Устойчивость растений к болезням


Устойчивость растений к болезням защищает растения от патогенов двумя способами: с помощью заранее сформированных структур и химических веществ, а также с помощью реакций иммунной системы, вызванных инфекцией. По отношению к восприимчивому растению устойчивость к болезням представляет собой снижение роста патогенов на растении или внутри него (и, следовательно, снижение заболеваемости), в то время как термин «толерантность к болезням» описывает растения, которые проявляют незначительный ущерб от болезней, несмотря на значительные уровни патогенов. Исход заболевания определяется трехсторонним взаимодействием возбудителя, растения и условий окружающей среды (взаимодействие, известное как треугольник болезни ).
Соединения, активирующие защиту, могут перемещаться от клетки к клетке и систематически через сосудистую систему растения. Однако у растений нет циркулирующих иммунных клеток , поэтому большинство типов клеток обладают широким набором антимикробной защиты. Хотя при сравнении нескольких образцов можно наблюдать очевидные качественные различия в устойчивости к болезням (что позволяет классифицировать их как «устойчивые» или «восприимчивые» после заражения одним и тем же штаммом патогена при одинаковых уровнях инокулята в аналогичных средах), градация количественных различий в устойчивости к болезням чаще наблюдается между штаммами или генотипами растений . Растения устойчиво сопротивляются одним патогенам, но уступают другим; устойчивость обычно специфична для определенных видов или штаммов патогенов.
Фон
[ редактировать ]Устойчивость растений к болезням имеет решающее значение для надежного производства продуктов питания и обеспечивает значительное сокращение использования земли, воды, топлива и других ресурсов в сельском хозяйстве. Растения как в естественных, так и в культурных популяциях обладают присущей им устойчивостью к болезням, но это не всегда их защищает.
Фитофтороз, Великий голод в Ирландии 1840-х годов, был вызван оомицетом Phytophthora infestans . Первый в мире массово выращиваемый бананов сорт Грос Мишель был потерян в 1920-х годах из-за панамской болезни, вызванной грибком Fusarium oxysporum . Нынешние эпидемии стеблевой ржавчины пшеницы , листовой ржавчины и желтой полосатой ржавчины, распространяющиеся из Восточной Африки на Индийский субконтинент, вызваны ржавчинными грибами Puccinia graminis и P. striiformis . Другие эпидемии включают гниль каштана , а также рецидивирующие тяжелые заболевания растений, такие как рисовая гниль , соевая нематода и язва цитрусовых . [ 1 ] [ 2 ]
Патогены растений могут быстро распространяться на большие расстояния, переносясь водой, ветром, насекомыми и людьми. По оценкам, в крупных регионах и многих видах сельскохозяйственных культур болезни обычно снижают урожайность растений на 10% каждый год в более развитых странах или сельскохозяйственных системах, но потери урожая из-за болезней часто превышают 20% в менее развитых странах. [ 1 ]
Тем не менее, борьба с болезнями достаточно успешна для большинства сельскохозяйственных культур. Борьба с болезнями достигается за счет использования растений, которые были выведены на хорошую устойчивость ко многим заболеваниям, а также с помощью таких подходов к выращиванию растений, как севооборот , семена, свободные от патогенов, подходящие даты посева и плотность посадки, контроль влажности полей и использование пестицидов .
Общие механизмы устойчивости к болезням
[ редактировать ]Предварительно сформированные структуры и соединения
[ редактировать ]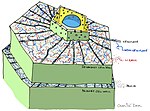
- растения Кутикула /поверхность
- растительных клеток Стенки
- Антимикробные химические вещества (например: полифенолы , сесквитерпеновые лактоны , сапонины )
- Антимикробные пептиды
- Ингибиторы ферментов
- Детоксифицирующие ферменты, расщепляющие токсины патогенного происхождения.
- Рецепторы, которые воспринимают присутствие патогенов и активируют индуцируемую защиту растений. [ 3 ]
Индуцируемая постинфекционная защита растений
[ редактировать ]- клеточной стенки Укрепление ( целлюлоза , лигнин , суберин , каллоза , белки клеточной стенки) [ 4 ]
- Антимикробные химические вещества, включая активные формы кислорода, такие как перекись водорода или пероксинитрит , или более сложные фитоалексины, такие как генистеин или камалексин.
- Антимикробные белки, такие как дефенсины , тионины или PR-1.
- Антимикробные ферменты, такие как хитиназы , бета- глюканазы или пероксидазы. [ 4 ]
- Гиперчувствительная реакция – быстрая реакция гибели клеток-хозяев, связанная с индукцией защиты.
Иммунная система
[ редактировать ]Иммунная система растений несет в себе два взаимосвязанных уровня рецепторов: один наиболее часто воспринимает молекулы вне клетки, а другой наиболее часто воспринимает молекулы внутри клетки. Обе системы распознают злоумышленника и реагируют активацией антимикробной защиты в инфицированной и соседних клетках. В некоторых случаях сигналы активации защиты распространяются на остальную часть растения или даже на соседние растения. Обе системы обнаруживают различные типы молекул патогенов и классы белков-рецепторов растений. [ 5 ] [ 6 ]
Первый уровень в первую очередь регулируется рецепторами распознавания образов , которые активируются путем распознавания эволюционно консервативных патогенов или молекулярных паттернов, связанных с микробами (PAMP или MAMP). Активация PRRs приводит к внутриклеточной передаче сигналов, транскрипционному перепрограммированию и биосинтезу сложного выходного ответа, который ограничивает колонизацию. Эта система известна как иммунитет, запускаемый PAMP, или иммунитет, запускаемый шаблоном (PTI). [ 7 ] [ 6 ] [ 8 ]
Второй уровень, в первую очередь регулируемый продуктами R-гена , часто называют эффекторно-запускаемым иммунитетом (ETI). ETI обычно активируется присутствием специфических «эффекторов» патогена, а затем вызывает сильный антимикробный ответ (см. раздел «Ген R» ниже).
Помимо PTI и ETI, защитные силы растений могут быть активированы путем обнаружения соединений, связанных с повреждением (DAMP), таких как части клеточной стенки растения, высвобождаемые во время патогенной инфекции. [ 9 ]
Реакции, активируемые рецепторами PTI и ETI, включают открытие ионных каналов , окислительный взрыв , окислительно-восстановительные изменения в клетках или каскады протеинкиназ , которые непосредственно активируют клеточные изменения (например, усиление клеточной стенки или выработку антимикробных препаратов) или активируют изменения в экспрессии генов , которые затем усиливают другие защитные функции. ответы.
Иммунные системы растений демонстрируют некоторое механическое сходство с иммунными системами насекомых и млекопитающих, но также обладают многими специфичными для растений характеристиками. [ 10 ] Два вышеописанных уровня играют центральную роль в иммунитете растений, но не полностью описывают иммунную систему растений. Кроме того, многие конкретные примеры явного PTI или ETI нарушают общепринятые определения PTI/ETI, что указывает на необходимость расширения определений и/или парадигм. [ 11 ]
Термин «количественная устойчивость» (обсуждаемый ниже) относится к устойчивости растений к болезням, которая контролируется множеством генов и множеством молекулярных механизмов, каждый из которых оказывает небольшое влияние на общий признак устойчивости. Количественную устойчивость часто противопоставляют устойчивости к ETI, опосредованной отдельными R-генами с основным эффектом.
Иммунитет, вызванный шаблонами
[ редактировать ]PAMP патогенов , консервативные молекулы, которые обитают в нескольких родах , многие исследователи называют MAMP. Защиты, вызванной восприятием MAMP, достаточны для отражения большинства патогенов. Однако эффекторные белки патогена (см. ниже) адаптированы для подавления базальной защиты, такой как PTI. Было обнаружено множество рецепторов MAMP (и DAMP). MAMP и DAMP часто обнаруживаются с помощью трансмембранных рецептор-киназ, которые несут LRR или внеклеточные домены LysM . [ 5 ]
Эффекторный иммунитет
[ редактировать ]Иммунитет, запускаемый эффекторами (ETI), активируется при наличии эффекторов патогена. Реакция ETI зависит от R-генов и активируется конкретными штаммами патогенов. Растительный ETI часто вызывает апоптозную гиперчувствительную реакцию .
