Диоксигеназа
| Диоксигеназа | |||
|---|---|---|---|
 Кристаллическая структура штамма Acinetobacter ADP1 протокатехуата 3,4-диоксигеназы в комплексе с 3,4-дигидроксибензоатом | |||
| Идентификаторы | |||
| Символ | Диоксигеназа_C | ||
| Пфам | PF00775 | ||
| Пфам Клан | CL0287 | ||
| ИнтерПро | IPR000627 | ||
| PROSITE | PDOC00079 | ||
| ОБЛАСТЬ ПРИМЕНЕНИЯ 2 | 2шт / ОБЛАСТЬ / СУПФАМ | ||
| |||
Диоксигеназы являются оксидоредуктазами ферментами . Аэробная жизнь , от простых одноклеточных видов бактерий до сложных эукариотических организмов, эволюционировала и стала зависеть от окислительной способности дикислорода в различных метаболических путях. От генерации энергетического аденозинтрифосфата (АТФ) до разложения ксенобиотиков , использование дикислорода в качестве биологического окислителя широко распространено и варьируется по точному механизму его использования. Ферменты используют множество различных схем использования дикислорода, и это во многом зависит от субстрата и реакции.
Сравнение с монооксигеназами
[ редактировать ]В монооксигеназах только один атом дикислорода включается в субстрат, а другой восстанавливается до молекулы воды. Диоксигеназы ( КФ 1.13.11 ) катализируют окисление субстрата без восстановления одного атома кислорода из дикислорода в молекулу воды. Однако это определение неоднозначно, поскольку не учитывает, сколько субстратов участвует в реакции. различные схемы кофакторов Большинство диоксигеназ полностью включают дикислород в один субстрат, и для достижения этого используются . Например, в α-кетоглутарат- зависимых ферментах один атом дикислорода включен в два субстрата, один из которых всегда представляет собой α-кетоглутарат, и эта реакция осуществляется моноядерным железным центром.
Железосодержащие ферменты
[ редактировать ]Наиболее широко наблюдаемым кофактором, участвующим в реакциях диоксигенирования, является железо , но каталитическая схема, используемая этими железосодержащими ферментами, весьма разнообразна. Железосодержащие диоксигеназы можно разделить на три класса в зависимости от того, как железо включается в активный центр: те, которые используют моноядерный железный центр, те, которые содержат кластер Риске [2Fe-2S] и те, которые используют простетическую группу гема .
Мононуклеарные диоксигеназы железа
[ редактировать ]Мононуклеарные железодиоксигеназы или негемовые железозависимые диоксигеназы, как их еще называют, все используют одно каталитическое железо для включения одного или обоих атомов дикислорода в субстрат. Несмотря на это обычное событие оксигенации, мононуклеарные диоксигеназы железа различаются по тому, как активация дикислорода используется для стимулирования определенных химических реакций. [ 1 ] Например, расщепление связи углерод-углерод, гидроперекисное окисление жирных кислот, разрыв связи углерод-сера и окисление тиола — все это реакции, катализируемые моноядерными диоксигеназами железа. [ 1 ] [ 2 ] [ 3 ]
Большинство мононуклеарных диоксигеназ железа являются членами суперсемейства купинов , в котором общая структура домена описывается как шестицепочечная β-бочкообразная складка (или мотив желеобразного рулета ). В центре этой бочкообразной структуры находится ион металла, чаще всего двухвалентного железа, координационное окружение которого часто обеспечивается остатками двух частично консервативных структурных мотивов: G(X) E ( X ) 5 HXH(X) 3–4 6 G и Г(Х) 5 - 7 РХГ(Х) 2 Н(Х) 3 Н. [ 4 ] [ 5 ]


