Зеленая серная бактерии
| Зеленая серная бактерии | |
|---|---|

| |
| Бактерии зеленого серы в колонке Winogradsky | |
| Научная классификация | |
| Домен: | Бактерии |
| (не вмешательство): | Бактероидота -хлорбиота группа |
| Филум: | Хлоробиот Iino et al. 2021 [ 3 ] |
| Сорт: | "Хлоробия" Garrity and Holt 2001 [ 2 ] |
| Заказ: | Хлорбиал Gibbons and Murray 1978 (утвержденные списки 1980) [ 1 ] |
| Семьи и роды | |
| Синонимы | |
| |
Зеленая серная бактерии представляют собой тип , хлорбиота , [ 4 ] обязательно анаэробных фотоавтотрофных бактерий , которые метаболизируют серную. [ 5 ]
Зеленая серная бактерии не мотильны (за исключением хлорпетонического талассии , который может скользить) и способен к аноксигенным фотосинтезу . [ 5 ] [ 6 ] Они живут в анаэробной водной среде. [ 7 ] В отличие от растений, бактерии зеленого серы в основном используют сульфидные ионы в качестве доноров электронов. [ 8 ] Это автотрофы , которые используют обратный цикл трикарбоновой кислоты для выполнения углеродной фиксации . [ 9 ] Они также являются миксотрофами и снижают азот. [ 10 ] [ 11 ]
Характеристики
[ редактировать ]Зеленая серная бактерии представляют собой грамотрицательные стержни или сферические бактерии. Некоторые виды бактерий зеленого серы имеют газовые вакуоли, которые позволяют двигаться. Это фотолитоавтотрофы, и используют световую энергию и уменьшенные соединения серы в качестве источника электрона. [ 12 ] Доноры электронов включают H 2 , H 2 S, S. Основным фотосинтетическим пигментом в этих бактериях является бактериохлорофиллы C или D в зеленых видах и E у коричневых видов и расположен в хлоросомах и плазматических мембранах. [ 7 ] Хлорсомы -это уникальная особенность, которая позволяет им собирать свет в условиях низкого освещения. [ 13 ]
Среда обитания
[ редактировать ]Большинство бактерий зеленого серы являются мезофильными , предпочитают умеренные температуры и все живут в водной среде. Они требуют анаэробных условий и уменьшения серы; Они обычно встречаются в верхних миллиметрах осадка. Они способны к фотосинтезу в условиях низкого освещения. [ 7 ]
Было обнаружено, что в Черном море , чрезвычайно нексичной среде, находится большая популяция бактерий зеленого серы на глубине около 100 м. Из -за отсутствия света, доступного в этом регионе моря, большинство бактерий были фотосинтетически неактивными. Фотосинтетическая активность, обнаруженная в хемоклинане сульфида , позволяет предположить, что бактериям нуждается в очень небольшой энергии для поддержания клеток. [ 14 ]
Был обнаружен вид зеленого серы -бактерий, живущего возле черного курильщика у побережья Мексики на глубине 2500 м в Тихом океане . На этой глубине бактерия, обозначенная GSB1, живет от тусклого свечения теплового вентилятора, поскольку ни один солнечный свет не может проникнуть в эту глубину. [ 15 ]
Зеленая серная бактерии также были найдены, живущие на колониях коралловых рифов на Тайване, они составляют большую часть «зеленого слоя» на этих колониях. Они, вероятно, играют роль в коралловой системе, и между бактериями и коралловым хозяином может быть симбиотическая связь. [ 16 ] Коралл может обеспечить анаэробную среду и источник углерода для бактерий. Бактерии могут обеспечивать питательные вещества и детоксифицировать коралл, окисляя сульфид. [ 17 ]
Один тип бактерий зеленого серы, хлорбакул Tepidum , был обнаружен в серы. Эти организмы термофильные , в отличие от большинства других зеленых бактерий серы. [ 7 ]
Филогения