R-гены и R-белки
[ редактировать ]Растения развили R-гены (гены устойчивости), продукты которых опосредуют устойчивость к определенным штаммам вирусов, бактерий, оомицетов, грибов, нематод или насекомых. Продукты гена R представляют собой белки, которые позволяют распознавать эффекторы конкретного патогена либо посредством прямого связывания, либо путем распознавания эффекторного изменения белка-хозяина. [ 6 ] Многие R-гены кодируют белки NB-LRR (белки с нуклеотидсвязывающими и богатыми лейцином повторяющимися доменами, также известные как белки NLR или белки STAND, среди других названий). Большинство иммунных систем растений несут репертуар из 100–600 различных гомологов гена R. Было продемонстрировано, что отдельные R-гены опосредуют устойчивость к конкретным штаммам вирусов, бактерий, оомицетов, грибов, нематод или насекомых. Продукты гена R контролируют широкий набор реакций устойчивости к болезням, индукции которых часто бывает достаточно, чтобы остановить дальнейший рост/распространение патогена.
Изученные R-гены обычно придают специфичность определенным штаммам вида патогена (тех, которые экспрессируют известный эффектор). Как впервые заметил Гарольд Флор в своей формулировке взаимосвязи ген-ген в середине 20-го века , ген R растения обладает специфичностью к гену авирулентности патогена (ген Avr). Теперь известно, что гены авирулентности кодируют эффекторы. Ген патогена Avr должен иметь такую же специфичность, что и ген R, чтобы этот ген R обеспечивал устойчивость, что позволяет предположить взаимодействие рецептора/ лиганда для генов Avr и R. [ 10 ] Альтернативно, эффектор может модифицировать свою клеточную мишень-хозяина (или молекулярную приманку этой мишени), а продукт гена R (белок NLR) активирует защитные силы, когда он обнаруживает модифицированную форму мишени-хозяина или приманки. [ 6 ] [ 12 ]
Эффекторная биология
[ редактировать ]Эффекторы играют центральную роль в патогенном или симбиотическом потенциале микробов и микроскопических животных, колонизирующих растения, таких как нематоды. [ 13 ] [ 14 ] [ 15 ] Эффекторы обычно представляют собой белки, которые доставляются за пределы микроба в клетку-хозяина. Эти эффекторы, полученные от колонистов, манипулируют физиологией и развитием клеток хозяина. Таким образом, эффекторы представляют собой примеры совместной эволюции (пример: грибной белок, который функционирует вне гриба, но внутри растительных клеток, эволюционировал и взял на себя функции, специфичные для растения). Круг хозяев возбудителя определяется, среди прочего, наличием соответствующих эффекторов, позволяющих колонизировать конкретного хозяина. [ 5 ] Эффекторы патогенного происхождения являются мощным инструментом для определения функций растений, которые играют ключевую роль в заболеваниях и устойчивости к болезням. Очевидно, что большинство эффекторов манипулируют физиологией хозяина, позволяя заболеванию возникнуть. Хорошо изученные бактериальные патогены растений обычно экспрессируют несколько десятков эффекторов, часто доставляемых в организм хозяина с помощью секреторного аппарата типа III . [ 13 ] Фитопатогены грибов, оомицетов и нематод, по-видимому, экспрессируют несколько сотен эффекторов. [ 14 ] [ 15 ]
Так называемые «основные» эффекторы определяются с функциональной точки зрения по их широкому распространению в популяции конкретного патогена и их существенному вкладу в вирулентность патогена. Геномика может использоваться для идентификации основных эффекторов, которые затем можно использовать для открытия новых аллелей гена R , которые можно использовать в селекции растений на устойчивость к болезням.
Малые РНК и РНК-интерференция
[ редактировать ]растений Предполагается, что пути мРНК являются важными компонентами иммунитета, запускаемого патоген-ассоциированным молекулярным паттерном (PAMP) (PTI) и эффекторного иммунитета (ETI). [ 16 ] [ 17 ] бактериально-индуцированные микроРНК (миРНК) у арабидопсиса влияют на передачу гормональных сигналов, включая ауксин, абсцизовую кислоту (АБК), жасмоновую кислоту (ЖК) и салициловую кислоту (СА). Было показано, что [ 18 ] [ 19 ] Достижения в полногеномных исследованиях выявили массовую адаптацию паттернов экспрессии микроРНК хозяина после заражения грибковыми патогенами Fusarium virguliforme . [ 20 ] трава Эризифовая [ 21 ] Вертициллиум георгин , [ 22 ] и Кронартии дубы , [ 23 ] и оомицета Phytophthora sojae . [ 24 ] Изменения экспрессии мРНК в ответ на грибковые патогены указывают на то, что в этом защитном пути может участвовать подавление генов. Однако есть также свидетельства того, что противогрибковая защитная реакция на Colletotrichum spp. Инфекция кукурузы не полностью регулируется индукцией специфической микроРНК, но вместо этого может действовать для точной настройки баланса между генетическими и метаболическими компонентами после заражения. [ нужна ссылка ]
Транспорту мРНК во время инфекции, вероятно, способствуют внеклеточные везикулы (EV) и мультивезикулярные тельца (MVB). [ 25 ] Состав РНК в растительных ЭВ полностью не изучен, но вполне вероятно, что они частично ответственны за транспортировку РНК. Растения могут системно переносить вирусные РНК, мРНК , микроРНК и малые интерферирующие РНК (миРНК) через флоэму. [ 26 ] Считается, что этот процесс происходит через плазмодесмы и включает РНК-связывающие белки, которые способствуют локализации РНК в клетках мезофилла. Хотя они были идентифицированы во флоэме с мРНК, нет убедительных доказательств того, что они опосредуют транспорт РНК на большие расстояния. [ 27 ] Таким образом, ЭВ могут способствовать альтернативному пути загрузки РНК во флоэму или, возможно, могут транспортировать РНК через апопласт. [ 28 ] Есть также доказательства того, что растительные ЭВ могут обеспечивать межвидовой перенос мРНК посредством РНК-интерференции, такой как индуцированное хозяином подавление генов (HIGS). [ 29 ] [ 30 ] Транспорт РНК между растениями и грибами, по-видимому, является двунаправленным, поскольку мРНК грибкового патогена Botrytis cinerea нацелены на гены защиты хозяина у арабидопсиса и томатов. было показано, что [ 31 ]
Устойчивость на видовом уровне
[ редактировать ]В небольшом числе случаев гены растений эффективны против целого вида патогена, даже если этот вид патогенен для других генотипов этого вида-хозяина. Примеры включают ячмень MLO против мучнистой росы , пшеницу Lr34 против листовой ржавчины и пшеницу Yr36 против полосатой ржавчины пшеницы . В зависимости от конкретного гена и комбинации растительного патогена может существовать множество механизмов этого типа устойчивости. Другие причины эффективного иммунитета растений могут включать отсутствие коадаптации (у патогена и/или растения отсутствуют многочисленные механизмы, необходимые для колонизации и роста внутри этого вида-хозяина) или особенно эффективный набор заранее сформированных защитных механизмов. [ нужна ссылка ]
Сигнальные механизмы
[ редактировать ]Восприятие присутствия патогена
[ редактировать ]Передача защитных сигналов растений активируется рецепторами, обнаруживающими патогены, которые описаны в разделе выше. [ 5 ] Активированные рецепторы часто вызывают выработку реактивного кислорода и оксида азота , кальция , калия и протонов поток ионов , изменение уровней салициловой кислоты и других гормонов, а также активацию MAP-киназ и других специфических протеинкиназ . [ 10 ] Эти события, в свою очередь, обычно приводят к модификации белков, которые контролируют транскрипцию генов , и активации экспрессии генов , связанных с защитой . [ 8 ]
Транскрипционные факторы и гормональный ответ
[ редактировать ]Были идентифицированы многочисленные гены и/или белки, а также другие молекулы, которые опосредуют передачу защитного сигнала растений. [ 32 ] [ 33 ] Динамика цитоскелета и движения пузырьков помогает направить защитные реакции растений на точку атаки патогена.
