Марчантиофита
| Печеночники | |
|---|---|

| |
| «Hepaticae» из Эрнста Геккеля книги «Художественные формы природы» , 1904 год. | |
| Научная классификация | |
| Королевство: | Растения |
| Клэйд : | Эмбриофиты |
| Клэйд : | Сетафита |
| Разделение: | Марчантиофита Стотлер и Стотл.-Выпускник, 1977 г. [ 2 ] купить 2000 г. [ 3 ] |
| Занятия и заказы | |
Marchantiophyta ( / m ɑːr ˌ k n æ t i ˈ ɒ f ə ə t , - oʊ ˈ f aɪ t ə / ) представляют собой подразделение несосудистых наземных растений, обычно называемых печеночными или печеночниками . Подобно мхам и роголистникам , они имеют жизненный цикл с доминированием гаметофита , в котором клетки растения несут только один набор генетической информации.
Подсчитано, что существует около 9000 видов печеночников. [ 4 ] Некоторые из наиболее известных видов растут в виде сплюснутого безлистного слоевища , но большинство видов покрыты листвой и по форме очень напоминают сплющенный мох . Листовые виды можно отличить от внешне сходных мхов по ряду особенностей, в том числе по одноклеточным ризоидам . Листовые печеночники также отличаются от большинства (но не всех) мхов тем, что их листья никогда не имеют ребер ( присутствуют у многих мхов) и могут нести краевые реснички (очень редко у мхов). Другие различия не универсальны для всех мхов и печеночников, но наличие трехрядного расположения листьев, наличие глубоких лопастей или сегментированных листьев или отсутствие четко дифференцированных стебля и листьев указывают на то, что растение является печеночником. Печеночники отличаются от мхов наличием уникальных сложных маслянистых тел с высоким показателем преломления.
Печеночники обычно небольшие, обычно 2–20 мм (0,079–0,787 дюйма) в ширину, а отдельные растения имеют длину менее 10 см (3,9 дюйма), и поэтому их часто упускают из виду. Однако некоторые виды могут покрывать большие участки земли, камней, деревьев или любого другого достаточно твердого субстрата, на котором они встречаются. Они распространены по всему миру почти во всех доступных средах обитания, чаще всего во влажных местах, хотя встречаются также пустынные и арктические виды. Некоторые виды могут причинять неудобства в тенистых теплицах или сорняки в садах. [ 5 ]
Физические характеристики
[ редактировать ]Описание
[ редактировать ]Большинство печеночников небольшие, их ширина составляет 2–20 миллиметров (0,08–0,8 дюйма), а длина отдельных растений составляет менее 10 сантиметров (4 дюйма). [ 6 ] поэтому их часто упускают из виду. Наиболее знакомые печеночники состоят из распростертой, уплощенной, лентовидной или разветвленной структуры, называемой слоевищем ( телом растения); эти печеночники называются слоевищными печеночниками . Однако у большинства печеночников стебли уплощены с перекрывающимися чешуйками или листьями в два или более рядов, причем средний ряд часто заметно отличается от внешних рядов; их называют листоватыми печеночниками или чешуйчатыми печеночниками . [ 7 ] [ 8 ] ( см . в галерее Примеры ниже. )

Печеночников наиболее достоверно отличить от внешне сходных мхов по одноклеточным ризоидам . [ 9 ] Другие различия не универсальны для всех мхов и всех печеночников; [ 8 ] но отсутствие четко дифференцированных стебля и листьев у слоевищных видов или у листоватых видов наличие глубоколопастных или сегментированных листьев и наличие листьев, расположенных в три ряда, [ 10 ] [ 11 ] а также частое дихотомическое ветвление указывают на то, что это растение является печеночником. За некоторыми исключениями, все печеночники подвергаются полипластидному мейозу, в отличие от мхов и роголистников, у которых наблюдается монопластидный мейоз. [ 12 ] В отличие от любых других эмбриофитов, большинство печеночников содержат уникальные мембраносвязанные масляные тельца , содержащие изопреноиды, по крайней мере, в некоторых их клетках, при этом липидные капли в цитоплазме всех других растений не закрыты. [ 13 ] Общее физическое сходство некоторых мхов и листоватых печеночников означает, что подтверждение идентификации некоторых групп может быть произведено с уверенностью только с помощью микроскопии или опытного бриолога .