[ редактировать ]| 16S RRNA на основе LTP _08_2023 [ 18 ] [ 19 ] [ 20 ] | 120 маркерных белков на основе GTDB 08-RS214 [ 21 ] [ 22 ] [ 23 ] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
|
Таксономия
[ редактировать ]- Семейство Chlorobiaceae Copeland 1956 ["Chlorobacteriaceae" Geitler & Pascher 1925 ]
- ? Ancalochloris Gorlenko and Lebedeva 1971
- Chlorobaculum Imhoff 2003
- Chlorobium Nadson 1906
- ? "" Chroloplana " Dubinina и Gorlenko 1975
- ? " Clathrochloris " Geitler 1925
- Протекохлорис Горленко 1970
- Семейство " Thermochlorobacteriaceae " Corrig. Лю и соавт. 2012 ["Chloroherpetonaceae" Bello et al. 2022 ]
- Chloroherpeton Gibson et al. 1985
- « Ca. Thermochlorobacter » Liu et al. 2012
Конкретные характеристики родов
[ редактировать ]Зеленая серная бактерии - это семейство Chlorobiaceae. Есть четыре рода; Хлорпетон , протекохлорис , хлорбиум и хлорбакулум . Характеристики, используемые для различения этих родов, включают некоторые метаболические свойства, пигменты, морфологию клеток и спектры поглощения. Тем не менее, трудно различить эти свойства, и поэтому таксономическое разделение иногда неясно. [ 24 ]
Как правило, хлорбиум - это стержня или вибердайка, а некоторые виды содержат газовые пузырьки. Они могут развиваться как отдельные или агрегатные ячейки. Они могут быть зелеными или темно -коричневыми. Зеленые штаммы используют фотосинтетические пигменты BCHL C или D с каротиноидами хлорбактена, а коричневые штаммы используют фотосинтетический пигмент BCHL E с каротиноидами изорениератена. Низкое количество соли требуется для роста. [ 24 ]
Просеткохлористы состоит из виброид, яйцеклеток или стержня. Они начинаются как отдельные клетки, которые образуют придатки, которые не отражаются, называемые неразборчивыми протезами. Они также могут сформировать газовые пузырьки. Присутствующие фотосинтетические пигменты включают BCHL C, D или E. Кроме того, соль необходима для роста. [ 24 ]
Chlorobaculum развивается как отдельные клетки и, как правило, является вибердайкой или стержней. Некоторые из них могут формировать газовые везикулы. Фотосинтетические пигменты в этом роде: BCHL , D или E. C Некоторые виды требуют NaCl (хлорид натрия) для роста. Члены этого рода были частью рода хлорбиума, но сформировали отдельную линию. [ 24 ]
Род хлорохерпетон уникален, потому что члены этого рода являются подвижными. Они сгибают длинные стержни и могут двигаться, скользя. Они зеленые цвета и содержат фотосинтетический пигмент BCHL C, а также γ-каротин . Соль требуется для роста. [ 24 ]
Метаболизм
[ редактировать ]Фотосинтез
[ редактировать ]Бактерии зеленого серы используют реакционный центр типа I для фотосинтеза. Центры реакции типа I являются бактериальным гомологом фотосистемы I (PSI) в растениях и цианобактериях . Центры реакции GSB содержат бактериохлорофилл A и известны как реакционные центры p840 из -за длины волны возбуждения 840 нм, который приводит в движение поток электронов. В бактериях зеленого серы реакционный центр связан с большим антеннским комплексом, называемым хлорсомой, которая захватывает и пробуждает световую энергию в реакционный центр. Хлорсомы имеют пиковое поглощение в дальней красной области спектра между 720 и 750 нм, потому что они содержат бактериохлорофилл C, D и E. [ 25 ] Белковой комплекс, называемый комплексом Фенна-Матфею-Олсона (FMO), физически расположен между хлорсомами и RC-RC P840. Комплекс FMO помогает эффективно перенести энергию, поглощаемую антенями в реакционный центр.
