Ауксин

В растениях имеется еще четыре эндогенно синтезируемых ауксина. [1] [2]
Все ауксины представляют собой соединения с ароматическим кольцом и группой карбоновой кислоты: [2] [3]

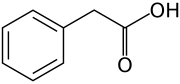


Ауксины ( множественное число от ауксина / ˈ ɔː k s ɪ n / ) представляют собой класс растительных гормонов (или регуляторов роста растений) с некоторыми морфогеноподобными характеристиками. Ауксины играют кардинальную роль в координации многих процессов роста и поведения в жизненных циклах растений и необходимы для развития организма растений. Голландский биолог Фриц Вармолт Вент впервые описал ауксины и их роль в росте растений в 1920-х годах. [4] Кеннет В. Тиманн стал первым, кто выделил один из этих фитогормонов и определил его химическую структуру как индол-3-уксусную кислоту (ИУК). Вент и Тиманн в 1937 году стали соавторами книги о растительных гормонах « Фитогормоны» .
Обзор [ править ]
Ауксины были первыми из основных растительных гормонов, которые были открыты. Свое название они получили от греческого слова αυξειν ( ауксеин – «расти/увеличиваться»). Ауксин присутствует во всех частях растения, хотя и в очень разных концентрациях. Концентрация в каждой позиции является важной информацией для развития, поэтому она подлежит жесткой регуляции посредством метаболизма и транспорта. В результате ауксин создает «паттерны» максимумов и минимумов концентрации ауксина в организме растения, которые, в свою очередь, направляют дальнейшее развитие соответствующих клеток и, в конечном итоге, растения в целом.
распределения ауксина внутри растения (динамический и чувствительный к окружающей среде) Характер является ключевым фактором для роста растения, его реакции на окружающую среду и, в частности, для развития органов растения. [5] [6] (например, листья или цветы ). Это достигается за счет очень сложного и слаженного активного транспорта молекул ауксина от клетки к клетке по всему растительному организму — путем так называемого полярного транспорта ауксина . [5] Таким образом, растение может (в целом) реагировать на внешние условия и приспосабливаться к ним, не требуя при этом нервной системы . Ауксины обычно действуют совместно с другими растительными гормонами или противоположно им. Например, соотношение ауксина и цитокинина в определенных тканях растения определяет зарождение корневых и побеговых почек.
На молекулярном уровне все ауксины представляют собой соединения с ароматическим кольцом и группой карбоновой кислоты . [3] Наиболее важным членом семейства ауксинов является индол-3-уксусная кислота (ИУК). [7] который вызывает большую часть эффектов ауксина в интактных растениях и является наиболее мощным природным ауксином. И что касается нативного ауксина, его равновесие в растениях контролируется многими способами: от синтеза через возможную конъюгацию до деградации его молекул, всегда в соответствии с требованиями ситуации.
- Пять встречающихся в природе (эндогенных) ауксинов в растениях включают индол-3-уксусную кислоту, 4-хлориндол-3-уксусную кислоту , фенилуксусную кислоту , индол-3-масляную кислоту и индол-3-пропионовую кислоту . [1] [2] Однако большая часть знаний, описанных до сих пор в области биологии ауксина и описанных в последующих разделах, в основном применима к ИУК; остальные три эндогенных ауксина, по-видимому, имеют незначительное значение для интактных растений в естественной среде. Помимо эндогенных ауксинов, ученые и производители разработали множество синтетических соединений, обладающих ауксиновой активностью.
- Синтетические ауксины делятся на четыре класса: [8] [1]
- дикамба
- хинолинкарбоновые кислоты, в состав которых входит хинклорак
- производные пиридинкарбоновых кислот , к которым относятся пиклорам , триклопир , клопиралид
- феноксиуксусная кислота , феноксипропионовая кислота и феноксимасляная кислота, 1-нафталинуксусной кислоты производные 2,4-Д , 2,4-ДП 2,4-ДБ 2,4,5-Т MCPA MCPB , мекопроп
Некоторые синтетические ауксины, такие как 2,4-Д и 2,4,5-трихлорфеноксиуксусная кислота (2,4,5-Т), продаются как гербициды . Широколистные растения ( двудольные ), такие как одуванчики , гораздо более восприимчивы к ауксинам, чем узколистные растения ( однодольные ), такие как травы и зерновые культуры, что делает эти синтетические ауксины ценными в качестве гербицидов.
