают входные данные из областей, расположенных в средней линии коры и ретросплениальной области . [61]
Стандартная точка зрения состоит в том, что V1 предоставляет «самые важные» входные данные для MT. [56] Тем не менее, несколько исследований показали, что нейроны МТ способны реагировать на визуальную информацию, часто избирательно по направлению, даже после того, как V1 был разрушен или инактивирован. [62] Более того, исследования Семира Зеки и его коллег показали, что определенные типы визуальной информации могут достигать MT еще до того, как она достигнет V1.
MT отправляет свой основной сигнал в области коры головного мозга, непосредственно окружающие его, включая области FST, MST и V4t (средний височный серп). Другие проекции МТ нацелены на области лобной и теменной долей, связанные с движением глаз (лобное поле глаза и латеральная внутритеменная область).
Функция
[ редактировать ]Первые исследования электрофизиологических свойств нейронов МТ показали, что значительная часть клеток настроена на скорость и направление движущихся зрительных стимулов. [63] [64]
Исследования повреждений также подтвердили роль МТ в восприятии движения и движениях глаз. [65] Нейропсихологические исследования пациента, неспособного видеть движение и вместо этого видящего мир в серии статичных «кадров», показали, что V5 у приматов гомологичен MT у человека. [66] [67]
Однако, поскольку нейроны в V1 также настроены на направление и скорость движения, эти ранние результаты оставили открытым вопрос о том, что именно MT может делать, а что не может V1. В этой области была проведена большая работа, поскольку она, по-видимому, интегрирует локальные визуальные сигналы движения в глобальное движение сложных объектов. [68] Например, поражение V5 приводит к нарушению восприятия движения и обработки сложных стимулов. Он содержит множество нейронов, избирательных к движению сложных зрительных объектов (концы линий, углы). Микростимуляция нейрона, расположенного в V5, влияет на восприятие движения. Например, если найти в V5 обезьяны нейрон, предпочитающий движение вверх, и стимулировать его электродом, то обезьяна с большей вероятностью сообщит о движении вверх, когда ей предъявлены стимулы, содержащие также слова «лево» и «право». как «восходящие» компоненты. [69]
До сих пор существует много споров по поводу точной формы вычислений, проводимых в области МТ. [70] и некоторые исследования показывают, что движение функций на самом деле уже доступно на более низких уровнях зрительной системы, таких как V1. [71] [72]
Функциональная организация
[ редактировать ]Было показано, что МТ организован в виде колонн направления. [73] ДеАнджелис утверждал, что МТ-нейроны также были организованы на основе их настройки на бинокулярное неравенство. [74]
V6
[ редактировать ]Дорсомедиальная область (DM), также известная как V6 , по-видимому, реагирует на зрительные стимулы, связанные с самодвижением. [75] и широкоугольная стимуляция. [76] V6 — подразделение зрительной коры приматов, впервые описанное Джоном Оллманом и Джоном Каасом в 1975 году. [77] V6 расположен в дорсальной части экстрастриарной коры , рядом с глубокой бороздой, проходящей через центр мозга ( медиальная продольная щель ), и обычно также включает в себя части медиальной коры, такие как теменно-затылочная борозда (POS). [76] : 7970 ДМ содержит топографически организованное представление всего поля зрения. [76] : 7970
Есть сходство между зрительной областью V5 и V6 обыкновенной игрунки . Обе области получают прямые связи от первичной зрительной коры . [76] : 7971 Оба имеют высокое содержание миелина — характеристику, которая обычно присутствует в структурах мозга, участвующих в быстрой передаче информации. [78]
В течение многих лет считалось, что СД существует только у обезьян Нового Света . Однако более поздние исследования показали, что СД также существует у обезьян Старого Света и людей. [76] : 7972 V6 также иногда называют теменно-затылочной областью (ПО), хотя соответствие неточно. [79] [80]
Характеристики
[ редактировать ]Нейроны области DM/V6 ночных мартышек и обыкновенных мартышек обладают уникальными ответными свойствами, включая чрезвычайно острую избирательность ориентации зрительных контуров и предпочтение длинным непрерывным линиям, охватывающим большие части поля зрения. [81] [82]
Однако по сравнению с областью МТ значительно меньшая доля клеток СД проявляет избирательность по направлению движения зрительных образов. [83] Еще одно заметное отличие области MT заключается в том, что клетки в DM настроены на низкочастотные компоненты изображения и плохо реагируют на движение текстурированных узоров, таких как поле случайных точек. [83] Эти свойства ответа позволяют предположить, что ДМ и МТ могут работать параллельно: первый анализирует собственное движение относительно окружающей среды, а второй анализирует движение отдельных объектов относительно фона. [83]
Недавно у человека была идентифицирована область, реагирующая на широкоугольные поля потока, и считается, что она является гомологом области V6 макака. [84]
Пути
[ редактировать ]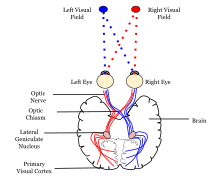
Связи и свойства ответа клеток в DM/V6 позволяют предположить, что эта область является ключевым узлом в подмножестве « дорсального потока », называемого некоторыми как «дорсомедиальный путь». [85] Этот путь, вероятно, важен для контроля скелетно-моторной активности, включая постуральные реакции и движения к объектам. [80] Основная «прямая» связь СД происходит с корой, непосредственно ростральной по отношению к нему, на границе между затылочной и теменной долями (V6A). [85] Эта область, в свою очередь, имеет относительно прямые связи с областями лобной доли, контролирующими движения рук, включая премоторную кору . [85] [86]
См. также
[ редактировать ]- Кортикальная область
- Корковая слепота
- Теория интеграции функций
- Список регионов человеческого мозга
- Ретинотопия
- Визуальная обработка
- Сложная ячейка
Ссылки
[ редактировать ]- ^ Мэзер Г. «Зрительная кора» . Школа естественных наук: Университет Сассекса . Проверено 6 марта 2017 г.