Механизмы транскрипционных факторов и гормонов
[ редактировать ]Активность иммунной системы растений частично регулируется сигнальными гормонами, такими как: [ 34 ] [ 35 ]
Между этими путями могут существовать существенные перекрестные помехи. [ 34 ]
Регулирование путем деградации
[ редактировать ]Как и во многих путях передачи сигнала, экспрессия растительных генов во время иммунных ответов может регулироваться путем деградации. Это часто происходит, когда связывание гормона с гормональными рецепторами стимулирует убиквитин -ассоциированную деградацию белков-репрессоров, которые блокируют экспрессию определенных генов. Конечным результатом является активируемая гормонами экспрессия генов. Примеры: [ 36 ]
- Ауксин : связывается с рецепторами, которые затем рекрутируют и разрушают репрессоры активаторов транскрипции, которые стимулируют экспрессию специфичных для ауксина генов.
- Жасмоновая кислота: аналогична ауксину, за исключением того, что жасмонатные рецепторы влияют на сигнальные медиаторы жасмонатного ответа, такие как белки JAZ.
- Гиббереллиновая кислота : Гиббереллин вызывает конформационные изменения рецепторов, а также связывание и деградацию белков Della .
- Этилен: Ингибирующее фосфорилирование активатора этиленового ответа EIN2 блокируется связыванием этилена. Когда это фосфорилирование снижается, белок EIN2 расщепляется, и часть белка перемещается в ядро, чтобы активировать экспрессию гена, отвечающего на этилен.
Убиквитин и передача сигналов E3
[ редактировать ]Убиквитинирование играет центральную роль в передаче сигналов в клетках, которая регулирует процессы, включая деградацию белка и иммунологический ответ. [ 37 ] Хотя одной из основных функций убиквитина является нацеливание на белки для разрушения, он также полезен в сигнальных путях, высвобождении гормонов, апоптозе и перемещении материалов по клетке. Убиквитинирование является компонентом нескольких иммунных реакций. Без правильного функционирования убиквитина вторжение патогенов и других вредных молекул резко увеличится из-за ослабления иммунной защиты. [ 37 ]

Сигнализация E3
[ редактировать ]Фермент Е3 убиквитинлигаза является основным компонентом, обеспечивающим специфичность путей деградации белков, включая пути передачи иммунных сигналов. [ 36 ] Компоненты фермента E3 можно сгруппировать по тому, какие домены они содержат, и включают несколько типов. [ 38 ]
К ним относятся одиночная субъединица Ring и U-box, HECT и CRL . [ 39 ] [ 40 ] Сигнальные пути растений, включая иммунные реакции, контролируются несколькими путями обратной связи, которые часто включают отрицательную обратную связь; и они могут регулироваться ферментами деубиквитинирования, деградацией факторов транскрипции и деградацией негативных регуляторов транскрипции. [ 36 ] [ 41 ]
Количественное сопротивление
[ редактировать ]Различия в устойчивости растений к болезням часто носят постепенный или количественный, а не качественный характер. Термин количественная устойчивость (QR) относится к устойчивости растений к болезням, которая контролируется множеством генов и множеством молекулярных механизмов, каждый из которых оказывает небольшое или незначительное влияние на общий признак устойчивости. [ 42 ] QR важен в селекции растений, поскольку получаемая в результате устойчивость часто оказывается более устойчивой (эффективной в течение большего количества лет) и с большей вероятностью будет эффективной против большинства или всех штаммов конкретного патогена. QR обычно эффективен против одного вида патогенов или группы близкородственных видов, а не в целом эффективен против нескольких патогенов. [ 42 ] QR часто получают путем селекции растений без знания причинно-следственных генетических локусов или молекулярных механизмов. QR, вероятно, зависит от многих компонентов иммунной системы растений, обсуждаемых в этой статье, а также от характеристик, которые являются уникальными для определенных пар растение-патоген (например, чувствительность к определенным эффекторам патогена), а также от общих характеристик растений, таких как листовая пластинка. характеристики поверхности, корневая система или архитектура растительного покрова. Термин QR является синонимом незначительной генной устойчивости . [ 43 ]
Устойчивость взрослых растений и устойчивость рассады
[ редактировать ]Устойчивость взрослых растений (APR) — это специальный термин, обозначающий количественную устойчивость, которая не эффективна на стадии рассады, но эффективна на многих оставшихся стадиях роста растений. [ 43 ] [ 44 ] [ 42 ] Разница между устойчивостью взрослых растений и устойчивостью проростков особенно важна для однолетних культур . [ 45 ] Устойчивость проростков – это устойчивость, которая начинается на стадии развития растения и продолжается на протяжении всей его жизни. В использовании специалистами этот термин не относится к устойчивости, которая активна только на стадии всходов. «Устойчивость проростков» является синонимом устойчивости основного гена или устойчивости на всех стадиях (ASR) и используется в отличие от «устойчивости взрослых растений». [ 43 ] Устойчивость сеянцев часто опосредуется отдельными R-генами, но не все R-гены кодируют устойчивость сеянцев.
Селекция растений на устойчивость к болезням
[ редактировать ]Селекционеры уделяют особое внимание отбору и созданию устойчивых к болезням линий растений. Болезни растений также можно частично контролировать с помощью пестицидов и таких методов выращивания, как севооборот , обработка почвы, плотность посадки, отсутствие болезней семян и очистка оборудования, но обычно предпочтительны сорта растений с присущей (генетически детерминированной) устойчивостью к болезням. [ 2 ] Селекция на устойчивость к болезням началась, когда растения были впервые одомашнены. Усилия по селекции продолжаются, поскольку популяции патогенов находятся под давлением отбора и приобретают повышенную вирулентность, патогены перемещаются (или перемещаются) в новые области, изменение методов выращивания или климата благоприятствует некоторым патогенам и может снизить эффективность устойчивости, а селекция растений по другим признакам может подорвать предыдущую устойчивость. . [ 46 ] Линия растений с приемлемой устойчивостью к одному патогену может не иметь устойчивости к другим.
Селекция на устойчивость обычно включает в себя:
- Идентификация растений, которые могут быть менее желательны по другим причинам, но которые обладают полезным признаком устойчивости к болезням, включая дикие линии растений, которые часто проявляют повышенную устойчивость.
- Скрещивание желательного, но восприимчивого к болезням сорта с растением, являющимся источником устойчивости.
- Выращивание кандидатов для разведения в условиях, способствующих заболеванию, возможно, включая инокуляцию патогена. Необходимо уделять внимание конкретным изолятам патогена, чтобы учитывать изменчивость внутри одного вида патогена.
- Отбор устойчивых к болезням особей, которые сохраняют другие желательные характеристики, такие как урожайность, качество, а также другие признаки устойчивости к болезням. [ 46 ]
Устойчивость называется устойчивой , если она продолжает оставаться эффективной в течение нескольких лет широкого использования по мере развития популяций патогенов. « Вертикальная устойчивость » специфична для определенных рас или штаммов видов патогенов, часто контролируется отдельными R-генами и может быть менее устойчивой. Горизонтальная устойчивость или устойчивость широкого спектра против целого вида патогена часто не полностью эффективна, но более устойчива и часто контролируется многими генами, которые разделяются в размножающихся популяциях. [ 2 ] Устойчивость устойчивости важна даже тогда, когда ожидается появление будущих улучшенных сортов: среднее время от признания человеком угрозы нового грибкового заболевания до появления устойчивого к этому патогену урожая составляет не менее двенадцати лет. [ 47 ] [ 48 ]
Такие культуры, как картофель, яблоко, банан и сахарный тростник, часто размножаются путем вегетативного размножения , чтобы сохранить весьма желательные сорта растений, поскольку ауткроссинг этих видов серьезно нарушает предпочтительные признаки. См. также бесполое размножение . Вегетативно размножаемые культуры могут быть одной из лучших мишеней для повышения устойчивости с помощью биотехнологического растений метода трансформации для управления генами, влияющими на устойчивость к болезням. [ 1 ]
Научная селекция устойчивости к болезням началась сэром Роулендом Биффеном , который идентифицировал единственный рецессивный ген устойчивости пшеницы к желтой ржавчине. Почти каждая культура затем была выведена с включением генов устойчивости к болезням (R), многие из которых были интрогрессированы от совместимых диких родственников. [ 1 ]
ГМ или трансгенно-инженерная устойчивость к болезням
[ редактировать ]Термин ГМ ( «генетически модифицированные» ) часто используется как синоним слова « трансгенный» для обозначения растений, модифицированных с использованием технологий рекомбинантной ДНК. Растения с устойчивостью к трансгенным/ГМ-болезням против насекомых-вредителей оказались чрезвычайно успешными в качестве коммерческой продукции, особенно кукурузы и хлопка, и ежегодно высаживаются на площади более 20 миллионов гектаров в более чем 20 странах мира. [ 49 ] (см. также генетически модифицированные культуры ). Устойчивость трансгенных растений к болезням против микробных патогенов была впервые продемонстрирована в 1986 году. Экспрессия оболочки вируса генных последовательностей белков обеспечивала устойчивость к вирусам посредством малых РНК . Это оказалось широко применимым механизмом ингибирования репликации вируса. [ 50 ] Объединив гены белков оболочки трех разных вирусов, ученые разработали гибриды тыквы с подтвержденной на практике мультивирусной устойчивостью. Подобный уровень устойчивости к этой разновидности вирусов не был достигнут с помощью традиционной селекции.
