Третичная структура нуклеиновой кислоты

Третичная структура нуклеиновой кислоты представляет собой трехмерную форму полимера нуклеиновой кислоты . [1] РНК и ДНК Молекулы способны выполнять разнообразные функции — от молекулярного распознавания до катализа . Такие функции требуют точной трехмерной структуры. Хотя такие структуры разнообразны и кажутся сложными, они состоят из повторяющихся, легко узнаваемых третичных структурных мотивов , которые служат молекулярными строительными блоками. Некоторые из наиболее распространенных мотивов третичной структуры РНК и ДНК описаны ниже, но эта информация основана на ограниченном количестве решенных структур. По мере структурной характеристики новых молекул РНК и ДНК будет обнаружено еще больше третичных структурных мотивов.
Спиральные структуры
[ редактировать ]
Двойная спираль
[ редактировать ]Двойная спираль является доминирующей третичной структурой биологической ДНК, а также возможной структурой РНК. Считается, что в природе встречаются три конформации ДНК: A-ДНК , B-ДНК и Z-ДНК . форма «B», описанная Джеймсом Д. Уотсоном и Фрэнсисом Криком . Считается, что в клетках преобладает [2] Джеймс Д. Уотсон и Фрэнсис Крик описали эту структуру как двойную спираль с радиусом 10 Å и шагом 34 Å , совершающую один полный оборот вокруг своей оси каждые 10 пар оснований последовательности. [3] Двойная спираль делает один полный оборот вокруг своей оси каждые 10,4–10,5 пар оснований в растворе. Эта частота скручивания (известная как шаг спирали ) во многом зависит от сил укладки, которые каждое основание оказывает на своих соседей в цепи. Двойная спиральная РНК принимает конформацию, аналогичную структуре А-формы.
Возможны и другие конформации; Фактически, теперь для описания любой новой структуры ДНК, которая может появиться в будущем, доступны только буквы F, Q, U, V и Y. [4] [5] Однако большинство этих форм созданы синтетически и не наблюдались в природных биологических системах.

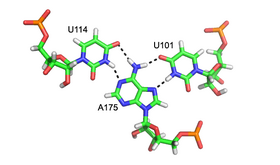
Триплексы с большими и малыми канавками
[ редактировать ]Триплекс малой бороздки представляет собой повсеместный РНК структурный мотив . Поскольку взаимодействие с малой бороздкой часто опосредовано 2'-ОН сахара рибозы , этот мотив РНК сильно отличается от своего ДНК- эквивалента. Наиболее распространенным примером тройки малой бороздки является мотив A-минор или вставка аденозиновых оснований в малую бороздку (см. Выше). Однако этот мотив не ограничивается аденозинами, поскольку другие нуклеиновые основания наблюдалось также, что взаимодействуют с малой бороздкой РНК.
Малая канавка представляет собой почти идеальное дополнение для вставленного основания. Это обеспечивает оптимальные контакты Ван-дер-Ваальса , обширные водородные связи и гидрофобную поверхность, а также создает очень энергетически выгодное взаимодействие. [8] [9] Поскольку тройки малых бороздок способны стабильно упаковывать свободную петлю и спираль, они являются ключевыми элементами в структуре крупных рибонуклеотидов , включая интрон группы I. [10] интрон группы II, [11] и рибосома .
Хотя большая бороздка стандартной А-формы РНК довольно узкая и, следовательно, менее доступна для триплексного взаимодействия, чем малая бороздка, триплексные взаимодействия с большой бороздкой можно наблюдать в некоторых структурах РНК. Эти структуры состоят из нескольких комбинаций пар оснований и взаимодействий Хугстина. Например, триплекс GGC (GGC амино(N-2)-N-7, имино-карбонил, карбонил-амино(N-4); Уотсон-Крик), наблюдаемый в 50S рибосоме , состоит из GC типа Уотсона-Крика . пара и входящий G, который образует псевдо-Хугстинову сеть взаимодействий водородных связей между обоими основаниями, участвующими в каноническом спаривании. [12] Другие примечательные примеры триплексов с большими бороздками включают (i) каталитическое ядро интрона группы II, показанное на рисунке слева. [6] (ii) каталитически необходимая тройная спираль, наблюдаемая в теломеразной РНК человека [7] (iii) рибопереключатель SAM-II [14] и (iv) элемент ядерной экспрессии (ENE), который действует как элемент стабилизации РНК посредством образования тройной спирали с поли(А)-хвостом. [15] [16]
Трехцепочечная ДНК также возможна из- за связей Хугстина или обратных водородных связей Хугстина в основной бороздке B-формы ДНК .
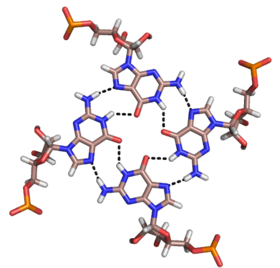
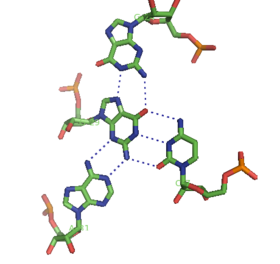
Квадруплексы
[ редактировать ]Помимо двойных спиралей и вышеупомянутых триплексов, РНК и ДНК могут также образовывать четверные спирали. Существуют разнообразные структуры квадруплексов оснований РНК. Четыре последовательных остатка гуанина могут образовывать квадруплекс в РНК за счет водородных связей Хугстина, образуя «кольцо Хугстина» (см. рисунок). [12] Пары GC и AU также могут образовывать базовый квадруплекс с комбинацией спаривания Уотсона-Крика и неканонического спаривания в малой бороздке . [17]
Ядро аптамера малахитового зеленого также представляет собой разновидность базового квадруплекса с другим паттерном водородных связей (см. Рисунок). [13] Квадруплекс может повторяться несколько раз подряд, создавая чрезвычайно стабильную структуру.
