Т-трубочка
| Т-трубочка | |
|---|---|
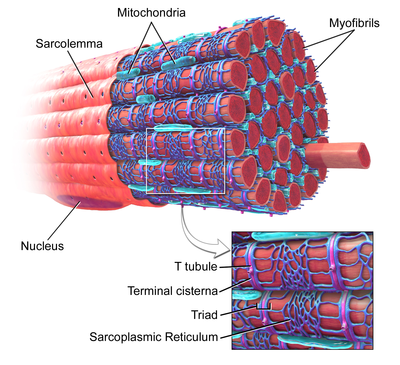 Скелетные мышечные волокна с обозначенными Т-трубочками в увеличенном изображении. | |
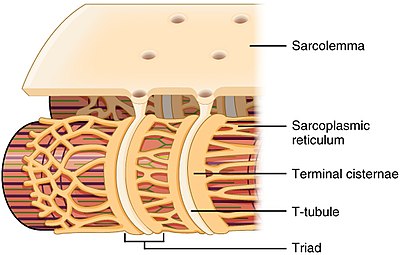 Структура Т-канальцев и связь с саркоплазматической сетью в скелетных мышцах | |
| Подробности | |
| Часть | Клеточная мембрана клеток скелетных и сердечных мышц |
| Идентификаторы | |
| латинский | поперечный каналец |
| ТД | Х2.00.05.2.01018, Х2.00.05.2.02013 |
| Анатомическая терминология | |
Т-трубочки ( поперечные канальцы ) представляют собой расширения клеточной мембраны , проникающие в центр клеток скелетных и сердечных мышц . Благодаря мембранам, которые содержат большие концентрации ионных каналов , транспортеров и насосов, Т-трубочки обеспечивают быструю передачу потенциала действия в клетку, а также играют важную роль в регулировании концентрации клеточного кальция.
Благодаря этим механизмам Т-трубочки позволяют клеткам сердечной мышцы сокращаться более сильно, синхронизируя высвобождение кальция из саркоплазматической сети по всей клетке. [1] На структуру и функцию Т-канальцев каждое сокращение кардиомиоцитов влияет. [2] а также заболеваниями, потенциально способствующими сердечной недостаточности и аритмиям . Хотя эти структуры были впервые обнаружены в 1897 году, исследования биологии Т-трубочек продолжаются.
Структура
[ редактировать ]Т-трубочки представляют собой канальцы, образованные из того же фосфолипидного бислоя, что и поверхностная мембрана или сарколемма клеток скелетных или сердечных мышц. [1] Они соединяются непосредственно с сарколеммой на одном конце, а затем перемещаются глубоко внутрь клетки, образуя сеть канальцев с секциями, идущими как перпендикулярно (поперечно), так и параллельно (аксиально) сарколемме. [1] Из-за этой сложной ориентации некоторые называют Т-трубочки поперечно-осевой трубчатой системой. [3] Внутренняя часть или просвет Т-трубочек открыта на поверхности клетки, а это означает, что Т-трубочки заполнены жидкостью, содержащей те же компоненты, что и раствор, окружающий клетку (внеклеточная жидкость). Мембрана, образующая Т-канальцы, является не просто пассивной соединительной трубкой, а очень активной: она усеяна белками, включая кальциевые каналы L-типа , натрий-кальциевые обменники , кальциевые АТФазы и бета-адренорецепторы . [1]
Т-трубочки обнаруживаются как в предсердий , так и в желудочках клетках сердечной мышцы ( кардиомиоцитах ), в которых они развиваются в первые несколько недель жизни. [4] Они обнаружены в мышечных клетках желудочков у большинства видов и в клетках предсердных мышц у крупных млекопитающих. [5] В клетках сердечной мышцы у разных видов Т-трубочки имеют диаметр от 20 до 450 нанометров и обычно расположены в областях, называемых Z-дисками , где актиновые миофиламенты закрепляются внутри клетки. [1] Т-трубочки в сердце тесно связаны с внутриклеточными запасами кальция, известными как саркоплазматический ретикулум, в определенных областях, называемых терминальными цистернами. Соединение Т-трубочек с терминальной цистерной известно как диада . [6]
В клетках скелетных мышц Т-трубочки в три-четыре раза уже, чем в клетках сердечной мышцы, и имеют диаметр от 20 до 40 нм. [1] Обычно они расположены по обе стороны от полоски миозина, в месте перекрытия (соединение AI) между полосами A и I. [7] Т-трубочки в скелетных мышцах связаны с двумя терминальными цистернами , известными как триада . [1] [8]
Регуляторы
