ДНК-связывающий белок

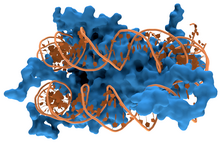


ДНК-связывающие белки — это белки , которые имеют ДНК-связывающие домены и, таким образом, обладают специфическим или общим сродством к одно- или двухцепочечной ДНК . [ 3 ] [ 4 ] [ 5 ] ДНК-связывающие белки, специфичные для последовательности, обычно взаимодействуют с большой бороздкой , B-ДНК поскольку она открывает больше функциональных групп , которые идентифицируют пару оснований . [ 6 ] [ 7 ]
Примеры
[ редактировать ]ДНК-связывающие белки включают факторы транскрипции , которые модулируют процесс транскрипции, различные полимеразы , нуклеазы , расщепляющие молекулы ДНК, и гистоны , которые участвуют в упаковке хромосом и транскрипции в ядре клетки . ДНК-связывающие белки могут включать в себя такие домены, как цинковый палец , спираль-поворот-спираль и лейциновую застежку (среди многих других), которые облегчают связывание с нуклеиновой кислотой. Есть и более необычные примеры, такие как активаторы транскрипции, подобные эффекторам .
Неспецифические ДНК-белковые взаимодействия
[ редактировать ]Структурные белки, связывающие ДНК, являются хорошо изученными примерами неспецифических взаимодействий ДНК-белок. Внутри хромосом ДНК содержится в комплексах со структурными белками. Эти белки организуют ДНК в компактную структуру, называемую хроматином . У эукариот эта структура включает связывание ДНК с комплексом небольших основных белков, называемых гистонами . У прокариот задействованы несколько типов белков. [ 8 ] [ 9 ] Гистоны образуют дискообразный комплекс, называемый нуклеосомой , который содержит два полных витка двухцепочечной ДНК, обернутых вокруг его поверхности. Эти неспецифические взаимодействия образуются посредством основных остатков в гистонах, образующих ионные связи с кислым сахарофосфатным остовом ДНК, и поэтому в значительной степени независимы от последовательности оснований. [ 10 ] Химические модификации этих основных аминокислотных остатков включают метилирование , фосфорилирование и ацетилирование . [ 11 ] Эти химические изменения изменяют силу взаимодействия между ДНК и гистонами, делая ДНК более или менее доступной для факторов транскрипции и изменяя скорость транскрипции. [ 12 ] Другие неспецифические ДНК-связывающие белки в хроматине включают белки группы высокой подвижности (HMG), которые связываются с изогнутой или искаженной ДНК. [ 13 ] Биофизические исследования показывают, что эти архитектурные белки HMG связывают, изгибают и образуют петли ДНК, выполняя ее биологические функции. [ 14 ] [ 15 ] Эти белки играют важную роль в изгибании массивов нуклеосом и их организации в более крупные структуры, образующие хромосомы. [ 16 ] Недавно было показано, что белок 25, связывающий FK506 (FBP25), неспецифически связывается с ДНК, что помогает в восстановлении ДНК. [ 17 ]
Белки, специфически связывающие одноцепочечную ДНК.
[ редактировать ]Отдельной группой ДНК-связывающих белков являются ДНК-связывающие белки, которые специфически связывают одноцепочечную ДНК. У людей репликационный белок А является наиболее изученным членом этого семейства и используется в процессах разделения двойной спирали, включая репликацию ДНК, рекомбинацию и репарацию ДНК. [ 18 ] Эти связывающие белки, по-видимому, стабилизируют одноцепочечную ДНК и защищают ее от образования петель или разрушения нуклеазами .