Уникальная структура квадруплексных участков РНК может выполнять различные функции в биологической системе. Двумя важными функциями являются потенциал связывания с лигандами или белками и его способность стабилизировать всю третичную структуру ДНК или РНК. Сильная структура может ингибировать или модулировать транскрипцию и репликацию , например, в теломерах хромосом и UTR мРНК. [18] Идентичность основания важна для связывания лиганда. G-квартет обычно связывает одновалентные катионы, такие как калий, в то время как другие основания могут связывать множество других лигандов, таких как гипоксантин, в квадруплексе UUCU. [17]
Наряду с этими функциями G-квадруплекс в мРНК вокруг областей связывания рибосом может служить регулятором экспрессии генов у бактерий . [19] Возможно, существуют еще более интересные структуры и функции, которые еще предстоит открыть in vivo .
Коаксиальная укладка
[ редактировать ]
Коаксиальная укладка, также известная как спиральная укладка, является основным фактором, определяющим третичную структуру РНК более высокого порядка. Коаксиальная укладка происходит, когда два дуплекса РНК образуют непрерывную спираль, которая стабилизируется за счет укладки оснований на границе раздела двух спиралей. отмечена коаксиальная укладка . В кристаллической структуре тРНКФе [21] Совсем недавно коаксиальная укладка наблюдалась в структурах более высокого порядка многих рибозимов , включая многие формы самосплайсинговых интронов группы I и группы II . Общие мотивы коаксиальной укладки включают взаимодействие целующейся петли и псевдоузла . Устойчивость этих взаимодействий можно предсказать путем адаптации «правил Тернера». [22]
В 1994 году Уолтер и Тернер определили вклад свободной энергии стекинг-взаимодействий ближайших соседей внутри границы раздела спираль-спираль, используя модельную систему, которая создавала интерфейс спираль-спираль между коротким олигомером и выступающим элементом из четырех нуклеотидов на конце шпильки . корень . Их эксперименты подтвердили, что термодинамический вклад укладки оснований между двумя спиральными вторичными структурами близко имитирует термодинамику образования стандартного дуплекса (взаимодействия ближайших соседей предсказывают термодинамическую стабильность образующейся спирали). Относительная стабильность взаимодействий ближайших соседей может быть использована для прогнозирования благоприятной коаксиальной упаковки на основе известной вторичной структуры. Уолтер и Тернер обнаружили, что в среднем точность предсказания структуры РНК улучшилась с 67% до 74%, когда были включены вклады коаксиального укладки. [23]
Большинство хорошо изученных третичных структур РНК содержат примеры коаксиальной укладки. Некоторыми яркими примерами являются тРНК-Phe, интроны группы I, интроны группы II и рибосомальные РНК. Кристаллические структуры тРНК выявили наличие двух удлиненных спиралей, образующихся в результате коаксиальной укладки акцепторного стебля аминокислот с Т-плечом, а также укладки D- и антикодонов. Эти взаимодействия внутри тРНК ориентируют стебель антикодона перпендикулярно стеблю аминокислоты, что приводит к функциональной L-образной третичной структуре. [21] Было показано, что в интронах группы I спирали P4 и P6 коаксиально укладываются друг на друга с помощью комбинации биохимических процессов. [24] и кристаллографические методы. Кристаллическая структура P456 дала подробное представление о том, как коаксиальная укладка стабилизирует упаковку спиралей РНК в третичные структуры. [25] В интроне группы самосплайсинга II Oceanobacillus iheyensis стебли IA и IB коаксиально укладываются друг на друга и способствуют относительной ориентации составляющих спиралей пятистороннего соединения. [6] Такая ориентация способствует правильному сворачиванию активного центра функционального рибозима. Рибосома содержит многочисленные примеры коаксиальной укладки, включая сложенные сегменты длиной до 70 п.н. [26]

Двумя распространенными мотивами, включающими коаксиальную укладку, являются петли поцелуев и псевдоузлы. При взаимодействии целующихся петель одноцепочечные области петель двух шпилек взаимодействуют посредством спаривания оснований, образуя сложную, коаксиально уложенную спираль. Примечательно, что эта структура позволяет всем нуклеотидам в каждой петле участвовать во взаимодействиях спаривания оснований и штабелирования. Этот мотив был визуализирован и изучен с помощью ЯМР-анализа Ли и Кротерсом. [27] Мотив псевдоузла возникает, когда одноцепочечная область петли шпильки соединяется с последовательностью выше или ниже в пределах одной и той же цепи РНК. Две образующиеся дуплексные области часто накладываются друг на друга, образуя стабильную коаксиально сложенную составную спираль. Одним из примеров мотива псевдоузла является высокостабильный рибозим вируса гепатита Дельта, основная цепь которого демонстрирует общую топологию двойного псевдоузла. [28]
Эффект, подобный коаксиальному стэкингу, наблюдался в рационально спроектированных структурах ДНК. Структуры ДНК-оригами содержат большое количество двойных спиралей с открытыми тупыми концами. Было замечено, что эти структуры слипаются по краям, содержащим эти открытые тупые концы, из-за гидрофобных взаимодействий при штабелировании. [29] Объединив эти рационально спроектированные наноструктуры ДНК и визуализацию суперразрешения DNA-PAINT, исследователи выявили индивидуальную силу энергий укладки между всеми возможными динуклеотидами. [30]