[ редактировать ]Форма системы Т-трубочек создается и поддерживается множеством белков. Белок амфифизин-2 кодируется геном BIN1 и отвечает за формирование структуры Т-трубочек и обеспечение расположения соответствующих белков (в частности, кальциевых каналов L-типа) внутри мембраны Т-трубочек. [9] Юнктофилин-2 кодируется геном JPH2 и помогает сформировать соединение между мембраной Т-канальцев и саркоплазматическим ретикулумом, жизненно важное для связи возбуждения-сокращения . [6] Белок, закрывающий титин, известный как телетонин, кодируется геном TCAP и помогает развитию Т-канальцев и потенциально ответственен за увеличение количества Т-канальцев, наблюдаемое по мере роста мышц. [6]
Функция
[ редактировать ]Связь возбуждения-сокращения
[ редактировать ]Т-трубочки являются важным звеном в цепи от электрического возбуждения клетки до ее последующего сокращения (связка возбуждения-сокращения). Когда необходимо сокращение мышцы, стимуляция нерва или соседней мышечной клетки вызывает характерный поток заряженных частиц через клеточную мембрану, известный как потенциал действия . В состоянии покоя на внутренней стороне мембраны меньше положительно заряженных частиц по сравнению с внешней стороной, и мембрана считается поляризованной. Во время потенциала действия положительно заряженные частицы (преимущественно ионы натрия и кальция) проходят через мембрану снаружи внутрь. Это меняет нормальный дисбаланс заряженных частиц и называется деполяризацией . Одна область мембраны деполяризует соседние области, и возникающая волна деполяризации затем распространяется вдоль клеточной мембраны. [10] Поляризация мембраны восстанавливается, когда ионы калия возвращаются через мембрану изнутри наружу клетки.
В клетках сердечной мышцы, когда потенциал действия проходит по Т-канальцам, он активирует кальциевые каналы L-типа в мембране Т-канальцев. Активация кальциевого канала L-типа позволяет кальцию проникать в клетку. Т-трубочки содержат более высокую концентрацию кальциевых каналов L-типа, чем остальная часть сарколеммы, и поэтому большая часть кальция, поступающего в клетку, происходит через Т-трубочки. [11] Этот кальций связывается и активирует рецептор, известный как рианодиновый рецептор , расположенный в собственном внутреннем хранилище кальция клетки, саркоплазматическом ретикулуме. Активация рианодинового рецептора вызывает высвобождение кальция из саркоплазматического ретикулума, вызывая сокращение мышечных клеток. [12] Однако в клетках скелетных мышц кальциевый канал L-типа непосредственно прикреплен к рианодиновому рецептору на саркоплазматическом ретикулуме, что позволяет активировать рианодиновый рецептор напрямую, без необходимости притока кальция. [13]
Важность Т-трубочек обусловлена не только концентрацией в них кальциевых каналов L-типа, но и их способностью синхронизировать высвобождение кальция внутри клетки. Быстрое распространение потенциала действия по сети Т-канальцев активирует все кальциевые каналы L-типа почти одновременно. Поскольку Т-трубочки приближают сарколемму к саркоплазматической сети во всех областях клетки, кальций может затем высвобождаться из саркоплазматической сети одновременно по всей клетке. Эта синхронизация высвобождения кальция позволяет мышечным клеткам сокращаться более сильно. [14] В клетках, лишенных Т-трубочек, таких как гладкомышечные клетки , больные кардиомиоциты или мышечные клетки, в которых Т-трубочки были искусственно удалены, кальций, поступающий через сарколемму, должен постепенно диффундировать по всей клетке, активируя рианодиновые рецепторы гораздо медленнее. как волна кальция, приводящая к менее сильному сокращению. [14]