У печеночников, как и у других мохообразных, в жизненном цикле преобладают гаметофиты , при этом спорофит зависит от гаметофита. [ 13 ] Спорофиты многих печеночников не фотосинтезируют, но есть и некоторые, которые в разной степени фотосинтезируют. [ 14 ] Каждая клетка типичного растения печеночника содержит только один набор генетической информации, поэтому клетки растения гаплоидны на протяжении большей части его жизненного цикла. Это резко контрастирует с картиной, наблюдаемой почти у всех животных и сосудистых растений. У более знакомых семенных растений гаплоидное поколение представлено только крошечной пыльцой и семязачатком , тогда как диплоидное поколение представляет собой знакомое дерево или другое растение. [ 15 ] Еще одной необычной особенностью жизненного цикла печеночника является то, что спорофиты (т.е. диплоидное тело) очень недолговечны и отмирают вскоре после высвобождения спор. [ 16 ] У мхов спорофит более устойчив, а у роголистников спорофит рассеивает споры в течение длительного периода времени. [ нужна ссылка ]
Жизненный цикл
[ редактировать ]
Жизнь печеночника начинается с прорастания гаплоидной споры с образованием протонемы , которая представляет собой либо массу нитевидных нитей, либо уплощенное слоевище. [ 17 ] [ 18 ] Протонема — переходный этап в жизни печеночника, из которого вырастает зрелое растение-гаметофор (« носитель гамет »), производящее половые органы. Мужские органы известны как антеридии ( единственное число: антеридии) и производят сперматозоиды. Скопления антеридиев окружены защитным слоем клеток, называемым перигонием ( множественное число: перигония). Как и у других наземных растений, женские органы известны как архегонии ( единственное число: архегониум) и защищены тонкой окружающей перихетумом ( множественное число: перихета). [ 8 ] Каждый архегоний имеет тонкую полую трубку, «шею», по которой сперматозоиды плывут к яйцеклетке.
Виды печеночников могут быть как двудомными , так и однодомными . У двудомных печеночников женские и мужские половые органы расположены на разных растениях-гаметофитах. У однодомных печеночников два типа репродуктивных структур расположены на разных ветвях одного и того же растения. [ 19 ] В любом случае сперматозоиды должны переместиться из антеридиев, где они производятся, в архегоний, где содержатся яйцеклетки. Сперматозоиды . печеночников двужгутиковые , т. е. имеют два хвостообразных жгутика , которые позволяют им плавать на короткие расстояния [ 20 ] при условии наличия хотя бы тонкой пленки воды. Их путешествию может способствовать плеск капель дождя. В 2008 году японские исследователи обнаружили, что некоторые печеночники способны выбрасывать содержащую сперму воду на высоту до 15 см в воздух, что позволяет им оплодотворять женские растения, растущие на расстоянии более метра от ближайшего самца. [ 21 ]
Когда сперматозоиды достигают архегониев, происходит оплодотворение , приводящее к образованию диплоидного спорофита. После оплодотворения у незрелого спорофита внутри архегония развиваются три отдельные области: (1) ножка , которая одновременно закрепляет спорофит на месте и получает питательные вещества от его «материнского» растения, (2) сферическая или эллипсоидная капсула , внутри которой споры будет произведен для распространения в новые места, и (3) щетинка (стебель), которая лежит между двумя другими областями и соединяет их. [ 20 ] У спорофита отсутствует апикальная меристема , чувствительная к ауксину точка расхождения с другими наземными растениями, возникшая где-то в позднем силуре / раннем девоне . [ 22 ] [ 23 ] Когда у спорофита развились все три области, щетинка удлиняется, выталкиваясь из архегония и разрывая его. Пока ножка остается прикрепленной к родительскому растению, капсула выталкивается щетинкой и вытягивается от растения в воздух. Внутри капсулы клетки делятся, образуя как более поздние клетки, так и клетки, продуцирующие споры. Элаты подобны пружинам и раздвигают стенку капсулы, чтобы разлететься, когда капсула лопнет. Клетки, производящие споры, подвергаются мейозу с образованием гаплоидных спор , которые затем рассеются, после чего жизненный цикл может начаться снова.
Бесполое размножение
[ редактировать ]Некоторые печеночники способны к бесполому размножению ; у мохообразных вообще «было бы почти верно сказать, что вегетативное размножение является правилом, а не исключением». [ 24 ] Например, у Riccia , когда старые части раздвоенных слоевищ отмирают, молодые кончики становятся отдельными особями. [ 24 ]
Некоторые слоевищные печеночники, такие как Marchantia Polymorpha и Lunularia Crusata, образуют небольшие дискообразные почки в неглубоких чашечках. [ 25 ] Marchantia gemmae может быть рассеяна на высоту до 120 см под действием дождя, попадающего в чашки. [ 26 ] У Мецгерии почки растут по краям слоевища. [ 27 ] Marchantia polymorpha – распространенный сорняк в теплицах, часто покрывающий всю поверхность контейнеров; [ 28 ] : 230 Распространение геммы - это «основной механизм, с помощью которого печеночник распространяется по питомнику или теплице». [ 28 ] : 231
Симбиоз
[ редактировать ]Таллоидные печеночники обычно являются местом обитания симбиотических грибов -гломеромицетов , которые имеют арбускулярные (несущие реснички) корешки, напоминающие корешки сосудистых растений. виды Aneuraceae связаны Однако с грибами -базидиомицетами, принадлежащими к роду Tulasnella , в то время как листовые печеночники обычно содержат симбиотические грибы-базидиомицеты, принадлежащие к роду Serendipita . [ 29 ]
Экология
[ редактировать ]Сегодня печеночников можно встретить во многих экосистемах по всей планете, за исключением моря и чрезмерно засушливых сред или тех, которые подвергаются высокому уровню прямой солнечной радиации. [ 30 ] Как и большинство групп живых растений, они наиболее распространены (как по численности, так и по видам) во влажных тропических районах. [ 31 ] Печеночники чаще встречаются в тени от умеренной до глубокой, хотя пустынные виды могут переносить прямой солнечный свет и периоды полного высыхания.