Связывание со специфическими последовательностями ДНК
[ редактировать ]Напротив, другие белки эволюционировали, чтобы связываться со специфическими последовательностями ДНК. Наиболее интенсивно изучаются различные факторы транскрипции — белки, регулирующие транскрипцию. Каждый фактор транскрипции связывается с одним конкретным набором последовательностей ДНК и активирует или ингибирует транскрипцию генов, которые имеют эти последовательности вблизи своих промоторов. Факторы транскрипции делают это двумя способами. Во-первых, они могут связываться с РНК-полимеразой, ответственной за транскрипцию, либо напрямую, либо через другие белки-медиаторы; это обнаруживает полимеразу у промотора и позволяет ей начать транскрипцию. [ 19 ] Альтернативно, факторы транскрипции могут связывать ферменты , которые модифицируют гистоны в промоторе. Это изменяет доступность матрицы ДНК для полимеразы. [ 20 ]
Эти мишени ДНК могут встречаться по всему геному организма. Таким образом, изменения активности одного типа транскрипционных факторов могут затронуть тысячи генов. [ 21 ] Таким образом, эти белки часто являются мишенями процессов передачи сигнала , которые контролируют реакцию на изменения окружающей среды или клеточную дифференцировку и развитие. Специфичность взаимодействия этих факторов транскрипции с ДНК обусловлена тем, что белки устанавливают множественные контакты с краями оснований ДНК, что позволяет им читать последовательность ДНК. Большинство этих взаимодействий с основаниями происходит в основной бороздке, где основания наиболее доступны. [ 22 ] Математическое описание связывания белок-ДНК с учетом специфичности последовательности, а также конкурентного и кооперативного связывания белков разных типов обычно выполняют с помощью решетчатых моделей . [ 23 ] Вычислительные методы для определения специфичности последовательности связывания ДНК были предложены для эффективного использования обильных данных о последовательностях в постгеномную эпоху. [ 24 ] Кроме того, достигнут прогресс в структурном прогнозировании специфичности связывания белковых семейств с использованием глубокого обучения. [ 25 ]
Взаимодействия белок-ДНК
[ редактировать ]Взаимодействия белок-ДНК происходят, когда связывает молекулу ДНК , часто для регулирования биологической функции ДНК, обычно экспрессии гена белок . Среди белков, которые связываются с ДНК, есть факторы транскрипции , которые активируют или подавляют экспрессию генов путем связывания с мотивами ДНК и гистонами , которые составляют часть структуры ДНК и связываются с ней менее специфично. белки, восстанавливающие ДНК, такие как урацил-ДНК-гликозилаза Также с ней тесно взаимодействуют .
Обычно белки связываются с ДНК в большой бороздке ; однако есть исключения. [ 26 ] Взаимодействие белок-ДНК бывает в основном двух типов: специфическое взаимодействие и неспецифическое взаимодействие. Недавние эксперименты с одиночными молекулами показали, что ДНК-связывающие белки подвергаются быстрому повторному связыванию, чтобы связаться в правильной ориентации для распознавания целевого сайта. [ 27 ]
Дизайн
[ редактировать ]Создание ДНК-связывающих белков, имеющих определенный сайт связывания ДНК, было важной целью биотехнологии. Белки с цинковыми пальцами были разработаны для связывания со специфическими последовательностями ДНК, и это является основой нуклеаз с цинковыми пальцами . Недавно эффекторные нуклеазы, подобные активатору транскрипции были созданы (TALEN), которые основаны на природных белках, секретируемых бактериями Xanthomonas через систему секреции типа III при заражении различных видов растений . [ 28 ]
Методы обнаружения
