Псевдоген

Псевдогены — это нефункциональные сегменты ДНК , напоминающие функциональные гены . Большинство из них возникают как избыточные копии функциональных генов либо непосредственно в результате дупликации генов , либо косвенно в результате обратной транскрипции транскрипта мРНК . Псевдогены обычно идентифицируются, когда анализ последовательности генома обнаруживает геноподобные последовательности, в которых отсутствуют регуляторные последовательности, необходимые для транскрипции или трансляции , или кодирующие последовательности которых явно дефектны из-за сдвигов рамки считывания или преждевременных стоп-кодонов . Псевдогены — это разновидность мусорной ДНК .
Большинство небактериальных геномов содержат множество псевдогенов, часто столько же, сколько и функциональных генов. Это неудивительно, поскольку ожидается, что различные биологические процессы случайно создадут псевдогены, а специализированных механизмов для их удаления из геномов не существует. В конечном итоге псевдогены могут быть удалены из своих геномов в результате ошибок репликации ДНК или репарации ДНК , или же они могут накопить столько мутационных изменений, что их больше нельзя будет распознать как прежние гены. Анализ этих событий дегенерации помогает прояснить эффекты неселективных процессов в геномах.
Последовательности псевдогенов могут транскрибироваться в РНК на низких уровнях благодаря промоторным элементам, унаследованным от предкового гена или возникающим в результате новых мутаций. Хотя большинство из этих транскриптов будут иметь не большее функциональное значение, чем случайные транскрипты из других частей генома, некоторые из них дали начало полезным регуляторным РНК и новым белкам.
Характеристики
[ редактировать ]В этом разделе отсутствует информация о более быстром накоплении мутаций. ( декабрь 2023 г. ) |
Псевдогены обычно характеризуются сочетанием сходства с известным геном и утратой некоторой функциональности. То есть, хотя каждый псевдоген имеет последовательность ДНК , аналогичную некоторому функциональному гену, они обычно не способны производить функциональные конечные белковые продукты. [ 1 ] Псевдогены иногда трудно идентифицировать и охарактеризовать в геномах, поскольку два требования — сходство и потеря функциональности — обычно подразумеваются посредством выравнивания последовательностей, а не доказываются биологически.
- Гомология подразумевается сходством последовательностей между последовательностями ДНК псевдогена и известного гена. После выравнивания процент идентичных пар оснований двух последовательностей вычисляется . Высокая идентичность последовательностей означает, что весьма вероятно, что эти две последовательности разошлись от общей предковой последовательности (гомологичны), и крайне маловероятно, что эти две последовательности развились независимо (см. Конвергентную эволюцию ).
- Нефункциональность может проявляться по-разному. Обычно ген должен пройти несколько этапов, чтобы стать полностью функциональным белком: транскрипция , процессинг пре-мРНК , трансляция и сворачивание белка — все это необходимые части этого процесса. Если какой-либо из этих шагов не удался, последовательность можно считать нефункциональной. При высокопроизводительной идентификации псевдогенов наиболее часто выявляемыми нарушениями являются преждевременные стоп-кодоны и сдвиги рамки считывания , которые почти всегда предотвращают трансляцию функционального белкового продукта.
Псевдогены генов РНК обычно труднее обнаружить, поскольку их не нужно транслировать и, следовательно, они не имеют «рамок считывания». Ряд псевдогенов рРНК был идентифицирован на основании изменений концов массива рДНК. [ 2 ]
Псевдогены могут усложнить молекулярно-генетические исследования. Например, амплификация гена с помощью ПЦР может одновременно амплифицировать псевдоген, имеющий схожие последовательности. Это известно как смещение ПЦР или смещение амплификации. Точно так же псевдогены иногда обозначаются как гены в генома последовательностях .
Обработанные псевдогены часто создают проблемы для программ прогнозирования генов , поскольку их часто ошибочно принимают за настоящие гены или экзоны. Было высказано предположение, что идентификация обработанных псевдогенов может помочь повысить точность методов прогнозирования генов. [ 3 ]
В 2014 году было показано, что транслируются 140 человеческих псевдогенов. [ 4 ] Однако функция белковых продуктов, если таковая имеется, неизвестна.