Открытие [ править ]
Чарльз Дарвин [ править ]
В 1881 году Чарльз Дарвин и его сын Фрэнсис провели эксперименты на колеоптилях — оболочках, покрывающих молодые листья прорастающих сеянцев травы. В ходе эксперимента колеоптиль подвергся воздействию света от однонаправленного источника и заметил, что они наклоняются в сторону света. [9] Покрыв различные части колеоптилей светонепроницаемым непрозрачным колпачком, Дарвины обнаружили, что свет воспринимается кончиком колеоптиля, но изгиб происходит в гипокотиле . Однако сеянцы не проявляли признаков развития к свету, если кончик был закрыт непрозрачным колпачком или если кончик был удален. Дарвины пришли к выводу, что кончик колеоптиля отвечает за восприятие света, и предположили, что посланник передается в нисходящем направлении от кончика колеоптиля, заставляя его изгибаться. [10]
Питер Бойсен Дженсен [ править ]
В 1910 году датский учёный Питер Бойсен Йенсен продемонстрировал, что фототропный стимул в овса колеоптиле может распространяться через разрез . [11] Эти эксперименты были расширены и опубликованы более подробно в 1911 и 1913 годах. [12] [13] Он обнаружил, что кончик можно отрезать и надеть обратно, а последующее одностороннее освещение все еще способно вызвать положительную фототропную кривизну в базальной части колеоптиля. Он продемонстрировал, что передача может происходить через тонкий слой желатина , отделяющий односторонне освещенный кончик от затененной культи. Вставив кусок слюды, он мог заблокировать передачу на освещенной и неосвещенной стороне иглы соответственно, что позволило ему показать, что передача происходила в затененной части иглы. Таким образом, тканью, получавшей стимул роста, была продольная половина колеоптиля, демонстрирующая большую скорость удлинения при фототропном искривлении. [14] [15]
В 1911 году Бойсен-Йенсен на основании результатов своих экспериментов пришел к выводу, что передача фототропного стимула была не физическим эффектом (например, из-за изменения давления), а результатом миграции вещества или ионов (была вызвана переносом вещества или ионов). [12] Эти результаты имели основополагающее значение для дальнейших работ по ауксиновой теории тропизмов .
Фриц Ушел [ править ]
В 1928 году голландский ботаник Фриц Вармолт Вент показал, что химический посланник диффундирует из кончиков колеоптилей. Эксперимент Вента выявил, как химическое вещество, способствующее росту, заставляет колеоптиль расти в направлении света. Он срезал кончики колеоптилей и поместил их в темноту, поместив несколько кончиков на блоки агара, которые, как он предсказал, впитают химическое вещество, способствующее росту. На контрольных колеоптилях он поместил блок, в котором отсутствовало химическое вещество. На других он размещал блоки, содержащие химическое вещество, либо по центру верхней части колеоптиля, чтобы распределить химическое вещество равномерно, либо смещенно, чтобы увеличить концентрацию с одной стороны. [10]
Когда химическое вещество, способствующее росту, было распределено равномерно, колеоптиль рос прямо. Если химическое вещество распределялось неравномерно, колеоптиль выгибался от стороны куба, как бы растя к свету, хотя и рос в темноте. Позже Вент предположил, что веществом-мессенджером является гормон, способствующий росту, который он назвал ауксином, который асимметрично распределяется в области сгибания. Вент пришел к выводу, что ауксин находится в более высокой концентрации на затененной стороне, что способствует удлинению клеток, что приводит к изгибу колеоптилей к свету. [15]
Гормональная активность [ править ]
Ауксины помогают развитие на всех уровнях растения, от клеточного уровня, через органы и, в конечном итоге, до всего растения.
Молекулярные механизмы [ править ]
Когда растительная клетка вступает в контакт с ауксином, это вызывает резкие изменения в экспрессии генов , при этом многие гены активируются или подавляются. Точные механизмы, с помощью которых это происходит, все еще являются областью активных исследований, но в настоящее время существует общий консенсус по крайней мере о двух путях передачи сигналов ауксина. [16] [17]
Восприятие [ править ]
семейства TIR1/AFB Наиболее изученными рецепторами ауксина являются белки F-бокса . Белки F-box нацелены на деградацию других белков по пути деградации убиквитина . Когда белки TIR1/AFB связываются с ауксином, молекула ауксина действует как « молекулярный клей » (термин, придуманный Нин Чжэном ), который позволяет этим белкам затем связываться со своими мишенями (см. ниже). Атомная структура механизма восприятия ауксина TIR1 была определена методом рентгеновской кристаллографии . [18]
Другой ауксин-связывающий белок, ABP1, в настоящее время часто рассматривается как рецептор ауксина (в апопласте ), но обычно считается, что он играет гораздо более второстепенную роль, чем сигнальный путь TIR1/AFB, и о передаче сигналов ABP1 известно гораздо меньше. [17]
Модули сигнализации Aux/IAA и ARF [ править ]

Факторы ответа на ауксин (ARF) представляют собой большую группу факторов транскрипции , которые участвуют в передаче сигналов ауксина. В отсутствие ауксина ARF связываются с классом репрессоров, известных как Aux/IAA. Aux/IAA подавляют способность ARF усиливать транскрипцию генов. Кроме того, связывание Aux/IAA с ARF приводит к контакту Aux/IAA с промоторами генов, регулируемых ауксином. Находясь на этих промоторах, Aux/IAA подавляют экспрессию этих генов за счет привлечения других факторов для внесения модификаций в структуру ДНК .