Аналогичная стратегия была использована для борьбы с вирусом кольцевой пятнистости папайи , который к 1994 году угрожал уничтожить Гавайях на индустрию папайи . Полевые испытания продемонстрировали отличную эффективность и высокое качество плодов. К 1998 году первая трансгенная папайя, устойчивая к вирусу, была одобрена для продажи. Устойчивость к болезням сохраняется более 15 лет. На трансгенную папайю приходится около 85% производства на Гавайях. Плод одобрен для продажи в США, Канаде и Японии.
Линии картофеля, экспрессирующие последовательности вирусных репликаз, которые придают устойчивость к вирусу скручивания листьев картофеля, продавались под торговыми названиями NewLeaf Y и NewLeaf Plus и широко использовались в коммерческом производстве в 1999–2001 годах, пока корпорация McDonald's не решила не закупать ГМ-картофель, а компания Monsanto не решила. закрыть свой картофельный бизнес NatureMark. [ 51 ] Картофель NewLeaf Y и NewLeaf Plus обладал двумя ГМ-признаками, поскольку они также выражали Bt-опосредованную устойчивость к колорадскому жуку.
К 2013 году на рынок не попала ни одна другая культура с искусственно созданной устойчивостью к болезням против микробных патогенов, хотя более десятка из них находились в той или иной стадии разработки и тестирования.
| Год публикации | Обрезать | Устойчивость к болезням | Механизм | Статус разработки |
|---|---|---|---|---|
| 2012 | Помидор | Бактериальное пятно | Ген R перца | 8 лет полевых испытаний |
| 2012 | Рис | Бактериальный ожог и бактериальная полоса | Разработанный ген E | Лаборатория |
| 2012 | Пшеница | Мучнистая роса | Сверхэкспрессированный ген R пшеницы | 2 года полевых испытаний на момент публикации |
| 2011 | Яблоко | парши яблони Грибок | тионина Ген из ячменя | 4 года полевых испытаний на момент публикации |
| 2011 | Картофель | Картофельный вирус Y | Устойчивость, обусловленная патогенами | 1 год полевых испытаний на момент публикации |
| 2010 | Яблоко | Огненный ожог | Антибактериальный протеин от моли | 12 лет полевых испытаний на момент публикации |
| 2010 | Помидор | Мультибактериальная резистентность | PRR из арабидопсиса | Лабораторный масштаб |
| 2010 | Банан | Ксантомонада увядает | Новый ген перца | Сейчас на полевых испытаниях |
| 2009 | Картофель | Фитофтороз | R-гены диких родственников | 3 года полевых испытаний |
| 2009 | Картофель | Фитофтороз | Ген R от дикого родственника | 2 года полевых испытаний на момент публикации |
| 2008 | Картофель | Фитофтороз | Ген R от дикого родственника | 2 года полевых испытаний на момент публикации |
| 2008 | слива | Вирус оспы сливы | Устойчивость, обусловленная патогенами | Одобрения регулирующих органов, без коммерческих продаж |
| 2005 | Рис | Бактериальная полоса | Ген R кукурузы | Лаборатория |
| 2002 | Ячмень | Стеблевая ржавчина | Ген киназы покоящихся лимфоцитов (RLK) устойчивого сорта ячменя | Лаборатория |
| 1997 | Папайя | Вирус кольцевой пятнистости | Устойчивость, обусловленная патогенами | Одобрено и продается на коммерческой основе с 1998 г., в Японии продается с 2012 г. |
| 1995 | Давить | Три мозаичных вируса | Устойчивость, обусловленная патогенами | Одобрено и продается на коммерческой основе с 1994 года. |
| 1993 | Картофель | Картофельный вирус X | Фермент, индуцируемый интерфероном млекопитающих | 3 года полевых испытаний на момент публикации |
передача ПРР
[ редактировать ]Исследования, направленные на искусственное сопротивление, основаны на нескольких стратегиях. Один из них — передать полезные PRR видам, у которых их нет. Идентификация функциональных PRR и их передача видам-реципиентам, у которых отсутствует ортологичный рецептор, может обеспечить общий путь к дополнительному расширению репертуара PRR. Например, Arabidopsis PRR рецептор EF-Tu (EFR) распознает бактериальный фактор элонгации трансляции EF-Tu . Исследования, проведенные в лаборатории Сейнсбери, показали, что внедрение EFR в Nicotiana benthamiana или Solanum lycopersicum (помидор), которые не могут распознавать EF-Tu , обеспечивает устойчивость к широкому спектру бактериальных патогенов. Экспрессия EFR в томатах была особенно эффективна против широко распространенной и разрушительной почвенной бактерии Ralstonia solanacearum . [ 52 ] ) томата И наоборот, ген PRR Verticillium 1 ( Ve1 может быть перенесен от томата к Arabidopsis , где он придает устойчивость к изолятам Verticillium расы 1 . [ 1 ]
Укладка
[ редактировать ]Вторая стратегия пытается одновременно использовать несколько генов NLR — стратегия разведения, известная как стекирование. с помощью ДНК Сорта, созданные с помощью молекулярной селекции или переноса генов , вероятно, будут демонстрировать более длительную устойчивость, поскольку патогенам придется мутировать несколько эффекторных генов. Секвенирование ДНК позволяет исследователям функционально «добывать» гены NLR из нескольких видов/штаммов. [ 1 ]
Эффекторный ген avrBs2 из Xanthomona perforans является возбудителем бактериальной пятнистости перца и томата. Первый «эффекторно-рационализированный» поиск потенциально устойчивого R-гена последовал за открытием того, что avrBs2 обнаружен у большинства болезнетворных видов Xanthomonas и необходим для приспособленности патогена. Ген Bs2 NLR из дикого перца Capsicum chacoense был перенесен в томат, где он ингибировал рост патогена. Полевые испытания продемонстрировали высокую устойчивость без бактерицидных химикатов. Однако редкие штаммы Xanthomonas преодолели Bs2 -опосредованную устойчивость перца за счет приобретения мутаций avrBs2 , которые избегают распознавания, но сохраняют вирулентность. Объединение R-генов, каждый из которых распознает свой основной эффектор, может задержать или предотвратить адаптацию. [ 1 ]
Более 50 локусов в штаммах пшеницы придают устойчивость к возбудителям стеблевой, листовой и желтой полосатой ржавчины пшеницы. Ген NLR стеблевой ржавчины 35 ( Sr35 ), клонированный из диплоидного родственника культивируемой пшеницы, Triticum monococcum , обеспечивает устойчивость к изоляту пшеничной ржавчины Ug99 . Аналогичным образом, Sr33 из родственника пшеницы Aegilops tauschii кодирует ортолог пшеницы ячменя Mla, гены устойчивости к мучнистой росе. Оба гена необычны для пшеницы и ее родственников. В сочетании с геном Sr2 , который действует аддитивно, по крайней мере, с Sr33, они могут обеспечить длительную устойчивость к болезням к Ug99 и его производным. [ 1 ]
Гены-исполнители
[ редактировать ]Другой класс генов устойчивости растений к болезням открывает «люк», который быстро убивает проникшие клетки, останавливая размножение патогенов. Xanthomonas и Ralstonia Эффекторы, подобные активатору транскрипции (TAL), представляют собой ДНК-связывающие белки, которые активируют экспрессию генов хозяина для повышения вирулентности патогена. Линии риса и перца независимо развили сайты связывания эффекторов TAL, которые вместо этого действуют как палач, вызывающий гибель сверхчувствительных клеток-хозяев при повышении регуляции. Xa27 из риса и Bs3 и Bs4c из перца являются такими генами-«исполнителями» (или «палачами»), которые кодируют негомологичные растительные белки с неизвестной функцией. Гены-исполнители экспрессируются только в присутствии специфического эффектора TAL. [ 1 ]
Сконструированные гены-исполнители были продемонстрированы путем успешной переработки промотора Bs3 перца , чтобы он содержал два дополнительных сайта связывания для эффекторов TAL из разных штаммов патогенов. Впоследствии сконструированный ген-исполнитель был внедрен в рис путем добавления пяти эффекторных сайтов связывания TAL к промотору Xa27 . Синтетическая конструкция Xa27 обеспечивала устойчивость к бактериальному ожогу Xanthomonas и бактериальным видам полосатости листьев. [ 1 ]