[ редактировать ]Существует множество методов in vitro и in vivo , которые полезны для обнаружения взаимодействий ДНК-белок. Ниже перечислены некоторые методы, используемые в настоящее время: [ 29 ] Анализ изменения электрофоретической подвижности (EMSA) является широко распространенным качественным методом изучения белок-ДНК-взаимодействий известных ДНК-связывающих белков. [ 30 ] [ 31 ] ДНК-белковое взаимодействие – иммуноферментный анализ (DPI-ELISA) позволяет качественно и количественно анализировать ДНК-связывающие предпочтения известных белков in vitro . [ 32 ] [ 33 ] Этот метод позволяет анализировать белковые комплексы, которые связываются с ДНК (DPI-Recruitment-ELISA) или подходит для автоматического скрининга нескольких нуклеотидных зондов благодаря стандартному формиату планшета ELISA. [ 34 ] [ 35 ] Анализ ДНКазного следа можно использовать для идентификации конкретных участков связывания белка с ДНК при разрешении пар оснований. [ 36 ] Иммунопреципитация хроматина используется для идентификации целевых областей ДНК in vivo известного фактора транскрипции. Этот метод в сочетании с высокопроизводительным секвенированием известен как ChIP-Seq , а в сочетании с микрочипами — как ChIP-чип . Дрожжевая одногибридная система (Y1H) используется для определения того, какой белок связывается с конкретным фрагментом ДНК. Бактериальная одногибридная система (B1H) используется для определения того, какой белок связывается с тем или иным фрагментом ДНК. Определение структуры с помощью рентгеновской кристаллографии использовалось для получения очень детального атомного представления о взаимодействиях белок-ДНК. другие методы, такие как SELEX, PBM (микрочипы, связывающие белки), скрининг микрочипов ДНК, DamID, FAIRE или, в последнее время, DAP-seq. Помимо этих методов, в лаборатории для исследования взаимодействия ДНК-белок in vivo и in vitro используются
Управление взаимодействиями
[ редактировать ]Взаимодействия белок-ДНК можно модулировать с помощью таких стимулов, как ионная сила буфера, скученность макромолекул, [ 27 ] температура, pH и электрическое поле. Это может привести к обратимой диссоциации/ассоциации комплекса белок-ДНК. [ 37 ] [ 38 ]
См. также
[ редактировать ]- домен bZIP
- ЧИП-экзо
- Сравнение программного обеспечения для моделирования нуклеиновых кислот
- ДНК-связывающий домен
- Спираль-петля-спираль
- Спираль-поворот-спираль
- HMG-коробка
- Лейциновая молния
- Лекситропсин (полусинтетический ДНК-связывающий лиганд)
- Дезоксирибонуклеопротеин
- Программное обеспечение для прогнозирования сайтов взаимодействия белок-ДНК
- РНК-связывающий белок
- Одноцепочечный связывающий белок
- Цинковый палец
Ссылки
[ редактировать ]- ^ Создано на основе PDB 1LMB.
- ^ Создано на основе PDB 1RVA.
- ^ Трэверс, А.А. (1993). ДНК-белковые взаимодействия . Лондон: Спрингер. ISBN 978-0-412-25990-6 .
- ^ Пабо CO, Sauer RT (1984). «Распознавание белка-ДНК». Анну. Преподобный Биохим . 53 (1): 293–321. дои : 10.1146/annurev.bi.53.070184.001453 . ПМИД 6236744 .
- ^ Дикерсон Р.Э. (1983). «Спираль ДНК и как ее читать». Sci Am . 249 (6): 94–111. Бибкод : 1983SciAm.249f..94D . doi : 10.1038/scientificamerican1283-94 .
- ^ Циммер С., Венерт Ю (1986). «Неинтеркалирующие ДНК-связывающие лиганды: специфика взаимодействия и их использование в качестве инструментов в биофизических, биохимических и биологических исследованиях генетического материала» . Прог. Биофиз. Мол. Биол . 47 (1): 31–112. дои : 10.1016/0079-6107(86)90005-2 . ПМИД 2422697 .
- ^ Дерван П.Б. (апрель 1986 г.). «Дизайн ДНК-связывающих молекул со специфичной последовательностью». Наука . 232 (4749): 464–71. Бибкод : 1986Sci...232..464D . дои : 10.1126/science.2421408 . ПМИД 2421408 .
- ^ Сэндман К., Перейра С., Рив Дж. (1998). «Разнообразие прокариотических хромосомных белков и происхождение нуклеосомы» . Cell Mol Life Sci . 54 (12): 1350–64. дои : 10.1007/s000180050259 . ПМЦ 11147202 . ПМИД 9893710 . S2CID 21101836 .