Связывание ауксина с TIR1/AFB позволяет им связываться с Aux/IAA. При связывании TIR1/AFB Aux/IAA помечаются как подлежащие деградации. Деградация Aux/IAA высвобождает белки ARF, которые затем способны активировать или репрессировать гены, с промоторами которых они связаны. [16] [17]
Считается, что большое количество возможных пар связывания Aux/IAA и ARF и их различное распределение между типами клеток и возрастом развития объясняют удивительно разнообразные ответы, которые вызывает ауксин.
В июне 2018 года было продемонстрировано, что растительные ткани могут чрезвычайно быстро реагировать на ауксин TIR1-зависимым образом (вероятно, слишком быстро, чтобы это можно было объяснить изменениями в экспрессии генов). Это побудило некоторых ученых предположить, что существует еще неидентифицированный TIR1-зависимый путь передачи сигналов ауксина, который отличается от хорошо известного транскрипционного ответа. [19]
На клеточном уровне [ править ]
На клеточном уровне ауксин необходим для роста клеток , влияя как на деление клеток , так и на их расширение. Уровень концентрации ауксина вместе с другими местными факторами способствует дифференцировке клеток и спецификации клеточной судьбы.
В зависимости от конкретной ткани ауксин может способствовать осевому удлинению (как в побегах), латеральному расширению (как при набухании корня) или изодиаметрическому расширению (как при росте плодов). В некоторых случаях (рост колеоптилей) клеточная экспансия, стимулируемая ауксином, происходит при отсутствии клеточного деления. В других случаях стимулируемое ауксином деление клеток и размножение клеток могут быть тесно секвенированы в одной и той же ткани (зарождение корней, рост плодов). В живом растении ауксины и другие растительные гормоны почти всегда взаимодействуют, определяя закономерности развития растений.
Органные паттерны [ править ]
Совместный рост и деление растительных клеток приводит к росту тканей , а специфический рост тканей способствует развитию органов растения .

Рост клеток способствует увеличению размеров растения, неравномерно локализованный рост вызывает изгиб, поворот и направленность органов — например, стебли поворачиваются к источникам света ( фототропизм ), корни растут в ответ на силу тяжести ( гравитропизм ) и другие тропизмы возникли потому, что клетки одна сторона растет быстрее, чем клетки на другой стороне органа. Таким образом, точный контроль распределения ауксина между различными клетками имеет первостепенное значение для конечной формы роста и организации растений.
ауксина и неравномерное распределение Транспорт ауксина
Чтобы вызвать рост в необходимых доменах, ауксины обязательно должны быть активны преимущественно в них. Локальные максимумы ауксина могут образовываться путем активного биосинтеза в определенных клетках тканей, например, по триптофан-зависимым путям. [20] но ауксины синтезируются не во всех клетках (даже если клетки сохраняют потенциальную способность к этому, только при определенных условиях в них активируется синтез ауксинов). Для этой цели ауксины должны быть не только транслоцированы в те сайты, где они необходимы, но также у них должен быть установленный механизм обнаружения этих сайтов. Транслокация осуществляется по всему телу растения, в первую очередь от вершин побегов к вершинам корней (сверху вниз).
На большие расстояния перемещение происходит посредством потока жидкости в сосудах флоэмы , но для транспорта на короткие расстояния используется уникальная система скоординированного полярного транспорта непосредственно от клетки к клетке. Этот активный транспорт на короткие расстояния проявляет некоторые морфогенетические свойства.
Этот процесс, полярный транспорт ауксина , является направленным, очень строго регулируемым и основан на неравномерном распределении переносчиков оттока ауксина на плазматической мембране, которые направляют ауксины в нужном направлении. Хотя PIN-FORMED (PIN) белки жизненно важны для полярной транспортировки ауксина, [6] [21] семейство генов AUXIN1/LIKE-AUX1 (AUX/LAX) кодирует носители неполярного притока ауксина. [22]
Регуляция локализации белка PIN в клетке определяет направление транспорта ауксина из клетки, а концентрированные усилия многих клеток создают пики ауксина, или ауксиновые максимумы (области, в которых имеются клетки с более высоким содержанием ауксина – максимум). [6] Правильные и своевременные максимумы ауксина в развивающихся корнях и побегах необходимы для организации развития органа. [5] [23] [24] PINs регулируются несколькими путями как на транскрипционном, так и на посттрансляционном уровнях. Белки PIN могут фосфорилироваться с помощью PINOID, что определяет их апикобазальную полярность и, тем самым, направленность потоков ауксина. Кроме того, другие киназы AGC, такие как D6PK, фосфорилируют и активируют переносчики PIN. Киназы AGC, включая PINOID и D6PK, нацелены на плазматическую мембрану посредством связывания с фосфолипидами. Выше D6PK 3'-фосфоинозитид-зависимая протеинкиназа 1 (PDK1) действует как главный регулятор. PDK1 фосфорилирует и активирует D6PK на базальной стороне плазматической мембраны, осуществляя деятельность PIN-опосредованного полярного транспорта ауксина и последующего развития растений. [25] Окружающие максимумы ауксина представляют собой клетки с низкими впадинами ауксина или минимумами ауксина. Например, было показано, что в плодах арабидопсиса минимум ауксина важен для развития его тканей. [26]
Ауксин оказывает существенное влияние на пространственную и временную экспрессию генов во время роста апикальных меристем. Эти взаимодействия зависят как от концентрации ауксина, так и от пространственной ориентации во время первичного позиционирования. Ауксин опирается на PIN1, который работает как переносчик оттока ауксина. Позиционирование PIN1 на мембранах определяет направление потока гормона от более высоких концентраций к более низким. [27] Зарождение зачатков в апикальных меристемах коррелирует с повышенным уровнем ауксина. [24] Гены, необходимые для определения идентичности клеток, располагаются и экспрессируются в зависимости от уровня ауксина. STM (SHOOT MERISTEMLESS), который помогает поддерживать недифференцированные клетки, снижается в присутствии ауксина. Это позволяет растущим клеткам дифференцироваться в различные растительные ткани. Гены CUC (Чашевидная семядоля) устанавливают границы для роста тканей и способствуют росту. [28] Их регуляция активируется за счет притока ауксина. [24] Эксперименты с использованием визуализации GFP (ЗЕЛЕНЫЙ ФЛУОРЕСЦЕНЦИОННЫЙ БЕЛОК) у арабидопсиса подтвердили эти утверждения.