Измерение коаксиальной укладки нуклеиновой кислоты
[ редактировать ]Ранние измерения коаксиального стэкинга проводились с использованием биохимических анализов, которые изучают относительную миграцию различных молекул нуклеиновых кислот в зависимости от их конформации и типа присутствующих взаимодействий. Короткие молекулы ДНК, содержащие разрывы, которые все еще могли укладываться коаксиально, мигрировали быстрее, чем молекулы ДНК, содержащие разрывы, и, следовательно, не имели коаксиальной укладки. Это можно объяснить полимерными свойствами ДНК: более жесткие стержнеобразные молекулы будут быстрее мигрировать по электрическому градиенту в матрице по сравнению с более гибкими молекулами. [31] Разработка новых методов, таких как оптические пинцеты и способность складывать наноструктуры ДНК, привела к измерению пучков ДНК и их способности складываться друг в друга. Затем можно проанализировать силу, необходимую для разрыва этих пучков с помощью оптического пинцета, для измерения энергий укладки пар оснований. [32] Эти измерения проводились в основном в неравновесных условиях, и были сделаны различные экстраполяции для получения точных значений коаксиальной укладки между основаниями. Недавние исследования одиночных молекул с использованием наноструктур ДНК и микроскопии сверхвысокого разрешения DNA-PAINT позволили измерить это взаимодействие между динуклеотидами с помощью углубленного кинетического анализа времени связывания коротких молекул ДНК с их комплементарными последовательностями в присутствии или отсутствии ДНК-нуклеотидов. штабелирование взаимодействий. [30]
Другие мотивы
[ редактировать ]Взаимодействие тетрапетли-рецептора
[ редактировать ]
Взаимодействия тетрапетля-рецептор сочетают в себе взаимодействия спаривания оснований и укладки между петлевыми нуклеотидами мотива тетрапетли и мотива рецептора, расположенного внутри дуплекса РНК, создавая третичный контакт, который стабилизирует глобальную третичную укладку молекулы РНК . Тетрапетли также являются возможными структурами в дуплексах ДНК. [34]
Стеблевые петли могут сильно различаться по размеру и последовательности, но тетрапетли из четырех нуклеотидов очень распространены и обычно относятся к одной из трех категорий в зависимости от последовательности. [35] Эти три семейства представляют собой тетралупы CUYG, UNCG и GNRA (см. рисунок справа) . [36] В каждом из этих семейств тетрапетли второй и третий нуклеотиды образуют поворот в цепи РНК, а пара оснований между первым и четвертым нуклеотидами стабилизирует структуру стебель-петли. В целом установлено, что стабильность тетрапетли зависит от состава оснований внутри петли и от состава этой «замыкающей пары оснований». [37] Семейство тетрапетлей GNRA чаще всего наблюдается во взаимодействиях тетрапетля-рецептор. Кроме того, известно, что тетрапетли UMAC являются альтернативными версиями петель GNRA, обе имеют схожие структуры основной цепи; несмотря на сходство, они различаются возможными дальнодействующими взаимодействиями, на которые способны. [38]

«Мотивы рецептора тетрапетли» представляют собой третичные взаимодействия дальнего действия. [39] состоящий из водородных связей между основаниями тетрапетли и последовательностями стебельковой петли в дистальных участках вторичной структуры РНК. [40] Помимо водородных связей, важным компонентом этих третичных взаимодействий являются стэкинг-взаимодействия. Например, во взаимодействиях GNRA-тетрапетля второй нуклеотид тетрапетли укладывается непосредственно на мотив A-платформы (см. Выше) внутри рецептора. [25] Последовательность тетрапетли и ее рецептора часто коварна, так что один и тот же тип третичного контакта может быть установлен с разными изоформами тетрапетли и родственного ей рецептора. [41]
Например, структура и функция самосплайсируемого интрона группы I зависят от мотивов тетрапетлевого рецептора. [25] [40] В частности, три адениновых остатка канонического мотива GAAA укладываются на вершину спирали рецептора и образуют с рецептором множественные стабилизирующие водородные связи. Первый аденин последовательности GAAA образует тройную пару оснований с основаниями AU рецептора. Второй аденин стабилизируется за счет водородных связей с тем же уридином, а также через его 2'-ОН с рецептором и за счет взаимодействий с гуанином тетрапетли GAAA. Третий аденин образует тройную пару оснований.
Мотив ля минор
[ редактировать ]

А-минорный мотив представляет собой повсеместно встречающийся третичный структурный мотив РНК. Он образуется путем вставки неспаренного нуклеозида в малую бороздку дуплекса РНК. По сути, это пример тройки минорных грувов . Хотя гуанозин, цитозин и уридин также могут образовывать тройные взаимодействия малых бороздок, взаимодействия малых бороздок с аденином очень распространены. В случае аденина край N1-C2-N3 внедряющего основания образует водородные связи с одним или обоими 2'-ОН дуплекса, а также с основаниями дуплекса (см. Рисунок: А-минорные взаимодействия ). Дуплекс хоста часто представляет собой базовую пару GC.
