Нотохорда
Эта статья может быть слишком технической для понимания большинства читателей . ( февраль 2017 г. ) |
| Нотохорда | |
|---|---|
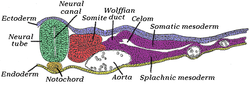 Поперечный срез куриного эмбриона после сорокапятичасовой инкубации. | |
| Подробности | |
| Предшественник | Осевая мезодерма |
| Дает начало | Студенистое ядро |
| Идентификаторы | |
| латинский | в беде |
| МеШ | С000000 |
| ТО | Е5.0.1.1.0.0.8 |
| ФМА | 85521 |
| Анатомическая терминология | |

В зоологии и анатомии развития хорда многих представляет собой эластичную стержнеобразную анатомическую структуру, встречающуюся у вторичноротых животных . Нотохорда — одна из пяти синапоморфий или характеристик, используемых для определения вида как хордового .
Хорда происходит из эмбриональной мезодермы и состоит из внутреннего ядра вакуолизированных клеток, заполненных гликопротеинами , покрытых двумя спиральными коллагеново - эластиновыми оболочками. Он расположен вдоль рострально - каудальной оси тела (т.е. продольно или « от головы к хвосту »), дорсально по отношению к кишечной трубке и вентрально по отношению к дорсальному нервному шнуру . У некоторых хордовых, таких как оболочники , хорда развивается на личиночной стадии, но теряется на последующих стадиях взрослой жизни.
Хорда важна для передачи сигналов о дорсо-вентральном паттерне клеток, происходящих из мезодермальных предшественников. Это помогает сформировать предшественники, необходимые для развития определенных органов и эмбриона. Таким образом, хорда играет важную роль в эмбриональном развитии.
Хорда обеспечивает направленную связь с окружающими тканями в качестве срединной структуры во время эмбрионального развития , действует как предшественник позвонков и примитивного осевого эндоскелета и может обеспечивать облегченное движение хвоста при плавании. [ 1 ]
Присутствие
[ редактировать ]У головохордовых ( ланцетников ) хорда сохраняется на протяжении всей жизни как основная структурная опора тела.
У оболочников хорда имеется только на личиночной стадии, у взрослого животного она полностью отсутствует, хорда не вакуолизирована. [ 2 ]
У всех позвоночных , кроме миксины , хорда присутствует только на ранних этапах эмбрионального развития и позже заменяется костным и/или хрящевым позвоночником , при этом ее первоначальная структура интегрируется в межпозвоночные диски в виде студенистого ядра . [ 3 ] [ 4 ]
Структура
[ редактировать ]Хорда представляет собой длинную палочковидную срединную структуру, которая развивается дорсально от кишечной трубки и вентрально от нервной трубки . Хорда состоит в основном из гликопротеинового ядра, окруженного оболочкой из коллагеновых волокон. Он закручен в две противоположные спирали . Гликопротеины хранятся в вакуолизированных, набухших клетках, покрытых кавеолами на клеточной поверхности. [ 5 ] Угол между этими волокнами определяет, приведет ли повышенное давление в ядре к укорочению и утолщению или к удлинению и истончению. [ 6 ]
Поочередное сокращение мышечных волокон, прикрепленных к каждой стороне хорды, приводит к движению из стороны в сторону, напоминающему корму , что позволяет хвосту плавать и волнообразно двигаться . Затвердевшая хорда предотвращает телескопическое движение , такое как у дождевого червя . [ 7 ]
Роль в передаче сигналов и развитии
[ редактировать ]Хорда играет ключевую роль в передаче сигналов и координации развития. Эмбрионы современных позвоночных образуют временные структуры хорды во время гаструляции . Хорда находится вентральнее нервной трубки .