Поскольку Т-канальцы являются основным местом взаимодействия возбуждения и сокращения, здесь сосредоточены ионные каналы и белки, участвующие в этом процессе: внутри мембраны Т-канальцев расположено в 3 раза больше кальциевых каналов L-типа по сравнению с остальными. из сарколеммы. Кроме того, бета-адренорецепторы также имеют высокую концентрацию в мембране Т-канальцев. [15] и их стимуляция увеличивает высвобождение кальция из саркоплазматической сети. [16]
Контроль кальция
[ редактировать ]Поскольку пространство внутри просвета Т-трубочек непрерывно с пространством, окружающим клетку (внеклеточным пространством), концентрации ионов между ними очень похожи. Однако из-за важности ионов внутри Т-канальцев (особенно кальция в сердечной мышце) очень важно, чтобы эти концентрации оставались относительно постоянными. Поскольку Т-трубочки очень тонкие, они по существу улавливают ионы. Это важно, поскольку, независимо от концентрации ионов в других частях клетки, Т-канальцы все еще содержат достаточно ионов кальция, чтобы обеспечить сокращение мышц. Следовательно, даже если концентрация кальция вне клетки падает ( гипокальциемия ), концентрация кальция внутри Т-канальцев остается относительно постоянной, что позволяет сердечному сокращению продолжаться. [6]
Т-трубочки не только являются местом поступления кальция в клетку, но и местом удаления кальция. Это важно, поскольку означает, что уровни кальция внутри клетки можно строго контролировать на небольшой площади (т. е. между Т-трубочками и саркоплазматической сетью, что называется локальным контролем). [17] Белки, такие как натрий-кальциевый обменник и сарколеммальная АТФаза, расположены главным образом в мембране Т-канальцев. [6] Натрий-кальциевый обменник пассивно удаляет один ион кальция из клетки в обмен на три иона натрия. Таким образом, будучи пассивным процессом, он может позволить кальцию проникать в клетку или выходить из нее в зависимости от комбинации относительных концентраций этих ионов и напряжения на клеточной мембране ( электрохимический градиент ). [10] Кальций-АТФаза активно удаляет кальций из клетки, используя энергию, полученную из аденозинтрифосфата (АТФ). [10]
детубуляция
[ редактировать ]Чтобы изучить функцию Т-трубочек, Т-трубочки можно искусственно отсоединить от поверхностной мембраны, используя метод, известный как детубуляция . Химические вещества, такие как глицерин [18] или формамид [14] (для скелетных и сердечных мышц соответственно) можно добавлять во внеклеточный раствор, окружающий клетки. Эти агенты повышают осмолярность внеклеточного раствора, вызывая сокращение клеток. Когда эти агенты прекращаются, клетки быстро расширяются и возвращаются к своему нормальному размеру. Это сжатие и повторное расширение клетки приводит к отделению Т-трубочек от поверхностной мембраны. [19] Альтернативно, осмолярность внеклеточного раствора можно снизить, используя, например, гипотонический солевой раствор, вызывая временное набухание клеток. Возвращение внеклеточного раствора к нормальной осмолярности позволяет клеткам вернуться к своему прежнему размеру, что снова приводит к детубуляции. [20]
История