Виды и происхождение
[ редактировать ]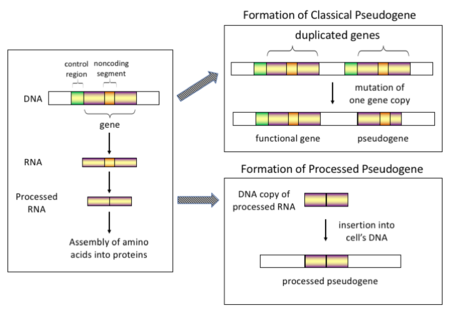
Существует четыре основных типа псевдогенов, каждый из которых имеет разные механизмы происхождения и характерные особенности. Классификация псевдогенов следующая:
Обработано
[ редактировать ]
У высших эукариот , особенно млекопитающих , ретротранспозиция — довольно распространенное событие, оказавшее огромное влияние на состав генома. Например, где-то от 30 до 44% генома человека состоит из повторяющихся элементов, таких как SINE и LINE (см. ретротранспозоны ). [ 7 ] [ 8 ] В процессе ретротранспозиции часть транскрипта мРНК или гнРНК гена спонтанно обратно транскрибируется обратно в ДНК и встраивается в хромосомную ДНК. было показано Хотя ретротранспозоны обычно создают копии самих себя, в системе in vitro , что они также могут создавать ретротранспонированные копии случайных генов. [ 9 ] Как только эти псевдогены встраиваются обратно в геном, они обычно содержат поли-А-хвост , а их интроны обычно расщепляются ; обе эти особенности являются отличительными чертами кДНК . Однако, поскольку они происходят из продукта РНК, процессированные псевдогены также лишены вышестоящих промоторов нормальных генов; таким образом, они считаются «мертвыми по прибытии» и становятся нефункциональными псевдогенами сразу после события ретротранспозиции. [ 10 ] Однако эти вставки иногда вносят экзоны в существующие гены, обычно через альтернативно сплайсированные транскрипты. [ 11 ] Еще одной характеристикой процессированных псевдогенов является обычное усечение 5'-конца относительно родительской последовательности, что является результатом относительно непроцессивного механизма ретротранспозиции, который создает процессированные псевдогены. [ 12 ] У приматов постоянно создаются процессированные псевдогены. [ 13 ] Например, человеческие популяции имеют отдельные наборы обработанных псевдогенов у своих особей. [ 14 ]
Показано, что обработанные псевдогены накапливают мутации быстрее, чем непроцессированные. [ 15 ]
Необработанные (дублированные)
[ редактировать ]
Дупликация генов — еще один распространенный и важный процесс в эволюции геномов. Копия функционального гена может возникнуть в результате события дупликации гена, вызванного гомологичной рекомбинацией , например, повторяющихся последовательностей SINE на смещенных хромосомах, и впоследствии приобрести мутации , которые заставляют копию терять функцию исходного гена. Дуплицированные псевдогены обычно имеют все те же характеристики, что и гены, включая интактную экзон - интронную структуру и регуляторные последовательности. Потеря функциональности дублированного гена обычно мало влияет на приспособленность организма , поскольку неповрежденная функциональная копия все еще существует. Согласно некоторым эволюционным моделям, общие дублированные псевдогены указывают на эволюционное родство человека и других приматов. [ 16 ] Если псевдогенизация вызвана дупликацией гена, она обычно происходит в первые несколько миллионов лет после дупликации гена, при условии, что ген не подвергался какому-либо давлению отбора . [ 17 ] Дупликация генов приводит к функциональной избыточности , и обычно невыгодно иметь два идентичных гена. Мутации, которые нарушают структуру или функцию любого из двух генов, не являются вредными и не будут удалены в процессе отбора. В результате мутировавший ген постепенно становится псевдогеном и либо неэкспрессируется, либо становится нефункциональным. Такая эволюционная судьба показана популяционно -генетическим моделированием. [ 18 ] [ 19 ] а также анализом генома . [ 17 ] [ 20 ] В соответствии с эволюционным контекстом эти псевдогены либо будут удалены, либо станут настолько отличными от родительских генов, что их больше нельзя будет идентифицировать. Относительно молодые псевдогены можно распознать благодаря сходству их последовательностей. [ 21 ]
Унитарные псевдогены
[ редактировать ]
Различные мутации (такие как инделы и нонсенс-мутации гена ) могут помешать нормальной транскрипции или трансляции , и, таким образом, ген может стать менее функциональным, нефункциональным или «деактивированным». Это те же механизмы, с помощью которых непроцессированные гены становятся псевдогенами, но разница в этом случае состоит в том, что ген не был дублирован до псевдогенизации. Обычно такой псевдоген вряд ли закрепится в популяции, но различные популяционные эффекты, такие как генетический дрейф , узкое место в популяции или, в некоторых случаях, естественный отбор , могут привести к фиксации. Классическим примером унитарного псевдогена является ген, который предположительно кодирует у приматов фермент L-гулоно-γ-лактоноксидазу (GULO). У всех изученных млекопитающих, кроме приматов (за исключением морских свинок), GULO способствует биосинтезу аскорбиновой кислоты (витамина С), но у людей и других приматов он существует в виде отключенного гена (GULOP). [ 22 ] [ 23 ] Другой, более свежий пример отключенного гена связывает деактивацию гена каспазы 12 (посредством нонсенс-мутации ) с положительным отбором у людей. [ 24 ]
Примеры псевдогенной функции
[ редактировать ]Хотя подавляющее большинство псевдогенов утратили свою функцию, возникли случаи, когда псевдоген либо вновь обретал свою первоначальную или аналогичную функцию, либо развил новую функцию. В геноме человека был идентифицирован ряд примеров, которые первоначально были классифицированы как псевдогены, но позже было обнаружено, что они играют функциональную, хотя и не обязательно кодирующую белок, роль. [ 25 ] [ 26 ]
Примеры включают следующее:
Кодирование белка: " псевдопсевдогены »
[ редактировать ]
Быстрое распространение технологий секвенирования ДНК привело к идентификации многих очевидных псевдогенов с использованием методов прогнозирования генов . Псевдогены часто идентифицируются по появлению преждевременного стоп-кодона в предсказанной последовательности мРНК, который теоретически предотвратил бы синтез ( трансляцию ) нормального белкового продукта исходного гена. Имелись сообщения о трансляционном считывании таких преждевременных стоп-кодонов у млекопитающих. Как указано на рисунке выше, небольшое количество белкового продукта такого прочтения все еще может быть распознаваемым и функционировать на определенном уровне. Если это так, то псевдоген может быть подвергнут естественному отбору . Похоже, это произошло во время эволюции дрозофил видов .