- ^ Фишек М., Херрманн Д., Эгеа-Вайс А., Кловс М., Бауэр Л., Ли Т.И. и др. (май 2023 г.). «Кортико-кортикальная обратная связь задействует активные дендриты зрительной коры» . Природа . 617 (7962): 769–776. Бибкод : 2023Natur.617..769F . дои : 10.1038/s41586-023-06007-6 . ПМЦ 10244179 . ПМИД 37138089 .
- ^ Бенсон Н.К., Юн Дж.М., Форенцо Д., Энгель С.А., Кей К.Н., Винавер Дж. (ноябрь 2022 г.). «Изменчивость площади поверхности карт V1, V2 и V3 у большой выборки людей-наблюдателей» . Журнал неврологии . 42 (46): 8629–8646. doi : 10.1523/jneurosci.0690-21.2022 . ПМЦ 9671582 . ПМИД 36180226 .
- ^ Браз Дж., Петтре Дж., Ричард П., Керрен А., Линсен Л., Баттиато С. и др. (11 февраля 2016 г.). «Алгоритмическая оптимизация модели HMAX, направленная на эффективное распознавание объектов» . В Битар А.В., Мансур М.М., Чехаб А. (ред.). Компьютерное зрение, обработка изображений и компьютерная графика, теория и приложения . Берлин, Германия: Springer . п. 377. ИСБН 978-3-319-29971-6 .
- ^ Унгерлейдер Л.Г., Мишкин М (1982). «Две корковые зрительные системы» . Ingle DJ, Гудейл М.А., Мэнсфилд Р.Дж. (ред.). Анализ визуального поведения . Бостон: MIT Press. стр. 549–586 . ISBN 978-0-262-09022-3 .
- ^ Гудейл М.А., Милнер А.Д. (январь 1992 г.). «Отдельные визуальные пути восприятия и действия». Тенденции в нейронауках . 15 (1): 20–25. CiteSeerX 10.1.1.207.6873 . дои : 10.1016/0166-2236(92)90344-8 . ПМИД 1374953 . S2CID 793980 .
- ^ Аглиоти С., ДеСуза Дж. Ф., Гудейл М. А. (июнь 1995 г.). «Иллюзия контраста размеров обманывает глаз, но не руку» . Современная биология . 5 (6): 679–685. Бибкод : 1995CBio....5..679A . дои : 10.1016/S0960-9822(95)00133-3 . ПМИД 7552179 . S2CID 206111613 .
- ^ Франц В.Х., Шарновски Ф., Гегенфуртнер К.Р. (декабрь 2005 г.). «Эффект иллюзии при хватании постоянен во времени, а не динамичен». Журнал экспериментальной психологии. Человеческое восприятие и производительность . 31 (6): 1359–1378. дои : 10.1037/0096-1523.31.6.1359 . ПМИД 16366795 .
- ^ Ганель Т., Гудейл Массачусетс (декабрь 2003 г.). «Визуальный контроль действия, а не восприятия, требует аналитической обработки формы объекта». Природа . 426 (6967): 664–667. Бибкод : 2003Natur.426..664G . дои : 10.1038/nature02156 . PMID 14668865 . S2CID 4314969 .
- ^ Ганель Т., Танзер М., Гудейл М.А. (март 2008 г.). «Двойная диссоциация между действием и восприятием в контексте зрительных иллюзий: противоположные эффекты реального и иллюзорного размера». Психологическая наука . 19 (3): 221–225. дои : 10.1111/j.1467-9280.2008.02071.x . ПМИД 18315792 . S2CID 15679825 .