Классификация
[ редактировать ]Отношения с другими растениями
[ редактировать ]Традиционно печеночники группировались вместе с другими мохообразными ( мхами и роголистниками ) в отдел Bryophyta, внутри которого печеночники составляли класс Hepaticae (также называемый Marchantiopsida). [ 8 ] [ 32 ] Несколько позже печеночникам был выделен отдельный отдел (Marchantiophyta), [ 33 ] поскольку мохообразные стали считаться парафилетическими . Однако самые последние филогенетические данные показывают, что печеночники действительно, вероятно, являются частью монофилетической клады («Bryophyta sensu lato » или «Bryophyta Schimp.») наряду со мхами и роголистниками. [ 34 ] [ 35 ] [ 36 ] [ 37 ] [ 38 ] [ 39 ] [ 40 ] [ 41 ] [ 42 ] [ чрезмерное цитирование ] Поэтому было предложено перевести печеночников в класс под названием Marchantiopsida. [ 36 ] Кроме того, существуют убедительные филогенетические данные, позволяющие предположить, что печеночники и мхи образуют монофилетический субклад Setaphyta . [ 35 ] [ 43 ] [ 44 ]
| Модель «Монофилетические мохообразные» | Модель «Печеночники плюс мхи – базальная» | |||||
|---|---|---|---|---|---|---|
| ||||||
| Две наиболее вероятные модели эволюции мохообразных. [ 44 ] | ||||||
Важным выводом из этой филогении является то, что предковые устьица, по-видимому, были утеряны в линии печеночников. [ 35 ] [ 39 ] Среди самых ранних окаменелостей, которые, как полагают, относятся к печеночникам, являются окаменелости Pallaviciniites компрессионные из верхнего девона Нью - Йорка . [ 45 ] Эти окаменелости напоминают современные виды Metzgeriales . [ 46 ] Еще одно девонское ископаемое, называемое Protosalvinia, также похоже на печеночник, но его связь с другими растениями до сих пор неясна, поэтому он может не принадлежать к Marchantiophyta. В 2007 году были объявлены самые старые окаменелости, относимые на тот момент к печеночникам, Metzgeriothallus sharonae из живета (средний девон ) Нью-Йорка , США. [ 47 ] Однако в 2010 году в Аргентине было обнаружено пять различных типов окаменелых спор печеночника, датируемых гораздо более ранним средним ордовиком , около 470 миллионов лет назад. [ 1 ] [ 48 ]
Внутренняя классификация
[ редактировать ]Бриологи относят печеночников к отделу Marchantiophyta . Это подразделенное название основано на названии наиболее общепризнанного рода печёночников Marchantia . [ 49 ] В дополнение к этому названию, основанному на таксоне , печеночников часто называют Hepaticophyta . Это название происходит от их общего латинского названия, поскольку латынь была языком, на котором ботаники публиковали свои описания видов. Это название привело к некоторой путанице, [ нужна ссылка ] отчасти потому, что это, по-видимому, таксонное название, полученное от рода Hepatica , который на самом деле является цветковым растением семейства лютиковых Ranunculaceae . Кроме того, название Hepaticophyta в учебниках часто пишется с ошибкой как Hepatophyta , что только усугубляет путаницу.
Хотя среди бриологов нет единого мнения относительно классификации печеночников выше семейного ранга, [ 50 ] Marchantiophyta можно разделить на три класса: [ 51 ] [ 52 ] [ 53 ] [ 54 ]
- Юнгерманниопсида : включает два порядка Metzgeriales (простые слоевища) и Jungermanniales (листистые печеночники).