[ редактировать ]Идея клеточной структуры, которая позже стала известна как Т-трубочка, была впервые предложена в 1881 году. Очень короткий временной интервал между стимуляцией поперечно-полосатой мышечной клетки и ее последующим сокращением был слишком коротким, чтобы быть вызванным сигнальным химическим веществом, перемещающимся на расстояние. между сарколеммой и саркоплазматической сетью. Поэтому было высказано предположение, что мембранные мешочки, проникающие в клетку, могут объяснить наблюдаемое очень быстрое начало сокращения. [21] [22] Лишь в 1897 году были обнаружены первые Т-трубочки с помощью световой микроскопии для изучения сердечной мышцы, которой инъецировали чернила . Технологии визуализации развивались, и с появлением трансмиссионной электронной микроскопии структура Т-трубочек стала более очевидной. [23] что привело к описанию продольного компонента сети Т-трубочек в 1971 году. [24] В 1990-х и 2000-х годах конфокальная микроскопия позволила провести трехмерную реконструкцию сети Т-канальцев и количественно оценить размер и распределение Т-канальцев. [25] и важные взаимоотношения между Т-канальцами и высвобождением кальция начали раскрываться с открытием кальциевых искр . [26] В то время как ранние работы были сосредоточены на сердечной мышце желудочков и скелетных мышцах, в 2009 году наблюдалась обширная сеть Т-трубочек в клетках сердечной мышцы предсердий. [27] Текущие исследования сосредоточены на регуляции структуры Т-канальцев и на том, как Т-канальцы влияют на сердечно-сосудистые заболевания и способствуют их развитию. [28]
Клиническое значение
[ редактировать ]Структура Т-канальцев может быть изменена в результате заболевания, которое в сердце может способствовать слабости сердечной мышцы или нарушению сердечного ритма. Изменения, наблюдаемые при заболевании, варьируются от полной потери Т-трубочек до более тонких изменений в их ориентации или характере ветвления. [29] Т-канальцы могут быть утрачены или разрушены после инфаркта миокарда . [29] а также нарушаются в желудочках пациентов с сердечной недостаточностью , способствуя уменьшению силы сокращения и потенциально снижая шансы на выздоровление. [30] Сердечная недостаточность также может привести к почти полной потере Т-канальцев предсердных кардиомиоцитов, снижая сократимость предсердий и потенциально способствуя фибрилляции предсердий . [27]
Структурные изменения в Т-трубочках могут привести к отходу кальциевых каналов L-типа от рианодиновых рецепторов. Это может увеличить время, необходимое для повышения уровня кальция в клетке, что приведет к ослаблению сокращений и аритмии . [6] [27] Однако нарушение структуры Т-канальцев не может быть постоянным, поскольку некоторые предполагают, что ремоделирование Т-канальцев можно обратить вспять с помощью интервальных тренировок . [6]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж г Хун, ТинТин; Шоу, Робин М. (01 января 2017 г.). «Микроанатомия и функция сердечных Т-трубочек» . Физиологические обзоры . 97 (1): 227–252. doi : 10.1152/physrev.00037.2015 . ISSN 0031-9333 . ПМК 6151489 . ПМИД 27881552 .
- ^ Рог-Зелинска Е.А. и др. (2021). «Поэтапная деформация Т-трубочек кардиомиоцитов приводит к обмену содержимым канальцев» . Цирк. Рез . 128 (2): 203–215. дои : 10.1161/CIRCRESAHA.120.317266 . ПМЦ 7834912 . ПМИД 33228470 .
- ^ Феррантини, Сесилия; Коппини, Рафаэле; Саккони, Леонардо; Тоси, Бенедетта; Чжан, Мэй Ло; Ван, Го Лян; Фрис, Эут де; Хоппенбрауэрс, Эрнст; Павоне, Франческо (1 июня 2014 г.). «Влияние детубуляции на силу и кинетику сокращения сердечной мышцы» . Журнал общей физиологии . 143 (6): 783–797. дои : 10.1085/jgp.201311125 . ПМК 4035744 . ПМИД 24863933 .
- ^ Хэддок, Питер С.; Кутзи, Уильям А.; Чо, Эмили; Портер, Лиза; Като, Хидеки; Берс, Дональд М.; Джафри, М. Салит; Артман, Майкл (3 сентября 1999 г.). «Субклеточные градиенты [Ca2+]i во время взаимодействия возбуждения-сокращения в миоцитах желудочков новорожденных кроликов» . Исследование кровообращения . 85 (5): 415–427. дои : 10.1161/01.RES.85.5.415 . ISSN 0009-7330 . ПМИД 10473671 .