- ^ Дама RT (2005). «Роль нуклеоид-ассоциированных белков в организации и уплотнении бактериального хроматина». Мол. Микробиол . 56 (4): 858–70. дои : 10.1111/j.1365-2958.2005.04598.x . ПМИД 15853876 .
- ^ Люгер К., Мэдер А., Ричмонд Р., Сарджент Д., Ричмонд Т. (1997). «Кристаллическая структура ядра нуклеосомы при разрешении 2,8 А». Природа . 389 (6648): 251–60. Бибкод : 1997Natur.389..251L . дои : 10.1038/38444 . ПМИД 9305837 . S2CID 4328827 .
- ^ Дженувейн Т., Эллис С. (2001). «Перевод гистонового кода». Наука . 293 (5532): 1074–80. CiteSeerX 10.1.1.453.900 . дои : 10.1126/science.1063127 . ПМИД 11498575 . S2CID 1883924 .
- ^ Ито Т (2003). «Сборка и ремоделирование нуклеосом». Белковые комплексы, модифицирующие хроматин . Актуальные темы микробиологии и иммунологии. Том. 274. стр. 1–22. дои : 10.1007/978-3-642-55747-7_1 . ISBN 978-3-642-62909-9 . ПМИД 12596902 .
{{cite book}}:|journal=игнорируется ( помогите ) - ^ Томас Дж (2001). «HMG1 и 2: архитектурные ДНК-связывающие белки». Биохим Соц Транс . 29 (Часть 4): 395–401. дои : 10.1042/BST0290395 . ПМИД 11497996 .
- ^ Муругесапиллай, Дивакаран; МакКоли, Мика Дж.; Хо, Ран; Нельсон Холт, Молли Х.; Степанянц, Армен; Махер, Л. Джеймс; Исраэлофф, Натан Э.; Уильямс, Марк К. (2014). «Соединение ДНК и образование петель с помощью HMO1 обеспечивает механизм стабилизации безнуклеосомного хроматина» . Исследования нуклеиновых кислот . 42 (14): 8996–9004. дои : 10.1093/nar/gku635 . ПМЦ 4132745 . ПМИД 25063301 .
- ^ Муругесапиллай, Дивакаран; МакКоли, Мика Дж.; Махер, Л. Джеймс; Уильямс, Марк К. (2017). «Одномолекулярные исследования высокомобильных белков, изгибающих архитектурную ДНК группы В» . Биофизические обзоры . 9 (1): 17–40. дои : 10.1007/s12551-016-0236-4 . ПМК 5331113 . ПМИД 28303166 .
- ^ Гроссшедль Р., Гизе К., Пейджел Дж. (1994). «Белки домена HMG: архитектурные элементы сборки нуклеопротеиновых структур». Тенденции Жене . 10 (3): 94–100. дои : 10.1016/0168-9525(94)90232-1 . ПМИД 8178371 .
- ^ Пракаш, Аджит; Шин, Джун; Раджан, Шрикант; Юн, Хо Соп (7 апреля 2016 г.). «Структурные основы распознавания нуклеиновых кислот FK506-связывающим белком 25 (FKBP25), ядерным иммунофилином» . Исследования нуклеиновых кислот . 44 (6): 2909–2925. дои : 10.1093/nar/gkw001 . ISSN 0305-1048 . ПМЦ 4824100 . ПМИД 26762975 .
- ^ Ифтоде С., Дэниели Ю., Боровец Дж. (1999). «Репликационный белок A (RPA): эукариотический SSB». Crit Rev Biochem Mol Biol . 34 (3): 141–80. дои : 10.1080/10409239991209255 . ПМИД 10473346 .
- ^ Майерс Л., Корнберг Р. (2000). «Медиатор регуляции транскрипции». Анну Рев Биохим . 69 (1): 729–49. doi : 10.1146/annurev.biochem.69.1.729 . ПМИД 10966474 .