- относятся К Marchantiopsida три порядка Marchantiales (сложно-слоевиковые печеночники) и Sphaerocarpales (бутылочные печеночники), а также Blasiales (ранее отнесенные к Metzgeriales). [ 51 ] [ 55 ] Сюда также входит проблемный род Monoclea , который иногда относят к отдельному отряду Monocleales. [ 56 ]
- Третий класс, Haplomitriopsida, недавно был признан сестринской группой других печеночников; [ 54 ] он включает роды Haplomitrium , Treubia и Apotreubia .
| Форрест 2006 г. [ 51 ] | Коул, Хильгер и Гоффине 2021 [ 57 ] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
|
Обновленная классификация Söderström et al. 2016 год [ 58 ]
- Marchantiophyta Stotler и Крэндалл-Стотлер, 2000 г.
- Гапломитриопсида Стотлер и Крэндалл-Стотлер, 1977 г.
- Калобрьялес Хэмлин 1972
- Треубиалес Шляков 1972 г.
- Маркантиопсида Кронквист, Тахтаян и Циммерманн, 1966 г.
- Blasiidae He-Nygrén et al. 2006 г.
- Блазиалес Стотлер и Крэндалл-Стотлер, 2000 г.
- Marchantiidae Engler 1893 sensu He-Nygrén et al. 2006 г.
- Лунуляриалес Лонг 2006
- Marchantiales Limpricht 1877 (сложные слоевища)
- Неоходгсониалес Лонг 2006
- Sphaerocarpales Cavers 1910 (печеночники бутылочные)
- Blasiidae He-Nygrén et al. 2006 г.
- Юнгерманниопсида Стотлер и Крэндалл-Стотлер, 1977 г.
- Jungermanniidae Engler 1893 (печеночники листовые)
- Юнгерманниалы Клинггреффа, 1858 г.
- Порельялес Шляков 1972 г.
- Птилидиалес Шляков 1972 г.
- Metzgeriidae Bartholomew-Начало 1990 г.
- Мецгериалес Шало 1930 г.
- Плеврозиалес Шляков 1972 г.
- Pelliidae He-Nygrén et al. 2006 г.
- Фоссомброниалы Шляков 1972 г.
- Паллавичиниалес Фрей и Стек, 2005 г.
- Пеллиалес Хе-Нигрен и др. 2006 г.
- Jungermanniidae Engler 1893 (печеночники листовые)
- Гапломитриопсида Стотлер и Крэндалл-Стотлер, 1977 г.
Подсчитано, что существует около 9000 видов печеночников, не менее 85% из которых относятся к листовой группе. [ 3 ] [ 59 ] Несмотря на этот факт, на сегодняшний день геномы печеночников не секвенированы, и лишь немногие гены идентифицированы и охарактеризованы. [ 60 ]
Экономическое значение
[ редактировать ]В древности считалось, что печеночник излечивает заболевания печени , отсюда и название. [ 61 ] В древнеанглийском языке слово «печеночник» буквально означает «печеночное растение» . [ 62 ] Вероятно, это произошло из-за внешнего вида некоторых слоевищных печеночников, которые по очертаниям напоминают печень, и привело к общему названию группы как печеночные , от латинского слова hēpaticus , означающего «принадлежащий печени». Неродственное цветущее растение Hepatica иногда также называют печеночником, поскольку когда-то оно также использовалось для лечения заболеваний печени. Эта архаичная связь формы растения с функцией была основана на «Доктрине сигнатур» . [ 63 ]
Сегодня печеночники не имеют прямого экономического значения. Наибольшее их влияние носит косвенный характер, за счет уменьшения эрозии берегов рек, сбора и удержания воды в тропических лесах, а также образования почвенной корки в пустынях и полярных регионах. Однако некоторые виды используются человеком напрямую. Некоторые виды, такие как Riccia fluitans , представляют собой водные слоевища печеночников, которые продаются для использования в аквариумах . Их тонкие, стройные ветви плавают на поверхности воды и обеспечивают среду обитания как мелким беспозвоночным, так и питающимся ими рыбам.
Галерея
[ редактировать ]Небольшая коллекция изображений, показывающих структуру и разнообразие печеночников:
-
 Marchantia polymorpha с антеридиальными и архегониальными стеблями.
Marchantia polymorpha с антеридиальными и архегониальными стеблями. -
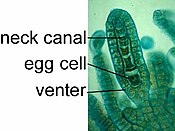 Архегоний Пореллы .
Архегоний Пореллы . -
 Спорофит пореллы, выходящий из архегония.
Спорофит пореллы, выходящий из архегония. -
 Комок Porella platyphylla , растущий на дереве.
Комок Porella platyphylla , растущий на дереве. -
 Pellia epiphylla , растет на влажной почве.
Pellia epiphylla , растет на влажной почве. -
 Plagiochila asplenioides — печеночник листовой .
Plagiochila asplenioides — печеночник листовой . -
 Riccia fluitans , водная слоевище печеночника .
Riccia fluitans , водная слоевище печеночника . -
 Conocephalum conicum — крупная слоевищница печеночника.