- ^ Ричардс, Массачусетс; Кларк, доктор медицинских наук; Сараванан, П.; Фойгт, Н.; Добрев, Д.; Эйснер, Д.А.; Траффорд, штат Аризона; Дибб, К.М. (ноябрь 2011 г.). «Поперечные канальцы являются общей чертой предсердных миоцитов крупных млекопитающих, включая человека» . Американский журнал физиологии. Физиология сердца и кровообращения . 301 (5): H1996–2005. дои : 10.1152/ajpheart.00284.2011 . ISSN 1522-1539 . ПМК 3213978 . ПМИД 21841013 .
- ^ Jump up to: а б с д и ж г Ибрагим, М.; Горелик Ю.; Якуб, Миннесота; Терраччано, CM (22 сентября 2011 г.). «Структура и функции сердечных Т-канальцев в норме и при заболеваниях» . Труды Королевского общества B: Биологические науки . 278 (1719): 2714–2723. дои : 10.1098/rspb.2011.0624 . ПМК 3145195 . ПМИД 21697171 .
- ^ ди Фьоре, Мариано С.Х.; Ерощенко, Виктор П (2008). Атлас гистологии Ди Фиоре: с функциональными корреляциями . Филадельфия: Wolters Kluwer Health/Lippincott Williams & Wilkins. п. 124. ИСБН 978-0-7817-7057-6 .
- ^ «4. Обратный захват и расслабление кальция» . www.bristol.ac.uk . Проверено 21 февраля 2017 г.
- ^ Колдуэлл, Джессика Л.; Смит, Шарлотта Э.Р.; Тейлор, Ребекка Ф.; Китмитто, Ашраф; Эйснер, Дэвид А.; Дибб, Кэтрин М.; Траффорд, Эндрю В. (5 декабря 2014 г.). «Зависимость поперечных канальцев сердца от белка домена BAR амфифизина II (BIN-1)» . Исследование кровообращения . 115 (12): 986–996. дои : 10.1161/CIRCRESAHA.116.303448 . ISSN 1524-4571 . ПМЦ 4274343 . ПМИД 25332206 .
- ^ Jump up to: а б с М., Берс Д. (2001). Связь возбуждения-сокращения и сократительная сила сердца (2-е изд.). Дордрехт: Kluwer Academic Publishers. ISBN 9780792371588 . OCLC 47659382 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Скривен, доктор медицинских наук; Дэн, П.; Мур, Эд (ноябрь 2000 г.). «Распределение белков, участвующих в соединении возбуждения-сокращения в миоцитах желудочков крысы» . Биофизический журнал . 79 (5): 2682–2691. Бибкод : 2000BpJ....79.2682S . дои : 10.1016/S0006-3495(00)76506-4 . ISSN 0006-3495 . ПМК 1301148 . ПМИД 11053140 .
- ^ Берс, Дональд М. (10 января 2002 г.). «Связь возбуждения и сокращения сердца». Природа . 415 (6868): 198–205. Бибкод : 2002Natur.415..198B . дои : 10.1038/415198a . ISSN 0028-0836 . ПМИД 11805843 . S2CID 4337201 .
- ^ Реббек, Робин Т.; Карунасекара, Ямуна; Совет, Филип Г.; Борода, Николь А.; Казаротто, Марко Г.; Далханти, Анджела Ф. (01 марта 2014 г.). «Соединение возбуждения-сокращения скелетных мышц: кто партнеры по танцу?». Международный журнал биохимии и клеточной биологии . 48 : 28–38. дои : 10.1016/j.biocel.2013.12.001 . ISSN 1878-5875 . ПМИД 24374102 .