В 2016 году сообщалось, что четыре предсказанных псевдогена у нескольких видов дрозофилы на самом деле кодируют белки с биологически важными функциями. [ 27 ] «предполагая, что такие« псевдопсевдогены »могут представлять собой широко распространенное явление». Например, функциональный белок ( обонятельный рецептор глутамата ) из гена Ir75a обнаружен только в нейронах . Это обнаружение тканеспецифичных биологически-функциональных генов, которые можно было бы классифицировать как псевдогены с помощью анализа in silico , усложняет анализ данных о последовательностях. [ 27 ] Другой дрозофилии псевдопсевдоген — цзинвэй , [ 28 ] [ 29 ] который кодирует функциональный алкогольдегидрогеназу фермент in vivo . [ 30 ]
По состоянию на 2012 год выяснилось, что в геноме человека насчитывается примерно 12 000–14 000 псевдогенов. [ 31 ] 2016 года Протеогеномный анализ с использованием масс-спектрометрии пептидов выявил по меньшей мере 19 262 человеческих белка, продуцируемых из 16 271 гена или кластеров генов, при этом идентифицировано 8 новых генов, кодирующих белки, которые ранее считались псевдогенами. [ 32 ] Более ранний анализ показал, что PGAM4 человека (фосфоглицератмутаза), [ 33 ] Ранее считавшийся псевдогеном, он не только функционален, но и вызывает бесплодие в случае мутации. [ 34 ] [ 35 ]
Ряд псевдопсевдогенов был также обнаружен у прокариот, у которых, по-видимому, сохраняются некоторые замены стоп-кодонов в основных генах, даже при положительном отборе. [ 36 ] [ 37 ]
Некодирующий белок
[ редактировать ]миРНК . Некоторые эндогенные siRNA , по-видимому, происходят из псевдогенов, и, таким образом, некоторые псевдогены играют роль в регуляции транскриптов, кодирующих белки, как указано в обзоре. [ 38 ] Одним из многих примеров является psiPPM1K. Процессинг РНК, транскрибированных с psiPPM1K, дает миРНК, которые могут подавлять наиболее распространенный тип рака печени — гепатоцеллюлярную карциному . [ 39 ] Это и многие другие исследования вызвали значительный ажиотаж по поводу возможности воздействия на псевдогены с помощью/в качестве терапевтических агентов. [ 40 ]
пиРНК . Некоторые piRNA происходят из псевдогенов, расположенных в кластерах piRNA. [ 41 ] Эти piRNA регулируют гены через путь piRNA в семенниках млекопитающих и имеют решающее значение для ограничения мобильными элементами . повреждения генома [ 42 ]

микроРНК . Имеется множество сообщений о транскриптах псевдогенов, действующих как ловушки микроРНК . Возможно, самым ранним окончательным примером такого псевдогена, участвующего в развитии рака, является псевдоген BRAF . Ген BRAF — это протоонкоген , мутация которого связана со многими видами рака. В норме количество белка BRAF находится под контролем в клетках за счет действия микроРНК. В нормальных ситуациях количество РНК из BRAF и псевдогена BRAFP1 конкурируют за микроРНК, но баланс двух РНК таков, что клетки растут нормально. Однако когда экспрессия РНК BRAFP1 увеличивается (экспериментально или в результате естественных мутаций), меньше микроРНК доступно для контроля экспрессии BRAF, а повышенное количество белка BRAF вызывает рак. [ 43 ] Этот вид конкуренции за регуляторные элементы со стороны РНК, которые являются эндогенными по отношению к геному, привел к появлению термина ce РНК .