- ^ Гудейл, Массачусетс (июль 2011 г.). «Превращение видения в действие» . Исследование зрения . 51 (13): 1567–1587. дои : 10.1016/j.visres.2010.07.027 . ПМИД 20691202 .
- ^ Коэн П., Сит Т.П., Уэллс М.Дж., Карандини М., Харрис К.Д. (август 2023 г.). «Лобная кора головного мозга мыши принимает аддитивные мультисенсорные решения» . Нейрон . 111 (15): 2432–2447.e13. дои : 10.1016/j.neuron.2023.05.008 . ПМЦ 10957398 . ПМИД 37295419 .
- ^ Гликштейн М., Риццолатти Дж. (1 декабря 1984 г.). «Франческо Дженнари и строение коры головного мозга». Тенденции в нейронауках . 7 (12): 464–467. дои : 10.1016/S0166-2236(84)80255-6 . S2CID 53168851 .
- ^ Хьюбель Д.Х., Визель Т.Н. (декабрь 1972 г.). «Ламинарное и столбчатое распределение коленно-кортикальных волокон у макак». Журнал сравнительной неврологии . 146 (4): 421–450. дои : 10.1002/cne.901460402 . ПМИД 4117368 . S2CID 6478458 .
- ^ Леуба Г., Крафцик Р. (октябрь 1994 г.). «Изменения объема, оценки поверхности, трехмерной формы и общего количества нейронов первичной зрительной коры человека от середины беременности до старости». Анатомия и эмбриология . 190 (4): 351–366. дои : 10.1007/BF00187293 . ПМИД 7840422 . S2CID 28320951 .
- ^ У Ф, Лу Ц, Конг Ю, Чжан Цз (сентябрь 2023 г.). «Всесторонний обзор роли нарушения функции зрительной коры при депрессивных расстройствах: возможности и проблемы». Неврологический бюллетень . 39 (9): 1426–1438. дои : 10.1007/s12264-023-01052-7 . PMC 10062279. PMID 36995569 .
- ^ Jump up to: а б Иоганн Кеплер (1604 г.) От Паралипомены до Витело, посредством которого рассматривается оптическая часть астрономии (Ad Vitellionem Paralipomena, quibus astronomae pars optica traditvr, 1604), цитируется А. Марком Смитом (2015) «От зрения к свету». Кеплер смоделировал глаз как стеклянную сферу, наполненную водой, и обнаружил, что каждая точка сцены, воспринимаемая глазом, проецируется на точку на задней стороне глаза (сетчатке).
- ^ Баргут Л. (1999). О различиях между маскировкой периферических и фовеальных паттернов (магистры). Беркли, Калифорния: Калифорнийский университет, Беркли.
- ^ Баргут Л. (2003). Видение: как глобальный контекст восприятия меняет обработку локального контраста (докторская диссертация). Ученая пресса. ISBN 978-3-639-70962-9 . Обновлено, чтобы включить методы компьютерного зрения.
- ^ Хюпе Дж.М., Джеймс А.С., Пейн Б.Р., Ломбер С.Г., Жирар П., Буллиер Дж. (август 1998 г.). «Корковая обратная связь улучшает различение фигуры и фона с помощью нейронов V1, V2 и V3». Природа . 394 (6695): 784–7. Бибкод : 1998Natur.394..784H . дои : 10.1038/29537 . ПМИД 9723617 .
- ^ Анджелуччи А., Буллер Дж. (2003). «Выход за пределы классического рецептивного поля нейронов V1: горизонтальные аксоны или аксоны обратной связи?». Журнал физиологии, Париж . 97 (2–3): 141–54. doi : 10.1016/j.jphysparis.2003.09.001 . ПМИД 14766139 .
- ^ Буллиер Дж., Хюпе Ж.М., Джеймс А.С., Жирар П. (2001). «Глава 13. Роль обратных связей в формировании ответов нейронов зрительной коры». Роль обратных связей в формировании ответов нейронов зрительной коры . Прогресс в исследованиях мозга. Том. 134. стр. 193–204. дои : 10.1016/s0079-6123(01)34014-1 . ISBN 978-0-444-50586-6 . ПМИД 11702544 .
- ^ Мюррей С.О., Шратер П., Керстен Д. (2004). «Перцептивная группировка и взаимодействие между зрительными областями коры». Нейронные сети: Официальный журнал Международного общества нейронных сетей . 17 (5–6): 695–705. дои : 10.1016/j.neunet.2004.03.010 . ПМИД 15288893 .