- ^ Шпигельман Б., Генрих Р. (2004). «Биологический контроль посредством регулируемых коактиваторов транскрипции» . Клетка . 119 (2): 157–67. дои : 10.1016/j.cell.2004.09.037 . ПМИД 15479634 . S2CID 14668705 .
- ^ Ли З, Ван Калкар С, Цюй С, Кавени В, Чжан М, Рен Б (2003). «Глобальная регуляторная роль транскрипции c-Myc в клетках лимфомы Беркитта» . Proc Natl Acad Sci США . 100 (14): 8164–9. Бибкод : 2003PNAS..100.8164L . дои : 10.1073/pnas.1332764100 . ПМК 166200 . ПМИД 12808131 .
- ^ Пабо С., Зауэр Р. (1984). «Распознавание белка-ДНК». Анну Рев Биохим . 53 (1): 293–321. дои : 10.1146/annurev.bi.53.070184.001453 . ПМИД 6236744 .
- ^ Тейф В.Б.; Риппе К. (2010). «Статистически-механические решетчатые модели связывания белка с ДНК в хроматине». Физический журнал: конденсированное вещество . 22 (41): 414105. arXiv : 1004.5514 . Бибкод : 2010JPCM...22O4105T . дои : 10.1088/0953-8984/22/41/414105 . ПМИД 21386588 . S2CID 103345 .
- ^ Вонг К.С., Чан ТМ, Пэн С., Ли Ю, Чжан З (2013). «Выяснение мотивов ДНК с использованием распространения убеждений» . Исследования нуклеиновых кислот . 41 (16): е153. дои : 10.1093/нар/gkt574 . ПМЦ 3763557 . ПМИД 23814189 .
- ^ Митра, Рактим; Ли, Джинсен; Сагендорф, Джаред М.; Цзян, Ибэй; Коэн, Ари С.; Чиу, Цу-Пей; Гласскок, Кэмерон Дж.; Рос, Ремо (5 августа 2024 г.). «Геометрическое глубокое изучение специфичности связывания белок-ДНК» . Природные методы . дои : 10.1038/s41592-024-02372-w . ISSN 1548-7091 .
- ^ Бьюли К.А., Гроненборн А.М., Клор Г.М. (1998). «Архитектурные белки, связывающие малые бороздки: структура, функции и распознавание ДНК» . Annu Rev Biophys Biomol Struct . 27 : 105–31. doi : 10.1146/annurev.biophys.27.1.105 . ПМЦ 4781445 . ПМИД 9646864 .
- ^ Jump up to: а б Ганджи, Махипал; Доктер, Маргрит; Ле Грайс, Стюарт Ф.Дж.; Аббонданциери, Элио А. (30 сентября 2016 г.). «ДНК-связывающие белки исследуют множество локальных конфигураций во время стыковки посредством быстрого повторного связывания» . Исследования нуклеиновых кислот . 44 (17): 8376–8384. дои : 10.1093/nar/gkw666 . ISSN 0305-1048 . ПМК 5041478 . ПМИД 27471033 .
- ^ Кларк К.Дж., Войтас Д.Ф., Эккер СК (сентябрь 2011 г.). «СКАЗКА о двух нуклеазах: нацеливание генов на массы?» . Рыбка данио . 8 (3): 147–9. дои : 10.1089/zeb.2011.9993 . ПМК 3174730 . ПМИД 21929364 .
- ^ Цай Ю., Хуан Х. (июль 2012 г.). «Достижения в изучении взаимодействия белка и ДНК». Аминокислоты . 43 (3): 1141–6. дои : 10.1007/s00726-012-1377-9 . ПМИД 22842750 . S2CID 310256 .
- ^ Фрид М., Кротерс Д.М. (1981). «Равновесия и кинетика lac-репрессорно-операторных взаимодействий при электрофорезе в полиакриламидном геле» . Нуклеиновые кислоты Рез . 9 (23): 6505–6525. дои : 10.1093/нар/9.23.6505 . ПМК 327619 . ПМИД 6275366 .