PSI и Центры реакции I типа способны уменьшить ферредоксин (FD), сильный редуктатор, который можно использовать для исправления CO
2 и уменьшить NAD + Полем Как только реакционный центр (RC) дал электрон FD, он становится окислительным агентом (p840 + ) с потенциалом снижения около +300 мВ. Хотя это недостаточно положительно, чтобы лишить электроны из воды, чтобы синтезировать o
2 ( и
0 = +820 мВ), он может принимать электроны из других источников, таких как h
2 с , тиосульфат или Fe 2+
ионы [ 26 ] Этот транспорт электронов от доноров, таких как H
2 с до акцептора FD называется линейным электронным потоком или линейным переносом электронов. Окисление сульфид -ионов приводит к производству серы как отходов, который накапливается как глобулы на внеклеточной стороне мембраны. Эти глобулы серы дают зеленым бактериям серы своего названия. Когда сульфид истощается, глобулы серы потребляются и дополнительно окисляются до сульфата. Тем не менее, путь окисления серы плохо понят. [ 8 ]
Вместо того, чтобы передавать электроны на FD, кластеры Fe-S в реакционном центре P840 могут переносить электроны в менахинон (MQ: MQH
2 ), который возвращает электроны в P840 + через электронную транспортную цепь (и т. Д.). На обратном пути к RC электроны от MQH2 проходят через комплекс цитохрома BC 1 (аналогичный комплексу III митохондрий), который накачивает H +
ионы через мембрану. Электрохимический потенциал протонов через мембрану используется для синтеза АТФ с помощью F O F 1 ATP -синтазы . Этот циклический транспорт электронов отвечает за преобразование энергии света в ячейку энергии в форме АТФ. [ 25 ]
Серная метаболизм
[ редактировать ]Зеленая серная бактерии окисляет неорганические соединения серы для использования в качестве доноров электронов для анаэробного фотосинтеза, в частности, при фиксации углекислого газа. Они обычно предпочитают использовать сульфид по другим соединениям серы в качестве донора электронов, однако они могут использовать тиосульфат или H 2 . [ 27 ] Промежуточное соединение обычно - это сера, которая откладывается за пределами клетки, [ 28 ] и конечный продукт сульфат. Сера, которая откладывается внеклеточно, находится в форме серы, которые могут быть позже полностью окислены. [ 27 ]
Механизмы окисления серы в бактериях зеленого серы плохо характеризуются. Некоторые ферменты, которые, как считается, участвуют в окислении сульфида, включают флавоцитохром C, сульфид: хинон -оксидоредуктаза и SO
X Система. Флавоцитохром может катализировать перенос электронов в цитохромы из сульфида, и эти цитохромы могут затем перемещать электроны в центр фотосинтеза. Однако не все бактерии зеленого серы продуцируют этот фермент, демонстрируя, что он не требуется для окисления сульфида. Сульфид: хинон -оксидоредуктаза (SQR) также помогает при транспортировке электронов, но, когда только было обнаружено, что приводит к снижению скорости окисления сульфида в бактериях зеленого серы, что позволяет предположить, что существует другой, более эффективный механизм. [ 27 ] Однако большинство бактерий зеленого серы содержат гомолог гена SQR. [ 29 ] Окисление тиосульфата в сульфат может быть катализировано ферментами в СО
X Система. [ 27 ]
Считается, что ферменты и гены, связанные с метаболизмом серы, были получены с помощью горизонтального переноса генов во время эволюции бактерий зеленого серы. [ 29 ]
Углеродная фиксация