Conocephalum conicum — крупная слоевищница печеночника.








См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б Уокер, Мэтт. «Окаменелости древнейших наземных растений, обнаруженные в Аргентине» [1] . (Би-би-си, Новости Земли, 2010).
- ^ Стотлер, Раймонд Э.; Барбара Дж. Кэндалл-Стотлер (1977). «Контрольный список печеночников и роголистников Северной Америки». Бриолог . 80 (3). Американское бриологическое и лихенологическое общество: 405–428. дои : 10.2307/3242017 . JSTOR 3242017 .
- ^ Перейти обратно: а б Крэндалл-Стотлер, Барбара; Стотлер, Раймонд Э. (2000). «Морфология и классификация Marchantiophyta». В А. Джонатане Шоу; Бернар Гоффине (ред.). Биология мохообразных . Кембридж: Издательство Кембриджского университета. п. 21. ISBN 0-521-66097-1 .
- ^ "Домашняя страница печеночников | UNB" . Архивировано из оригинала 24 июля 2021 года . Проверено 10 июня 2020 г.
- ^ Шустер, Рудольф М. (1992). Hepaticae и Anthocerotae Северной Америки . Том. VI. Чикаго: Полевой музей естественной истории. п. 19. ISBN 0-914868-21-7 .
- ^ Шустер, Рудольф М. Hepaticae и Anthocerotae Северной Америки , том. Я, стр. 243–244. (Нью-Йорк: издательство Колумбийского университета, 1966)
- ^ Кашьяп, Шив Рам. Печеночники Западных Гималаев и Пенджабской равнины , вып. я, с. 1. (Нью-Дели: The Chronica Botanica, 1929).
- ^ Перейти обратно: а б с д Шофилд, Всемирный банк. Введение в бриологию , стр. 135–140. (Нью-Йорк: Макмиллан, 1985). ISBN 0-02-949660-8 .
- ^ Нехира, Кунито. «Прорастание спор, развитие протонем и развитие спорлингов», с. 347 в Рудольфе М. Шустере (ред.), Новом руководстве по бриологии , том I. (Ничинан, Миядзаки, Япония: Ботаническая лаборатория Хаттори, 1983). ISBN 49381633045 .
- ^ Эллисон, К.В. и Джон Чайлд. Печеночники Новой Зеландии , стр. 13–14. (Данедин: Университет Отаго Press, 1975).
- ^ Конард, Генри С. и Пол Л. Редфирн-младший. « Как узнать мхи и печеночники» , исправленное издание, стр. 12–23. (Дубьюк, Айова: компания William C. Brown Co., 1979) ISBN 0-697-04768-7
- ^ Спорогенез у Physcomitrium patens: сотрудничество между поколениями и развитие стенки и апертуры спор.
- ^ Перейти обратно: а б Гарольд С. Болд, Си Джей Алексопулос и Т. Делевориас. Морфология растений и грибов , 5-е изд., с. 189. (Нью-Йорк: Харпер-Коллинз, 1987). ISBN 0-06-040839-1 .
- ^ Том 1, Глава 11-1: Фотосинтез: процесс
- ^ Фоскет, Дональд Э. Рост и развитие растений: молекулярный подход , с. 27. (Сан-Диего: Academic Press, 1994). ISBN 0-12-262430-0 .
- ^ Хикс, Мари Л. Путеводитель по печеночникам Северной Каролины , стр. 10. (Дарем: Издательство Университета Дьюка, 1992). ISBN 0-8223-1175-5 .
- ^ Нехира, Кунито. «Прорастание спор, развитие протонем и развитие спорлингов», стр. 358–374 в книге Рудольфа М. Шустера (ред.), Новое руководство по бриологии , том I. (Ничинан, Миядзаки, Япония: Ботаническая лаборатория Хаттори, 1983). ISBN 49381633045 .
- ^ Чопра, Р.Н. и ПК Кумра. Биология мохообразных , стр. 1–38. (Нью-Йорк: Джон Уайли и сыновья, 1988). ISBN 0-470-21359-0 .
- ^ Малькольм, Билл и Нэнси Малкольм. Мхи и другие мохообразные: иллюстрированный глоссарий , стр. 6 и 128. (Новая Зеландия: Micro-Optics Press, 2000). ISBN 0-473-06730-7 .
- ^ Перейти обратно: а б Кэмпбелл, Дуглас Х. Структура и развитие мхов и папоротников , стр. 73–74. (Лондон: The Macmillan Co., 1918)
- ^ Боль, С. (2010). «Ботаническая баллистика». Новый учёный . 208 (2792/3): 45–47. дои : 10.1016/s0262-4079(10)63177-6 .