ПТЭН . Ген PTEN является известным геном-супрессором опухолей . Псевдоген PTEN, PTENP1, представляет собой процессированный псевдоген, который по своей генетической последовательности очень похож на ген дикого типа. Однако PTENP1 имеет миссенс-мутацию, которая устраняет кодон инициирующего метионина и, таким образом, предотвращает трансляцию нормального белка PTEN. [ 44 ] Несмотря на это, PTENP1, по-видимому, играет роль в онкогенезе . 3'- UTR мРНК PTENP1 действует как ловушка для мРНК PTEN, нацеливаясь на микроРНК из-за ее сходства с геном PTEN, а сверхэкспрессия 3'-UTR приводила к увеличению уровня белка PTEN. [ 45 ] То есть сверхэкспрессия 3'-UTR PTENP1 приводит к усилению регуляции и подавлению раковых опухолей. Биология этой системы по сути является противоположностью описанной выше системы BRAF.
Потогенес . Псевдогены могут в течение эволюционного времени участвовать в конверсии генов и других мутационных событиях, которые могут привести к появлению новых или вновь функциональных генов. Это привело к появлению концепции, согласно которой псевдогены можно рассматривать как горшечные гены : потенциальные гены для эволюционной диверсификации. [ 46 ]
Бактериальные псевдогены
[ редактировать ]Псевдогены встречаются у бактерий . [ 47 ] Большинство из них обнаружено у бактерий, которые не являются свободноживущими; то есть они либо симбионты , либо облигатные внутриклеточные паразиты . Таким образом, им не требуются многие гены, которые необходимы свободноживущим бактериям, например гены, связанные с метаболизмом и восстановлением ДНК. Однако не существует порядка, при котором функциональные гены теряются в первую очередь. Например, самые старые псевдогены у Mycobacterium leprae связаны с РНК-полимеразой и биосинтезом , вторичных метаболитов тогда как самые старые псевдогены у Shigella flexneri и Shigella typhi связаны с ДНК репликацией , рекомбинацией и репарацией . [ 48 ]
Поскольку большинство бактерий, несущих псевдогены, являются либо симбионтами, либо облигатными внутриклеточными паразитами, размер генома со временем уменьшается. Крайним примером является геном Mycobacterium leprae — облигатного паразита и возбудителя проказы . Сообщается, что он имеет 1133 псевдогена, которые дают начало примерно 50% его транскриптома . [ 48 ] Эффект псевдогенов и редукции генома можно дополнительно увидеть при сравнении с Mycobacterium marinum , патогеном из того же семейства. Mycobacteirum marinum имеет больший геном по сравнению с Mycobacterium leprae , поскольку может выживать вне хозяина; следовательно, геном должен содержать необходимые для этого гены. [ 49 ]
Хотя редукция генома фокусируется на том, какие гены не нужны, путем избавления от псевдогенов, селективное давление со стороны хозяина может повлиять на то, что останется. В случае симбионта из филума Verrucomicrobiota имеется семь дополнительных копий гена, кодирующего путь манделалида. [ 50 ] Хозяин, виды из Lissoclinum , используют манделалиды как часть своего защитного механизма. [ 50 ]
Связь между эпистазом и теорией потери генов «домино» наблюдалась у Buchnera aphidicola . Теория домино предполагает, что если один ген клеточного процесса инактивируется, то отбор других вовлеченных генов ослабевает, что приводит к потере генов. [ 48 ] При сравнении Buchnera aphidicola и Escherichia coli . было обнаружено, что положительный эпистаз способствует потере генов, а отрицательный эпистаз препятствует этому

В этой статье отсутствует информация об археях. ( декабрь 2023 г. ) |
См. также
[ редактировать ]- Список псевдогенов человека-инвалида
- Молекулярная эволюция
- Молекулярная палеонтология
- Псевдоген (база данных)
- ретропозон
- Ретротранспозон
Ссылки
[ редактировать ]- ^ Майгелл А.Дж., Смит Н.Р., Робинсон П.А., Маркхэм А.Ф. (февраль 2000 г.). «Псевдогены позвоночных» . Письма ФЭБС . 468 (2–3): 109–114. дои : 10.1016/S0014-5793(00)01199-6 . ПМИД 10692568 . S2CID 42204036 .
- ^ Робишо Б.М., Суско Э., Харриган А.М., Снайдер М. (февраль 2017 г.). «Гены рибосомальной РНК способствуют образованию псевдогенов и мусорной ДНК в геноме человека» . Геномная биология и эволюция . 9 (2): 380–397. дои : 10.1093/gbe/evw307 . ПМК 5381670 . ПМИД 28204512 .