Нотогенез – это развитие хорды эпибластами , образующими дно полости амниона . [ 8 ] Прародительская хорда происходит из клеток, мигрирующих из примитивного узла и ямки. [ 9 ] Хорда формируется во время гаструляции и вскоре после этого вызывает образование нервной пластинки ( нейруляцию ), синхронизируя развитие нервной трубки . На вентральной стороне нервной борозды осевое утолщение энтодермы происходит . (У двуногих хордовых, например у человека, эту поверхность правильно называют передней поверхностью). Это утолщение выглядит как борозда (хордальная борозда), края которой анастомозируют (вступают в контакт) и таким образом превращают ее в твердый стержень из клеток многоугольной формы (хорда), который затем отделяется от энтодермы. [ нужна ссылка ]
У позвоночных он распространяется на всю длину будущего позвоночного столба и доходит до переднего конца среднего мозга , где заканчивается крючковидным окончанием в области будущей дорсальной поверхности клиновидной кости . Первоначально он существует между нервной трубкой и энтодермой желточного мешка; вскоре хорда отделяется от них мезодермой , которая разрастается медиально и окружает ее. Из мезодермы, окружающей нервную трубку и хорду, череп , позвоночный столб, оболочки головного . и спинного мозга развиваются [ 10 ] Поскольку он происходит из примитивного узла и в конечном итоге располагается в мезодермальном пространстве, его считают производным мезодермы. [ 11 ]
Постэмбриональный остаток хорды обнаруживается в студенистом ядре межпозвоночных дисков. Изолированные нотохордальные остатки могут ускользать от своего специфического для линии места назначения в студенистом ядре и вместо этого прикрепляться к внешним поверхностям тел позвонков , из которых нотохордальные клетки в значительной степени регрессируют. [ 12 ]
У амфибий и рыб
[ редактировать ]Во время развития земноводных и рыб хорда индуцирует развитие гипохорда за счет секреции фактора роста эндотелия сосудов . Гипохорд представляет собой временную структуру вентральнее хорды и в первую очередь отвечает за правильное развитие дорсальной аорты. [ 13 ]
Сгибание хорды , когда хорда сгибается, образуя часть развивающегося хвостового плавника, является признаком ранней стадии роста некоторых рыб. [ 14 ] [ 15 ] [ нужен лучший источник ]
У людей
[ редактировать ]К 4 годам все остатки хорды замещаются популяцией хондроцитоподобных клеток неясного происхождения. [ 16 ] Сохранение нотохордальных клеток внутри позвонка может вызвать патологическое состояние: персистирующий хордальный канал . [ 17 ] Если хорда и носоглотка не разделяются должным образом во время эмбрионального развития, может образоваться депрессия (бурса Торнвальдта) или киста Торнвальдта . [ 18 ] Эти клетки являются вероятными предшественниками редкого рака, называемого хордомой . [ 19 ]
Неврология
[ редактировать ]Исследования хорды сыграли ключевую роль в понимании развития центральной нервной системы . Трансплантировав и экспрессировав вторую хорду рядом с дорсальной нервной трубкой, на 180 градусов напротив нормального расположения хорды, можно индуцировать образование мотонейронов в дорсальной трубке. Формирование мотонейронов обычно происходит в вентральной части нервной трубки, тогда как в дорсальной трубке обычно образуются сенсорные клетки . [ 20 ]
Хорда секретирует белок, называемый звуковым ежом (SHH), ключевой морфоген, регулирующий органогенез и играющий решающую роль в передаче сигналов развитию мотонейронов. [ 21 ] Секреция SHH хордой устанавливает вентральный полюс дорсально-вентральной оси развивающегося эмбриона.
Эволюция хордовых
[ редактировать ]
Нотохорда является определяющей особенностью ( синапоморфией ) хордовых и присутствовала на протяжении всей жизни у многих самых ранних хордовых. Хотя гомологичен или желудок полухордовых когда-то считалось, что имеет общее линейное происхождение, теперь его рассматривают как аналогичный, конвергентный или происходящий от другого линейного происхождения. [ 22 ] Пикайя , по-видимому, имеет прото-нотохорду, а хорды присутствуют у нескольких базальных хордовых, таких как Haikouella , Haikouichthys и Myllokunmingia , все из кембрия .