А-минорные мотивы разделены на четыре класса: [8] [9] типы от 0 до III, в зависимости от положения вставляющего основания относительно двух 2'-ОН пары оснований Уотсона-Крика . В A-минорных мотивах типа I и II N3 аденина вставлен глубоко в малую бороздку дуплекса (см. Рисунок: Взаимодействия A-минор - взаимодействие типа II), и имеется хорошая комплементарность формы с парой оснований. В отличие от типов 0 и III, взаимодействия типа I и II специфичны для аденина за счет взаимодействий по водородным связям. При взаимодействии типа III и O2', и N3 вставочного основания менее тесно связаны с малой бороздкой дуплекса. Мотивы типа 0 и III более слабы и неспецифичны, поскольку они опосредованы взаимодействиями с одним 2'-ОН (см. Рисунок: А-минорные взаимодействия - взаимодействия типа 0 и типа III).
А-минорный мотив является одним из наиболее распространенных структурных мотивов РНК в рибосоме, где он способствует связыванию тРНК с субъединицей 23S. [43] Чаще всего они стабилизируют дуплексные взаимодействия РНК в петлях и спиралях, например, в ядре интронов группы II. [6]
Интересным примером А-минора является его роль в распознавании антикодонов . Рибосома должна различать правильные и неправильные пары кодон-антикодон. Частично это происходит за счет внедрения адениновых оснований в малую бороздку. Неправильные пары кодон-антикодон будут иметь искаженную спиральную геометрию, что не позволит А-минорному взаимодействию стабилизировать связывание и увеличит скорость диссоциации неправильной тРНК. [44]
Анализ A-минорных мотивов в 23S рибосомальной РНК выявил иерархическую сеть структурных зависимостей, которые, как предполагается, связаны с эволюцией рибосом и порядком событий, которые привели к развитию современной бактериальной большой субъединицы. [45]
Сообщается, что A-минорный мотив и его новый подкласс, WC/H A-минорные взаимодействия, укрепляют другие третичные структуры РНК, такие как тройные спирали с большой бороздкой, идентифицированные в элементах стабилизации РНК. [16] [15]
Молния рибоза
[ редактировать ]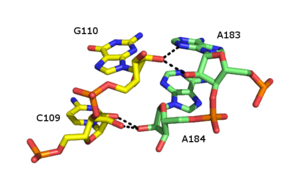
Рибозная молния представляет собой третичный структурный элемент РНК , в котором две цепи РНК удерживаются вместе за счет взаимодействий водородных связей с участием 2'OH сахаров рибозы на разных цепях. 2'OH может вести себя как донор и акцептор водородной связи, что позволяет образовывать раздвоенные водородные связи с другим 2'OH. [46] [47]
Сообщалось о многочисленных формах рибозной застежки, но общий тип включает четыре водородные связи между 2'-ОН-группами двух соседних сахаров. Рибозные молнии обычно встречаются в виде массивов, которые стабилизируют взаимодействия между отдельными цепями РНК. [48] Застежки-молнии рибозы часто наблюдаются как взаимодействия стебель-петля с очень низкой специфичностью последовательности. Однако в малых и больших субъединицах рибосом существует склонность к образованию рибозных застежек последовательности CC/AA — два цитозина в первой цепи соединены с двумя аденинами во второй цепи.
Роль ионов металлов
[ редактировать ]
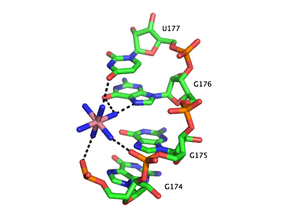
Функциональные РНК часто представляют собой свернутые стабильные молекулы трехмерной формы, а не гибкие линейные нити. [50] Катионы необходимы для термодинамической стабилизации третичных структур РНК. Катионы металлов, связывающие РНК, могут быть одновалентными, двухвалентными и трехвалентными. Калий (К + ) — обычный одновалентный ион, связывающий РНК. Обычным двухвалентным ионом, связывающим РНК, является магний (Mg 2+ ). Другие ионы, включая натрий (Na + ), кальций (Ca 2+ ) и марганец (Mn 2+ ) было обнаружено, что они связывают РНК in vivo и in vitro . Многовалентные органические катионы, такие как спермидин или спермин, также обнаруживаются в клетках и вносят важный вклад в сворачивание РНК. Трехвалентные ионы, такие как гексамин кобальта, или ионы лантаноидов, такие как тербий (Tb 3+ ) являются полезными экспериментальными инструментами для изучения связывания металлов с РНК. [51] [52]
Ион металла может взаимодействовать с РНК несколькими способами. Ион может диффузно ассоциироваться с основной цепью РНК, экранируя в противном случае неблагоприятные электростатические взаимодействия . Такое экранирование заряда часто осуществляется одновалентными ионами. Ионы, связанные с сайтом, стабилизируют определенные элементы третичной структуры РНК. Взаимодействия, связанные с сайтом, можно разделить на две категории в зависимости от того, опосредует ли вода связывание металла. Взаимодействия «внешней сферы» опосредуются молекулами воды, окружающими ион металла. Например, гексагидрат магния взаимодействует и стабилизирует определенные мотивы третичной структуры РНК посредством взаимодействия с гуанозином в большой бороздке. И наоборот, взаимодействия «внутренней сферы» напрямую опосредуются ионом металла. РНК часто сворачивается в несколько этапов, и эти этапы можно стабилизировать различными типами катионов. На ранних стадиях РНК образует вторичные структуры , стабилизированные за счет связывания одновалентных катионов, двухвалентных катионов и полианионных аминов с целью нейтрализации полианионной основной цепи. Более поздние стадии этого процесса включают формирование третичной структуры РНК, которая стабилизируется почти в основном за счет связывания двухвалентных ионов, таких как магний, с возможным вкладом связывания калия.