Двумя важными группами мононуклеарных негемовых диоксигеназ железа являются катехолдиоксигеназы и 2-оксоглутарат (2OG)-зависимые диоксигеназы . [ 6 ] Катехолдиоксигеназы кольцевой , некоторые из наиболее хорошо изученных ферментов диоксигеназ, используют дикислород для расщепления углерод-углеродной связи в системе ароматических катехолов . [ 4 ] Катехолдиоксигеназы далее классифицируются как «экстрадиол» или «интрадиол», и это различие основано на механистических различиях в реакциях (рис. 1 и 2). Ферменты интрадиола расщепляют углерод-углеродную связь между двумя гидроксильными группами. Активный центр железа координируется четырьмя белковыми лигандами — двумя гистидиновыми и двумя тирозинатными остатками — по тригонально-бипирамидальному принципу, причем пятое координационное место занимает молекула воды. [ 3 ] Как только катехолатный субстрат связывается с металлическим центром бидентатно через депротонированные гидроксильные группы, трехвалентное железо «активирует» субстрат посредством отрыва электрона с образованием радикала на субстрате. Затем это позволяет осуществить реакцию с дикислородом и последующее расщепление итрадиола через промежуточный циклический ангидрид. [ 2 ] [ 4 ] Члены экстрадиола используют двухвалентное железо в качестве активного окислительно-восстановительного состояния, и этот центр обычно координируется октаэдрически через мотив 2-His-1-Glu с лабильными водными лигандами, занимающими пустые позиции. Как только субстрат связывается с железистым центром, это способствует связыванию дикислорода и последующей активации. [ 2 ] [ 4 ] [ 7 ] Затем эта активированная форма кислорода вступает в реакцию с субстратом, в конечном итоге расщепляя связь углерод-углерод, прилегающую к гидроксильным группам, посредством образования промежуточного соединения α-кетолактона. [ 3 ]
В 2OG-зависимых диоксигеназах двухвалентное железо ( Fe(II) ) также координируется мотивом (His)2(Glu/Asp)1 «лицевой триады». Бидентатная координация 2OG и воды завершает псевдооктаэдрическую координационную сферу. После связывания субстрата водный лиганд высвобождается, образуя открытый координационный сайт для активации кислорода. [ 6 ] При связывании кислорода происходит плохо изученное превращение, во время которого 2OG окислительно декарбоксилируется до сукцината, а связь OO расщепляется с образованием промежуточного соединения Fe(IV)-оксо( феррил ). Этот мощный окислитель затем используется для проведения различных реакций, включая гидроксилирование, галогенирование и деметилирование. [ 8 ] В наиболее изученном случае гидроксилазы феррильное промежуточное соединение отрывает атом водорода от целевого положения субстрата, образуя радикал субстрата и Fe (III)-OH. Затем этот радикал соединяется с гидроксидным лигандом, образуя гидроксилированный продукт и состояние покоя фермента Fe (II). [ 8 ]
Риске диоксигеназы
[ редактировать ]Диоксигеназы Риске обычно катализируют цис-дигидроксилирование аренов до продуктов цис-дигидродиолов. Эти диоксигеназы также катализируют сульфоксидирование, десатурацию и окисление бензила. [ 2 ] Эти ферменты широко обнаружены в почвенных бактериях, таких как Pseudomonas , [ 3 ] и их реакции представляют собой начальный этап биоразложения ароматических углеводородов. [ 2 ] Диоксигеназы Риске структурно более сложны, чем другие диоксигеназы, из-за необходимости эффективного пути переноса электронов (рисунок 2), чтобы опосредовать дополнительное одновременное двухэлектронное восстановление ароматического субстрата.