Аллели восприимчивости хозяина
[ редактировать ]Большинство патогенов растений перепрограммируют модели экспрессии генов хозяина, чтобы принести прямую пользу патогену. Перепрограммированные гены, необходимые для выживания и размножения патогенов, можно назвать «генами предрасположенности к болезням». Рецессивные гены устойчивости являются кандидатами на восприимчивость к болезням. Например, мутация отключила ген Arabidopsis, кодирующий пектатлиазу (участвующую в деградации клеточной стенки), что обеспечило устойчивость к возбудителю мучнистой росы Golovinomyces cichoracearum . ячменя Аналогичным образом, ген MLO гороха и томата и спонтанно мутированные ортологи MLO также придают устойчивость к мучнистой росе. [ 1 ]
Lr34 — ген, обеспечивающий частичную устойчивость пшеницы к листовой, желтой ржавчине и мучнистой росе. Lr34 кодирует транспортер аденозинтрифосфатной (АТФ)-связывающей кассеты (ABC). Доминантный аллель, обеспечивающий устойчивость к болезням, недавно был обнаружен у культурной пшеницы (не у диких сортов) и, как и MLO , обеспечивает устойчивость широкого спектра у ячменя. [ 1 ]
Природные аллели факторов инициации элонгации трансляции хозяина eif4e и eif4g также являются рецессивными генами вирусной устойчивости. Некоторые из них были использованы для борьбы с потивирусами в ячмене, рисе, томатах, перце, горохе, салате и дыне. Это открытие привело к успешному скринингу мутантов на химически индуцированные аллели eif4e в томатах. [ 1 ]
Естественная вариация промотора может привести к эволюции рецессивных аллелей устойчивости к болезням. Например, рецессивный ген устойчивости xa13 у риса является аллелем Os-8N3 . Os-8N3 транскрипционно активируется Xanthomonas oryzae pv. oryzae , которые экспрессируют эффектор TAL PthXo1 . Ген xa13 имеет мутированный эффектор-связывающий элемент в своем промоторе, который устраняет связывание PthXo1 и делает эти линии устойчивыми к штаммам, основанным на PthXo1 . Это открытие также продемонстрировало, что Os-8N3 необходим для чувствительности. [ 1 ]
Xa13/Os-8N3 необходим для развития пыльцы, показывая, что такие мутантные аллели могут быть проблематичными, если фенотип восприимчивости к болезням изменит функцию в других процессах. Однако мутации в эффектор-связывающем элементе TAL Os11N3 (OsSWEET14) были сделаны путем слияния эффекторов TAL с нуклеазами ( TALEN ). Растения риса с отредактированным геномом и измененными сайтами связывания Os11N3 оставались устойчивыми к Xanthomonas oryzae pv. oryzae , но при этом обеспечивал нормальную функцию разработки. [ 1 ]
Замалчивание генов
[ редактировать ]Устойчивость, основанная на подавлении РНК, является мощным инструментом для создания устойчивых культур. Преимущество РНКи как новой генной терапии против грибковых, вирусных и бактериальных инфекций у растений заключается в том, что она регулирует экспрессию генов посредством деградации информационной РНК , репрессии трансляции и ремоделирования хроматина с помощью небольших некодирующих РНК. Механически процессы молчания управляются продуктами процессинга триггера двухцепочечной РНК ( дцРНК ), которые известны как малые интерферирующие РНК и микроРНК . [ 53 ]
Диапазон хостов
[ редактировать ]Среди тысяч видов фитопатогенных микроорганизмов лишь незначительное меньшинство способно инфицировать широкий спектр видов растений. Вместо этого большинство патогенов проявляют высокую степень специфичности к хозяину. Часто говорят, что виды растений, не являющиеся хозяевами, обладают устойчивостью, не являющейся хозяином . Термин «резистентность хозяина» используется, когда вид патогена может быть патогенным для вида-хозяина, но определенные штаммы этого вида растений устойчивы к определенным штаммам вида патогена. Причины резистентности хозяина и резистентности нехозяина могут перекрываться. Круг хозяев возбудителя определяется, среди прочего, наличием соответствующих эффекторов, позволяющих колонизировать конкретного хозяина. [ 5 ] Диапазон хозяев патогена может измениться совершенно внезапно, если, например, способность патогена синтезировать специфичный для хозяина токсин или эффектор достигается за счет перетасовки/мутации генов или горизонтального переноса генов . [ 54 ] [ 55 ]
Эпидемии и популяционная биология
[ редактировать ]Аборигенные популяции часто характеризуются значительным генотипическим разнообразием и рассредоточенной популяцией (рост в смеси со многими другими видами растений). Они также претерпели коэволюцию растительных патогенов . Следовательно, пока новые патогены не интродуцируются/не развиваются, в таких популяциях обычно наблюдается лишь низкий уровень эпидемий тяжелых заболеваний . [ 56 ]
Монокультурные сельскохозяйственные системы обеспечивают идеальную среду для эволюции патогенов, поскольку они предлагают высокую плотность целевых образцов со схожими/идентичными генотипами. [ 56 ] Рост мобильности, обусловленный современными транспортными системами, обеспечивает патогенам доступ к большему количеству потенциальных целей. [ 56 ] Изменение климата может изменить жизнеспособный географический ареал видов патогенов и привести к тому, что некоторые заболевания станут проблемой в районах, где раньше эти заболевания не имели такого значения. [ 56 ]
Эти факторы делают современное сельское хозяйство более подверженным эпидемиям болезней. Общие решения включают постоянную селекцию на устойчивость к болезням, использование пестицидов, использование пограничных проверок и ограничений на импорт растений, поддержание значительного генетического разнообразия в генофонде сельскохозяйственных культур (см. Разнообразие сельскохозяйственных культур ) и постоянный надзор для ускорения принятия соответствующих ответных мер. Некоторые виды патогенов обладают гораздо большей способностью преодолевать устойчивость растений к болезням, чем другие, часто из-за их способности быстро развиваться и широко распространяться. [ 56 ]
Тематическое исследование болезни американского каштана
[ редактировать ]Фитофтороз каштанов был впервые замечен на американских каштанах, которые росли на территории, ныне известной как Зоопарк Бронкса, в 1904 году. подход к его контролю. Самые ранние попытки решить проблему с каштаном включали химические или физические растворы. Они пытались использовать фунгициды, срезать ветки деревьев, чтобы остановить заражение, и полностью удалять зараженные деревья из жилищ, чтобы не позволить им заразить другие. Все эти стратегии закончились неудачей. Были даже приняты карантинные меры, чему способствовало принятие Закона о карантине растений. Фитофтороз каштанов по-прежнему представлял собой огромную проблему, поскольку он быстро распространялся по густонаселенным каштановым лесам. В 1914 году рассматривалась идея вызвать у деревьев устойчивость к фитофторозу с помощью различных средств и механизмов размножения. [ 57 ]
См. также
[ редактировать ]- Отношения ген-за-геном
- Индуцированная системная резистентность
- Защита растений от травоядных
- Патология растений
- Использование растениями эндофитных грибов для защиты
- Системная приобретенная резистентность
Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж г час я дж к л м н тот п Дангл, Дж.Л.; Хорват, DM; Стаскавич, Б.Дж. (2013). «Переход иммунной системы растений от рассечения к развертыванию» . Наука . 341 (6147): 746–751. Бибкод : 2013Sci...341..746D . дои : 10.1126/science.1236011 . ПМЦ 3869199 . ПМИД 23950531 .