Организация завода [ править ]
Поскольку ауксины способствуют формированию органов, [5] [6] они также принципиально необходимы для правильного развития самого растения. [5] Без гормональной регуляции и организации растения были бы просто размножающимися кучами одинаковых клеток. Использование ауксина начинается в зародыше растения, где направленное распределение ауксина способствует последующему росту и развитию первичных полюсов роста, а затем формирует почки будущих органов. Затем он помогает координировать правильное развитие возникающих органов, таких как корни, семядоли и листья, и передает сигналы на большие расстояния между ними, способствуя тем самым общей архитектуре растения. [5] На протяжении всей жизни растения ауксин помогает растению поддерживать полярность роста. [5] и фактически «узнать», где соединены его ветви (или какой-либо орган).
Важным принципом организации растений, основанным на распределении ауксина, является апикальное доминирование , что означает, что ауксин, вырабатываемый верхушечной почкой (или растущей верхушкой), диффундирует (и транспортируется) вниз и подавляет развитие задних боковых почек, которые в противном случае конкурировали бы с апикальный кончик для света и питательных веществ. Удаление верхушечного кончика и его подавляющего действия ауксина позволяет развиваться нижним спящим боковым почкам, а почки между стеблем листа и стеблем дают новые побеги, которые конкурируют за право стать ведущим ростом. На самом деле этот процесс довольно сложен, поскольку для достижения этого явления ауксин, транспортируемый вниз от верхушки свинцового побега, должен взаимодействовать с несколькими другими растительными гормонами (такими как стриголактоны или цитокинины ) в различных положениях вдоль оси роста в теле растения. Такое поведение растений используется при обрезке садоводами .
Наконец, на степень роста корней влияет сумма ауксинов, поступающих от стеблей к корням. Если кончики побегов удалить, растение не реагирует только ростом боковых почек, которые должны заменить исходный свинец. Отсюда также следует, что меньшее количество ауксина, поступающего к корням, приводит к более медленному росту корней, а питательные вещества впоследствии в большей степени передаются в верхнюю часть растения, которая, следовательно, начинает расти быстрее.
Эффекты [ править ]


Ауксин участвует в фототропизме , геотропизме , гидротропизме и других изменениях развития. Неравномерное распределение ауксина из-за факторов окружающей среды, таких как однонаправленный свет или сила тяжести, приводит к неравномерному росту тканей растения, и, как правило, ауксин определяет форму и форму тела растения, направление и силу роста всех органов и их взаимное взаимодействие. [6] Когда клетки увеличиваются в размерах, их объем увеличивается по мере увеличения внутриклеточной концентрации растворенных веществ по мере поступления воды в клетки из внеклеточной жидкости. Это стимулируемое ауксином потребление воды вызывает тургорное давление на клеточные стенки, заставляя растение изгибаться.
Ауксин стимулирует удлинение клеток, стимулируя факторы, способствующие разрыхлению стенок, такие как экспансины , которые ослабляют клеточные стенки . Эффект сильнее, если гиббереллины присутствуют еще и . Ауксин также стимулирует деление клеток, если цитокинины присутствуют . Когда ауксин и цитокинин наносятся на каллус , укоренение может быть получено с более высоким соотношением ауксина к цитокинину, рост побегов индуцируется более низким соотношением ауксина к цитокинину, а каллус формируется с промежуточными соотношениями, при этом точные пороговые соотношения зависят от вида и исходная ткань.Ауксин также вызывает накопление сахара и минералов в месте применения.
