ДНК -адениновая метилаза
| Специфичная для сайта ДНК-метилтрансфераза (аденин-специфическая) | |||
|---|---|---|---|
| Идентификаторы | |||
| ЕС №. | 2.1.1.72 | ||
| CAS №. | 69553-52-2 | ||
| Базы данных | |||
| Intenz | Intenz View | ||
| Бренда | Бренда вход | ||
| Расширение | Вид Nicezyme | ||
| Кегг | Кегг вход | ||
| Метатический | Метаболический путь | ||
| Напрямую | профиль | ||
| PDB Структуры | RCSB PDB PDBE PDBSUM | ||
| |||
ДНК -адениновая метилаза , (плотина) [ 1 ] (Также сайт-специфическая ДНК-метилтрансфераза (аденин-специфическая) , EC 2.1.1.72 , модификация метилаза , система ограничения )-это фермент , который добавляет метильную группу в аденин последовательности 5'-GATC-3 синтезированная ДНК . [ 2 ] [ 3 ] Сразу после синтеза ДНК дочь в течение короткого времени остается неметилированной. [ 4 ] Это сирот метилтрансфераза, которая не является частью системы ограничения модификации и регулирует экспрессию генов. [ 5 ] [ 6 ] [ 7 ] [ 8 ] Этот фермент катализирует следующую химическую реакцию
- S-Аденозил-L-метионин + ДНК аденин S-аденозил-L-гомоцистеин + ДНК 6-метиламинопурин

Это большая группа ферментов, уникальных для прокариот и бактериофагов. [ 9 ]
Э. Коли ДНК-аденин-метилтрансфераза фермент (плотина) широко используется для метода профилирования хроматина DAMID , в которой плотина сливается с ДНК-связывающим белком и экспрессируется в качестве трансгена в генетически действующем модели организме для идентификации белка сайты связывания. [ 10 ]
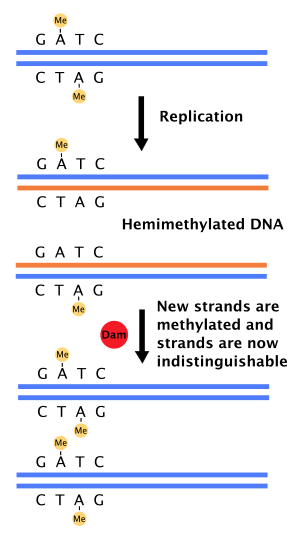
Роль в восстановлении несоответствия ДНК
[ редактировать ]Когда ДНК-полимераза допускает ошибку, приводящую к несоответствующей базовой паре или небольшой вставке или делеции во время синтеза ДНК , клетка будет восстанавливать ДНК путем, называемого восстановлением несоответствия . Тем не менее, ячейка должна быть в состоянии различать нити шаблона и вновь синтезированную прядь. У некоторых бактерий цепи ДНК метилируются метилазой плотины, и, следовательно, сразу после репликации ДНК будет гемиметилизирована. [ 4 ] Репарационный фермент, MUTS , связывается с несоответствиями в ДНК и рекрутирует MUTL, который впоследствии активирует эндонуклеазу MUTH. MUTH связывает гемиметилированные сайты GATC и при активации будет избирательно расщеплять неметилированную дочернюю прядь, позволяя геликазе и экзонуклеазам активировать зарождающуюся прядь в области, окружающей несоответствие. [ 4 ] [ 11 ] Затем прядь повторно синтезируется ДНК-полимеразой III .
Роль в регуляции репликации
[ редактировать ]Выстрел происхождения репликации (ORIC) в клетках бактерий высоко контролируется, чтобы гарантировать, что репликация ДНК происходит только один раз во время каждого деления клеток. Часть этого может быть объяснена медленным гидролизом АТФ с помощью ДНКа, белка, который связывается с повторами в ORIC, чтобы инициировать репликацию. Метилаза плотины также играет роль, потому что ORIC имеет 11 5'-GATC-3 'последовательностей (в E. coli ). Сразу после репликации ДНК ORIC гемиметилизируется и изолируется в течение определенного периода времени. Только после этого ORIC высвобождается и должен полностью метилироваться метилазой DAM до того, как произошло связывание DNAA.