Ордовикские , которые обладали хордами либо с прикрепленными костными элементами , океаны включали множество разнообразных видов Agnatha и ранних Gnathostomata либо без них, в первую очередь конодонтами . [ 23 ] плакодермы , [ 24 ] и остракодермы . Даже после эволюции позвоночного столба у chondrichthyes и Osteichthyes эти таксоны оставались обычными и хорошо представлены в летописи окаменелостей. Некоторые виды (см. список ниже) вернулись в примитивное состояние, сохранив хорду и во взрослом возрасте, хотя причины этого до конца не изучены.
Сценарии эволюционного происхождения хорды были всесторонне рассмотрены Анноной, Холландом и Д'Аньелло (2015). [ 25 ] Они отмечают, что, хотя многие из этих идей не получили должного подтверждения достижениями молекулярной филогенетики и генетики развития, две из них фактически были возрождены под воздействием современных молекулярных подходов (первый предполагает, что хорда развилась de novo у хордовых). , а второй происходит от гомологичной структуры, аксохорды, которая присутствовала у кольчатоподобных предков хордовых). Выбор между этими двумя сценариями (или, возможно, еще одним, который еще предстоит предложить) должен быть облегчен гораздо более тщательными исследованиями сетей регуляции генов у широкого спектра животных.
Постэмбриональная ретенция
[ редактировать ]У большинства позвоночных хорда развивается во вторичные структуры. У других хордовых хорда сохраняется как важнейшая анатомическая структура. Эволюция хорды внутри типа Chordata подробно рассмотрена Холландом и Соморжаем (2020). У позвоночных теперь есть шипы, поэтому хорда им не нужна. [ 26 ]
Следующие организмы сохраняют постэмбриональную хорду:
- Acipenseriformes ( веслонос и осетр ) [ 27 ]
- Ланцетник ( Амфиоксус )
- Оболочник (только личиночная стадия)
- миксина
- Минога
- Целаканты
- Африканская двоякодышащая рыба
- Головастики
- Остракодермы (вымершие)
Внутри Амфиоксуса
[ редактировать ]Хорда ланцетника выступает за передний конец нервной трубки. Этот выступ служит второй цели, позволяя животному зарываться в отложениях на мелководье. Там амфиоксус является фильтратором и большую часть своей жизни проводит частично погруженным в осадок. [ 7 ]
Дополнительные изображения
[ редактировать ]-
 Вид на поверхность эмбриона гиббона Concolor ( Hylobates concolor ).
Вид на поверхность эмбриона гиббона Concolor ( Hylobates concolor ). -
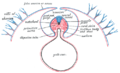 Схема поперечного среза, показывающая способ формирования амниона у цыпленка.
Схема поперечного среза, показывающая способ формирования амниона у цыпленка. -
 Разрез головы человеческого эмбриона двенадцатидневного возраста в области заднего мозга.
Разрез головы человеческого эмбриона двенадцатидневного возраста в области заднего мозга. -
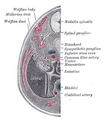 Поперечный срез человеческого эмбриона в возрасте от восьми с половиной до девяти недель.
Поперечный срез человеческого эмбриона в возрасте от восьми с половиной до девяти недель.




Ссылки
[ редактировать ]- ^ Шифферл, Д., Шольце-Виттлер, М., Вильяронга Луке, А., Пустет, М., Виттлер, Л., Венвлит, Й.В., Кох, Ф., и Херрманн, Б.Г. (2023). Полногеномная идентификация энхансеров хорды, составляющих регуляторный ландшафт локуса brachyury у мышей. Развитие (Кембридж, Англия), 150 (22). https://doi.org/10.1242/dev.202111
- ^ Ван, Ф., Чжан, К., Ши, Р., Се, З.-Ю., Чен, Л., Ван, К., Ван, Ю.-Т., Се, Х.-Х., И Ву, X.-T. (2018). Эмбриональные и эволюционные границы между хордой и хрящом: новый взгляд на маркеры, специфичные для студенистого ядра. Остеоартрит и хрящ , 26 (10), 1274–1282. https://doi.org/10.1016/j.joca.2018.05.022
- ^ Кремер, Юрген (2009). Заболевания межпозвоночных дисков: причины, диагностика, лечение и профилактика . Тиме. стр. 15–17. ISBN 978-3-13-582403-1 .