Реакция на рану [ править ]
Ауксин индуцирует образование и организацию флоэмы и ксилемы . При повреждении растения ауксин может индуцировать дифференцировку клеток и регенерацию сосудистых тканей. [29]
Рост и развитие корней [ править ]
Ауксины способствуют закладыванию корней. [30] Ауксин индуцирует как рост уже существующих корней, так и ветвление корней (зарождение бокового корня), а также образование придаточных корней. Поскольку больше нативного ауксина транспортируется вниз по стеблю к корням, стимулируется общее развитие корней. Если источник ауксина удален, например, путем обрезки кончиков стеблей, корни соответственно меньше стимулируются, и вместо этого поддерживается рост стебля.
В садоводстве ауксины, особенно NAA и IBA , широко применяются для стимуляции закладывания корней при укоренении черенков растений. Однако высокие концентрации ауксина подавляют удлинение корня и вместо этого усиливают образование придаточных корней. Удаление кончика корня может привести к торможению образования вторичных корней.
доминирование Апикальное
Ауксин индуцирует доминирование верхушки побега ; пазушные почки ингибируются ауксином, поскольку высокая концентрация ауксина непосредственно стимулирует синтез этилена в пазушных почках, вызывая торможение их роста и усиление апикального доминирования. При удалении верхушки растения угнетающее действие снимается и рост боковых почек усиливается. Это называется обезглавливанием, которое обычно проводится на чайных плантациях и при строительстве живых изгородей. Ауксин направляется в часть растения, обращенную от света, где он способствует удлинению клеток, заставляя растение наклоняться к свету. [31]
Рост и развитие плодов [ править ]
Ауксин необходим для роста и развития плодов и замедляет их старение . При удалении семян из клубники рост плодов прекращается; экзогенный ауксин стимулирует рост плодов с удаленными семенами. Для плодов с неоплодотворенными семенами экзогенный ауксин приводит к партенокарпии («росту нетронутых плодов»).
Плоды образуют аномальную морфологию, когда нарушается транспорт ауксина. [32] В плодах арабидопсиса ауксин контролирует выход семян из плода (стручка). Края клапана представляют собой специализированную ткань в капсулах, которая регулирует время открытия капсулы (раскрытие). Ауксин необходимо удалить из клеток края клапана, чтобы края клапана могли сформироваться. Этот процесс требует модификации переносчиков ауксина (белков PIN). [26]
Эволюционный переход от диплоидного к триплоидному эндосперма - и образование антиподальных клеток - мог произойти из-за сдвига в развитии гаметофитов , который вызвал новое взаимодействие с ауксин-зависимым механизмом, возникшим у самых ранних покрытосеменных растений . [33]
Цветение [ править ]
Ауксин также играет незначительную роль в инициировании цветения и развитии репродуктивных органов. В низких концентрациях он может замедлить старение цветов. Описан ряд мутантов растений, которые влияют на цветение и имеют дефицит либо синтеза, либо транспорта ауксина. У кукурузы одним из примеров является бесплодное соцветие bif22 . [34]
Биосинтез этилена [ править ]
В низких концентрациях ауксин может ингибировать образование этилена и транспорт предшественника в растениях; однако высокие концентрации могут индуцировать синтез этилена. [35] Следовательно, высокая концентрация может вызвать у некоторых видов женственность цветов. [ нужна ссылка ]
Ауксин ингибирует опадение до образования опадающего слоя и, таким образом, подавляет старение листьев.





Синтетические ауксины [ править ]
В ходе исследований по биологии ауксина было синтезировано множество соединений с заметной ауксиновой активностью. Было обнаружено, что многие из них обладают экономическим потенциалом для контролируемого человеком выращивания и развития растений в агрономии.
Ауксины токсичны для растений в больших концентрациях; они наиболее токсичны для двудольных и менее токсичны для однодольных . [36] Благодаря этому свойству синтетические ауксиновые гербициды, в том числе 2,4-дихлорфеноксиуксусная кислота (2,4-Д) и 2,4,5-трихлорфеноксиуксусная кислота были разработаны и используются для сорняками борьбы с (2,4,5-Т). .
Однако некоторые экзогенно синтезированные ауксины, особенно 1-нафталинуксусная кислота (NAA) и индол-3-масляная кислота (IBA), также широко применяются для стимуляции роста корней при черенках растений или в различных сельскохозяйственных целях, например, для предотвращения заглянуть в фруктовые сады .
При использовании в высоких дозах ауксин стимулирует выработку этилена , также нативного растительного гормона. Избыток этилена может препятствовать росту удлинения, вызывать листьев опадение ( опадение ) и даже гибель растения. Некоторые синтетические ауксины, такие как 2,4-Д и 2,4,5-Т, продаются также как гербициды . Двудольные растения , такие как одуванчики , гораздо более восприимчивы к ауксинам, чем однодольные растения , такие как травы и зерновые культуры. Таким образом, эти синтетические ауксины ценны как синтетические гербициды. 2,4-Д был первым широко используемым гербицидом, и он используется до сих пор. [37] 2,4-D был впервые коммерциализирован компанией Sherwin-Williams и начал использоваться в конце 1940-х годов. Его легко и недорого изготовить.