- ^ Jump up to: а б с Агриос, Джордж Н. (2005). Патология растений, пятое издание . Академическая пресса. ISBN 978-0120445653 .
- ^ Лутц, Диана (2012). Обнаружена ключевая часть системы быстрого реагирования растений. Архивировано 18 апреля 2023 г. в Wayback Machine . Вашингтонский университет в Сент-Луисе.
- ^ Jump up to: а б Дадакова, К.; Гавелкова, М.; Куркова Б.; Тлолкова И.; Каспаровский Т.; Здрагал, З.; Лохман, Дж. (24 апреля 2015 г.). «Анализ протеома и транскриптов культур клеток Vitis vinifera, подвергнутых инфекции Botrytis cinerea». Журнал протеомики . 119 : 143–153. дои : 10.1016/j.jprot.2015.02.001 . ПМИД 25688916 .
- ^ Jump up to: а б с д и Доддс, штат Пенсильвания; Ратьен, JP (2010). «Иммунитет растений: к комплексному взгляду на взаимодействие растений и патогенов». Обзоры природы Генетика . 11 (8): 539–548. дои : 10.1038/nrg2812 . hdl : 1885/29324 . ПМИД 20585331 . S2CID 8989912 .
- ^ Jump up to: а б с д Джонс, доктор медицинских наук; Дангл, Дж. Л. (2006). «Имунная система растений» . Природа . 444 (7117): 323–329. Бибкод : 2006Natur.444..323J . дои : 10.1038/nature05286 . ПМИД 17108957 .
- ^ Коуто, Д.; Зипфель, К. (2016). «Регуляция передачи сигналов рецепторов распознавания образов у растений» . Обзоры природы Иммунология . 16 (9): 537–552. дои : 10.1038/nri.2016.77 . ПМИД 27477127 . S2CID 21610374 .
- ^ Jump up to: а б Ли, Бо; Мэн, Сянцзун; Шан, Либо; Хэ, Пинг (май 2016 г.). «Транкрипционная регуляция паттерн-зависимого иммунитета у растений» . Клетка-хозяин и микроб . 19 (5): 641–650. дои : 10.1016/j.chom.2016.04.011 . ПМК 5049704 . ПМИД 27173932 .
- ^ Молина, Энтони; Боюсь, Ева; Басет, Лаура; Родригес, Тингуаро; Мелида, Хью; Денансе, Николас; Санчес-Валле, Андреа; Ривер, Мэри Питер; Лопес, Джемма; Фрейдье, Амандин; Барлет, Ксавье; Паттатил, Сивакумар; Хан, Майкл; Гоффнер, Дебора (2 февраля 2021 г.). «Состав клеточной стенки арабидопсиса определяет специфичность и пригодность устойчивости к болезням» . Труды Национальной академии наук . 118 (5):e2010243118. Бибкод : 2021PNAS..11810243M . дои : 10.1073/pnas.2010243118 . ISSN 0027-8424 . ПМЦ 7865177 . ПМИД 33509925 .
- ^ Jump up to: а б с Нюрнбергер, Торстен; Бруннер, Фредерик; Кеммерлинг, Биргит; Пиатер, Лизель (апрель 2004 г.). «Врожденный иммунитет у растений и животных: поразительное сходство и очевидные различия». Иммунологические обзоры . 198 (1): 249–266. дои : 10.1111/j.0105-2896.2004.0119.x . ПМИД 15199967 . S2CID 32547933 .
- ^ Томма, Б.; Нюрнбергер, Т.; Йостен, М. (2011). «О PAMP и эффекторах: размытая дихотомия PTI-ETI» . Растительная клетка . 23 (4): 4–15. дои : 10.1105/tpc.110.082602 . ПМК 3051239 . ПМИД 21278123 .
- ^ Даксбери, Зейн; Ма, Ян; Фурзер, Оливер Дж.; Ха, Сун Ын; Чевик, Волкан; Джонс, Джонатан Д.Г.; Саррис, Панайотис Ф. (24 июня 2016 г.). «Восприятие патогенов с помощью NLR у растений и животных: параллельные миры». Биоэссе . 38 (8). Уайли : 769–781. doi : 10.1002/bies.201600046 . ISSN 0265-9247 . ПМИД 27339076 . S2CID 3810233 .
- ^ Jump up to: а б Линдеберг, Магдалина; Куннак, Себастьян; Колмер, Алан (апрель 2012 г.). «Эффекторный репертуар Pseudomonas syringae типа III: последние слова в бесконечных спорах». Тенденции в микробиологии . 20 (4): 199–208. дои : 10.1016/j.tim.2012.01.003 . ПМИД 22341410 .
- ^ Jump up to: а б Рафики, Марьям; Эллис, Джеффри Дж.; Людовичи, Виктория А.; Хардхэм, Эдриенн Р.; Доддс, Питер Н. (2012). «Проблемы и прогресс в понимании роли эффекторов во взаимодействиях растений и грибов». Современное мнение в области биологии растений . 15 (4): 477–482. Бибкод : 2012COPB...15..477R . дои : 10.1016/j.pbi.2012.05.003 . ПМИД 22658704 .
- ^ Jump up to: а б Хевези, Тарек; Баум, Томас Дж. (январь 2013 г.). «Манипулирование растительными клетками с помощью эффекторов цист и корневых нематод» . Молекулярные растительно-микробные взаимодействия . 26 (1): 9–16. doi : 10.1094/MPMI-05-12-0106-FI . ПМИД 22809272 .
- ^ Джин, Приветствую (11 июля 2008 г.). «Эндогенные малые РНК и антибактериальный иммунитет растений» . Письма ФЭБС . 582 (18): 2679–2684. Бибкод : 2008FEBSL.582.2679J . doi : 10.1016/j.febslet.2008.06.053 . ISSN 0014-5793 . ПМЦ 5912937 . ПМИД 18619960 .
- ^ Падманабхан, Челлаппан; Чжан, Сяомин; Джин, Приветствую (август 2009 г.). «Малые РНК хозяина вносят большой вклад в врожденный иммунитет растений». Современное мнение в области биологии растений . 12 (4): 465–472. Бибкод : 2009COPB...12..465P . дои : 10.1016/j.pbi.2009.06.005 . ISSN 1369-5266 . ПМИД 19608454 .
- ^ Чжан, Вэйсюн; Гао, Шан; Чжоу, Сян; Челлаппан, Падманабхан; Чен, Чжэн; Чжоу, Сюэфэн; Чжан, Сяомин; Фромут, Нисса; Коутино, Габриэла (12 декабря 2010 г.). «Чувствительные к бактериям микроРНК регулируют врожденный иммунитет растений, модулируя сети гормонов растений» . Молекулярная биология растений . 75 (1–2): 93–105. дои : 10.1007/s11103-010-9710-8 . ISSN 0167-4412 . ПМК 3005105 . ПМИД 21153682 .
- ^ Боккара, Мартина; Саразин, Алексей; Тибо, Одон; Джей, Флоренция; Вуанне, Оливье; Наварро, Лайонел; Колот, Винсент (16 января 2014 г.). «Путь молчания miR472-RDR6 Arabidopsis модулирует иммунитет, запускаемый PAMP и эффекторами, посредством посттранскрипционного контроля генов устойчивости к болезням» . ПЛОС Патогены . 10 (1): e1003883. дои : 10.1371/journal.ppat.1003883 . ISSN 1553-7374 . ПМЦ 3894208 . ПМИД 24453975 . (Ошибка: дои : 10.1371/journal.ppat.1004814 , PMID 25859662 , часы втягивания )
- ^ Радван, Осман; Лю, Ю; Клаф, Стивен Дж. (август 2011 г.). «Транскрипционный анализ реакции корня сои на Fusarium virguliforme, возбудителя синдрома внезапной смерти» . Молекулярные растительно-микробные взаимодействия . 24 (8): 958–972. дои : 10.1094/mpmi-11-10-0271 . ISSN 0894-0282 . ПМИД 21751852 .
- ^ Синь, Минмин; Ван, Ю; Яо, Иньинь; Се, Чаоцзе; Пэн, Хуйру; Ни, Чжунфу; Сунь, Цисинь (2010). «Разнообразный набор микроРНК реагирует на заражение мучнистой росой и тепловой стресс у пшеницы ( Triticum aestivum L.)» . Биология растений BMC . 10 (1): 123. дои : 10.1186/1471-2229-10-123 . ISSN 1471-2229 . ПМК 3095282 . ПМИД 20573268 .