- ^ Стемпл, Дерек Л. (1 июня 2005 г.). «Структура и функция хорды: важный орган хордовых» . Разработка . 132 (11): 2503–2512. дои : 10.1242/dev.01812 . ISSN 0950-1991 . ПМИД 15890825 .
- ^ Лим, Йе-Вин; Ло, Харриет П.; Холл, Томас Э.; Партон, Роберт Г. (2020), «Живая конфокальная визуализация клеток хорды данио в условиях механического стресса in vivo», Caveolae , Methods in Molecular Biology, vol. 2169, Нью-Йорк, штат Нью-Йорк: Springer US, стр. 175–187, doi : 10.1007/978-1-0716-0732-9_16 , ISBN. 978-1-0716-0731-2 , PMID 32548829 , S2CID 219725868 , получено 14 января 2023 г.
- ^ МАР Кёль (2000). «Механическое проектирование гидравлических скелетов с волокнистой обмоткой: усиление и выпрямление эмбриональных хорд» . Американский зоолог . 40 : 28–041. дои : 10.1093/icb/40.1.28 .
- ^ Jump up to: а б Хомбергер, Доминик Г. (2004). Рассечение позвоночных . Уокер, Уоррен Ф. (Warren Franklin), Уокер, Уоррен Ф. (Warren Franklin). (9-е изд.). Бельмонт, Калифорния: Томсон Брукс/Коул. ISBN 0-03-022522-1 . OCLC 53074665 .
- ^ «Треламинарный зародышевый диск (3-я неделя)» . www.embryology.ch . Архивировано из оригинала 31 мая 2017 г. Проверено 9 января 2012 г.
- ^ Худ, Руссо, Блейкли, Рональд Д., Колин Г., Патрисия М. (29 мая 2007 г.). «Эмбрион и плод». Справочник по токсикологической патологии (второе издание) . 2 . Academic Press, издательство Elsevier Inc: 895–936. дои : 10.1016/b978-0-12-330215-1.50047-8 . ISBN 9780123302151 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Генри Грей (1918). Анатомия человеческого тела . Леа и Фебигер. стр. 52–54.
- ^ Гэри К. Шенвульф; Стивен Б. Блейл; Филип Р. Брауэр; Филиппа Х. Фрэнсис-Уэст (1 декабря 2014 г.). Электронная книга Ларсена по эмбриологии человека . Elsevier Науки о здоровье. стр. 71–72. ISBN 978-1-4557-2791-9 .
- ^ Чой, К.; Кон, Мартин Дж.; Харф, Брайан Д. (2009). «Идентификация клеток-предшественников студенистого ядра и остатков хорды у мышей: последствия для дегенерации диска и формирования хордомы» . Динамика развития . 237 (12): 3953–3958. дои : 10.1002/dvdy.21805 . ПМК 2646501 . ПМИД 19035356 .
- ^ Кливер, Ундина (2000). «Формирование паттерна эндодермы с помощью хорды: развитие гипохорда у Xenopus» (PDF) . Разработка . 127 (4): 869–979. дои : 10.1242/dev.127.4.869 . ПМИД 10648245 .
- ^ Пакстон, Джон Р.; Джонсон, Дж. Дэвид; Трнски, Томас (2001). «Личинки и молодь глубоководных «китов» Barbourisia и Rondeletia (Stephanoberyciformes: Barbourisiidae, Rondeletiidae) с комментариями о семейных отношениях» (PDF) . Записи Австралийского музея . 53 (3): 407–425. дои : 10.3853/j.0067-1975.53.2001.1352 . Архивировано из оригинала (PDF) 26 сентября 2003 г.