- ^ ван Барен М.Дж., Брент М.Р. (май 2006 г.). «Итеративное предсказание генов и удаление псевдогенов улучшают аннотацию генома» . Геномные исследования . 16 (5): 678–685. дои : 10.1101/гр.4766206 . ПМК 1457044 . ПМИД 16651666 .
- ^ Ким М.С., Пинто С.М., Гетнет Д., Нируджоги Р.С., Манда С.С., Чаеркади Р. и др. (май 2014 г.). «Проект карты протеома человека» . Природа . 509 (7502): 575–581. Бибкод : 2014Natur.509..575K . дои : 10.1038/nature13302 . ПМЦ 4403737 . ПМИД 24870542 .
- ^ Макс Э.Э. (1986). «Ошибки плагиата и молекулярная генетика» . Журнал эволюции творения . 6 (3): 34–46.
- ^ Чандрасекаран С., Бетран Э. (2008). «Происхождение новых генов и псевдогенов» . Природное образование . 1 (1): 181.
- ^ Юрка Дж. (декабрь 2004 г.). «Эволюционное влияние повторяющихся элементов Alu человека». Текущее мнение в области генетики и развития . 14 (6): 603–608. дои : 10.1016/j.где.2004.08.008 . ПМИД 15531153 .
- ^ Деваннье М., Хайдман Т. (2005). «LINE, SINE и обработанные псевдогены: паразитические стратегии моделирования генома». Цитогенетические и геномные исследования . 110 (1–4): 35–48. дои : 10.1159/000084936 . ПМИД 16093656 . S2CID 25083962 .
- ^ Деваннье М., Эсно К., Хайдманн Т. (сентябрь 2003 г.). «LINE-опосредованная ретротранспозиция меченых последовательностей Alu». Природная генетика . 35 (1): 41–48. дои : 10.1038/ng1223 . ПМИД 12897783 . S2CID 32151696 .
- ^ Граур Д., Шуали Ю., Ли В.Х. (апрель 1989 г.). «Делеции в обработанных псевдогенах накапливаются у грызунов быстрее, чем у людей». Журнал молекулярной эволюции . 28 (4): 279–285. Бибкод : 1989JMolE..28..279G . дои : 10.1007/BF02103423 . ПМИД 2499684 . S2CID 22437436 .
- ^ Берч Р., Диканс М., Кент В.Дж., Хаусслер Д., Брозиус Дж. (октябрь 2008 г.). «Вклад ретрокопии в эволюцию человеческого генома» . БМК Геномика . 9 : 466. дои : 10.1186/1471-2164-9-466 . ПМК 2584115 . ПМИД 18842134 .
- ^ Павличек А., Пачес Дж., Зика Р., Хейнар Дж. (октябрь 2002 г.). «Распределение по длине длинных вкраплений нуклеотидных элементов (LINE) и процессированных псевдогенов эндогенных ретровирусов человека: значение для ретротранспозиции и обнаружения псевдогенов». Джин . 300 (1–2): 189–194. дои : 10.1016/S0378-1119(02)01047-8 . ПМИД 12468100 .
- ^ Наварро, Галанте, Пенсильвания (июль 2015 г.). «Общегеномный ландшафт ретрокопий в геномах приматов» . Геномная биология и эволюция . 7 (8): 2265–2275. дои : 10.1093/gbe/evv142 . ПМЦ 4558860 . ПМИД 26224704 .
- ^ Шрайдер Д.Р., Наварро ФК, Галанте П.А., Пармиджани Р.Б., Камарго А.А., Хан М.В., де Соуза С.Дж. (24 января 2013 г.). «Полиморфизм числа копий гена, вызванный ретротранспозицией у людей» . ПЛОС Генетика . 9 (1): e1003242. дои : 10.1371/journal.pgen.1003242 . ПМЦ 3554589 . ПМИД 23359205 .
- ^ Чжэн Д., Франкиш А., Берч Р., Капранов П., Реймонд А., Чу С.В. и др. (июнь 2007 г.). «Псевдогены в регионах ENCODE: консенсусная аннотация, анализ транскрипции и эволюция» . Геномные исследования . 17 (6): 839–851. дои : 10.1101/гр.5586307 . ЧВК 1891343 . ПМИД 17568002 .
- ^ Макс Э.Э. (5 мая 2003 г.). «Ошибки плагиата и молекулярная генетика» . Архив TalkOrigins . Проверено 22 июля 2008 г.
- ^ Перейти обратно: а б Линч М., Конери Дж.С. (ноябрь 2000 г.). «Эволюционная судьба и последствия дублирования генов». Наука . 290 (5494): 1151–1155. Бибкод : 2000Sci...290.1151L . дои : 10.1126/science.290.5494.1151 . ПМИД 11073452 .