Было показано, что триклопир (3,5,6-ТПА), известный как гербицид, также увеличивает размер плодов растений. При повышенных концентрациях гормон может быть смертельным. Было показано, что снижение дозы до правильной концентрации изменяет пути фотосинтеза. Это препятствие для растения вызывает реакцию, которая увеличивает выработку углеводов, что приводит к увеличению размеров плодов. [38]
Производство гербицидов [ править ]
Синтетические ауксины используются как своего рода гербициды, и передозировка ауксинов прерывает рост растений и приводит к их гибели. [39] Дефолиант « Агент Оранж» , широко использовавшийся британскими войсками во время чрезвычайной ситуации в Малайзии и американскими войсками во время войны во Вьетнаме , представлял собой смесь 2,4-Д и 2,4,5-Т. Соединение 2,4-Д все еще используется и считается безопасным, но 2,4,5-Т был более или менее запрещен Агентством по охране окружающей среды США в 1979 году. Диоксин ТХДД является неизбежным загрязнителем, образующимся в производство 2,4,5-Т. В результате интегрального загрязнения диоксином употребление продуктов 2,4,5-Т связано с лейкемией , выкидышами , врожденными дефектами , повреждениями печени и другими заболеваниями .
См. также [ править ]
- Ауксин-связывающий белок
- Фузикокцин
- Гербицид ; в частности, см. раздел: §Ауксин
- Обрезка плодовых деревьев
- Тропизм
- Ведьмина метла
- Тосио Мурасиге
- Фолке К. Скуг
- Кеннет В. Тиманн
Ссылки [ править ]
- ^ Jump up to: Перейти обратно: а б с Саймон С., Петрашек Дж. (март 2011 г.). «Почему растениям нужно более одного типа ауксина» . Наука о растениях . 180 (3): 454–460. doi : 10.1016/j.plantsci.2010.12.007 . ПМИД 21421392 .
- ^ Jump up to: Перейти обратно: а б с Людвиг-Мюллер Дж (март 2011 г.). «Конъюгаты ауксина: их роль в развитии растений и в эволюции наземных растений» . Журнал экспериментальной ботаники . 62 (6): 1757–1773. дои : 10.1093/jxb/erq412 . ПМИД 21307383 .
Помимо ИУК, существует несколько других молекул с ауксиновой активностью, таких как индол-3-масляная кислота (ИМК) (рис. 1), 4-Cl-ИУК и индол-3-пропионовая кислота (ИПК). ... Индольный фрагмент (например, IAA, IBA, IPA, 4-Cl-IAA), а также партнер по конъюгату могут варьироваться, так что растение может производить множество различных комбинаций конъюгатов (Байгуз и Пиотровска, 2009). Кроме того, могут быть конъюгированы другие молекулы типа ауксина, такие как фенилуксусная кислота (PAA; Ludwig-Müller and Cohen, 2002) (Jentschel et al., 2007).
- ^ Jump up to: Перейти обратно: а б Таиз Л., Зейгер Э. (1998). Физиология растений (2-е изд.). Массачусетс: Sinauer Associates.
- ^ Фриц Вармолт Ушел
- ^ Jump up to: Перейти обратно: а б с д и ж г Фримл Дж. (февраль 2003 г.). «Транспорт ауксина – формирование растения» . Современное мнение в области биологии растений . 6 (1): 7–12. дои : 10.1016/S1369526602000031 . ПМИД 12495745 .
- ^ Jump up to: Перейти обратно: а б с д и Бенкова Э., Мичневич М., Зауэр М., Тейхманн Т., Зайфертова Д., Юргенс Г., Фримл Дж. (ноябрь 2003 г.). «Локальные, зависящие от оттока градиенты ауксина как общий модуль формирования органов растений» . Клетка . 115 (5): 591–602. дои : 10.1016/S0092-8674(03)00924-3 . ПМИД 14651850 . S2CID 16557565 .
- ^ Саймон Л., Буске Ж., Левеск Р.К., Лалонд М. (1993). «Происхождение и разнообразие эндомикоризных грибов и совпадение с сосудистыми наземными растениями». Природа . 363 (6424): 67–69. Бибкод : 1993Natur.363...67S . дои : 10.1038/363067a0 . S2CID 4319766 .
- ^ Сун, Ялин (2014). «Понимание механизма действия 2,4-дихлорфеноксиуксусной кислоты (2,4-Д) как гербицида». Журнал интегративной биологии растений . 56 (2): 106–113. дои : 10.1111/jipb.12131 . ПМИД 24237670 .
- ^ Хом Т., Пройтен Т., Фанкхаузер С. (январь 2013 г.). «Фототропизм: перевод света в направленный рост» (PDF) . Американский журнал ботаники . 100 (1): 47–59. дои : 10.3732/ajb.1200299 . ПМИД 23152332 . S2CID 25209232 .
- ^ Jump up to: Перейти обратно: а б Whippo CW, Hangarter RP (май 2006 г.). «Фототропизм: стремление к просветлению» . Растительная клетка . 18 (5): 1110–1119. дои : 10.1105/tpc.105.039669 . ПМЦ 1456868 . ПМИД 16670442 .