[ редактировать ]Зеленая серная бактерии - это фотоавтотрофы : они не только получают энергию от света, но и могут расти, используя углекислый газ в качестве единственного источника углерода. Они фиксируют диоксид углерода, используя цикл обратной трикарбоновой кислоты (RTCA) [ 9 ] где потребляется энергия для уменьшения углекислого газа, а не окислять, как видно в прямом цикле TCA , [ 9 ] Чтобы синтезировать пируват и ацетат . Эти молекулы используются в качестве сырья для синтеза всех строительных блоков, которые необходимо для создания макромолекул . Цикл RTCA очень энергоэффективен, позволяя бактериям расти в условиях низкого освещения. [ 30 ] Однако он имеет несколько чувствительных к кислороду ферментов, которые ограничивают его эффективность в аэробных условиях. [ 30 ]
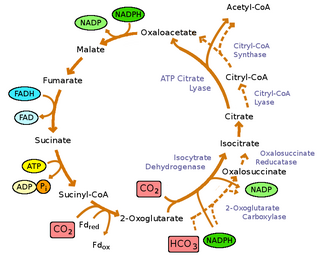
Реакции обращения цикла окислительного трикарбонового кислоты катализируются четырьмя ферментами: [ 9 ]
- пируват: ферредоксин (FD) оксидоредуктаза:
- ацетил-коа + co 2 + 2fdred + 2h + ⇌ pyruvate + coa + 2fdox
- ATP Citrate Lyase:
- ACL, ацетил-CoA + oxaloAcetate + ADP + PI ⇌ Цитрат + COA + ATP
- α-кето-глутарат: оксидоредуктаза ферредоксин:
- Сукцинил-CoA + CO 2 + 2FDRED + 2H + ⇌ α-кетоглутарат + COA + 2FDOX
- Fumarare Reductase
- Succinate + акцептор ⇌ Fumarate + уменьшенный акцептор
Тем не менее, окислительный цикл TCA (OTCA) по -прежнему присутствует в бактериях зеленой серы. OTCA может ассимилировать ацетат, однако OTCA, по -видимому, является неполной в бактериях зеленого серы из -за расположения и падения регуляции гена во время фототрофического роста. [ 9 ]
Миксотрофия
[ редактировать ]Бактерии зеленого серы часто называют облигационными фотоавтотрофами, поскольку они не могут расти при отсутствии света, даже если им обеспечивается органическое вещество. [ 9 ] [ 26 ] Однако они демонстрируют форму миксотрофии , где они могут потреблять простые органические соединения в присутствии света и CO 2 . [ 9 ] В присутствии CO 2 или HCO 3 − , Некоторые бактерии зеленого серы могут использовать ацетат или пируват. [ 9 ]
Миксотрофия в бактериях зеленой серы лучше всего смоделируется репрезентативной зеленой серной бактерией хлорбакулам тепидум . [ 31 ] Миксотрофия возникает во время биосинтеза аминокислот/утилизации углерода и энергетического метаболизма. [ 32 ] Бактерия использует электроны, генерируемые из окисления серы, и энергии, которую она захватывает от света для запуска RTCA. C. Tepidum также демонстрирует использование как пирувата, так и ацетата в качестве источника органического углерода. [ 32 ]
Примером миксотрофии в C. tepidum , который сочетает в себе автотрофию и гетеротрофию, является его синтез ацетил-КоА. C. Tepidum может автотрофически генерировать ацетил-COA через цикл RTCA, или он может гетеротрофически генерировать его от поглощения ацетата. Подобная миксотрофическая активность возникает, когда пируват используется для биосинтеза аминокислоты, но миксотрофный рост с использованием ацетата дает более высокие скорости роста. [ 31 ] [ 32 ]
В энергетическом метаболизме C. tepidum опирается на световые реакции для получения энергии (NADPH и NADH), потому что пути, обычно ответственные за производство энергии (окислительный пентозофосфатный путь и нормальный цикл TCA) лишь частично функциональны. [ 32 ] Фотоны, поглощенные светом, используются для получения NADPH и NADH, кофакторов энергетического метаболизма. C. Tepidum также генерирует энергию в форме АТФ, используя мотив протона, полученную из сульфидного окисления. [ 31 ] Производство энергии как от сульфидного окисления, так и поглощения фотонов посредством бактериохлорофиллов . [ 32 ]
Фиксация азота