- ^ «Коричневая рыба-призрак» (PDF) . Информационная система ихтиопланктона. Научный центр рыболовства Аляски. Национальное управление океанографии и атмосферы . Июль 2008 года . Проверено 14 марта 2009 г.
- ^ Урбан, JPG (2000). «Ядро межпозвоночного диска от развития до дегенерации» . Интегративная и сравнительная биология . 40 : 53–061. дои : 10.1093/icb/40.1.53 .
- ^ Кристоферсон, младший; Рабин, Бм; Халлам, Дания; Рассел, Эй Джей (1 января 1999 г.). «Сохранение нотохордального канала: МРТ и обычный вид пленки» (полный текст) . Американский журнал нейрорадиологии . 20 (1): 33–6. ISSN 0195-6108 . ПМИД 9974055 .
- ^ Муди М.В., Чи Д.Х., Чи Д.М., Мейсон Дж.К., Филлипс К.Д., Гросс К.В. и др. (2007). «Киста Торнвальдта: заболеваемость и клинический случай» . Ухо Нос Горло Дж . 86 (1): 45–7, 52. doi : 10.1177/014556130708600117 . ПМИД 17315835 .
- ^ Пиллаи С., Говендер С. (2018). «Сакральная хордома: Обзор литературы» . Дж Ортоп . 15 (2): 679–684. дои : 10.1016/j.jor.2018.04.001 . ПМК 5990241 . ПМИД 29881220 .
- ^ Уилсон, Ли; Маден, Малькольм (2005). «Механизмы формирования дорсовентрального паттерна в нервной трубке позвоночных» . Биология развития . 282 (1). Эльзевир Б.В.: 1–13. дои : 10.1016/j.ydbio.2005.02.027 . ISSN 0012-1606 . ПМИД 15936325 .
- ^ Эшелард, Ю; Эпштейн, диджей; Сен-Жак, Б; Шен, Л; Молер, Дж; Макмахон, Джа; Макмахон, Ап (декабрь 1993 г.). «Sonic hedgehog, член семейства предполагаемых сигнальных молекул, участвует в регуляции полярности ЦНС». Клетка . 75 (7): 1417–30. дои : 10.1016/0092-8674(93)90627-3 . ПМИД 7916661 . S2CID 6732599 .
- ^ Кардонг, Кеннет В. (1995). Позвоночные животные: сравнительная анатомия, функции, эволюция . МакГроу-Хилл. стр. 55, 57. ISBN. 978-0-697-21991-6 .
- ^ «Палеопозвоночные 30 000 Conodonta: обзор» . Архивировано из оригинала 13 марта 2006 г. Проверено 5 сентября 2007 г.
- ^ «Обзор плакодерм 60 000 палеопозвоночных» . Архивировано из оригинала 20 декабря 2010 года . Проверено 21 ноября 2009 г.
- ^ Аннона, Г.; Голландия, Северная Дакота; Д'Аньелло, С. (2015). «Эволюция хорды» . ЭвоДево . 6 . статья 30. doi : 10.1186/s13227-015-0025-3 . ПМЦ 4595329 . ПМИД 26446368 .
- ^ Голландия, Северная Дакота; Соморжай, IML (2020). «Серийная СЭМ-сканер предполагает, что стволовые клетки могут участвовать в росте хорды взрослого беспозвоночного хордового, багамского ланцетника» . ЭвоДево . 11 . статья 22. doi : 10.1186/s13227-020-00167-6 . ПМЦ 7568382 . PMID 33088474 .
- ^ Джозеф Дж. Лушкович; Филип Дж. Мотта; Стивен Ф. Нортон; Карел Ф. Лием (17 апреля 2013 г.). Экоморфология рыб . Springer Science & Business Media. п. 201. ИСБН 978-94-017-1356-6 .