- ^ Хуан Цзюй, Ван С, Дреер Б (март 2007 г.). «Влияние обратимой инактивации задне-височной зрительной коры на активность нейронов в области 17 кошки». Исследования мозга . 1138 : 111–28. дои : 10.1016/j.brainres.2006.12.081 . ПМИД 17276420 .
- ^ Уильямс М.А., Бейкер С.И., Оп де Бек Х.П., Шим В.М., Данг С., Триантафиллу С. и др. (декабрь 2008 г.). «Обратная связь информации о зрительных объектах с фовеальной ретинотопической корой» . Природная неврология . 11 (12): 1439–45. дои : 10.1038/nn.2218 . ПМЦ 2789292 . ПМИД 18978780 .
- ^ Кессервани Х (октябрь 2020 г.). «Биофизика визуального обнаружения границ: обзор основных принципов» . Куреус . 12 (10): е11218. дои : 10.7759/cureus.11218 . ПМК 7706146 . ПМИД 33269147 .
- ^ Линдеберг Т. (декабрь 2013 г.). «Вычислительная теория зрительных рецептивных полей» . Биологическая кибернетика . 107 (6): 589–635. дои : 10.1007/s00422-013-0569-z . ПМК 3840297 . ПМИД 24197240 .
- ^ Jump up to: а б Линдеберг Т. (январь 2021 г.). «Нормативная теория зрительных рецептивных полей» . Гелион . 7 (1): e05897. Бибкод : 2021Heliy...705897L . дои : 10.1016/j.heliyon.2021.e05897 . ПМЦ 7820928 . ПМИД 33521348 .
- ^ Jump up to: а б Линдеберг Т. (2023). «Ковариационные свойства при естественных преобразованиях изображений для обобщенной модели производной Гаусса для зрительных рецептивных полей» . Границы вычислительной нейронауки . 17 : 1189949. дои : 10.3389/fncom.2023.1189949 . ПМЦ 10311448 . ПМИД 37398936 .
- ^ ДеАнджелис Г.К., Озава И., Фриман Р.Д. (октябрь 1995 г.). «Динамика рецептивных полей в центральных зрительных путях». Тенденции в нейронауках . 18 (10): 451–458. дои : 10.1016/0166-2236(95)94496-р . ПМИД 8545912 . S2CID 12827601 .
- ^ ДеАнджелис Г.К., Анзай А. (21 ноября 2003 г.). «Современный взгляд на классическое рецептивное поле: линейная и нелинейная пространственно-временная обработка нейронами V1» . В Чалупа Л.М., Вернер Дж.С. (ред.). Визуальные нейронауки, 2 т. Набор . Том. 1. Кембридж: Массачусетский технологический институт. стр. 704–719. дои : 10.7551/mitpress/7131.003.0052 . ISBN 978-0-262-27012-0 .
- ^ Линдеберг Т. (2013). «Инвариантность зрительных операций на уровне рецептивных полей» . ПЛОС ОДИН . 8 (7): e66990. arXiv : 1210.0754 . Бибкод : 2013PLoSO...866990L . дои : 10.1371/journal.pone.0066990 . ПМК 3716821 . ПМИД 23894283 .
- ^ Чжаопин Л. (2014). «Гипотеза V1 - создание восходящей карты значимости для предварительного внимательного отбора и сегментации». Понимание видения: теория, модели и данные . Издательство Оксфордского университета. стр. 189–314. doi : 10.1093/acprof:oso/9780199564668.003.0005 . ISBN 9780199564668 .
- ^ Шварцкопф Д.С., Сонг С., Рис Дж. (январь 2011 г.). «Площадь поверхности человеческого V1 предсказывает субъективное восприятие размера объекта» . Природная неврология . 14 (1): 28–30. дои : 10.1038/nn.2706 . ПМК 3012031 . ПМИД 21131954 .
- ^ Газзанига М.С., Иври Р.Б., Мангун Г.Р. (2002). Когнитивная нейронаука: биология разума (2-е изд.). WW Norton & Co Inc. ISBN 978-0-393-97777-6 .
- ^ Тейлор, Кэтрин. и Жанетт Родригес. «Визуальная дискриминация». StatPearls, StatPearls Publishing, 19 сентября 2022 г.
- ^ фон дер Хейдт Р., Петеранс Э., Баумгартнер Г. (июнь 1984 г.). «Иллюзорные контуры и реакции корковых нейронов». Наука . 224 (4654): 1260–1262. Бибкод : 1984Sci...224.1260V . дои : 10.1126/science.6539501 . ПМИД 6539501 .