Сайты связывания металлов часто локализуются в глубокой и узкой большой бороздке дуплекса РНК, координируясь с хугстиновыми краями пуринов . В частности, катионы металлов стабилизируют места скручивания основной цепи, где плотная упаковка фосфатов приводит к образованию области плотного отрицательного заряда. В дуплексах РНК имеется несколько мотивов, связывающих ионы металлов, которые были идентифицированы в кристаллических структурах. Например, в домене P4-P6 Tetrahymena thermophila интрона группы I несколько сайтов связывания ионов состоят из тандемных пар GU-колебаний и тандемных несоответствий GA , в которых двухвалентные катионы взаимодействуют с гуанозиновым краем Хугстина через O6 и N7. [53] [54] [55] Другим ион-связывающим мотивом в интроне группы I тетрахимены является мотив платформы AA, в котором последовательные аденозины в одной и той же цепи РНК образуют неканоническую пару псевдооснований. [56] В отличие от тандемного мотива GU, мотив платформы AA преимущественно связывается с одновалентными катионами. Во многих из этих мотивов отсутствие одновалентных или двухвалентных катионов приводит либо к большей гибкости, либо к потере третичной структуры.
ионы двухвалентных металлов, особенно магния Было обнаружено, что , важны для структуры соединений ДНК, таких как промежуточное соединение Холлидея в генетической рекомбинации . Ион магния экранирует отрицательно заряженные фосфатные группы в соединении и позволяет им располагаться ближе друг к другу, создавая сложную конформацию, а не несложенную конформацию. [57] Магний жизненно важен для стабилизации такого рода соединений в искусственно созданных структурах, используемых в нанотехнологиях ДНК , таких как мотив двойного кроссовера. [58]
История
[ редактировать ]Самые ранние работы в области структурной биологии РНК более или менее совпадали с работами по ДНК, проводившимися в начале 1950-х годов. В своей основополагающей статье 1953 года Уотсон и Крик предположили, что скученность Ван-дер-Ваальса 2`ОН-группой рибозы не позволит РНК принять двойную спиральную структуру, идентичную предложенной ими модели – то, что мы теперь знаем как ДНК B-формы. [59] Это вызвало вопросы о трехмерной структуре РНК: может ли эта молекула образовывать спиральную структуру того или иного типа, и если да, то как?
В середине 1960-х годов интенсивно изучалась роль тРНК в синтезе белка. В 1965 году Холли и др. очистили и секвенировали первую молекулу тРНК, первоначально предположив, что она приняла структуру клеверного листа, основанную в основном на способности определенных участков молекулы образовывать структуры стебельковой петли. [60] Выделение тРНК оказалось первой крупной находкой в структурной биологии РНК. В 1971 году Ким и др. добился еще одного прорыва, получив кристаллы дрожжевой тРНК. ПТО это дифрагировало до разрешения 2-3 ангстрема с использованием спермина, природного полиамина , который связывался и стабилизировал тРНК. [61]
В течение значительного времени после появления первых структур тРНК область структуры РНК существенно не продвинулась вперед. Возможность изучения структуры РНК зависела от возможности выделить РНК-мишень. На протяжении многих лет это ограничивало область исследования, отчасти потому, что другие известные мишени, например, рибосому , было значительно сложнее изолировать и кристаллизовать. Таким образом, в течение примерно двадцати лет после первой публикации тРНК ПТО структура, были решены структуры лишь нескольких других мишеней РНК, причем почти все из них принадлежали к семейству транспортных РНК. [62]
Это досадное отсутствие возможностей в конечном итоге будет преодолено во многом благодаря двум крупным достижениям в исследованиях нуклеиновых кислот: идентификации рибозимов и способности производить их посредством транскрипции in vitro . После публикации Тома Чеха, в которой интрон группы I тетрахимены является автокаталитическим рибозимом, [63] и отчет Сидни Альтмана о катализе рибонуклеазой P РНК, [64] несколько других каталитических РНК были идентифицированы в конце 1980-х годов, [65] включая рибозим «головка молотка». В 1994 году Маккей и др. опубликовали структуру «комплекса рибозим-ингибитор РНК-ДНК с головкой молотка» с разрешением 2,6 ангстрема, в которой автокаталитическая активность рибозима нарушалась за счет связывания с ДНК-субстратом. [66] Помимо успехов в определении глобальной структуры с помощью кристаллографии, в начале 1990-х годов также был внедрен ЯМР как мощный метод в структурной биологии РНК. Подобные исследования позволили более точно охарактеризовать взаимодействия спаривания и укладки оснований, которые стабилизировали глобальные складки больших молекул РНК.