Диоксигеназы Риске состоят из трех компонентов: НАДН-зависимой редуктазы ФАД , ферредоксина с двумя кластерами Риске [2Fe-2S] и оксигеназы α3β3, каждая α-субъединица которой содержит моноядерный центр железа и кластер [2Fe-2S] Риске. [ 2 ] Внутри каждой α-субъединицы железо-серный кластер и моноядерный железный центр разделены расстоянием ~ 43 Å, что слишком далеко для эффективного переноса электронов . Вместо этого предполагается, что перенос электронов осуществляется через эти два центра в соседних субъединицах, при этом железо-серный кластер одной субъединицы переносит электроны в моноядерный центр железа соседней субъединицы, который удобно разделен на ~ 12 Å. Хотя это расстояние кажется оптимальным для эффективного переноса электронов, замена мостикового остатка аспартата приводит к потере функции фермента, что позволяет предположить, что перенос электронов вместо этого происходит через сеть водородных связей, удерживаемых этим остатком аспартата. [ 3 ]
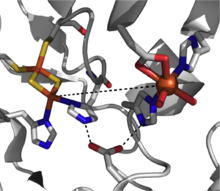
механизм активации О 2 этим классом диоксигеназ. Описан [ 7 ] Этот вид может представлять собой активный окислитель или может подвергаться гемолитическому разрыву связи OO с образованием промежуточного соединения железа (V)-оксо в качестве рабочего окислителя. [ 3 ] [ 7 ]
Гемсодержащие диоксигеназы
[ редактировать ]В то время как большинство железозависимых диоксигеназ используют негемовый кофактор железа, окисление L-(и D-)триптофана до N-формилкинуренина катализируется либо триптофан-2,3-диоксигеназой (ТДО), либо индоламин-2,3-диоксигеназой . IDO), которые представляют собой гемодиоксигеназы, которые используют железо, координированное простетической группой гема B. [ 9 ] [ 10 ] Хотя эти диоксигеназы представляют интерес отчасти потому, что они уникальным образом используют гем для катализа, они также представляют интерес из-за их важности в регуляции триптофана в клетке, что имеет многочисленные физиологические последствия. [ 11 ] Считается, что первоначальная ассоциация субстрата с двухкислородным железом в активном центре фермента происходит либо посредством радикального, либо электрофильного присоединения, требуя либо двухвалентного железа, либо трехвалентного железа соответственно. [ 9 ] Хотя точный механизм реакции гем-зависимых диоксигеназ все еще обсуждается, предполагается, что реакция протекает либо по диоксетановому механизму, либо по механизму Криги (рис. 4, 5). [ 9 ] [ 11 ]
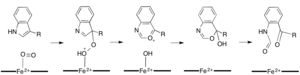
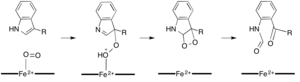
Камбиалистические диоксигеназы
[ редактировать ]Хотя железо на сегодняшний день является наиболее распространенным кофактором, используемым для ферментативного диоксигенирования, оно не требуется всем диоксигеназам для катализа. Кверцетин-2,3-диоксигеназа (кверцетиназа, QueD) катализирует диоксигенолитическое расщепление кверцетина до 2-протокатехоилфлороглюцинкарбоновой кислоты и монооксида углерода . [ 12 ] Наиболее охарактеризованный фермент Aspergillus japonicus требует присутствия меди . [ 4 ] и были обнаружены бактериальные кверцетиназы, которые являются весьма беспорядочными (камбиалистическими) [ 13 ] В их требованиях металлоцентр с различной степенью активности сообщается с замещением двухвалентного марганца , кобальта , железа, никеля и меди. [ 12 ] (Кверцетин, роль в обмене веществ). Ациредуктон (1,2-дигидрокси-5-(метилтио)пент-1-ен-3-он)диоксигеназа (ARD) обнаружена как у прокариот , так и у эукариот . [ 4 ] [ 12 ] [ 14 ] Ферменты ARD большинства видов связывают двухвалентное железо и катализируют окисление ациредуктона до 4-(метилтио)-2-оксобутаноата, α-кетокислоты метионина и муравьиной кислоты . Однако ARD из Klebsiella oxytoca катализирует дополнительную реакцию при связывании никеля (II): вместо этого он образует 3-(метилтио) пропионат, формиат и окись углерода в результате реакции ациредуктона с дикислородом. Активность Fe-ARD тесно переплетена с путем утилизации метионина, в котором метилтиоаденозиновый продукт клеточных реакций S-аденозилметионина (SAM) в конечном итоге превращается в ациредуктон.