- ^ Jump up to: а б Анзай А., Пэн X, Ван Эссен, округ Колумбия (октябрь 2007 г.). «Нейроны зрительной области V2 обезьяны кодируют комбинации ориентаций». Природная неврология . 10 (10): 1313–1321. дои : 10.1038/nn1975 . ПМИД 17873872 . S2CID 6796448 .
- ^ фон дер Хейдт Р., Чжоу Х., Фридман Х.С. (2000). «Представление стереоскопических границ в зрительной коре обезьяны» . Исследование зрения . 40 (15): 1955–1967. дои : 10.1016/s0042-6989(00)00044-4 . ПМИД 10828464 . S2CID 10269181 .
- ^ Цю FT, фон дер Хейдт Р. (июль 2005 г.). «Фигура и фон в зрительной коре: v2 сочетает в себе стереоскопические сигналы с правилами гештальта» . Нейрон . 47 (1): 155–166. дои : 10.1016/j.neuron.2005.05.028 . ПМК 1564069 . ПМИД 15996555 .
- ^ Маруко И., Чжан Б., Тао X, Тонг Дж., Смит Э.Л., Чино Ю.М. (ноябрь 2008 г.). «Постнатальное развитие несоответствия чувствительности в зрительной области 2 (v2) макак» . Журнал нейрофизиологии . 100 (5): 2486–2495. дои : 10.1152/jn.90397.2008 . ПМЦ 2585398 . ПМИД 18753321 .
- ^ Басси Т.Дж., Саксида Л.М. (2007). «Память, восприятие и вентральный зрительно-периринально-гиппокампальный поток: нестандартное мышление». Гиппокамп . 17 (9): 898–908. дои : 10.1002/hipo.20320 . ПМИД 17636546 . S2CID 13271331 .
- ^ Степневска I, Каас Дж. Х. (июль 1996 г.). «Топографические закономерности корковых связей V2 у макак». Журнал сравнительной неврологии . 371 (1): 129–152. doi : 10.1002/(SICI)1096-9861(19960715)371:1<129::AID-CNE8>3.0.CO;2-5 . ПМИД 8835723 . S2CID 8500842 .
- ^ Гаттасс Р., Соуза А.П., Мишкин М., Унгерлейдер Л.Г. (март 1997 г.). «Корковые проекции области V2 у макаки» . Кора головного мозга . 7 (2): 110–129. дои : 10.1093/cercor/7.2.110 . ПМИД 9087820 .
- ^ Хегде Дж., Ван Эссен, округ Колумбия (март 2000 г.). «Избирательность сложных форм в зрительной зоне V2 приматов» . Журнал неврологии . 20 (5): РК61. doi : 10.1523/JNEUROSCI.20-05-j0001.2000 . ПМК 6772904 . ПМИД 10684908 .
- ^ Хегде Дж., Ван Эссен, округ Колумбия (ноябрь 2004 г.). «Временная динамика анализа формы в зрительной области V2 макак». Журнал нейрофизиологии . 92 (5): 3030–3042. дои : 10.1152/jn.00822.2003 . ПМИД 15201315 . S2CID 6428310 .
- ^ Лопес-Аранда М.Ф., Лопес-Теллес Х.Ф., Наварро-Лобато И, Масмуди-Мартин М., Гутьеррес А., Хан ЗУ (июль 2009 г.). «Роль 6-го слоя зрительной коры V2 в памяти распознавания объектов». Наука . 325 (5936): 87–89. Бибкод : 2009Sci...325...87L . дои : 10.1126/science.1170869 . ПМИД 19574389 . S2CID 23990759 .
- ^ Брэддик О.Дж., О'Брайен Дж.М., Уоттам-Белл Дж., Аткинсон Дж., Хартли Т., Тернер Р. (2001). «Области мозга, чувствительные к связному зрительному движению». Восприятие . 30 (1): 61–72. дои : 10.1068/p3048 . ПМИД 11257978 . S2CID 24081674 .
- ^ Роза М.Г., Твидейл Р. (июль 2000 г.). «Зрительные области в латеральной и вентральной экстрастриарной коре мартышки». Журнал сравнительной неврологии . 422 (4): 621–651. doi : 10.1002/1096-9861(20000710)422:4<621::AID-CNE10>3.0.CO;2-E . ПМИД 10861530 . S2CID 25982910 .
- ^ Годдард Э., Мэннион DJ, Макдональд Дж.С., Соломон С.Г., Клиффорд К.В. (апрель 2011 г.). «Восприимчивость к цвету противоречит дорсальному компоненту человеческого V4» . Журнал видения . 11 (4): 3. дои : 10.1167/11.4.3 . ПМИД 21467155 .
- ^ Моран Дж., Дезимона Р. (август 1985 г.). «Избирательное внимание регулирует визуальную обработку в экстрастриарной коре». Наука . 229 (4715): 782–784. Бибкод : 1985Sci...229..782M . CiteSeerX 10.1.1.308.6038 . дои : 10.1126/science.4023713 . ПМИД 4023713 .