[ редактировать ]Большинство бактерий зеленого серы представляют собой диазотрофы : они могут уменьшить азот до аммиака, который затем используется для синтеза аминокислот. [ 33 ] Фиксация азота среди бактерий зеленого серы, как правило, типична для аноксигенного фототрофа и требует наличия света. Бактерии зеленого серы демонстрируют активность из системы секреции типа 1 и ферредоксин-NADP+ оксидоредуктазы для генерации пониженного железа, черты, которая развивалась для поддержки фиксации азота. [ 34 ] Подобно фиолетовым бактериям серы, они могут регулировать активность нитрогеназы посттрансляционно в ответ на концентрации аммиака. Их владение генами NIF , хотя и эволюционно различны, может предположить, что их способности фиксации азота возникают в двух разных событиях или благодаря общему очень отдаленному предку. [ 33 ]
Примеры бактерий зеленого серы, способных к фиксации азота, включают род хлорбиума и пелодион , исключая P. phaeoclathratiforme . Prothecochloris aestuarii и Chloroherpeton Thalassium также попадают в эту категорию. [ 33 ] Их фиксация N 2 широко распространена и играет важную роль в общей доступности азота для экосистем. Зеленая серная бактерии, живущие на коралловых рифах, таких как протекохлорис , имеют решающее значение для создания доступного азота в и без того ограниченной питательными веществами. [ 16 ]
Смотрите также
[ редактировать ]- Аноксическое событие
- Пурпурные бактерии серы
- Зеленые нельфурские бактерии
- Список родов бактерий
- Список бактериального порядка
Ссылки
[ редактировать ]- ^ Gibbons NE, Murray Rge. (1978). «Предложения, касающиеся более высоких таксонов бактерий» . Международный журнал систематической бактериологии . 28 : 1–6. doi : 10.1099/00207713-28-1-1 .
- ^ Garrity GM, Holt JG (2001). "Phylum bxi. Chlorobi Phy. Nov.". В Boone Dr, Castenholz RW, Garrity Gm (Eds.). Руководство по систематической бактериологии Берджи . Тол. 1 (Археи и глубоко ветвящая и фототрофная бактерии) (2 -е изд.). Нью -Йорк, Нью -Йорк: Springer -Verlag. С. 601–623.
- ^ Орен А, Гаррити Г.М. (октябрь 2021 г.). «Действительная публикация названий сорока двух фил прокариот» . Международный журнал систематической и эволюционной микробиологии . 71 (10): 5056. doi : 10.1099/ijsem.0.005056 . PMID 34694987 . S2CID 239887308 .
- ^ "Phylum Chlorobiota " . Список прокариотических имен с стоянием в номенклатуре . 25907 . Получено 22 августа 2023 года .
- ^ Jump up to: а беременный Брайант Д.А., Фригаард Ню (ноябрь 2006 г.). «Прокариотический фотосинтез и фототрофия освещены». Тенденции в микробиологии . 14 (11): 488–496. doi : 10.1016/j.tim.2006.09.001 . PMID 16997562 .
- ^ Зеленый Бр (2003). Светопоглощающие антенны в фотосинтезе . Спрингер. п. 8. ISBN 0792363353 .
- ^ Jump up to: а беременный в дюймовый Kushkevych I, Procházka J, Gajdács M, Rittmann SK, Vítězová M (июнь 2021 г.). «Молекулярная физиология анаэробных фототрофических фиолетовых и зеленых серных бактерий» . Международный журнал молекулярных наук . 22 (12): 6398. DOI : 10.3390/IJMS22126398 . PMC 8232776 . PMID 34203823 .
- ^ Jump up to: а беременный Sakurai H, Ogawa T, Shiga M, Inoue K (июнь 2010 г.). «Система окисления неорганической серы в бактериях зеленой серы». Фотосинтез исследования . 104 (2–3): 163–176. doi : 10.1007/s11120-010-9531-2 . PMID 20143161 . S2CID 1091791 .
- ^ Jump up to: а беременный в дюймовый и фон глин час Tang KH, Blankenship Re (ноябрь 2010 г.). «Как вперед, так и обратные циклы TCA работают в бактериях зеленого серы» . Журнал биологической химии . 285 (46): 35848–35854. doi : 10.1074/jbc.m110.157834 . PMC 2975208 . PMID 20650900 .
