Саркомер
| Саркомер | |
|---|---|
 Изображение саркомера | |
| Подробности | |
| Часть | Поперечно-полосатая мышца |
| Идентификаторы | |
| латинский | саркомер |
| МеШ | Д012518 |
| ТД | Х2.00.05.0.00008 |
| ФМА | 67895 |
| Анатомические термины микроанатомии | |
Саркомер (греч . σάρξ sarx «плоть», μέρος meros «часть») — наименьшая функциональная единица поперечно-полосатой мышечной ткани . [ 1 ] Это повторяющаяся единица между двумя Z-линиями. Скелетные мышцы состоят из трубчатых мышечных клеток (называемых мышечными волокнами или миофибрами), которые образуются во время эмбрионального миогенеза . Мышечные волокна содержат многочисленные трубчатые миофибриллы . Миофибриллы состоят из повторяющихся участков саркомеров, которые под микроскопом выглядят как чередующиеся темные и светлые полосы. Саркомеры состоят из длинных волокнистых белков в виде нитей, которые скользят друг мимо друга, когда мышца сокращается или расслабляется. Костамер — это другой компонент, который соединяет саркомер с сарколеммой .
Двумя важными белками являются миозин , образующий толстую нить, и актин , образующий тонкую нить. Миозин имеет длинный волокнистый хвост и шаровидную головку, которая связывается с актином. также Головка миозина связывается с АТФ , который является источником энергии для движения мышц. Миозин может связываться с актином только тогда, когда места связывания актина подвергаются воздействию ионов кальция.
Молекулы актина связаны с Z-линией, которая образует границы саркомера. Другие полосы появляются, когда саркомер расслаблен. [ 2 ]
Миофибриллы гладкомышечных клеток не образуют саркомеры.
Группы
[ редактировать ]Саркомеры придают скелетным и сердечным мышцам вид поперечно-полосатый . [ 2 ] который впервые описал Ван Левенгук . [ 3 ]
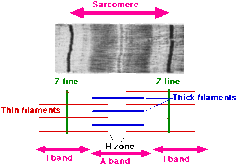
- Саркомер определяется как сегмент между двумя соседними Z-линиями (или Z-дисками). На электронных микрофотографиях поперечно-полосатой мышцы Z-линия (от немецкого «zwischen», что означает «между » ) появляется между I-полосами в виде темной линии, которая закрепляет актиновые миофиламенты.
- Вокруг Z-линии находится область I-полосы (для изотропного ). I-диапазон — зона тонких нитей, не перекрытая толстыми нитями (миозином).
- За I-диапазоном следует A-диапазон ( анизотропный ). Назван в честь своих свойств под микроскопом в поляризованном свете . A-диапазон содержит всю длину одной толстой нити. Анизотропная полоса содержит как толстые, так и тонкие нити.
- Внутри А-диапазона находится более бледная область, называемая Н-зоной (от немецкого «heller» — более яркий ). Названы в честь их более светлого вида под поляризационным микроскопом. H-диапазон — это зона толстых филаментов, не содержащая актина.
- Внутри Н-зоны находится тонкая М-линия (от немецкого «mittel», что означает «середина » ), возникающая посередине саркомера, образованная из поперечно соединяющихся элементов цитоскелета.
Взаимоотношения между белками и участками саркомера следующие:
- Актиновые нити, тонкие нити, являются основным компонентом I-диапазона и доходят до А-диапазона.
- Миозиновые нити, толстые нити, являются биполярными и простираются по всему А-диапазону. В центре они сшиты М-полосой.
- Гигантский белок тайтин (коннектин) простирается от Z-линии саркомера, где он связывается с системой толстых филаментов (миозина), до М-полосы, где, как полагают, он взаимодействует с толстыми нитями. Титин (и его сплайс-изоформы) — это самый крупный высокоэластичный белок, встречающийся в природе. Он обеспечивает места связывания для многочисленных белков и, как полагают, играет важную роль в качестве линейки саркомеров и в качестве образца для сборки саркомера.
- Предполагается, что другой гигантский белок, небулин , распространяется вдоль тонких нитей и всего I-диапазона. Считается, что, как и тайтин, он действует как молекулярная линейка при сборке тонких нитей.
- Несколько белков, важных для стабильности структуры саркомера, обнаружены как в Z-линии, так и в М-полосе саркомера.
- Актиновые нити и молекулы тайтина сшиты в Z-диске посредством белка Z-линии альфа-актинина.
- Белки М-диапазона миомезин, а также С-белок сшивают систему толстых филаментов (миозины) и часть М-диапазона тайтина (эластические нити).
- М-линия также связывает креатинкиназу, которая облегчает реакцию АДФ и фосфокреатина с образованием АТФ и креатина.
- Взаимодействие между актиновыми и миозиновыми нитями в А-диапазоне саркомера отвечает за сокращение мышц (на основе модели скользящих нитей ). [ 2 ]
Сокращение
[ редактировать ]Белок тропомиозин покрывает миозин-связывающие участки молекул актина в мышечной клетке. Чтобы мышечная клетка сократилась, тропомиозин должен переместиться, чтобы открыть места связывания актина. Ионы кальция связываются с молекулами тропонина С (которые рассеяны по белку тропомиозина) и изменяют структуру тропомиозина, заставляя его обнажить сайт связывания поперечных мостиков на актине.
Концентрация кальция в мышечных клетках контролируется саркоплазматическим ретикулумом , уникальной формой эндоплазматического ретикулума в саркоплазме .