- ^ Шмид М.К., Шмидт Дж.Т., Петерс А.Дж., Сондерс Р.К., Майер А., Леопольд Д.А. (ноябрь 2013 г.). «Чувствительные к движению реакции в зрительной области V4 при отсутствии первичной зрительной коры» . Журнал неврологии . 33 (48): 18740–18745. doi : 10.1523/JNEUROSCI.3923-13.2013 . ПМЦ 3841445 . ПМИД 24285880 .
- ^ Мур Т. , Армстронг К.М. (январь 2003 г.). «Селективное пропускание зрительных сигналов путем микростимуляции лобной коры». Природа . 421 (6921): 370–373. Бибкод : 2003Natur.421..370M . дои : 10.1038/nature01341 . ПМИД 12540901 . S2CID 4405385 .
- ^ Козловский С, Рогачев А (2021). «Как взаимодействуют области вентрального зрительного потока, когда мы запоминаем информацию о цвете и форме». В Величковский Б.М., Балабан П.М., Ушаков В.Л. (ред.). Достижения в области когнитивных исследований, искусственного интеллекта и нейроинформатики. Интеркогници 2020 . Достижения в области интеллектуальных систем и вычислений. Том. 1358. Чам: Международное издательство Springer. стр. 95–100. дои : 10.1007/978-3-030-71637-0_10 . ISBN 978-3-030-71636-3 .
- ^ Козловский С., Рогачев А. (октябрь 2021 г.). «Вентральные области зрительной коры и обработка цвета и формы в зрительной рабочей памяти». Международный журнал психофизиологии . 168 (Дополнение): С155–С156. doi : 10.1016/j.ijpsycho.2021.07.437 . S2CID 239648133 .
- ^ Jump up to: а б с Родился RT, Брэдли, округ Колумбия (2005). «Структура и функция зрительной области МТ». Ежегодный обзор неврологии . 28 : 157–189. дои : 10.1146/annurev.neuro.26.041002.131052 . ПМИД 16022593 .
- ^ Феллеман DJ, Ван Эссен, округ Колумбия (1991). «Распределенная иерархическая обработка данных в коре головного мозга приматов» . Кора головного мозга . 1 (1): 1–47. дои : 10.1093/cercor/1.1.1-a . ПМИД 1822724 .
- ^ Унгерлейдер Л.Г., Дезимона Р. (июнь 1986 г.). «Корковые связи зрительной области МТ у макаки». Журнал сравнительной неврологии . 248 (2): 190–222. дои : 10.1002/cne.902480204 . ПМИД 3722458 . S2CID 1876622 .
- ^ Синчич Л.К., Парк К.Ф., Вольгемут М.Дж., Хортон Дж.К. (октябрь 2004 г.). «Обход V1: прямой коленчатый вход в область МТ». Природная неврология . 7 (10): 1123–1128. дои : 10.1038/nn1318 . ПМИД 15378066 . S2CID 13419990 .
- ^ Уорнер CE, Голдшмит Ю., Борн Дж.А. (2010). «Синапс афферентов сетчатки с релейными клетками, нацеленными на среднюю височную область в пульвинарном и латеральном коленчатых ядрах» . Границы нейроанатомии . 4 :8. дои : 10.3389/нейро.05.008.2010 . ПМК 2826187 . ПМИД 20179789 .
- ^ Палмер С.М., Роза М.Г. (октябрь 2006 г.). «Четкая анатомическая сеть областей коры для анализа движения дальним периферическим зрением». Европейский журнал неврологии . 24 (8): 2389–2405. дои : 10.1111/j.1460-9568.2006.05113.x . ПМИД 17042793 . S2CID 21562682 .
- ^ Родман Х.Р., Гросс К.Г., Олбрайт Т.Д. (июнь 1989 г.). «Афферентные основы свойств зрительных реакций в области МТ макаки. I. Эффекты удаления стриарной коры» . Журнал неврологии . 9 (6): 2033–2050. doi : 10.1523/JNEUROSCI.09-06-02033.1989 . ПМК 6569731 . ПМИД 2723765 .
- ^ Дубнер Р., Зеки С.М. (декабрь 1971 г.). «Свойства ответа и рецептивные поля клеток в анатомически определенной области верхней височной борозды у обезьяны». Исследования мозга . 35 (2): 528–532. дои : 10.1016/0006-8993(71)90494-X . ПМИД 5002708 . .