Хотя точная роль Ni-ARD неизвестна, предполагается, что он помогает регулировать уровень метионина, действуя как шунт на пути спасения. Этот фермент K. oxytoca представляет собой уникальный пример того, как присутствующий ион металла определяет, какая реакция катализируется. Кверцетиназы и ферменты ARD являются членами суперсемейства купинов , к которому также принадлежат ферменты мононуклеарного железа. [ 15 ] Схема координации металлов для ферментов QueD представляет собой либо 3-His, либо 3-His-1-Glu, причем точное расположение зависит от организма. [ 4 ] Все ферменты ARD хелатируют каталитический металл (Ni или Fe) посредством мотива 3-His-1-Glu. [ 15 ] В этих диоксигеназах координирующие лиганды представлены обоими типичными купиновыми мотивами. В ферментах ARD металл существует в октаэдрическом расположении с тремя гистидиновыми остатками, образующими лицевую триаду. [ 14 ] Металлоцентры бактериальной кверцетиназы обычно имеют тригонально-бипирамидальную или октаэдрическую координационную среду, когда имеется четыре белковых лиганда; Металлические центры медь-зависимых ферментов QueD обладают искаженной тетраэдрической геометрией, в которой только три консервативных остатка гистидина обеспечивают координационные лиганды. [ 4 ] [ 12 ] Пустые координационные места во всех металлоцентрах заняты аквалигандами до тех пор, пока они не будут вытеснены поступающим субстратом.
Способность этих диоксигеназ сохранять активность в присутствии других металлов-кофакторов с широким диапазоном окислительно-восстановительных потенциалов позволяет предположить, что металлцентр не играет активной роли в активации дикислорода. Скорее, считается, что функция металлического центра заключается в удержании субстрата в правильной геометрии для его реакции с дикислородом. В этом отношении эти ферменты напоминают итрадиолкатехиндиоксигеназы , при которых металлоцентры активируют субстрат для последующей реакции с дикислородом.
Кофактор-независимые диоксигеназы
[ редактировать ]
Диоксигеназы, которые катализируют реакции без необходимости в кофакторе, встречаются в природе гораздо реже, чем те, которые в них нуждаются. две диоксигеназы, 1H-3-гидрокси-4-оксохинолин-2,4-диоксигеназа (QDO) и 1H-3-гидрокси-4-оксохинальдин-2,4-диоксигеназа Было показано, что (HDO), не требуют ни органических, ни металлический кофактор. [ 16 ] Эти ферменты катализируют деградацию хинолоновых гетероциклов аналогично кверцетиндиоксигеназе , но, как полагают, опосредуют радикальную реакцию молекулы дикислорода с карбанионом на субстрате (рисунок 5). [ 17 ] И HDO, и QDO принадлежат к суперсемейству ферментов α/β-гидролаз , хотя каталитические остатки в HDO и QDO, по-видимому, не выполняют ту же функцию, что и в остальных ферментах суперсемейства α/β-гидролаз. [ 16 ]
Клиническое значение
[ редактировать ]Разнообразие в семействе диоксигеназ означает широкий спектр биологических ролей:
- Триптофан-2,3-диоксигеназа (ТДО) помогает регулировать уровень триптофана в организме и экспрессируется во многих опухолях человека. [ 18 ] Другая гемовая железозависимая диоксигеназа, IDO, также имеет отношение к здоровью человека, поскольку она участвует в воспалительных реакциях в контексте некоторых заболеваний. [ 19 ] IDO влияет как на триптофан, так и на кинуренин и связан с депрессией у людей. [ 20 ]
- Алкаптонурия — генетическое заболевание, приводящее к дефициту гомогентизат-1,2-диоксигеназы , которая катализирует образование 4-малеилацетоацетата из гомогентизата . [ 21 ] Накопление гомогентизиновой кислоты может привести к повреждению сердечного клапана, образованию камней в почках и повреждению хрящей в организме. [ 22 ]
- Нейродегенерация, связанная с пантотенаткиназой (PKAN), представляет собой аутосомно-рецессивное заболевание, которое может приводить к развитию гранул железа и телец Леви в нейронах . Исследование показало, что у пациентов с диагнозом ПКАН обнаружено повышение уровня цистеина в бледном шаре вследствие дефицита цистеиндиоксигеназы . [ 23 ] У пациентов с ПКАН часто развиваются симптомы деменции , и они часто умирают в раннем взрослом возрасте.
- При репарации ДНК Fe (II)/2-оксоглутарат-зависимая диоксигеназа AlkB участвует в окислительном удалении алкилированных повреждений ДНК. Неспособность устранить повреждение алкилирования ДНК может привести к цитотоксичности или мутагенезу во время репликации ДНК.
- Циклооксигеназы (ЦОГ), которые отвечают за образование простаноидов в организме человека, являются мишенью многих НПВП . обезболивающих [ 10 ] Ингибирование ЦОГ приводит к уменьшению воспаления и оказывает анальгетический эффект за счет снижения уровня синтеза простагландинов и тромбоксана.