Мышечные клетки стимулируются, когда двигательный нейрон высвобождает нейротрансмиттер ацетилхолин , который проходит через нервно-мышечное соединение (синапс между конечной кнопкой нейрона и мышечной клеткой). Ацетилхолин связывается с постсинаптическим никотиновым ацетилхолиновым рецептором . Изменение конформации рецептора обеспечивает приток ионов натрия и инициацию постсинаптического потенциала действия . Затем потенциал действия перемещается по Т-трубочкам (поперечным канальцам), пока не достигнет саркоплазматического ретикулума. Здесь деполяризованная мембрана активирует потенциалзависимые кальциевые каналы L-типа , присутствующие в плазматической мембране. Кальциевые каналы L-типа тесно связаны с рианодиновыми рецепторами, присутствующими на саркоплазматическом ретикулуме. Поток кальция внутрь из кальциевых каналов L-типа активирует рианодиновые рецепторы, высвобождая ионы кальция из саркоплазматического ретикулума. Этот механизм называется кальций-индуцированным высвобождением кальция (CICR). Неясно, вызывает ли открытие рианодиновых рецепторов физическое открытие кальциевых каналов L-типа или присутствие кальция. Отток кальция обеспечивает головкам миозина доступ к местам связывания поперечных мостиков актина, что обеспечивает сокращение мышц. [ 4 ]
Сокращение мышц заканчивается, когда ионы кальция закачиваются обратно в саркоплазматический ретикулум, позволяя сократительному аппарату и, следовательно, мышечным клеткам расслабиться.
При сокращении мышц А-диски не меняют своей длины (1,85 микрометра в скелетных мышцах млекопитающих). [ 4 ] тогда как I-диапазоны и H-зона укорачиваются. Это заставляет Z-линии сближаться.
Отдых
[ редактировать ]В состоянии покоя головка миозина связана с молекулой АТФ в низкоэнергетической конфигурации и не может получить доступ к сайтам связывания поперечных мостиков актина. Однако головка миозина может гидролизовать АТФ до аденозиндифосфата (АДФ) и неорганического фосфат-иона. Часть энергии, выделяющаяся при этой реакции, изменяет форму головки миозина и переводит ее в высокоэнергетическую конфигурацию. В процессе связывания с актином головка миозина высвобождает АДФ и неорганический ион фосфата, изменяя свою конфигурацию обратно на низкоэнергетическую. Миозин остается прикрепленным к актину в состоянии, известном как строгость , до тех пор, пока новый АТФ не свяжет головку миозина. Это связывание АТФ с миозином высвобождает актин путем диссоциации поперечных мостиков. АТФ-ассоциированный миозин готов к следующему циклу, начинающемуся с гидролиза АТФ.
Диапазон А виден в виде темных поперечных линий, пересекающих миофибриллы; I-полоса видна как слегка окрашенные поперечные линии, а Z-линия видна как темные линии, разделяющие саркомеры на уровне светового микроскопа.
Хранение энергии
[ редактировать ]Большинство мышечных клеток могут хранить достаточное количество АТФ только для небольшого количества мышечных сокращений. Хотя мышечные клетки также хранят гликоген , большая часть энергии, необходимой для сокращения, поступает из фосфагенов . Один из таких фосфагенов, креатинфосфат , используется для обеспечения АДФ фосфатной группой для синтеза АТФ у позвоночных . [ 4 ]
Сравнительная структура
[ редактировать ]Структура саркомера влияет на его функцию несколькими способами. Перекрытие актина и миозина приводит к появлению кривой длины-напряжения , которая показывает, как выходная сила саркомера уменьшается, если мышца растягивается так, что меньше поперечных мостиков может образовываться или сжиматься до тех пор, пока актиновые нити не начнут мешать друг другу. Длина актиновых и миозиновых нитей (в совокупности длина саркомера) влияет на силу и скорость: более длинные саркомеры имеют больше поперечных мостиков и, следовательно, большую силу, но имеют меньший диапазон укорочения. У позвоночных наблюдается очень ограниченный диапазон длин саркомеров с примерно одинаковой оптимальной длиной (длина при пиковом напряжении) во всех мышцах индивидуума, а также между видами. Однако у членистоногих наблюдаются огромные различия (более чем в семь раз) в длине саркомеров как между видами, так и между мышцами у одной особи. Причины отсутствия значительной изменчивости саркомеров у позвоночных до конца не известны. [ нужна ссылка ]
Ссылки
[ редактировать ]- ^ Бига, Линдси М.; Доусон, Сьерра; Харвелл, Эми (2019). «10.2 Скелетные мышцы» . Анатомия и физиология . OpenStax/Университет штата Орегон . Проверено 22 мая 2021 г.
- ^ Jump up to: а б с Рис, Джейн; Кэмпбелл, Нил (2002). Биология . Сан-Франциско: Бенджамин Каммингс . ISBN 0-8053-6624-5 .
- ^ Мартоноси, АН (1 января 2000 г.). «Животное электричество, Ca2+ и сокращение мышц. Краткая история исследований мышц» . Акта Биохимика Полоника . 47 (3): 493–516. дои : 10.18388/abp.2000_3974 . ISSN 0001-527X . ПМИД 11310955 .
- ^ Jump up to: а б с Либер (2002). Структура, функция и пластичность скелетных мышц: физиологические основы реабилитации (2-е изд.). Липпинкотт Уильямс и Уилкинс. ISBN 978-0781730617 .
Внешние ссылки
[ редактировать ]- MBInfo: Саркомер
- MBInfo: Сократительное волокно
- Видео о мышечных тканях
- Гистологическое изображение: 21601ooa - Система обучения гистологии Бостонского университета - «Ультраструктура клетки: саркоплазма скелетных мышц»
- Medical Mnemonics .com : 50 379 107
- Изображения, созданные антителами к полосам
- Сокращение мышц для чайников
- Модельное изображение саркомера