- ^ Jump up to: а б с Феррантини, Сесилия; Коппини, Рафаэле; Саккони, Леонардо; Тоси, Бенедетта; Чжан, Мэй Ло; Ван, Го Лян; де Врис, Эут; Хоппенбрауэрс, Эрнст; Павоне, Франческо (1 июня 2014 г.). «Влияние детубуляции на силу и кинетику сокращения сердечной мышцы» . Журнал общей физиологии . 143 (6): 783–797. дои : 10.1085/jgp.201311125 . ISSN 1540-7748 . ПМК 4035744 . ПМИД 24863933 .
- ^ Лафламм, Массачусетс; Беккер, Польша (1 ноября 1999 г.). «G (s) и аденилатциклаза в поперечных канальцах сердца: значение для цАМФ-зависимой передачи сигналов». Американский журнал физиологии . 277 (5, часть 2): H1841–1848. дои : 10.1152/ajpheart.1999.277.5.H1841 . ISSN 0002-9513 . ПМИД 10564138 .
- ^ Берс, Дональд М. (15 мая 2006 г.). «Фосфорилирование сердечных рианодиновых рецепторов: места-мишени и функциональные последствия» . Биохимический журнал . 396 (Часть 1): e1–3. дои : 10.1042/BJ20060377 . ISSN 0264-6021 . ПМК 1450001 . ПМИД 16626281 .
- ^ Хинч Р., Гринштейн Дж. Л., Тансканен А. Дж., Сюй Л. и Уинслоу Р. Л. (2004) «Упрощенная модель локального контроля высвобождения кальция, индуцированного кальцием, в миоцитах сердечного желудочка», 87 (6).
- ^ Фрейзер, Джеймс А.; Скеппер, Джереми Н.; Хокадей, Остин Р.; Хуанг1, Кристофер Л.-Х. (1 августа 1998 г.). «Процесс трубчатой вакуолизации в скелетных мышцах земноводных». Журнал исследований мышц и подвижности клеток . 19 (6): 613–629. дои : 10.1023/А:1005325013355 . ISSN 0142-4319 . ПМИД 9742446 . S2CID 12312117 .
{{cite journal}}: CS1 maint: числовые имена: список авторов ( ссылка ) - ^ Каваи М., Хуссейн М., Орчард CH (1999). «Сочетание возбуждения-сокращения в миоцитах желудочков крысы после детубуляции, вызванной формамидом». Am J Physiol . 277 (2): Н603-9. дои : 10.1152/ajpheart.1999.277.2.H603 . ПМИД 10444485 .
- ^ Мёнч, И.; Микхоф, Кентукки; Ченг, Л.Ф.; Лопатин А.Н. (июль 2013 г.). «Разрешение гипоосмотического стресса в изолированных миоцитах желудочков мыши вызывает запечатывание Т-трубочек» . Экспериментальная физиология . 98 (7): 1164–1177. doi : 10.1113/expphysicalol.2013.072470 . ISSN 1469-445X . ПМЦ 3746342 . ПМИД 23585327 .
- ^ Хаксли, А.Ф. (15 июня 1971 г.). «Активация поперечно-полосатых мышц и ее механическая реакция». Труды Лондонского королевского общества. Серия Б, Биологические науки . 178 (1050): 1–27. дои : 10.1098/rspb.1971.0049 . ISSN 0950-1193 . ПМИД 4397265 . S2CID 30218942 .
- ^ Хилл, А.В. (октябрь 1949 г.). «Резкий переход от покоя к активности мышц». Труды Лондонского королевского общества. Серия Б, Биологические науки . 136 (884): 399–420. Бибкод : 1949РСПСБ.136..399Н . дои : 10.1098/rspb.1949.0033 . ISSN 0950-1193 . ПМИД 18143369 . S2CID 11863605 .
- ^ Линднер, Э. (1957). «[Субмикроскопическая морфология сердечной мышцы]». Журнал клеточных исследований и микроскопической анатомии . 45 (6): 702–746. ISSN 0340-0336 . ПМИД 13456982 .