- ^ Кук, Тодд Дж; Поли, ДоротиБелль; Коэн, Джерри Д. (2003). «Сыграл ли ауксин решающую роль в эволюции нового строения тела во время позднесилурийской-раннедевонской радиации наземных растений?». Эволюция физиологии растений . Эльзевир . стр. 85–107. дои : 10.1016/b978-012339552-8/50006-8 . ISBN 978-0-12-339552-8 .
- ^ Фридман, Уильям Э.; Мур, Ричард С.; Пуруганан, Майкл Д. (2004). «Эволюция развития растений» . Американский журнал ботаники . 91 (10). Ботаническое общество Америки ( Уайли ): 1726–1741. дои : 10.3732/ajb.91.10.1726 . ISSN 0002-9122 . ПМИД 21652320 .
- ^ Перейти обратно: а б Лепп, Хейно (15 апреля 2008 г.). «Вегетативное размножение» . Австралийские мохообразные . Австралийский национальный ботанический сад . Проверено 22 декабря 2011 г.
- ^ Смит, AJE (1989) Печеночники Великобритании и Ирландии , Издательство Кембриджского университета, Кембридж.
- ^ Экихуа, К. (1987). «Распространение гемм в печеночнице Marchantia-Polymorpha» . Криптогамия, Бриология, Лихенология . 8 (3): 199–217. Архивировано из оригинала 26 апреля 2012 года.
- ^ Лепп, Хейно (28 февраля 2008 г.). «Воспроизведение и распространение» . Австралийские мохообразные . Австралийский национальный ботанический сад . Проверено 22 декабря 2011 г.
- ^ Перейти обратно: а б Ньюби, Адам; Альтланд, Джеймс Э.; Гиллиам, Чарльз Х.; Ветье, Гленн (декабрь 2006 г.). «Борьба с послевсходовой печеночницей в питомниковых культурах, выращенных в контейнерах1» (PDF) . Дж. Энвайрон. Хорт . 24 (4). НИИ садоводства: 230–236. Архивировано из оригинала (PDF) 24 июля 2012 года . Проверено 24 декабря 2011 г.
- ^ Бидартондо, Мартин И.; Дакетт, Джеффри Г. (7 февраля 2010 г.). «Консервативные экологические и эволюционные закономерности в печеночно-грибных симбиозах» . Труды Королевского общества B: Биологические науки . 277 (1680): 485–492. дои : 10.1098/rspb.2009.1458 . ПМЦ 2842645 . ПМИД 19812075 .
- ^ Шустер, Рудольф М. Hepaticae и Anthocerotae Северной Америки , том. Я, стр. 243–249. (Нью-Йорк: издательство Колумбийского университета, 1966).
- ^ Поч, Тамаш. «Тропические лесные мохообразные», с. 59 в AJE Smith (Ed.) Экология мохообразных . (Лондон: Чепмен и Холл, 1982). ISBN 0-412-22340-6 .
- ^ Крэндалл-Стотлер, Барбара. и Стотлер, Раймонд Э. «Морфология и классификация Marchantiophyta». стр. 36–38 в книге А. Джонатана Шоу и Бернарда Гоффине (ред.), Bryophyte Biology . (Кембридж: Издательство Кембриджского университета: 2000). ISBN 0-521-66097-1
- ^ Гоффине, Бернар. «Происхождение и филогенетические связи мохообразных». стр. 124–149 в книге А. Джонатана Шоу и Бернарда Гоффине (ред.), Bryophyte Biology . (Кембридж: Издательство Кембриджского университета:!2000). ISBN 0-521-66097-1
- ^ Кокс, Саймон Дж.; и др. (2014). «Противоречивые филогении ранних наземных растений вызваны предвзятостью состава синонимических замен» . Систематическая биология . 63 (2): 272–279. дои : 10.1093/sysbio/syt109 . ПМЦ 3926305 . ПМИД 24399481 .
- ^ Перейти обратно: а б с Путтик, Марк Н.; и др. (март 2018 г.). «Взаимоотношения наземных растений и природа предкового эмбриофита». Современная биология . 28 (5): 733–745.e2. Бибкод : 2018CBio...28E.733P . дои : 10.1016/j.cub.2018.01.063 . hdl : 1983/ad32d4da-6cb3-4ed6-add2-2415f81b46da . ПМИД 29456145 . S2CID 3269165 .
- ^ Перейти обратно: а б де Соуза, Филипе; и др. (2019). «Филогения ядерных белков поддерживает монофилию трех групп мохообразных (Bryophyta Schimp.)». Новый фитолог . 222 (1): 565–575. дои : 10.1111/nph.15587 . hdl : 1983/0b471d7e-ce54-4681-b791-1da305d9e53b . ПМИД 30411803 . S2CID 53240320 .