Возрождение структурной биологии РНК в середине 1990-х годов вызвало настоящий взрыв в области структурных исследований нуклеиновых кислот. С момента публикации структур «головка-молот» и P 4-6 в эту область было внесено множество крупных вкладов. Некоторые из наиболее примечательных примеров включают структуры интронов группы I и группы II . [6] и Рибосома . [42] Первые три структуры были созданы с использованием транскрипции in vitro , и тот факт, что ЯМР сыграл роль в исследовании частичных компонентов всех четырех структур, свидетельствует о незаменимости обоих методов для исследования РНК. Нобелевская премия по химии 2009 года была присуждена Аде Йонат , Венкатраману Рамакришнану и Томасу Стейтцу за их структурные работы над рибосомой , продемонстрировавшие выдающуюся роль, которую структурная биология РНК сыграла в современной молекулярной биологии.
См. также
[ редактировать ]- Стебель-петля
- Псевдоузел
- Прогнозирование вторичной структуры
- Базовая пара
- Колебание базовой пары
- Базовая пара Хугстина
- Рибосома
- Рибопереключатель
- Рибозим
- Рибозим молотоголового типа
- Каталитический интрон группы I
- Интрон группы II
- тРНК
- G-квадруплекс
- i-мотив ДНК
- Тетралуп
- Скользкая последовательность
- Поцелуй стебель-петля
Ссылки
[ редактировать ]- ^ ИЮПАК , Сборник химической терминологии , 2-е изд. («Золотая книга») (1997). Исправленная онлайн-версия: (2006–) « Третичная структура ». два : 10.1351/goldbook.T06282
- ^ Ричмонд Т.Дж., Дэйви, Калифорния (май 2003 г.). «Строение ДНК в ядре нуклеосомы». Природа . 423 (6936): 145–50. Бибкод : 2003Natur.423..145R . дои : 10.1038/nature01595 . ПМИД 12736678 . S2CID 205209705 .
- ^ Уотсон Дж. Д., Крик Ф. Х. (апрель 1953 г.). «Молекулярная структура нуклеиновых кислот; структура нуклеиновой кислоты дезоксирибозы» (PDF) . Природа . 171 (4356): 737–8. Бибкод : 1953Natur.171..737W . дои : 10.1038/171737a0 . ПМИД 13054692 . S2CID 4253007 .
- ^ Бансал М (2003). «Структура ДНК: возвращение к двойной спирали Уотсона-Крика». Современная наука . 85 (11): 1556–1563.
- ^ Гош А., Бансал М. (2003). «Словарь структур ДНК от А до Я». Акта Кристаллогр Д. 59 (4): 620–626. дои : 10.1107/S0907444903003251 . ПМИД 12657780 .
- ^ Jump up to: а б с д и ВВП : 3BWP ; Тор Н., Китинг К.С., Тейлор С.Д., Пайл А.М. (апрель 2008 г.). «Кристаллическая структура самосплайсируемого интрона группы II» . Наука . 320 (5872): 77–82. Бибкод : 2008Sci...320...77T . дои : 10.1126/science.1153803 . ПМК 4406475 . ПМИД 18388288 . ; визуализируется с помощью PyMOL
- ^ Jump up to: а б ПКБ : 2К95 ; Ким Н.К., Чжан К., Чжоу Дж., Теймер К.А., Петерсон Р.Д., Фейгон Дж. (декабрь 2008 г.). «Структура раствора и динамика псевдоузла теломеразной РНК дикого типа человека» . Дж. Мол. Биол . 384 (5): 1249–61. дои : 10.1016/j.jmb.2008.10.005 . ПМК 2660571 . ПМИД 18950640 . ; визуализируется с помощью PyMOL
- ^ Jump up to: а б Ниссен П., Ипполито Дж.А., Бан Н., Мур П.Б., Стейц Т.А. (апрель 2001 г.). «Третичные взаимодействия РНК в большой субъединице рибосомы: А-минорный мотив» . Учеб. Натл. акад. наук. США . 98 (9): 4899–903. Бибкод : 2001PNAS...98.4899N . дои : 10.1073/pnas.081082398 . ПМК 33135 . ПМИД 11296253 .
- ^ Jump up to: а б Доэрти Э.А., Бэти Р.Т., Маскида Б., Дудна Дж.А. (апрель 2001 г.). «Универсальный способ упаковки спирали в РНК». Нат. Структура. Биол . 8 (4): 339–43. дои : 10.1038/86221 . ПМИД 11276255 . S2CID 213577 .
- ^ Шевчак А.А., Ортолева-Доннелли Л., Райдер С.П., Монкер Э., Штробель С.А. (декабрь 1998 г.). «Тройная спираль РНК с малой бороздкой внутри каталитического ядра интрона группы I». Нат. Структура. Биол . 5 (12): 1037–42. дои : 10.1038/4146 . ПМИД 9846872 . S2CID 10908125 .
- ^ Будвиллен М., де Ленкастр А., Пайл А.М. (июль 2000 г.). «Третичное взаимодействие, которое связывает домены активного сайта с 5'-сайтом сплайсинга интрона группы II». Природа . 406 (6793): 315–8. Бибкод : 2000Natur.406..315B . дои : 10.1038/35018589 . ПМИД 10917534 . S2CID 4336795 .