Ссылки
[ редактировать ]- ^ Jump up to: а б Лейтгеб С., Нидецкий Б. (декабрь 2008 г.). «Структурное и функциональное сравнение 2-His-1-карбоксилата и 3-His металлоцентров в негемовых железо(II)-зависимых ферментах». Труды Биохимического общества . 36 (Часть 6): 1180–6. дои : 10.1042/BST0361180 . ПМИД 19021520 .
- ^ Jump up to: а б с д и ж Рунда, Майкл Э.; Де Кок, Нильс А.В.; Шмидт, Сэнди (2023). «Оксигеназы Риске и другие ферредоксин-зависимые ферменты: принципы переноса электрона и каталитические возможности». ХимБиоХим . 24 (15): e202300078. дои : 10.1002/cbic.202300078 . ПМИД 36964978 .
- ^ Jump up to: а б с д и ж Самуэль де Виссер; Девеш Кумар (2011). Железосодержащие ферменты — универсальные катализаторы реакций гидроксилирования в природе . Королевское химическое общество. ISBN 978-1-84973-298-7 .
- ^ Jump up to: а б с д и ж г час Фетцнер С. (апрель 2012 г.). «Диоксигеназы, расщепляющие кольцо, с купиновой складкой» . Прикладная и экологическая микробиология . 78 (8): 2505–14. Бибкод : 2012ApEnM..78.2505F . дои : 10.1128/АЕМ.07651-11 . ПМК 3318818 . ПМИД 22287012 .
- ^ Стипанук М.Х., Симмонс Ч.Р., Карплюс П.А., Домини Дж.Е. (июнь 2011 г.). «Тиолдиоксигеназы: уникальные семейства купиновых белков» . Аминокислоты . 41 (1): 91–102. дои : 10.1007/s00726-010-0518-2 . ПМК 3136866 . ПМИД 20195658 .
- ^ Jump up to: а б Соломон Э.И., Брунольд Т.С., Дэвис М.И., Кемсли Дж.Н., Ли С.К., Ленерт Н. и др. (январь 2000 г.). «Геометрические и электронные корреляции структуры/функции в ферментах негемового железа». Химические обзоры . 100 (1): 235–350. дои : 10.1021/cr9900275 . ПМИД 11749238 .
- ^ Jump up to: а б с Барри, Сара М.; Чаллис, Грегори Л. (2013). «Механизм и каталитическое разнообразие негемовых железозависимых оксигеназ Риске» . АКС-катализ . 3 (10): 2362–2370. дои : 10.1021/cs400087p . ПМЦ 3827946 . ПМИД 24244885 .
- ^ Jump up to: а б Кребс С., Галонич Фухимори Д., Уолш К.Т., Боллинджер Дж.М. (июль 2007 г.). «Негемовые Fe(IV)-оксо-промежуточные соединения» . Отчеты о химических исследованиях . 40 (7): 484–92. дои : 10.1021/ar700066p . ПМК 3870002 . ПМИД 17542550 .
- ^ Jump up to: а б с Ефимов И., Басран Дж., Текрей С.Дж., Ханда С., Моват К.Г., Рэйвен Э.Л. (апрель 2011 г.). «Строение и механизм реакции гемодиоксигеназ» . Биохимия . 50 (14): 2717–24. дои : 10.1021/bi101732n . ПМК 3092302 . ПМИД 21361337 .
- ^ Jump up to: а б Соно М., член парламента от Роуча, Коултер Э.Д., Доусон Дж.Х. (ноябрь 1996 г.). «Гемсодержащие оксигеназы». Химические обзоры . 96 (7): 2841–2888. дои : 10.1021/cr9500500 . ПМИД 11848843 .
- ^ Jump up to: а б Текрей С.Дж., Моват К.Г., Чепмен С.К. (декабрь 2008 г.). «Изучение механизма действия триптофан-2,3-диоксигеназы» . Труды Биохимического общества . 36 (Часть 6): 1120–3. дои : 10.1042/BST0361120 . ПМЦ 2652831 . ПМИД 19021508 .