- ^ Либенс-Мак, Джеймс Х.; и др. (2019). «Тысяча растительных транскриптомов и филогеномика зеленых растений» . Природа . 574 (7780): 679–685. дои : 10.1038/s41586-019-1693-2 . ПМК 6872490 . PMID 31645766 .
- ^ Чжан, Цзянь; и др. (2020). «Геном роголистника и ранняя эволюция наземных растений» . Природные растения . 6 (2): 107–118. дои : 10.1038/s41477-019-0588-4 . ПМК 7027989 . ПМИД 32042158 .
- ^ Перейти обратно: а б Харрис, Броган Дж.; и др. (2020). «Филогеномные доказательства монофилии мохообразных и редуктивной эволюции устьиц». Современная биология . 30 (11): P2201–2012.E2. Бибкод : 2020CBio...30E2001H . дои : 10.1016/j.cub.2020.03.048 . hdl : 1983/fbf3f371-8085-4e76-9342-e3b326e69edd . ПМИД 32302587 . S2CID 215798377 .
- ^ Ли, Фэй-Вэй; и др. (2020). «Геномы Anthoceros проливают свет на происхождение наземных растений и уникальную биологию роголистников» . Природные растения . 6 (3): 259–272. дои : 10.1038/s41477-020-0618-2 . hdl : 10261/234303 . ПМЦ 8075897 . ПМИД 32170292 .
- ^ Соуза, Филипе; и др. (2020). «Филогения наземных растений хлоропластов: анализ с использованием более подходящих моделей неоднородного состава деревьев и участков» . Границы в науке о растениях . 11 : 1062. дои : 10.3389/fpls.2020.01062 . ПМЦ 7373204 . ПМИД 32760416 .
- ^ Су, Данян; и др. (2021). «Крупномасштабный филогеномный анализ выявляет монофилию мохообразных и неопротерозойское происхождение наземных растений» . Молекулярная биология и эволюция . 38 (8): 3332–3344. дои : 10.1093/molbev/msab106 . ПМЦ 8321542 . ПМИД 33871608 .
- ^ Соуза, Филипе; и др. (2020). «Митохондриальная филогения наземных растений демонстрирует поддержку Setaphyta в рамках моделей гетерогенного замещения по составу» . ПерДж . 8 (4): е8995. дои : 10.7717/peerj.8995 . ПМК 7194085 . ПМИД 32377448 .
- ^ Перейти обратно: а б Кокс, Саймон Дж. (2018). «Молекулярная филогенетика наземных растений: обзор с комментариями по оценке несоответствия между филогениями». Критические обзоры по наукам о растениях . 37 (2–3): 113–127. Бибкод : 2018CRvPS..37..113C . дои : 10.1080/07352689.2018.1482443 . hdl : 10400.1/14557 . S2CID 92198979 .
- ^ Тейлор, Томас Н. и Эдит Л. Тейлор. Биология и эволюция ископаемых растений , с. 139. (Энглвуд Клиффс, Нью-Джерси: Прентис Холл, 1993). ISBN 0-13-651589-4 .
- ^ Остендорп, Кора. Мохообразные палеозоя и мезозоя , стр. 70–71. ( Bryophytum Bibliotheca , Band 34, 1987). ISBN 3-443-62006-X .
- ^ ВанАллер Херник, Л.; Лендинг, Э.; Бартовски, К.Е. (2008). «Самые старые печеночники на Земле - Metzgeriothallus sharonae sp. nov. из среднего девона (живе) восточного Нью-Йорка, США». Обзор палеоботаники и палинологии . 148 (2–4): 154–162. Бибкод : 2008RPaPa.148..154H . дои : 10.1016/j.revpalbo.2007.09.002 .
- ^ Рубинштейн, CV; Джерриенн, П.; Де Ла Пуэнте, GS; Астини, РА; Стиманс, П. (2010). «Свидетельства существования наземных растений в раннем среднем ордовике в Аргентине (восточная Гондвана)» . Новый фитолог . 188 (2): 365–369. дои : 10.1111/j.1469-8137.2010.03433.x . hdl : 11336/55341 . ПМИД 20731783 .
- ^ Крэндалл-Стотлер, Барбара. и Стотлер, Раймонд Э. «Морфология и классификация Marchantiophyta». п. 63 в книге А. Джонатана Шоу и Бернарда Гоффине (ред.), Биология мохообразных . (Кембридж: Издательство Кембриджского университета: 2000). ISBN 0-521-66097-1
- ^ Джонс, EW (2004). Флора печеночников и роголистников Западной Африки . Скрипта Ботника Бельгика. Том. 30. Мейзе: Национальный ботанический сад (Бельгия). п. 30. ISBN 90-72619-61-7 .