- ^ Jump up to: а б с ПДБ : 1РАУ ; Чеонг С., Мур П.Б. (сентябрь 1992 г.). «Структура раствора необычайно стабильного тетраплекса РНК, содержащего структуры G- и U-квартета». Биохимия . 31 (36): 8406–14. дои : 10.1021/bi00151a003 . ПМИД 1382577 . ; визуализируется с помощью PyMOL
- ^ Jump up to: а б ВВП : 1FIT ; Боуг С., Грейт Д., Уилсон С. (август 2000 г.). «2.8 Кристаллическая структура аптамера малахитового зеленого». Дж. Мол. Биол . 301 (1): 117–28. дои : 10.1006/jmbi.2000.3951 . ПМИД 10926496 . ; визуализируется с помощью PyMOL
- ^ Гилберт С.Д., Рэмбо Р.П., Ван Тайн Д., Бэти RT (февраль 2008 г.). «Структура рибопереключателя SAM-II, связанного с S-аденозилметионином». Нат. Структура. Мол. Биол . 15 (2): 177–82. дои : 10.1038/nsmb.1371 . ПМИД 18204466 . S2CID 40791601 .
- ^ Jump up to: а б Миттон-Фрай Р.М., ДеГрегорио С.Дж., Ван Дж., Стейц Т.А., Стейц Дж.А. (ноябрь 2010 г.). «Распознавание хвоста поли(А) элементом вирусной РНК посредством сборки тройной спирали» . Наука . 330 (6008): 1244–7. Бибкод : 2010Sci...330.1244M . дои : 10.1126/science.1195858 . ПМК 3074936 . ПМИД 21109672 .
- ^ Jump up to: а б Тораби С.Ф., Вайдья А.Т., Тыковски К.Т., ДеГрегорио С.Дж., Ван Дж., Шу, М.Д. и др. (январь 2021 г.). «Стабилизация РНК с помощью кармана связывания 3'-конца поли(А)-хвоста и других способов взаимодействия поли(А)-РНК» . Наука . 371 (6529). дои : 10.1126/science.abe6523 . ISSN 0036-8075 . ПМЦ 9491362 . ПМИД 33414189 . S2CID 231195473 .
- ^ Jump up to: а б Бэти Р.Т., Гилберт С.Д., Монтанж Р.К. (ноябрь 2004 г.). «Структура природного рибопереключателя, реагирующего на гуанин, в комплексе с метаболитом гипоксантином». Природа . 432 (7015): 411–5. Бибкод : 2004Natur.432..411B . дои : 10.1038/nature03037 . ПМИД 15549109 . S2CID 2462025 .
- ^ Артханари Х., Болтон, П.Х. (март 2001 г.). «Функциональная и дисфункциональная роль квадруплексной ДНК в клетках» . хим. Биол . 8 (3): 221–30. дои : 10.1016/S1074-5521(01)00007-2 . ПМИД 11306347 .
- ^ Оливер А.В., Богдарина И., Шредер Э., Тейлор И.А., Нил Г.Г. (август 2000 г.). «Преимущественное связывание белка гена 5 fd со структурами тетраплексных нуклеиновых кислот». Дж. Мол. Биол . 301 (3): 575–84. дои : 10.1006/jmbi.2000.3991 . ПМИД 10966771 .
- ^ ВВП : 6 тонн ; Сассман Дж.Л., Холбрук С.Р., Ордер Р.В., Генеральный директор Черча, Ким Ш.Х. (август 1978 г.). «Кристаллическая структура дрожжевой РНК-переносчика фенилаланина. I. Кристаллографическое уточнение». Дж. Мол. Биол . 123 (4): 607–30. дои : 10.1016/0022-2836(78)90209-7 . ПМИД 357742 . ; рендерится через PyMOL .
- ^ Jump up to: а б Куигли Дж.Дж., Рич А. (ноябрь 1976 г.). «Структурные домены молекул транспортных РНК». Наука . 194 (4267): 796–806. Бибкод : 1976Sci...194..796Q . дои : 10.1126/science.790568 . ПМИД 790568 .
- ^ «Дуглас Х. Тернер» . Правила Тернера . Химический факультет Рочестерского университета.
- ^ Уолтер А.Э., Тернер Д.Х., Ким Дж., Литтл М.Х., Мюллер П., Мэтьюз Д.Х., Цукер М. (сентябрь 1994 г.). «Коаксиальная укладка спиралей усиливает связывание олигорибонуклеотидов и улучшает предсказания сворачивания РНК» . Учеб. Натл. акад. наук. США . 91 (20): 9218–22. Бибкод : 1994PNAS...91.9218W . дои : 10.1073/pnas.91.20.9218 . ПМК 44783 . ПМИД 7524072 .
- ^ Мерфи Флорида, Ван Ю.Х., Гриффит Дж.Д., Чех Т.Р. (сентябрь 1994 г.). «Коаксиально уложенные спирали РНК в каталитическом центре рибозима Tetrahymena». Наука . 265 (5179): 1709–12. Бибкод : 1994Sci...265.1709M . дои : 10.1126/science.8085157 . ПМИД 8085157 .