- ^ Уолш Дж. Б. (январь 1995 г.). «Как часто дублированные гены приобретают новые функции?» . Генетика . 139 (1): 421–428. дои : 10.1093/генетика/139.1.421 . ПМЦ 1206338 . ПМИД 7705642 .
- ^ Линч М., О'Хели М., Уолш Б., Force A (декабрь 2001 г.). «Вероятность сохранения вновь возникшего дубликата гена» . Генетика . 159 (4): 1789–1804. дои : 10.1093/генетика/159.4.1789 . ПМЦ 1461922 . ПМИД 11779815 .
- ^ Харрисон П.М., Хеги Х., Баласубраманиан С., Ласкомб Н.М., Бертоне П., Эколс Н. и др. (февраль 2002 г.). «Молекулярные окаменелости в геноме человека: идентификация и анализ псевдогенов в хромосомах 21 и 22» . Геномные исследования . 12 (2): 272–280. дои : 10.1101/гр.207102 . ПМК 155275 . ПМИД 11827946 .
- ^ Чжан Дж (2003). «Эволюция путем дупликации генов: обновление». Тенденции экологии и эволюции . 18 (6): 292–298. дои : 10.1016/S0169-5347(03)00033-8 .
- ^ Нисикими М., Каваи Т., Яги К. (октябрь 1992 г.). «Морские свинки обладают сильно мутированным геном L-гулоно-гамма-лактоноксидазы, ключевого фермента биосинтеза L-аскорбиновой кислоты, отсутствующего у этого вида» . Журнал биологической химии . 267 (30): 21967–21972. дои : 10.1016/S0021-9258(19)36707-9 . ПМИД 1400507 .
- ^ Нишикими М., Фукуяма Р., Миношима С., Симидзу Н., Яги К. (май 1994 г.). «Клонирование и хромосомное картирование человеческого нефункционального гена L-гулоно-гамма-лактоноксидазы, фермента биосинтеза L-аскорбиновой кислоты, отсутствующего у человека» . Журнал биологической химии . 269 (18): 13685–13688. дои : 10.1016/S0021-9258(17)36884-9 . ПМИД 8175804 .
- ^ Сюэ Ю, Дейли А, Ингвадоттир Б, Лю М, Куп Г, Ким Ю и др. (апрель 2006 г.). «Распространение неактивной формы каспазы-12 у людей связано с недавним положительным отбором» . Американский журнал генетики человека . 78 (4): 659–670. дои : 10.1086/503116 . ПМЦ 1424700 . ПМИД 16532395 .
- ^ Читам С.В., Фолкнер Г.Дж., Динджер М.Э. (март 2020 г.). «Преодоление проблем и догм для понимания функций псевдогенов». Обзоры природы. Генетика . 21 (3): 191–201. дои : 10.1038/s41576-019-0196-1 . ПМИД 31848477 . S2CID 209393216 .
- ^ Зербино Д.Р., Франкиш А., Фличек П. (август 2020 г.). «Прогресс, проблемы и сюрпризы в аннотировании генома человека» . Ежегодный обзор геномики и генетики человека . 21 (1): 55–79. doi : 10.1146/annurev-genom-121119-083418 . ПМК 7116059 . ПМИД 32421357 .
- ^ Перейти обратно: а б Прието-Годино Л.Л., Ритц Р., Баржетон Б., Абуин Л., Аргуэлло Дж.Р., Пераро М.Д., Бентон Р. (ноябрь 2016 г.). «Псевдопсевдогены обонятельных рецепторов» . Природа . 539 (7627): 93–97. Бибкод : 2016Natur.539...93P . дои : 10.1038/nature19824 . ПМК 5164928 . ПМИД 27776356 .
- ^ Джеффс П., Эшбернер М. (май 1991 г.). «Процессированные псевдогены у дрозофилы». Слушания. Биологические науки . 244 (1310): 151–159. Бибкод : 1991RSPSB.244..151J . дои : 10.1098/rspb.1991.0064 . ПМИД 1679549 . S2CID 1665885 .
- ^ Ван В., Чжан Дж., Альварес С., Ллопарт А., Лонг М. (сентябрь 2000 г.). «Происхождение гена Jingwei и сложная модульная структура его родительского гена, желтого императора, у Drosophila melanogaster» . Молекулярная биология и эволюция . 17 (9): 1294–1301. doi : 10.1093/oxfordjournals.molbev.a026413 . ПМИД 10958846 .
- ^ Лонг М., Лэнгли CH (апрель 1993 г.). «Естественный отбор и происхождение цзинвэй, химерного процессированного функционального гена у дрозофилы». Наука . 260 (5104): 91–95. Бибкод : 1993Sci...260...91L . дои : 10.1126/science.7682012 . ПМИД 7682012 .