- ^ Wahlund TM, Madagan Mt (январь 1993 г.). «Фиксация нитрогена термофической зеленой серной бактерией хлороробий tepidum » Журнал бактераологии 175 (2): 474–4 Doi : 10.1128/ jb.175.2.474-478.1 196162PMC 8093448PMID
- ^ Feng X, Tang KH, Blankenship Re, Tang YJ (декабрь 2010 г.). «Анализ метаболического потока миксотрофических метаболизмов в зеленой бактерии серы хлорбакулум тепидум» . Журнал биологической химии . 285 (50): 39544–39550. doi : 10.1074/jbc.m110.162958 . PMC 2998096 . PMID 20937805 .
- ^ «Зеленая серная бактерии - обзор | темы ScienceDirect» . www.sciencedirect.com . Получено 2022-04-22 .
- ^ Элс (1 изд.). John Wiley & Sons, Ltd. 2001-05-30. doi : 10.1002/9780470015902.a0000458.pub2 . ISBN 978-0-470-01617-6 Полем S2CID 82067054 .
- ^ Marschall E, Jogler M, Hessge U, Overmann J (май 2010). «Крупномасштабные схемы распределения и активности чрезвычайно адаптированной популяции бактерий зеленого серы в Черном море». Экологическая микробиология . 12 (5): 1348–1362. doi : 10.1111/j.1462-2920.2010.02178.x . PMID 20236170 .
- ^ Битти JT, Overmann J, Lince MT, Manske AK, Lang AS, Blankenship RE, et al. (Июнь 2005 г.). «Обязательно фотосинтетический бактериальный анаэроб из глубоководного гидротермального вентиляционного отверстия» . Труды Национальной академии наук Соединенных Штатов Америки . 102 (26): 9306–9310. Bibcode : 2005pnas..102.9306B . doi : 10.1073/pnas.0503674102 . PMC 1166624 . PMID 15967984 .
- ^ Jump up to: а беременный Yang SH, Lee ST, Huang CR, Tseng CH, Chiang PW, Chen CP, et al. (2016-02-26). «Распространенность потенциальных азотных фиксирующих бактерий с зелеными серы в скелете рифового строительства кораллов Isopora palifera» . Лимнология и океанография . 61 (3): 1078–1086. Bibcode : 2016limoc..61.1078y . doi : 10.1002/lno.10277 . ISSN 0024-3590 . S2CID 87463811 .
- ^ Cai L, Zhou G, Tian RM, Tong H, Zhang W, Sun J, et al. (Август 2017). «Метагеномный анализ выявляет зеленую бактерию серы в качестве потенциального кораллового симбионта» . Научные отчеты . 7 (1): 9320. Bibcode : 2017natsr ... 7.9320c . doi : 10.1038/s41598-017-09032-4 . PMC 5571212 . PMID 28839161 .
- ^ "LTP" . Получено 20 ноября 2023 года .
- ^ «Дерево LTP_ALL в формате Newick» . Получено 20 ноября 2023 года .
- ^ «LTP_08_2023 Notes» (PDF) . Получено 20 ноября 2023 года .
- ^ "GTDB Release 08-RS214" . База данных таксономии генома . Получено 10 мая 2023 года .
- ^ "BAC120_R214.sp_label" . База данных таксономии генома . Получено 10 мая 2023 года .
- ^ «История таксонов» . База данных таксономии генома . Получено 10 мая 2023 года .
- ^ Jump up to: а беременный в дюймовый и Bryantseva IA, Tarasov AL, Kostrikina NA, Gaisin VA, Grouzdev DS, Gorlenko VM (декабрь 2019). «Протекохлорис Марина Сп. Ноябрь, новая зеленая бактерия серы из прибрежной зоны Южно -Китайского моря». Архив микробиологии . 201 (10): 1399–1404. doi : 10.1007/s00203-019-01707-y . PMID 31338544 . S2CID 198190182 .