- ^ Бойсен Дж. П. (1910). «О проведении фототропного раздражителя у проростков Авены» . Отчеты Немецкого ботанического общества . 28 :118-120. дои : 10.1111/j.1438-8677.1910.tb06871.x . S2CID 257824886 .
- ^ Jump up to: Перейти обратно: а б Бойсен Дж. П. (1911). «Передача фототропного раздражения в Авене» (PDF) . Королевский Труды Датского общества наук . 1 :1–24.
- ^ Бойсен Дж. П. (1913). «О проведении фототропного раздражения в колеоптиле Авены» . Отчеты Немецкого ботанического общества . 31 :559-566. дои : 10.1111/j.1438-8677.1913.tb06990.x . S2CID 257819964 .
- ^ Ларсен П. (1960). «Питер Бойсен-Йенсен 1883-1959» . Физиология растений . 35 : 986–988. дои : 10.1104/стр.35.6.986 .
- ^ Jump up to: Перейти обратно: а б Крокер С., Макмиллан Дж., Гаскин П. «Природа ауксина» . Мендипвеб . Архивировано из оригинала 9 сентября 2002 г. Проверено 7 января 2009 г.
- ^ Jump up to: Перейти обратно: а б Лейзер О (январь 2018 г.). «Ауксиновая сигнализация» . Физиология растений . 176 (1): 465–479. дои : 10.1104/стр.17.00765 . ПМЦ 5761761 . ПМИД 28818861 .
- ^ Jump up to: Перейти обратно: а б с Эндерс Т.А., Стрейдер Л.С. (февраль 2015 г.). «Ауксиновая активность: прошлое, настоящее и будущее» . Американский журнал ботаники . 102 (2): 180–196. дои : 10.3732/ajb.1400285 . ПМЦ 4854432 . ПМИД 25667071 .
- ^ Тан, Сюй; Кальдерон-Вильялобос, Луз Ирина А.; Шарон, Михал; Чжэн, Чансюэ; Робинсон, Кэрол В.; Эстель, Марк; Чжэн, Нин (апрель 2007 г.). «Механизм восприятия ауксина убиквитинлигазой TIR1» . Природа . 446 (7136): 640–645. дои : 10.1038/nature05731 . ISSN 1476-4687 .
- ^ Фендрич М., Ахманова М., Меррин Дж., Гланк М., Хагихара С., Такахаши К. и др. (июль 2018 г.). «Быстрое и обратимое ингибирование роста корней посредством передачи сигналов ауксина TIR1» . Природные растения . 4 (7): 453–459. дои : 10.1038/s41477-018-0190-1 . ПМК 6104345 . ПМИД 29942048 .
- ^ Машигути К., Танака К., Сакаи Т., Сугавара С., Кавайде Х., Нацумэ М. и др. (ноябрь 2011 г.). «Основной путь биосинтеза ауксина у Arabidopsis» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (45): 18512–18517. Бибкод : 2011PNAS..10818512M . дои : 10.1073/pnas.1108434108 . ПМК 3215075 . ПМИД 22025724 .
- ^ Петрасек Дж., Мравец Дж., Бушар Р., Блейксли Дж.Дж., Абас М., Сейфертова Д. и др. (май 2006 г.). «Белки PIN выполняют функцию ограничения скорости клеточного оттока ауксина» . Наука . 312 (5775): 914–918. Бибкод : 2006Sci...312..914P . дои : 10.1126/science.1123542 . ПМИД 16601150 . S2CID 28800759 .
- ^ Сваруп Р., Перет Б (1 января 2012 г.). «Семейство носителей притока ауксина AUX / LAX - обзор» . Границы в науке о растениях . 3 : 225. дои : 10.3389/fpls.2012.00225 . ПМЦ 3475149 . ПМИД 23087694 .
- ^ Сабатини С., Бейс Д., Волкенфельт Х., Мерфетт Дж., Гилфойл Т., Малами Дж. и др. (ноябрь 1999 г.). «Ауксин-зависимый дистальный организатор структуры и полярности в корне Arabidopsis». Клетка . 99 (5): 463–472. дои : 10.1016/S0092-8674(00)81535-4 . hdl : 1874/21099 . ПМИД 10589675 . S2CID 8041065 .
- ^ Jump up to: Перейти обратно: а б с Хейслер М.Г., Оно С., Дас П., Зибер П., Редди Г.В., Лонг Дж.А., Мейеровиц Э.М. (ноябрь 2005 г.). «Закономерности транспорта ауксина и экспрессии генов во время развития зачатка, выявленные с помощью живого изображения меристемы соцветия арабидопсиса» . Современная биология . 15 (21): 1899–1911. дои : 10.1016/j.cub.2005.09.052 . ПМИД 16271866 . S2CID 14160494 .
- ^ Тан С., Чжан X, Конг В., Ян XL, Мольнар Г., Вондракова З. и др. (май 2020 г.). «Зависимый от липидного кода фосфопереключатель PDK1-D6PK активирует PIN-опосредованный отток ауксина у Arabidopsis». Природные растения . 6 (5): 556–569. дои : 10.1038/s41477-020-0648-9 . ПМИД 32393881 . S2CID 218593545 .