- ^ Инь, Цзуцзюнь; Ли, Ян; Хан, Сюлань; Шен, Фафу (25 апреля 2012 г.). «Полногеномное профилирование микроРНК и других малых некодирующих РНК в Verticillium dahliae – инокулированных корнях хлопчатника» . ПЛОС ОДИН . 7 (4): e35765. Бибкод : 2012PLoSO...735765Y . дои : 10.1371/journal.pone.0035765 . ISSN 1932-6203 . ПМЦ 3338460 . ПМИД 22558219 .
- ^ Лу, Шанфа; Сунь, Ин-Сюань; Амерсон, Генри; Чанг, Винсент Л. (7 августа 2007 г.). «МикроРНК в сосне лоблолли ( Pinus taeda L.) и их связь с развитием веретенообразных ржавчинных галлов» . Заводской журнал . 51 (6): 1077–1098. дои : 10.1111/j.1365-313x.2007.03208.x . ISSN 0960-7412 . ПМИД 17635765 .
- ^ Го, На; Йе, Вэнь-Ву; Ву, Сяо-Лин; Шен, Дан-Ю; Ван, Юань-Чао; Син, Хан; Доу, Дао-Лонг (ноябрь 2011 г.). «Профилирование микрочипов выявило микроРНК, влияющие на устойчивость сои к Phytophthora sojae ». Геном . 54 (11): 954–958. дои : 10.1139/g11-050 . ISSN 0831-2796 . ПМИД 21995769 .
- ^ Микали, Кристина О.; Нойманн, Улла; Грюневальд, Дорит; Панструга, Ральф; О'Коннелл, Ричард (28 октября 2010 г.). «Биогенез специализированного растительно-грибного интерфейса во время интернализации клетки-хозяина Golovinomyces orontii haustoria» . Клеточная микробиология . 13 (2): 210–226. дои : 10.1111/j.1462-5822.2010.01530.x . ISSN 1462-5814 . ПМИД 20880355 . S2CID 39002035 .
- ^ Кер, Дж.; Буц, А. (18 декабря 2007 г.). «Транспорт на большие расстояния и движение РНК через флоэму» . Журнал экспериментальной ботаники . 59 (1): 85–92. дои : 10.1093/jxb/erm176 . ISSN 0022-0957 . ПМИД 17905731 .
- ^ Гомес, Г.; Торрес, Х.; Паллас, В. (29 ноября 2004 г.). «Идентификация транслоцируемых РНК-связывающих белков флоэмы дыни, потенциальных компонентов системы транспорта РНК на большие расстояния» . Заводской журнал . 41 (1): 107–116. дои : 10.1111/j.1365-313x.2004.02278.x . ISSN 0960-7412 . ПМИД 15610353 .
- ^ Раттер, Брайан Д; Иннес, Роджер В. (август 2018 г.). «Внеклеточные везикулы как ключевые медиаторы растительно-микробных взаимодействий» . Современное мнение в области биологии растений . 44 : 16–22. Бибкод : 2018COPB...44...16R . дои : 10.1016/j.pbi.2018.01.008 . ISSN 1369-5266 . ПМИД 29452903 . S2CID 3377305 .
- ^ Гаг, Сиддхеш Б. (декабрь 2017 г.). «Вынужденное хозяином подавление генов - новая наука, направленная на создание устойчивости сельскохозяйственных культур к вредным патогенам растений». Физиологическая и молекулярная патология растений . 100 : 242–254. Бибкод : 2017PMPP..100..242G . дои : 10.1016/j.pmpp.2017.10.003 . ISSN 0885-5765 .
- ^ Новара, Даниэла; Гей, Александра; Лакомм, Кристоф; Шоу, Джейн; Ридаут, Кристофер; Душков, Димитр; Хензель, Гетц; Кумлен, Йохен; Швейцер, Патрик (сентябрь 2010 г.). «HIGS: индуцированное хозяином подавление генов у облигатного биотрофного грибкового патогена Blumeria graminis» . Растительная клетка . 22 (9): 3130–3141. дои : 10.1105/tpc.110.077040 . ISSN 1040-4651 . ПМЦ 2965548 . ПМИД 20884801 .
- ^ Вайберг, Арне; Ван, Мин; Линь, Фэн-Мао; Чжао, Хунвэй; Чжан, Чжихун; Калошян, Исгуи; Хуан, Сянь-Да; Джин, Приветствую (03.10.2013). «Грибковые малые РНК подавляют иммунитет растений, перехватывая пути взаимодействия РНК-хозяина» . Наука . 342 (6154): 118–123. Бибкод : 2013Sci...342..118W . дои : 10.1126/science.1239705 . ISSN 0036-8075 . ПМК 4096153 . ПМИД 24092744 .
- ^ Хаммонд-Козак, Ким Э; Паркер, Джейн Э. (апрель 2003 г.). «Расшифровка связи между растением и патогеном: новые перспективы селекции молекулярной устойчивости». Современное мнение в области биотехнологии . 14 (2): 177–193. дои : 10.1016/s0958-1669(03)00035-1 . ПМИД 12732319 .
- ^ Дадакова, Катерина; Клемпова, Житка; Ендрисакова, Тереза; Лохман, Ян; Каспаровский, Томас (01 декабря 2013 г.). «Выяснение сигнальных молекул, участвующих в восприятии эргостерина в табаке». Физиология и биохимия растений . 73 : 121–127. Бибкод : 2013PlPB...73..121D . дои : 10.1016/j.plaphy.2013.09.009 . ПМИД 24095918 .
- ^ Jump up to: а б Мур, Джон В.; Лоук, Гэри Дж.; Споул, Стивен Х. (август 2011 г.). «Динамика транскрипции в иммунитете растений» . Растительная клетка . 23 (8): 2809–2820. дои : 10.1105/tpc.111.087346 . ПМК 3180793 . ПМИД 21841124 .
- ^ Бюргер, Марко; Чори, Джоан (14 августа 2019 г.). «Стресс из-за гормонов: как растения управляют иммунитетом» . Клетка-хозяин и микроб . 26 (2): 163–172. дои : 10.1016/j.chom.2019.07.006 . ISSN 1934-6069 . ПМЦ 7228804 . ПМИД 31415749 .
- ^ Jump up to: а б с Саданандом, Ари; Бейли, Марк; Юэн, Ричард; Ли, Джек; Нелис, Стюарт (октябрь 2012 г.). «Система убиквитин-протеасома: центральный модификатор передачи сигналов растений» . Новый фитолог . 196 (1): 13–28. дои : 10.1111/j.1469-8137.2012.04266.x . ПМИД 22897362 .
- ^ Jump up to: а б Трухильо, М.; Ширасу, К. (август 2010 г.). «Убиквитинирование в иммунитете растений». Современное мнение в области биологии растений . 13 (4): 402–408. Бибкод : 2010COPB...13..402T . дои : 10.1016/j.pbi.2010.04.002 . ПМИД 20471305 .
- ^ Крейг, А.; Юэн, Р.; Месмар, Дж.; Гудипати, В.; Саданандом, А. (1 марта 2009 г.). «Убиквитинлигазы Е3 и врожденный иммунитет растений» . Журнал экспериментальной ботаники . 60 (4): 1123–1132. дои : 10.1093/jxb/erp059 . ПМИД 19276192 .
- ^ Мун, Дж. (1 декабря 2004 г.). «Путь убиквитин-протеасома и развитие растений» . Растительная клетка онлайн . 16 (12): 3181–3195. дои : 10.1105/tpc.104.161220 . ПМЦ 535867 . ПМИД 15579807 .
- ^ Трухильо, Марко; Ширасу, Кен (1 августа 2010 г.). «Убиквитинирование в иммунитете растений». Современное мнение в области биологии растений . 13 (4): 402–408. Бибкод : 2010COPB...13..402T . дои : 10.1016/j.pbi.2010.04.002 . ПМИД 20471305 .