- ^ Пей Б., Сису С., Франкиш А., Ховальд С., Хабеггер Л., Му XJ и др. (сентябрь 2012 г.). «Ресурс псевдогена GENCODE» . Геномная биология . 13 (9): С51. дои : 10.1186/gb-2012-13-9-r51 . ПМЦ 3491395 . ПМИД 22951037 .
- ^ Райт Дж.К., Мадж Дж., Вайссер Х., Барзин М.П., Гонсалес Дж.М., Бразма А. и др. (июнь 2016 г.). «Улучшение аннотации эталонного гена GENCODE с использованием строгого рабочего процесса протеогеномики» . Природные коммуникации . 7 : 11778. Бибкод : 2016NatCo...711778W . дои : 10.1038/ncomms11778 . ПМЦ 4895710 . ПМИД 27250503 .
- ^ Дирик Х.А., Мерсер Дж.Ф., Гловер Т.В. (октябрь 1997 г.). «Псевдоген изоформы фосфоглицератмутазы головного мозга (PGAM 1) локализован в гене болезни Менкеса человека (ATP7 A)». Джин . 198 (1–2): 37–41. дои : 10.1016/s0378-1119(97)00289-8 . ПМИД 9370262 .
- ^ Бетран Э., Ван В., Джин Л., Лонг М. (май 2002 г.). «Эволюция гена, обработанного фосфоглицератмутазой, у человека и шимпанзе, раскрывающая происхождение нового гена приматов» . Молекулярная биология и эволюция . 19 (5): 654–663. doi : 10.1093/oxfordjournals.molbev.a004124 . ПМИД 11961099 .
- ^ Окуда Х., Цудзимура А., Ири С., Ямамото К., Фукухара С., Мацуока Ю. и др. (2012). «Один нуклеотидный полиморфизм в новом сцепленном с полом ретротранспонированном гене PGAM4, специфичном для семенников, влияет на мужскую фертильность человека» . ПЛОС ОДИН . 7 (5): e35195. Бибкод : 2012PLoSO...735195O . дои : 10.1371/journal.pone.0035195 . ПМЦ 3348931 . ПМИД 22590500 .
- ^ Белинский Ф., Гангулы И., Поляков Е., Юрченко В., Рогозин И.Б. (февраль 2021 г.). «Анализ стоп-кодонов в генах, кодирующих прокариотические белки, предполагает частые события чтения» . Международный журнал молекулярных наук . 22 (4): 1876. doi : 10.3390/ijms22041876 . ПМЦ 7918605 . PMID 33672790 .
- ^ Фэн Ю, Ван З, Чиен Кюй, Чен ХЛ, Лян ЮХ, Хуа Х, Чиу Ч. (май 2022 г.). « «Псевдопсевдогены» в бактериальных геномах: протеогеномика обнаруживает широкую, но низкую белковую экспрессию псевдогенов у Salmonella enterica» . Исследования нуклеиновых кислот . 50 (9): 5158–5170. дои : 10.1093/nar/gkac302 . ПМЦ 9122581 . ПМИД 35489061 .
- ^ Чан В.Л., Чанг Дж.Г. (2014). «Эндогенные миРНК псевдогенного происхождения и их функции». Псевдогены . Методы молекулярной биологии. Том. 1167. стр. 227–39. дои : 10.1007/978-1-4939-0835-6_15 . ISBN 978-1-4939-0834-9 . ПМИД 24823781 .
- ^ Чан В.Л., Юо С.И., Ян В.К., Хунг С.Ю., Чанг Ю.С., Чиу CC и др. (апрель 2013 г.). «Транскрибируемый псевдоген ψPPM1K генерирует эндогенную миРНК для подавления роста онкогенных клеток при гепатоцеллюлярной карциноме» . Исследования нуклеиновых кислот . 41 (6): 3734–3747. дои : 10.1093/нар/gkt047 . ПМК 3616710 . ПМИД 23376929 .
- ^ Робертс Т.К., Моррис К.В. (декабрь 2013 г.). «Уже не так псевдо: псевдогены как терапевтические мишени» . Фармакогеномика . 14 (16): 2023–2034. дои : 10.2217/стр.13.172 . ПМК 4068744 . ПМИД 24279857 .
- ^ Оловников И., Ле Томас А., Аравин А.А. (2014). «Схема манипулирования кластерами piRNA». PIWI-взаимодействующие РНК . Методы молекулярной биологии. Том. 1093. стр. 47–58. дои : 10.1007/978-1-62703-694-8_5 . ISBN 978-1-62703-693-1 . ПМИД 24178556 .
- ^ Сиоми MC, Сато К., Пезич Д., Аравин А.А. (апрель 2011 г.). «PIWI-взаимодействующие малые РНК: авангард защиты генома». Обзоры природы. Молекулярно-клеточная биология . 12 (4): 246–258. дои : 10.1038/nrm3089 . ПМИД 21427766 . S2CID 5710813 .