- ^ Перейти обратно: а б с Форрест, Лаура Л.; Кристин Э. Дэвис; Дэвид Г. Лонг; Барбара Дж. Крэндалл-Стотлер; Александра Кларк; Мишель Л. Холлингсворт (2006). «Разгадка эволюционной истории печеночников (Marchantiophyta): многочисленные таксоны, геномы и анализ». Бриолог . 109 (3): 303–334. doi : 10.1639/0007-2745(2006)109[303:UTEHOT]2.0.CO;2 . S2CID 85912159 .
- ^ Генрихс, Йохен; С. Робберт Градштейн; Розмари Уилсон; Харальд Шнайдер (2005). «К естественной классификации печеночников (Marchantiophyta) на основе гена хлоропластов rbcL». Криптогамия Bryologie . 26 (2): 131–150.
- ^ Хэ-Нигрэн, Сяолань; Айно Жуслен; Инкери Ахонен; Дэвид Гленни; Блу Пайпер (2006). «Освещение эволюционной истории печеночников (Marchantiophyta) – к естественной классификации» . Кладистика . 22 (1): 1–31. дои : 10.1111/j.1096-0031.2006.00089.x . ПМИД 34892891 . S2CID 86082381 .
- ^ Перейти обратно: а б Рензалья, Карен С.; Скотт Шуэтт; Р. Джоэл Дафф; Роберто Лигроне; А. Джонатан Шоу; Брент Д. Мишлер; Джеффри Г. Дакетт (2007). «Филогения мохообразных: продвижение молекулярных и морфологических границ». Бриолог . 110 (2): 179–213. doi : 10.1639/0007-2745(2007)110[179:BPATMA]2.0.CO;2 . S2CID 85788756 .
- ^ Форрест, Лаура Л.; Барбара Дж. Крэндалл-Стотлер (2004). «Филогения простых слоевищных печеночников (Jungermanniopsida, Metzgeriidae), выведенная на основе пяти генов хлоропластов». Монографии по систематической ботанике . Молекулярная систематика мохообразных. 98 . Издательство Ботанического сада Миссури: 119–140.
- ^ Шустер, Рудольф М. Hepaticae и Anthocerotae Северной Америки , том. VI, с. 26. (Чикаго: Полевой музей естественной истории, 1992). ISBN 0-914868-21-7 .
- ^ Коул, Теодор CH; Хильгер, Хартмут Х.; Гоффине, Бернар. «Плакат по филогении мохообразных: систематика и характеристика несосудистых наземных растений (мхов, печеночников, роголистников)» . 2021 . Проверено 6 декабря 2022 г.
- ^ Сёдерстрем; и др. (2016). «Всемирный контрольный список роголистников и печеночников» . ФитоКлючи (59): 1–826. дои : 10.3897/phytokeys.59.6261 . ПМЦ 4758082 . ПМИД 26929706 .
- ^ Садава, Давид; Дэвид М. Хиллис; Х. Крейг Хеллер; Мэй Беренбаум (2009). Жизнь: наука биологии (9-е изд.). Нью-Йорк: WH Freeman. п. 599. ИСБН 978-1429246446 .
- ^ Сероцка, Я; Козловский, Л.П.; Буйницкий, Дж. М.; Ярмоловский, А; Швейковска-Кулинска, З (2014). «Экспрессия женских генов у двудомного печеночника Pellia endiviifolia регулируется на этапе развития и связана с производством архегониев» . Биология растений BMC . 14 :168. дои : 10.1186/1471-2229-14-168 . ПМК 4074843 . ПМИД 24939387 .
- ^ Диттмер, Ховард Дж. Филогения и формы в царстве растений , с. 286. (Торонто: Д. Ван Ностранд Ко., 1964).
- ^ Рэйвен, PH, RF Эверт и SE Eichhorn. Биология растений , 7-е изд., с. 351. (Нью-Йорк: WH Freeman, 2005). ISBN 0-7167-1007-2 .
- ^ Стерн, Кингсли Р. Введение в биологию растений , 5-е изд., с. 338. (Дубьюк, Айова: Wm. C. Brown Publishers, 1991). ISBN 0-697-09947-4 .
Внешние ссылки
[ редактировать ] СМИ, связанные с Marchantiophyta, на Викискладе?
СМИ, связанные с Marchantiophyta, на Викискладе?  Данные, относящиеся к Marchantiophyta , в Wikispecies
Данные, относящиеся к Marchantiophyta , в Wikispecies - Строение печеночника в картинках
- LiToL: Сборка печеночникового Древа Жизни (примечание: вместо «500 000 миллионов лет назад» читайте «480 миллионов лет назад»).
- Взаимоотношения мхов, печеночников и роголистников
- Дополнительная информация о печеночниках
- Печеночники
Классификация Archaeplastida или Plantae sl |
|---|
| Базы данных органов управления : Национальные |
|---|