- ^ Jump up to: Перейти обратно: а б Сорефан К., Гирин Т., Лильегрен С.Дж., Люнг К., Роблес П., Гальван-Ампудиа К.С. и др. (май 2009 г.). «Для распространения семян арабидопсиса необходим регулируемый минимум ауксина». Природа . 459 (7246): 583–586. Бибкод : 2009Natur.459..583S . дои : 10.1038/nature07875 . ПМИД 19478783 . S2CID 4411776 .
- ^ Кречек П., Скупа П., Либус Дж., Нарамото С., Техос Р., Фримл Дж., Зазималова Э. (29 декабря 2009 г.). «Семейство белков-переносчиков ауксина PIN-FORMED (PIN)» . Геномная биология . 10 (12): 249. doi : 10.1186/gb-2009-10-12-249 . ПМК 2812941 . ПМИД 20053306 .
- ^ «ЧАШЕЧНО-ОБРАЗНАЯ СЯТОЛЕДОН3 арабидопсиса регулирует постэмбриональную меристему побега и формирование границ органов» . Викигенес. 2006.
- ^ Алони Р., Алони Э., Лангханс М., Ульрих К.И. (май 2006 г.). «Роль цитокинина и ауксина в формировании архитектуры корня: регуляция сосудистой дифференцировки, инициации бокового корня, доминирования апикальной части корня и гравитропизма корня» . Анналы ботаники . 97 (5): 883–893. дои : 10.1093/aob/mcl027 . ПМК 2803412 . ПМИД 16473866 .
- ^ Чемберс (1999). Научно-технический словарь . ISBN 978-0-550-14110-1 .
- ^ «Вот почему растения растут навстречу свету!» . Лаборатория Иржи Фримла . VIB (Фландрийский институт биотехнологии. 2012. Архивировано из оригинала 15 декабря 2018 г.).
- ^ Немхаузер Дж. Л., Фельдман Л. Дж., Замбриски ПК (сентябрь 2000 г.). «Ауксин и ЭТТИН в морфогенезе гинецея Arabidopsis». Разработка . 127 (18): 3877–3888. дои : 10.1242/dev.127.18.3877 . ПМИД 10952886 .
- ^ Фридман В.Е. (июнь 2009 г.). «Эволюция. Ауксин на перекрестке эво-дево». Наука . 324 (5935). Американская ассоциация содействия развитию науки : 1652–1653. Бибкод : 2009Sci...324.1652F . дои : 10.1126/science.1176526 . ПМИД 19556491 . S2CID 206521265 .
- ^ МакСтин П. , Малкомбер С., Скирпан А., Лунде С., Ву Икс, Келлог Э., Хейк С. (июнь 2007 г.). «бесплодное соцветие2» Кодирует аналог серин/треониновой киназы PINOID и необходим для органогенеза во время соцветия и вегетативного развития кукурузы . Физиология растений . 144 (2): 1000–1011. дои : 10.1104/стр.107.098558 . ЧВК 1914211 . ПМИД 17449648 .
- ^ Ю Ю.Б., Ян С.Ф. (декабрь 1979 г.). «Ауксин-индуцированное производство этилена и его ингибирование аминоэтиоксивинилглицином и ионом кобальта» . Физиология растений . 64 (6): 1074–1077. дои : 10.1104/стр.64.6.1074 . ПМЦ 543194 . ПМИД 16661095 .
- ^ МакСтин П. (март 2010 г.). «Развитие ауксина и однодольных растений» . Перспективы Колд-Спринг-Харбор в биологии . 2 (3): а001479. doi : 10.1101/cshperspect.a001479 . ПМК 2829952 . ПМИД 20300208 .
- ^ Отраслевая целевая группа II по данным исследований 2,4-D
- ^ Месехо С., Росито С., Рейг С., Мартинес-Фуэнтес А., Агусти М. (2012). «Синтетический ауксин 3,5,6-ТПА провоцирует опадение плодов Citrus clementina (Hort. ex Tan) за счет снижения доступности фотосинтата». Журнал регулирования роста растений . 31 (2): 186–194. дои : 10.1007/s00344-011-9230-z . S2CID 8338429 .
- ^ Гроссманн К. (сентябрь 2007 г.). «Действие гербицида ауксина: шаг за шагом поднимаем завесу» . Сигнализация и поведение растений . 2 (5): 421–423. дои : 10.4161/psb.2.5.4417 . ПМЦ 2634233 . ПМИД 19704620 .
Дальнейшее чтение [ править ]
- Локасио А, Ройг-Вилланова И, Бернарди Дж, Варотто С (25 августа 2014 г.). «Современные перспективы гормонального контроля развития семян арабидопсиса и кукурузы: акцент на ауксине» . Границы в науке о растениях . 5 . Границы : 412. doi : 10.3389/fpls.2014.00412 . ПМЦ 4142864 . ПМИД 25202316 .