- ^ Маунселл Дж. Х., Ван Эссен, округ Колумбия (май 1983 г.). «Функциональные свойства нейронов средневисочной зрительной области обезьяны-макаки. I. Избирательность по направлению, скорости и ориентации стимула». Журнал нейрофизиологии . 49 (5): 1127–1147. дои : 10.1152/jn.1983.49.5.1127 . ПМИД 6864242 . S2CID 8708245 .
- ^ Дюрстелер М.Р., Вурц Р.Х., Ньюсом В.Т. (май 1987 г.). «Нарушение направленного преследования после поражения фовеального представительства в верхней височной борозде макаки». Журнал нейрофизиологии . 57 (5): 1262–1287. CiteSeerX 10.1.1.375.8659 . дои : 10.1152/jn.1987.57.5.1262 . ПМИД 3585468 .
- ^ Хесс Р.Х., Бейкер К.Л., Зиль Дж. (май 1989 г.). «Пациент, слепой к движению: пространственные и временные фильтры низкого уровня» . Журнал неврологии . 9 (5): 1628–1640. doi : 10.1523/JNEUROSCI.09-05-01628.1989 . ПМК 6569833 . ПМИД 2723744 .
- ^ Бейкер К.Л., Хесс Р.Ф., Зиль Дж. (февраль 1991 г.). «Остаточное восприятие движения у пациента, «слепого к движению», оцененное с помощью случайных точечных стимулов с ограниченным сроком службы» . Журнал неврологии . 11 (2): 454–461. doi : 10.1523/JNEUROSCI.11-02-00454.1991 . ПМК 6575225 . ПМИД 1992012 .
- ^ Мовшон, Дж. А., Адельсон, Э. Х., Гиззи, М. С., и Ньюсом, WT (1985). Анализ движущихся зрительных образов. В: К. Чагас, Р. Гаттасс и К. Гросс (ред.), Механизмы распознавания образов (стр. 117–151), Рим: Vatican Press.
- ^ Бриттен К.Х., ван Везель Р.Дж. (май 1998 г.). «Электрическая микростимуляция кортикальной области MST искажает восприятие направления у обезьян». Природная неврология . 1 (1): 59–63. дои : 10.1038/259 . ПМИД 10195110 . S2CID 52820462 .
- ^ Уилсон Х.Р., вице-президент Феррера, Йо К. (июль 1992 г.). «Психофизически мотивированная модель двумерного восприятия движения». Визуальная нейронаука . 9 (1): 79–97. дои : 10.1017/s0952523800006386 . ПМИД 1633129 . S2CID 45196189 .
- ^ Тинсли С.Дж., Уэбб Б.С., Барракла Н.Э., Винсент С.Дж., Паркер А., Деррингтон А.М. (август 2003 г.). «Природа нейронных ответов V1 на двумерные движущиеся модели зависит от структуры рецептивного поля мартышки». Журнал нейрофизиологии . 90 (2): 930–937. дои : 10.1152/jn.00708.2002 . ПМИД 12711710 . S2CID 540146 .
- ^ Pack CC, родился RT, Ливингстон MS (февраль 2003 г.). «Двумерная субструктура стерео- и двигательных взаимодействий в зрительной коре макак» . Нейрон . 37 (3): 525–535. дои : 10.1016/s0896-6273(02)01187-x . ПМИД 12575958 .
- ^ Олбрайт Т.Д. (декабрь 1984 г.). «Направление и ориентационная избирательность нейронов зрительной области МТ макаки». Журнал нейрофизиологии . 52 (6): 1106–1130. дои : 10.1152/jn.1984.52.6.1106 . ПМИД 6520628 .
- ^ ДеАнджелис Г.К., Ньюсом WT (февраль 1999 г.). «Организация диспаратно-селективных нейронов в области МТ макак» . Журнал неврологии . 19 (4): 1398–1415. doi : 10.1523/JNEUROSCI.19-04-01398.1999 . ПМК 6786027 . ПМИД 9952417 .
- ^ Кардин В., Смит А.Т. (август 2010 г.). «Чувствительность зрительных и вестибулярных областей коры человека к визуальной стимуляции, совместимой с эгомоциями» . Кора головного мозга . 20 (8): 1964–1973. дои : 10.1093/cercor/bhp268 . ПМК 2901022 . ПМИД 20034998 .
- ^ Jump up to: а б с д и Питцалис С., Галлетти С., Хуанг Р.С., Патрия Ф., Коммиттери Г., Галати Г. и др. (июль 2006 г.). «Широкоугольная ретинотопия определяет кортикальную зрительную область v6 человека» . Журнал неврологии . 26 (30): 7962–7973. doi : 10.1523/jneurosci.0178-06.2006 . ПМК 6674231 . ПМИД 16870741 .