Роль в регуляции экспрессии белка
[ редактировать ]Дамба также играет роль в продвижении и репрессии транскрипции РНК . В E. coli вниз по течению GATC последовательности являются метилированными , способствуя транскрипции. Например, пиелонефритом связанное с PILI (PAP), изменение фазы , в уропатогенной кишечной палочке контролируется плотиной посредством метилирования двух сайтов GATC, проксимальных и дистальных по отношению к промотору PAP . [ 12 ] Учитывая его роль регуляции белка в кишечной палочке , ген метилазы плотины является несущественным, поскольку нокаут гена все еще оставляет бактерии жизнеспособными. [ 13 ] Поддержание жизнеспособности, несмотря на нокаут гена плотины, также наблюдается у сальмонеллы и агрегатибактерных актиномицеемкометров . [ 14 ] [ 15 ] Тем не менее, у организмов, таких как Vibrio cholerae и Yersinia pseudotuberculosis , ген плотины необходим для жизнеспособности. [ 16 ] Нокаут гена плотины в агрегатибактерных актиномицеемкомитанах приводил к нарушению регулируемого уровня белка, лейкотоксина, а также снижал способность микроба вторгаться в пероральные эпителиальные клетки. [ 15 ] Кроме того, исследование по дефициту метилазы плотины Streptococcus mutans , стоматологического патогена, выявило дисрегуляцию 103 генов, некоторые из которых включают кариогенный потенциал. [ 16 ]
Структурные особенности
[ редактировать ]Сходство в каталитических доменах C5-цитозин метилтрансфераз и N6 и N4-аденин-метилтрансферазы обеспечило большой интерес к пониманию основы для функциональных сходств и различий. Метилтрансферазы или метилазы классифицируются на три группы (группы α, β и γ) на основе последовательного порядка определенных 9 мотивов и домена распознавания цели (TRD). [ 17 ] Мотив I состоит из трипептида Gly-X-Gly и называется G-петлей и участвует в связывании S-аденозилметионинового кофактора. [ 18 ] Мотив II высоко консервативен среди метилаз N4 и N6-аденина и содержит отрицательно заряженную аминокислоту, за которой следует гидрофобная боковая цепь в последних положениях цепи β2, чтобы связывать адомету . [ 17 ] Мотив III также вовлечен в связывание Adomet. Мотив IV особенно важен и хорошо известен в характеристиках метилазы. Он состоит из дипролильного компонента и высоко консервативен среди N6-адениновых метилтрансфераз в качестве мотива DPPY, однако этот мотив может варьироваться для N4-аденина и C5-цитозин метилтрансферазы. Было установлено, что мотив DPPY имеет важное значение для связывания Adomet. [ 19 ] Мотивы IV-VIII играют роль в каталитической активности, в то время как мотивы 1-III и X играют роль в связывании кофактора. Для N6 -адениновых метилаз последовательный порядок для этих мотивов как таковой: N -концевой - x - I - II - III - TRD - IV - V - VI - VII - VIII - C -концевой E. и Колиме метилаза следует этой структурной последовательности. [ 17 ] Эксперимент по кристаллографии 2015 года показал, что e . Метилаза Coli Dam была способна связывать не GATC ДНК с той же последовательности обсуждаемых мотивов; Авторы утверждают, что полученная структура может служить основанием для репрессии транскрипции, которая не основана на метилировании. [ 20 ]
Бактериальные бактериальные и бактериофаги метилазы
[ редактировать ]Метилаза плотины представляет собой сирот метилтрансфераза, которая не является частью системы ограничения ограничения, но работает независимо для регулирования экспрессии генов, восстановления несоответствия и репликации бактерий среди многих других функций. Это не единственный пример орфан-метилтрансферазы, поскольку существует клеточный цикл, регулируемая метилтрансфераза (CCRM), которая метилат 5'-г-г-г-гемиметилированной геми-метилированной ДНК для контроля жизненного цикла Caulobacter Crescentus и других родственных видов. [ 21 ]
В отличие от их бактериальных аналогов, существуют также существуют фаговые метилтрансферазы и, в частности, в T2, T4 и других бактериофагах T-Even, которые заражают E. coli. [ 5 ] В исследовании было идентифицировано, что, несмотря на то, что аминокислотные последовательности E. coli и плотина T4 Methylase Amino Acdes имеют идентичность последовательности до 64% в четырех областях от 11 до 33 остатков, что предполагает общее эволюционное происхождение для бактерий и гены фаговой метилазы. [ 22 ] Метилазы T2 и T4 отличаются от метилазы плотины E. coli не только от их способности метилировать 5-гидроксиметилцитозин, но и метилировать неканонические ДНК-сайты. Несмотря на обширную характеристику in vitro этих избранных нескольких фага -сирот метилтрансферазы, их биологическая цель до сих пор не ясна. [ 5 ]
Смотрите также
[ редактировать ]Ссылки
[ редактировать ]- ^ Браун, Теренс (2002). «Глава 14: Мутация, восстановление и рекомбинация. Раздел 2.3». Геномы . Гарлендская наука. ISBN 0-471-25046-5 .