- ^ Jump up to: а беременный Hauska G, Schoedl T, Remigy H, Tsiotis G (октябрь 2001 г.). «Реакционный центр бактерий зеленого серы (1)» . Biochimica et Biophysica Acta . 1507 (1–3): 260–277. doi : 10.1016/s0005-2728 (01) 00200-6 . PMID 11687219 .
- ^ Jump up to: а беременный Лигрон Р. (2019). «Переезд на свет: эволюция фотосинтеза» . В Лигроне р (ред.). Биологические инновации, которые построили мир: четыре миллиона лет путешествия по жизни и истории Земли . Cham: Springer International Publishing. С. 99–127. doi : 10.1007/978-3-030-16057-9_4 . ISBN 978-3-030-16057-9 Полем S2CID 189992218 . Получено 2021-01-29 .
- ^ Jump up to: а беременный в дюймовый Frigaard Nu, Dahl C (январь 2008 г.). «Сервный метаболизм в фототрофных бактериях серы» . В Пуле Р.К. (ред.). Достижения в области микробной физиологии . Тол. 54. Академическая пресса. С. 103–200 . Получено 2022-04-22 .
- ^ Ван Гемерден H (октябрь 1986 г.). «Производство элементарной серы зелеными и фиолетовыми бактериями серы» . Архив микробиологии . 146 (1): 52–56. doi : 10.1007/bf00690158 . ISSN 1432-072X . S2CID 30812886 .
- ^ Jump up to: а беременный Грегерсен Л.Х., Брайант Д.А., Фригаард Ню (2011). «Механизмы и эволюция окислительного метаболизма серы в бактериях зеленого серы» . Границы в микробиологии . 2 : 116. doi : 10.3389/fmicb.2011.00116 . PMC 3153061 . PMID 21833341 .
- ^ Jump up to: а беременный Bar-Evon A, Noor E, Milo R (март 2012 г.). «Обследование путей фиксации углерода через количественную линзу» . Журнал экспериментальной ботаники . 63 (6): 2325–2342. doi : 10.1093/jxb/err417 . PMID 22200662 .
- ^ Jump up to: а беременный в Frigaard Nu, Chew AG, Li H, Maresca JA, Bryant DA (2003). «Chlorobium tepidum: понимание структуры, физиологии и метаболизма зеленой бактерии серы, полученной из полной последовательности генома». Фотосинтез исследования . 78 (2): 93–117. doi : 10.1023/b: pres.0000004310.96189.b4 . PMID 16245042 . S2CID 30218833 .
- ^ Jump up to: а беременный в дюймовый и Feng X, Tang KH, Blankenship Re, Tang YJ (декабрь 2010 г.). «Анализ метаболического потока миксотрофических метаболизмов в зеленой бактерии серы хлорбакулум тепидум» . Журнал биологической химии . 285 (50): 39544–39550. doi : 10.1074/jbc.m110.162958 . PMC 2998096 . PMID 20937805 .
- ^ Jump up to: а беременный в Мэдиган М.Т. (1995). «Микробиология азотной фиксации с помощью аноксигенных фотосинтетических бактерий». В Blankenship Re , Madigan Mt, Bauer CE (ред.). Аноксигенные фотосинтетические бактерии . Достижения в области фотосинтеза и дыхания. Тол. 2. Дордрехт: Спрингер Нидерланды. С. 915–928. doi : 10.1007/0-306-47954-0_42 . ISBN 978-0-306-47954-0 .
- ^ Mus F, Colman DR, Peters JW, Boyd ES (август 2019 г.). «Геобиологические отзывы, кислород и эволюция нитрогеназы» . Свободная радикальная биология и медицина . Ранняя жизнь на Земле и окислительный стресс. 140 : 250–259. doi : 10.1016/j.freeradbiomed.2019.01.050 . PMID 30735835 . S2CID 73433517 .
Внешние ссылки
[ редактировать ]- "Семейство хлорбиааа" . Прокариоты . Архивировано с оригинала 17 ноября 2003 года . Получено 5 июля 2005 года .