- ^ Оллман Дж. М., Каас Дж. Х. (декабрь 1975 г.). «Дорсомедиальная кортикальная зрительная область: область третьего уровня в затылочной доле совы-обезьяны (Aotus trivirgatus)». Исследования мозга . 100 (3): 473–487. дои : 10.1016/0006-8993(75)90153-5 . ПМИД 811327 . S2CID 22980932 .
- ^ Питцалис С., Фаттори П., Галлетти С. (2013). «Функциональная роль медиальной области движения V6» . Границы поведенческой нейронауки . 6 : 91. дои : 10.3389/fnbeh.2012.00091 . ПМЦ 3546310 . ПМИД 23335889 .
- ^ Галлетти С., Гамберини М., Куц Д.Ф., Балдинотти И., Фаттори П. (февраль 2005 г.). «Взаимосвязь между V6 и PO в экстрастриарной коре макак» (PDF) . Европейский журнал неврологии . 21 (4): 959–970. CiteSeerX 10.1.1.508.5602 . дои : 10.1111/j.1460-9568.2005.03911.x . ПМИД 15787702 . S2CID 15020868 . Архивировано из оригинала (PDF) 8 августа 2017 г. Проверено 14 сентября 2018 г.
- ^ Jump up to: а б Галлетти С., Куц Д.Ф., Гамберини М., Бревельери Р., Фаттори П. (ноябрь 2003 г.). «Роль медиальной теменно-затылочной коры в контроле тянущихся и хватательных движений». Экспериментальное исследование мозга . 153 (2): 158–170. дои : 10.1007/s00221-003-1589-z . ПМИД 14517595 . S2CID 1821863 .
- ^ Бейкер Дж. Ф., Петерсен С. Е., Ньюсом В. Т., Оллман Дж. М. (март 1981 г.). «Свойства зрительной реакции нейронов в четырех экстрастриарных зрительных областях совы-обезьяны (Aotus trivirgatus): количественное сравнение медиальной, дорсомедиальной, дорсолатеральной и средней височной областей». Журнал нейрофизиологии . 45 (3): 397–416. дои : 10.1152/jn.1981.45.3.397 . ПМИД 7218008 . S2CID 9865958 .
- ^ Луи Л.Л., Борн Дж.А., Роза М.Г. (февраль 2006 г.). «Свойства функционального ответа нейронов дорсомедиальной зрительной области обезьян Нового Света (Callithrix jacchus)» . Кора головного мозга . 16 (2): 162–177. дои : 10.1093/cercor/bhi094 . ПМИД 15858163 .
- ^ Jump up to: а б с Дюкре Д. «Калькариновая (зрительная) кора» . База знаний Коннектопедии . Архивировано из оригинала 20 января 2018 г. Проверено 25 января 2018 г.
{{cite web}}: CS1 maint: неподходящий URL ( ссылка ) - ^ Питцалис С., Серено М.И., Коммиттери Дж., Фаттори П., Галати Г., Патрия Ф. и др. (февраль 2010 г.). «Человек v6: область медиального движения» . Кора головного мозга . 20 (2): 411–424. дои : 10.1093/cercor/bhp112 . ПМЦ 2803738 . ПМИД 19502476 .
- ^ Jump up to: а б с Фаттори П., Раос В., Бревельери Р., Боско А., Марцокки Н., Галлетти К. (январь 2010 г.). «Дорсомедиальный путь предназначен не только для достижения: захватывания нейронов медиальной теменно-затылочной коры макак» . Журнал неврологии . 30 (1): 342–349. doi : 10.1523/JNEUROSCI.3800-09.2010 . ПМК 6632536 . ПМИД 20053915 .
- ^ Монако С., Малфатти Г., Зендрон А., Пелленсен Е., Турелла Л. (декабрь 2019 г.). «Прогнозирующее кодирование намерений действий в дорсальном и вентральном зрительном потоке основано на зрительных ожиданиях, информации, основанной на памяти, и моторной подготовке». Структура и функции мозга . 224 (9): 3291–3308. дои : 10.1007/s00429-019-01970-1 . ПМИД 31673774 . S2CID 207811473 .
Внешние ссылки
[ редактировать ]- «Первичная зрительная кора Мэтью Шмолески» . Университет Юты . Архивировано из оригинала 29 декабря 2004 г.
- «Архитектура зрительной коры Дэвида Хьюбела» . Гарвардский университет . Архивировано из оригинала 01 марта 2017 г.
- ancil-415 в NeuroNames - полосатая область 17
- ancil-699 в NeuroNames - зона Бродмана 17 в Геноне
- Окрашенные изображения срезов мозга, включающие «визуальную кору%20», в проекте BrainMaps.
- Симулятор компьютерного моделирования карт зрительной коры на сайте topographica.org