- ^ Marinus MG, Morris NR (июнь 1973 г.). «Выделение дезоксирибонуклеиновой кислоты-метилазы мутантов Escherichia coli K-12» . Журнал бактериологии . 114 (3): 1143–50. doi : 10.1128/jb.114.3.1143-1150.1973 . PMC 285375 . PMID 4576399 .
- ^ Geier GE, Modrich P (февраль 1979 г.). «Последовательность распознавания метилазы плотины Escherichia coli K12 и способ расщепления эндонуклеазы DPN I». Журнал биологической химии . 254 (4): 1408–13. PMID 368070 .
- ^ Jump up to: а беременный в Баррас Ф., Маринус М.Г. (1989). «Великий GATC: метилирование ДНК в E. coli». Тенденции в генетике . 5 (5): 139–143. doi : 10.1016/0168-9525 (89) 90054-1 . PMID 2667217 .
- ^ Jump up to: а беременный в Мерфи Дж., Махони Дж., Эйнсворт С., Наута А., Ван Синдерен Д. (декабрь 2013 г.). «Бактериофаг -сирот ДНК метилтрансферазы: понимание их бактериального происхождения, функции и возникновения» . Прикладная и экологическая микробиология . 79 (24): 7547–55. doi : 10.1128/aem.02229-13 . PMC 3837797 . PMID 24123737 .
- ^ Кесслер С., Манта В (август 1990 г.). «Специфичность рестрикции эндонуклеазы и модификации ДНК метилтрансферазирует обзор (издание 3)». Ген . 92 (1–2): 1–248. doi : 10.1016/0378-1119 (90) 90486-b . PMID 2172084 .
- ^ Робертс Р.Дж. (апрель 1990 г.). «Ограничительные ферменты и их изосхизомеры» . Исследование нуклеиновых кислот . 18 Suppl: 2331–65. doi : 10.1093/nar/18.suppl.2331 . PMC 331877 . PMID 2159140 .
- ^ Юань Р (1981). «Структура и механизм многофункциональных эндонуклеаз ограничений». Ежегодный обзор биохимии . 50 : 285–319. doi : 10.1146/annurev.bi.50.070181.001441 . PMID 6267988 .
- ^ Робертс RJ, Macelis D (Eds.). «База данных рестрикционных ферментов» . Рефейс . Получено 22 февраля 2020 года .
- ^ Aughey GN, Southall TD (январь 2016 г.). «Плотина, это хорошо! Дамидное профилирование взаимодействия белка-ДНК» . Wiley междисциплинарные обзоры. Биология развития . 5 (1): 25–37. doi : 10.1002/wdev.205 . PMC 4737221 . PMID 26383089 .
- ^ Løbner-Olesen A, Skovgaard O, Marinus MG (апрель 2005 г.). «Метилирование плотины: координация клеточных процессов». Текущее мнение о микробиологии . 8 (2): 154–60. doi : 10.1016/j.mib.2005.02.009 . PMID 15802246 .