- ^ Jump up to: а б с д Шааб М.Р., Барни Б.М., Франциско В.А. (январь 2006 г.). «Кинетические и спектроскопические исследования кверцетин-2,3-диоксигеназы Bacillus subtilis». Биохимия . 45 (3): 1009–16. дои : 10.1021/bi051571c . ПМИД 16411777 .
- ^ «Одиночная супероксиддисмутаза Rhodobacter capsulatus представляет собой камбиалистический марганецсодержащий фермент» . Jb.asm.org . Проверено 11 марта 2014 г.
- ^ Jump up to: а б Марони М.Дж., Чюрли С. (апрель 2014 г.). «Неокислительно-восстановительные никелевые ферменты» . Химические обзоры . 114 (8): 4206–28. дои : 10.1021/cr4004488 . ПМЦ 5675112 . ПМИД 24369791 .
- ^ Jump up to: а б Бур Дж.Л., Малруни С.Б., Хаузингер Р.П. (февраль 2014 г.). «Никельзависимые металлоферменты» . Архив биохимии и биофизики . 544 : 142–52. дои : 10.1016/j.abb.2013.09.002 . ПМЦ 3946514 . ПМИД 24036122 .
- ^ Jump up to: а б Фетцнер С. (ноябрь 2002 г.). «Оксигеназы без необходимости в кофакторах или ионах металлов». Прикладная микробиология и биотехнология . 60 (3): 243–57. дои : 10.1007/s00253-002-1123-4 . ПМИД 12436305 . S2CID 24908037 .
- ^ Багг ТД (сентябрь 2003 г.). «Ферменты диоксигеназы: каталитические механизмы и химические модели». Тетраэдр . 59 (36): 7075–7101. дои : 10.1016/S0040-4020(03)00944-X .
- ^ Пилотт Л., Ларрье П., Строобант В., Колау Д., Долусик Е., Фредерик Р., Де План Е., Юттенхове С., Воутерс Дж., Мазерил Б., Ван ден Эйнде Б.Дж. (февраль 2012 г.). «Обращение опухолевой иммунной резистентности путем ингибирования триптофан-2,3-диоксигеназы» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (7): 2497–502. Бибкод : 2012PNAS..109.2497P . дои : 10.1073/pnas.1113873109 . ПМК 3289319 . ПМИД 22308364 .
- ^ Мураками Ю, Хоши М, Имамура Ю, Ариока Ю, Ямамото Ю, Сайто К (2013). «Замечательная роль индоламин-2,3-диоксигеназы и метаболитов триптофана при инфекционных заболеваниях: потенциальная роль в воспалительных заболеваниях, опосредованных макрофагами» . Медиаторы воспаления . 2013 : 391984. doi : 10.1155/2013/391984 . ПМЦ 3588179 . ПМИД 23476103 .
- ^ Субаренда ME, Postolache TT (сентябрь 2012 г.). «Нейровоспаление и депрессия: роль индоламин-2,3-диоксигеназы (IDO) как молекулярного пути». Психосоматическая медицина . 74 (7): 668–72. дои : 10.1097/PSY.0b013e318268de9f . ПМИД 22923699 .
- ^ Фут D, Фут JG (2011). Биохимия (4-е изд.). Хобокен, Нью-Джерси: John Wiley & Sons. п. 1045. ИСБН 978-0470917459 .
- ^ Форнпхуткул С., Интрон В.Дж., Перри М.Б., Бернардини И., Мерфи М.Д., Фитцпатрик Д.Л., Андерсон П.Д., Хейзинг М., Аникстер Ю., Гербер Л.Х., Гал В.А. (декабрь 2002 г.). «Естественная история алкаптонурии» . Медицинский журнал Новой Англии . 347 (26): 2111–21. doi : 10.1056/NEJMoa021736 . ПМИД 12501223 .
- ^ Стипанюк, М.Х.; Домини-младший, JE; Ли, Джи; Колозо, РМ (июнь 2006 г.). «Метаболизм цистеина млекопитающих: новый взгляд на регуляцию метаболизма цистеина» . Журнал питания . 136 (дополнение 6): 1652S–1659S. дои : 10.1093/jn/136.6.1652S . ПМИД 16702335 . Проверено 5 апреля 2021 г.