- ^ Каррет Ф.А., Решке М., Руокко А., Нг С., Чапуй Б., Леопольд В. и др. (апрель 2015 г.). «Псевдоген BRAF действует как конкурентная эндогенная РНК и индуцирует лимфому in vivo» . Ячейка 161 (2): 319–332. дои : 10.1016/j.cell.2015.02.043 . ПМК 6922011 . ПМИД 25843629 .
- ^ Дахия П.Л., Фитцджеральд М.Г., Чжан Х, Марш Д.Д., Чжэн З., Питч Т. и др. (май 1998 г.). «Высококонсервативный процессированный псевдоген PTEN расположен на участке хромосомы 9p21» . Онкоген . 16 (18): 2403–2406. дои : 10.1038/sj.onc.1201762 . ПМИД 9620558 .
- ^ Полисено Л., Салмена Л., Чжан Дж., Карвер Б., Хавеман В.Дж., Пандольфи П.П. (июнь 2010 г.). «Независимая от кодирования функция мРНК генов и псевдогенов регулирует биологию опухоли» . Природа . 465 (7301): 1033–1038. Бибкод : 2010Natur.465.1033P . дои : 10.1038/nature09144 . ПМК 3206313 . ПМИД 20577206 .
- ^ Балакирев Е.С., Аяла Ф.Дж. (2003). «Псевдогены: это «мусор» или функциональная ДНК?». Ежегодный обзор генетики . 37 : 123–151. дои : 10.1146/annurev.genet.37.040103.103949 . ПМИД 14616058 .
- ^ Гудхед I, Дарби AC (февраль 2015 г.). «Извлечение псевдогенов из псевдогенов» (PDF) . Современное мнение в микробиологии . 23 : 102–109. дои : 10.1016/j.mib.2014.11.012 . ПМИД 25461580 .
- ^ Перейти обратно: а б с Даган Т., Блехман Р., Граур Д. (февраль 2006 г.). «Теория домино» гибели генов: события постепенного и массового вымирания генов в трех линиях облигатных симбиотических бактериальных патогенов» . Молекулярная биология и эволюция . 23 (2): 310–316. дои : 10.1093/molbev/msj036 . ПМИД 16237210 .
- ^ Малхотра С., Ведити СК, Бланделл ТЛ (август 2017 г.). «Расшифровка сходств и различий между видами микобактерий» . PLOS Забытые тропические болезни . 11 (8): e0005883. дои : 10.1371/journal.pntd.0005883 . ПМЦ 5595346 . ПМИД 28854187 .
- ^ Перейти обратно: а б Лопера Дж., Миллер И.Дж., Макфэйл К.Л., Кван Дж.К. (21 ноября 2017 г.). «Увеличенная дозировка биосинтетических генов в защитном бактериальном симбионте с уменьшенным геномом» . mSystems . 2 (6): 1–18. дои : 10.1128/msystems.00096-17 . ПМК 5698493 . ПМИД 29181447 .
- ^ Коул С.Т., Эйглмайер К., Паркхилл Дж., Джеймс К.Д., Томсон Н.Р., Уиллер П.Р. и др. (февраль 2001 г.). «Массовый распад генов бациллы проказы». Природа . 409 (6823): 1007–1011. Бибкод : 2001Natur.409.1007C . дои : 10.1038/35059006 . ПМИД 11234002 . S2CID 4307207 .
Дальнейшее чтение
[ редактировать ]- Герштейн М., Чжэн Д. (август 2006 г.). «Настоящая жизнь псевдогенов». Научный американец . 295 (2): 48–55. Бибкод : 2006SciAm.295b..48G . doi : 10.1038/scientificamerican0806-48 . ПМИД 16866288 .
- Торрентс Д., Суяма М., Здобнов Е., Борк П. (декабрь 2003 г.). «Полногеномное исследование псевдогенов человека» . Геномные исследования . 13 (12): 2559–2567. дои : 10.1101/гр.1455503 . ПМК 403797 . ПМИД 14656963 .
- Бишоф Дж.М., Чанг А.П., Шитц Т.Е., Стоун Э.М., Казавант Т.Л., Шеффилд В.К., Браун Т.А. (июнь 2006 г.). «Полногеномная идентификация псевдогенов, способных вызывать конверсию генов, вызывающих заболевания» . Человеческая мутация . 27 (6): 545–552. дои : 10.1002/humu.20335 . ПМИД 16671097 . S2CID 20219423 .
- Сиберг-Олсен М.Дж., Гарбер А.И., Килинг П.Дж., Маккатчеон Дж.П., Хусник Ф. (июль 2022 г.). «Псевдоискатель: обнаружение псевдогенов в геномах прокариот» . Молекулярная биология и эволюция . 39 (7). дои : 10.1093/molbev/msac153 . ПМЦ 9336565 . ПМИД 35801562 .