- ^ Casadesús J, Low D (сентябрь 2006 г.). «Эпигенетическая регуляция генов в бактериальном мире» . Микробиология и молекулярная биология обзоры . 70 (3): 830–56. doi : 10.1128/mmbr.00016-06 . PMC 1594586 . PMID 16959970 .
- ^ Bale A, D'Alarcao M, Marinus MG (февраль 1979 г.). «Характеристика метилирования ДНК -метилирования Escherichia coli K12». Мутационные исследования . 59 (2): 157–65. doi : 10.1016/0027-5107 (79) 90153-2 . PMID 375073 .
- ^ Николсон Б., Лоу Д (февраль 2000 г.). «ДНК-метилирование-зависимая регуляция экспрессии PEF в сальмонелле Typhimurium». Молекулярная микробиология . 35 (4): 728–42. doi : 10.1046/j.1365-2958.2000.01743.x . PMID 10692151 .
- ^ Jump up to: а беременный Wu H, Lippmann JE, Oza JP, Zeng M, Fives-Taylor P, Reich No (август 2006 г.). «Инактивация ДНК -аденинметилтрансферазы изменяет факторы вирулентности в актинобацилле актиномицеемкомитанов». Пероральная микробиология и иммунология . 21 (4): 238–44. doi : 10.1111/j.1399-302x.2006.00284.x . PMID 16842508 .
- ^ Jump up to: а беременный Хулио С.М., Хейтофф Д.М., Провенцано Д., Клозе К.Е., Синшаймер Р.Л., Лоу Д.А., Махан М.Дж. (декабрь 2001 г.). «ДНК -аденин -метилаза необходима для жизнеспособности и играет роль в патогенезе псевдотуберкулеза иерсинии и холеры вибриона» . Инфекция и иммунитет . 69 (12): 7610–5. doi : 10.1128/iai.69.12.7610-7615.2001 . PMC 98854 . PMID 11705940 .
- ^ Jump up to: а беременный в Мэлоун Т., Блюменталь Р.М., Ченг Х (ноябрь 1995 г.). «Анализ с структурой выявляет девять мотивов последовательности, консервативные среди аминометилтрансфераз ДНК, и предполагает каталитический механизм для этих ферментов». Журнал молекулярной биологии . 253 (4): 618–32. doi : 10.1006/jmbi.1995.0577 . PMID 7473738 .
- ^ Schluckebier G, O'gara M, Saenger W, Cheng X (март 1995 г.). «Универсальная структура каталитического домена адометозависимых метилтрансфераз». Журнал молекулярной биологии . 247 (1): 16–20. doi : 10.1006/jmbi.1994.0117 . PMID 7897657 .
- ^ Kossykh VG, Schlagman SL, Hattman S (июль 1993 г.). «Консервативный мотив последовательности DPPY в области IV ДНК плотины фага T4 [N-Аденин] -метилтрансфераза важен для связывания S-аденозил-L-метионина» . Исследование нуклеиновых кислот . 21 (15): 3563–6. doi : 10.1093/nar/21.15.3563 . PMC 331459 . PMID 16617501 .
- ^ Horton JR, Zhang X, Blumenthal RM, Cheng X (апрель 2015 г.). «Структуры Escherichia coli ДНК-метилтрансферазы (DAM) в комплексе с не GATC-последовательности: потенциальные последствия для метилирования-независимой репрессии транскрипции» . Исследование нуклеиновых кислот . 43 (8): 4296–308. doi : 10.1093/nar/gkv251 . PMC 4417163 . PMID 25845600 .
- ^ Zweiger G, Marczynski G, Shapiro L (январь 1994). «Калобактерная ДНК метилтрансфераза, которая функционирует только в предсказательной клетке». Журнал молекулярной биологии . 235 (2): 472–85. doi : 10.1006/jmbi.1994.1007 . PMID 8289276 .
- ^ Хаттман С., Уилкинсон Дж., Суинтон Д., Шлагман С., Макдональд П.М., Мосиг Г. (ноябрь 1985). «Общее эволюционное происхождение плотины фага T4 и хозяина генов DNA-адениновой метилтрансферазы Escherichia coli» . Журнал бактериологии . 164 (2): 932–7. doi : 10.1128/jb.164.2.932-937.1985 . PMC 214344 . PMID 3902803 .