- ^ Сперелакис, Н.; Рубио, Р. (август 1971 г.). «Упорядоченная решетка осевых канальцев, которые соединяют соседние поперечные канальцы в миокарде желудочков морской свинки». Журнал молекулярной и клеточной кардиологии . 2 (3): 211–220. дои : 10.1016/0022-2828(71)90054-х . ISSN 0022-2828 . ПМИД 5117216 .
- ^ Савио-Галимберти, Элеонора; Фрэнк, Джой; Иноуэ, Масаси; Гольдхабер, Джошуа И.; Каннелл, Марк Б.; Бридж, Джон Х.Б.; Саксе, Фрэнк Б. (август 2008 г.). «Новые особенности поперечной трубчатой системы кролика, выявленные путем количественного анализа трехмерных реконструкций по конфокальным изображениям» . Биофизический журнал . 95 (4): 2053–2062. Бибкод : 2008BpJ....95.2053S . дои : 10.1529/biophysj.108.130617 . ISSN 1542-0086 . ПМЦ 2483780 . ПМИД 18487298 .
- ^ Ченг, Х.; Ледерер, В.Дж.; Каннелл, МБ (29 октября 1993 г.). «Кальциевые искры: элементарные события, лежащие в основе связи возбуждения-сокращения в сердечной мышце». Наука . 262 (5134): 740–744. Бибкод : 1993Sci...262..740C . дои : 10.1126/science.8235594 . ISSN 0036-8075 . ПМИД 8235594 .
- ^ Jump up to: а б с Дибб, Кэтрин М.; Кларк, Джессика Д.; Хорн, Марго А.; Ричардс, Марк А.; Грэм, Хелен К.; Эйснер, Дэвид А.; Траффорд, Эндрю В. (сентябрь 2009 г.). «Характеристика обширной поперечной канальцевой сети в предсердных миоцитах овцы и ее истощение при сердечной недостаточности» . Кровообращение: Сердечная недостаточность . 2 (5): 482–489. doi : 10.1161/CIRCHEARTFAILURE.109.852228 . ISSN 1941-3297 . ПМИД 19808379 .
- ^ Эйснер, Дэвид А.; Колдуэлл, Джессика Л.; Кистамас, Корнель; Траффорд, Эндрю В. (07 июля 2017 г.). «Кальций и связь возбуждения-сокращения в сердце» . Исследование кровообращения . 121 (2): 181–195. дои : 10.1161/CIRCRESAHA.117.310230 . ISSN 1524-4571 . ПМК 5497788 . ПМИД 28684623 .
- ^ Jump up to: а б Пинали, Кристиан; Малик, Надим; Давенпорт, Дж. Бернард; Аллан, Лоуренс Дж.; Мерфитт, Люси; Икбал, Мохаммед М.; Бойетт, Марк Р.; Райт, Элизабет Дж.; Уокер, Рэйчел (4 мая 2017 г.). «Т-трубочки после инфаркта миокарда образуют увеличенные разветвленные структуры с нарушением регуляции юнктофилина-2 и мостового интегратора 1 (BIN-1)» . Журнал Американской кардиологической ассоциации . 6 (5). дои : 10.1161/JAHA.116.004834 . ISSN 2047-9980 . ПМК 5524063 . ПМИД 28473402 .
- ^ Зейдель, Томас; Наванкасаттусас, Сутип; Ахмад, Азми; Диакос, Николаос А.; Сюй, Вейнинг Дэвид; Тристани-Фирузи, Мартин; Бониос, Майкл Дж.; Талеб, Иосиф; Ли, Дин Ю. (25 апреля 2017 г.). «Листчатое ремоделирование поперечной трубчатой системы при сердечной недостаточности человека нарушает связь возбуждения-сокращения и функциональное восстановление путем механической разгрузки» . Тираж . 135 (17): 1632–1645. doi : 10.1161/CIRCULATIONAHA.116.024470 . ISSN 1524-4539 . ПМК 5404964 . ПМИД 28073805 .