- ^ Ширсекар, Гаутама; Дай, Лянъин; Ху, Яджун; Ван, Сюэцзюнь; Цзэн, Лижун; Ван, Го-Лян (февраль 2010 г.). «Роль убиквитинирования во врожденном иммунитете растений и вирулентности патогена». Журнал биологии растений . 53 (1): 10–18. Бибкод : 2010JPBio..53...10S . дои : 10.1007/s12374-009-9087-x . S2CID 36185464 .
- ^ Jump up to: а б с Каугер, Кристина; Браун, Джеймс К.М. (25 августа 2019 г.). «Долговечность количественной устойчивости сельскохозяйственных культур: больше, чем мы знаем?». Ежегодный обзор фитопатологии . 57 (1). Годовые обзоры : 253–277. doi : 10.1146/annurev-phyto-082718-100016 . ISSN 0066-4286 . ПМИД 31206351 . S2CID 190533925 .
- ^ Jump up to: а б с Парк, Роберт Ф.; Голегаонкар, Прашант Г.; Деревнина, Лида; Сандху, Каранджит С.; Караоглу, Хайдар; Эльмансур, Худа М.; Дракатос, Питер М.; Сингх, Давиндер (4 августа 2015 г.). «Листовая ржавчина культурного ячменя: патология и борьба». Ежегодный обзор фитопатологии . 53 (1). Годовые обзоры : 565–589. doi : 10.1146/annurev-phyto-080614-120324 . ISSN 0066-4286 . ПМИД 26047566 .
- ^ Бхавани, Шридхар (18 июня 2021 г.). «Устойчивость взрослых растений (APR): стратегия борьбы с устойчивыми патогенами» . CIMMYT (Международный центр улучшения кукурузы и пшеницы) . Проверено 6 июля 2021 г.
- ^ Сингх, Рави П.; Сингх, Паван К.; Руткоски, Джессика; Ходсон, Дэвид П.; Он, Синьяо; Йоргенсен, Лиза Н.; Ховмёллер, Могенс С.; Уэрта-Эспино, Хулио (4 августа 2016 г.). «Влияние болезни на потенциал урожайности пшеницы и перспективы генетического контроля». Ежегодный обзор фитопатологии . 54 (1). Годовые обзоры : 303–322. doi : 10.1146/annurev-phyto-080615-095835 . ISSN 0066-4286 . ПМИД 27296137 . S2CID 4603818 .
- ^ Jump up to: а б Статман, Д.Д.; Леонард, К.Дж.; Миллер-Гарвин, Дж. (2007). «Селекция культур на длительную устойчивость к болезням». Достижения в агрономии . 95 : 319–367. дои : 10.1016/S0065-2113(07)95004-X . ISBN 9780123741653 .
- ^ Шимелис, Х.; Лэнг, М. «Сроки улучшения традиционных сельскохозяйственных культур: процедуры предварительной селекции и селекции». Австралийский журнал растениеводства : 1542–1549. eISSN 1835-2707 . ISSN 1835-2693 . S2CID 55486617 .
- ^ Махлейн, А.-К.; Кушка, Монтана; Беманн, Дж.; Польдер, Г.; Уолтер, А. (25 августа 2018 г.). «Гиперспектральные датчики и технологии визуализации в фитопатологии: современное состояние». Ежегодный обзор фитопатологии . 56 (1). Годовые обзоры : 535–558. doi : 10.1146/annurev-phyto-080417-050100 . ISSN 0066-4286 . ПМИД 30149790 . S2CID 52096158 .
- ^ Табашник, Брюс Э.; Брево, Тьерри; Карьер, Ив (2013). «Устойчивость насекомых к Bt-культурам: уроки первого миллиарда акров». Природная биотехнология . 31 (6): 510–521. дои : 10.1038/nbt.2597 . ПМИД 23752438 . S2CID 205278530 .
- ^ Кавана, штат Техас; Спиллейн, К. (1 февраля 1995 г.). «Стратегии создания устойчивости к вирусам у трансгенных растений». Эвфитика . 85 (1–3): 149–158. дои : 10.1007/BF00023943 . ISSN 0014-2336 . S2CID 20940279 .
- ^ Каневский, Войцех К.; Томас, Питер Э. (2004). «Картофельная история». АгБиоФорум . 7 (1 и 2): 41–46.
- ^ Лакомб, Северин; Ругон-Кардозо, Алехандра; Шервуд, Эмма; Питерс, Немо; Дальбек, Дуглас; ван Эссе, Х. Питер; Смокер, Мэтью; Раллапалли, Ганасьям; Томма, Барт PHJ; Стаскавич, Брайан; Джонс, Джонатан Д.Г.; Зипфель, Кирилл (17 апреля 2010 г.). «Межсемейный перенос растительного рецептора распознавания образов обеспечивает бактериальную устойчивость широкого спектра» . Природная биотехнология . 28 (4): 365–369. дои : 10.1038/nbt.1613 . ПМИД 20231819 . S2CID 7260214 – через www.nature.com.
- ^ Картикеян А.; Дейвамани, М.; Шобхана, В.Г.; Судха, М.; Анандхан, Т. (2013). «РНК-интерференция: эволюция и применение в борьбе с болезнями растений». Архив фитопатологии и защиты растений . 46 (12): 1430–1441. Бибкод : 2013ArPPP..46.1430K . дои : 10.1080/03235408.2013.769315 . S2CID 85060938 .
- ^ Беттгенхойзер, Ян; Гилберт, Брайан; Эйлифф, Майкл; Москва, Мэтью Дж. (11 декабря 2014 г.). «Нехозяинная устойчивость к возбудителям ржавчины – продолжение континуума» . Границы в науке о растениях . 5 : 664. doi : 10.3389/fpls.2014.00664 . ПМЦ 4263244 . ПМИД 25566270 .
- ^ Рестрепо, Сильвия; Табима, Хавьер Ф.; Мидерос, Мария Ф.; Грюнвальд, Никлаус Дж.; Матуте, Дэниел Р. (4 августа 2014 г.). «Видообразование грибковых и оомицетовых патогенов растений» . Ежегодный обзор фитопатологии . 52 (1). Годовые обзоры: 289–316. doi : 10.1146/annurev-phyto-102313-050056 . ISSN 0066-4286 . ПМИД 24906125 .
- ^ Jump up to: а б с д и Макдональд, бакалавр; Линде, К. (2002). «Генетика популяции патогенов, эволюционный потенциал и устойчивая устойчивость». Ежегодный обзор фитопатологии . 40 : 349–379. doi : 10.1146/annurev.phyto.40.120501.101443 . ПМИД 12147764 . S2CID 23726106 .
- ^ Барнс, Джессика С; Делборн, Джейсон А. (сентябрь 2022 г.). «Политика генетической технонауки в целях сохранения: случай устойчивого к фитофторозу американского каштана» . Окружающая среда и планирование E: Природа и космос . 5 (3): 1518–1540. Бибкод : 2022EnPlE...5.1518B . дои : 10.1177/25148486211024910 .
Дальнейшее чтение
[ редактировать ]- Лукас, Дж. А., «Защита растений». Глава 9 патологии растений и патогенов растений, 3-е изд. 1998 Блэквелл Наука. ISBN 0-632-03046-1
- Хаммонд-Косак К. и Джонс Дж.Д. «Реакция на патогены растений». В: Бьюкенен, Груиссем и Джонс, ред. Биохимия и молекулярная биология растений, второе издание. 2015. Уайли-Блэквелл, Хобокен, Нью-Джерси. ISBN 9780470714218
- Доддс, П.; Ратьен, Дж. (2010). «Иммунитет растений: к комплексному взгляду на взаимодействие растений и патогенов». Обзоры природы Генетика . 11 (8): 539–548. дои : 10.1038/nrg2812 . hdl : 1885/29324 . ПМИД 20585331 . S2CID 8989912 .
- Мишельмор, Ричард В.; Христопулу, Марилена; Колдуэлл, Кэтрин С. (4 августа 2013 г.). «Влияние генетики, функции и эволюции генов устойчивости на прочное будущее». Ежегодный обзор фитопатологии . 51 (1). Годовые обзоры : 291–319. doi : 10.1146/annurev-phyto-082712-102334 . ISSN 0066-4286 . ПМИД 23682913 . S2CID 22234708 .
- Шуман, Г. Болезни растений: их биология и социальное воздействие. 1991 APS Press, Сент-Пол, Миннесота. ISBN 0890541167