- ^ Гарнер М.М., Ревзин А. (1981). «Метод гель-электрофореза для количественной оценки связывания белков со специфическими участками ДНК: применение к компонентам регуляторной системы лактозного оперона Escherichia coli» . Нуклеиновые кислоты Рез . 9 (13): 3047–3060. дои : 10.1093/нар/9.13.3047 . ПМК 327330 . ПМИД 6269071 .
- ^ Брэнд Л.Х., Кирхлер Т., Хаммель С., Чабан С., Ванке Д. (2010). «DPI-ELISA: быстрый и универсальный метод определения связывания факторов транскрипции растений с ДНК in vitro» . Растительные методы . 25 (6): 25. дои : 10.1186/1746-4811-6-25 . ПМЦ 3003642 . ПМИД 21108821 .
- ^ Фишер С.М., Бёзер А., Хирш Дж.П., Ванке Д. (2016). «Количественный анализ взаимодействия белка и ДНК с помощью qDPI-ELISA». Растительные синтетические промоторы . Методы Мол. Биол. Том. 1482. стр. 49–66. дои : 10.1007/978-1-4939-6396-6_4 . ISBN 978-1-4939-6394-2 . ПМИД 27557760 .
- ^ Хекер А., Бранд Л.Х., Питер С., Симончелло Н., Килиан Дж., Хартер К., Годен В., Ванке Д. (2015). «ГАГА-связывающий фактор арабидопсиса БАЗОВЫЙ ПЕНТАЦИСТЕИН6 привлекает компонент ПОЛИКОМБ-РЕПРЕССИВНЫЙ КОМПЛЕКС1, КАК ГЕТЕРОХРОМАТИНОВЫЙ БЕЛОК1, к мотивам ДНК ГАГА» . Физиол растений . 163 (3): 1013–1024. дои : 10.1104/стр.15.00409 . ПМЦ 4741334 . ПМИД 26025051 .
- ^ Брэнд Л.Х., Хеннегес С., Шюсслер А., Колукисаоглу Хю, Кох Г., Вальмерот Н., Хеккер А., Туроу К., Зелл А., Хартер К., Ванке Д. (2013). «Скрининг взаимодействий белок-ДНК с помощью автоматизированного ИФА на взаимодействие ДНК-белок» . ПЛОС ОДИН . 8 (10): е75177. Бибкод : 2013PLoSO...875177B . дои : 10.1371/journal.pone.0075177 . ПМЦ 3795721 . ПМИД 24146751 .
- ^ Гала-диджей, Шмитц А (1978). «Отпечаток ДНКазы: простой метод определения специфичности связывания белок-ДНК» . Нуклеиновые кислоты Рез . 5 (9): 3157–3170. дои : 10.1093/нар/5.9.3157 . ПМЦ 342238 . ПМИД 212715 .
- ^ Хианик Т., Ван Дж (2009). «Электрохимические аптасенсоры – последние достижения и перспективы». Электроанализ . 21 (11): 1223–1235. дои : 10.1002/elan.200904566 .
- ^ Госай А. и др. (2016). «Связывание/рассвязывание комплекса тромбин-аптамер человека, контролируемое электрическим стимулом» . наук. Представитель . 6 : 37449. Бибкод : 2016NatSR...637449G . дои : 10.1038/srep37449 . ПМК 5118750 . ПМИД 27874042 .
Внешние ссылки
[ редактировать ]- Связывание белок-ДНК: данные, инструменты и модели (аннотированный список, постоянно обновляемый)
- Инструмент Abalone для моделирования взаимодействий ДНК-лиганд.
- База данных предсказанных факторов транскрипции DBD. Использует тщательно подобранный набор ДНК-связывающих доменов для прогнозирования факторов транскрипции во всех полностью секвенированных геномах.
- ДНК-связывание + белки Национальной медицинской библиотеки США по медицинским предметным рубрикам (MeSH)