- ^ Jump up to: а б с Кейт Дж. Х., Гудинг А. Р., Поделл Э., Чжоу К., Голден Б. Л., Кундрот CE, Чех Т. Р., Дудна Дж. А. (сентябрь 1996 г.). «Кристаллическая структура домена рибозима группы I: принципы упаковки РНК». Наука . 273 (5282): 1678–85. Бибкод : 1996Sci...273.1678C . дои : 10.1126/science.273.5282.1678 . ПМИД 8781224 . S2CID 38185676 .
- ^ Ноллер HF (сентябрь 2005 г.). «Структура РНК: чтение рибосомы». Наука . 309 (5740): 1508–14. Бибкод : 2005Sci...309.1508N . дои : 10.1126/science.1111771 . ПМИД 16141058 . S2CID 16577145 .
- ^ Ли Эй Джей, Кротерс DM (август 1998 г.). «Структура решения комплекса петля-петля РНК: последовательность инвертированной петли ColE1» . Структура . 6 (8): 993–1005. дои : 10.1016/S0969-2126(98)00101-4 . ПМИД 9739090 .
- ^ Ферре-Д'Амаре А.Р., Чжоу К., Дудна Дж.А. (октябрь 1998 г.). «Кристаллическая структура рибозима вируса гепатита дельта». Природа . 395 (6702): 567–74. Бибкод : 1998Natur.395..567F . дои : 10.1038/26912 . ПМИД 9783582 . S2CID 4359811 .
- ^ Ротемунд П.В. (март 2006 г.). «Складывание ДНК для создания наноразмерных форм и узоров» (PDF) . Природа . 440 (7082): 297–302. Бибкод : 2006Natur.440..297R . дои : 10.1038/nature04586 . ПМИД 16541064 . S2CID 4316391 .
- ^ Jump up to: а б Банерджи, Абхинав; Ананд, Микки; Калита, Симанта; Ганджи, Махипал (декабрь 2023 г.). «Одномолекулярный анализ энергетики укладки оснований ДНК с использованием структурированных наноструктур ДНК» . Природные нанотехнологии . 18 (12): 1474–1482. Бибкод : 2023NatNa..18.1474B . дои : 10.1038/s41565-023-01485-1 . ПМЦ 10716042 . ПМИД 37591937 .
- ^ Яковчук, Петр; Протозанова Екатерина; Франк-Каменецкий, Максим Д. (2006). «Вклад укладки оснований и спаривания оснований в термическую стабильность двойной спирали ДНК» . Исследования нуклеиновых кислот . 34 (2): 564–574. дои : 10.1093/nar/gkj454 . ПМЦ 1360284 . ПМИД 16449200 .
- ^ КИЛХЕРР, ФАБИАН; ПРОСЫПАЙСЯ, КРИСТИАН; ФУР, БЕНДЖАМИН; РИФ, МАТТИАС; ЗАХАРИЯ, МАРТИН; ДИТЦ, ХЕНДРИК (2016). «Одномолекулярное рассечение сил укладки в ДНК». Наука . 353 (6304). doi : 10.1126/science.aaf5508 . ПМИД 27609897 .
- ^ Jump up to: а б с д PDB : 1GID ; Кейт Дж. Х., Гудинг А. Р., Поделл Э., Чжоу К., Голден Б. Л., Кундрот CE, Чех Т. Р., Дудна Дж. А. (сентябрь 1996 г.). «Кристаллическая структура домена рибозима группы I: принципы упаковки РНК». Наука . 273 (5282): 1678–85. Бибкод : 1996Sci...273.1678C . дои : 10.1126/science.273.5282.1678 . ПМИД 8781224 . S2CID 38185676 . ; визуализируется с помощью PyMOL
- ^ Накано М., Муди Э.М., Лян Дж., Бевилаква ПК (декабрь 2002 г.). «Отбор термодинамически стабильных тетрапетлей ДНК с использованием гель-электрофореза в градиенте температуры выявляет четыре мотива: d (cGNNAg), d (cGNABg), d (cCNNGg) и d (gCNNGc)». Биохимия . 41 (48): 14281–92. дои : 10.1021/bi026479k . ПМИД 12450393 .
- ^ Мур П.Б. (1999). «Структурные мотивы в РНК». Анну. Преподобный Биохим . 68 (1): 287–300. doi : 10.1146/annurev.biochem.68.1.287 . ПМИД 10872451 .
- ^ Абрамовиц Д.Л., Пайл AM (февраль 1997 г.). «Замечательная морфологическая изменчивость общего мотива сворачивания РНК: взаимодействие тетрапетли GNRA-рецептора» . Дж. Мол. Биол . 266 (3): 493–506. дои : 10.1006/jmbi.1996.0810 . ПМИД 9067606 .
- ^ Муди Э.М., Феррар Дж.К., Бевилаква ПК (июнь 2004 г.). «Доказательства того, что сворачивание шпильки тетрапетли РНК менее кооперативно, чем ее аналог ДНК». Биохимия . 43 (25): 7992–8. дои : 10.1021/bi049350e . ПМИД 15209494 .
- ^ Чжао Ц, Хуан ХК, Нагасвами Ю, Ся Ю, Гао Х, Фокс ГЭ (август 2012 г.). «Тетралупы UNAC: в какой степени они имитируют тетралупы GNRA?». Биополимеры . 97 (8): 617–628. дои : 10.1002/bip.22049 . ПМИД 22605553 .
- ^ Уильямс Д.Х., Гейт М.Дж., Лоукс Д. (2006). Нуклеиновые кислоты в химии и биологии . Кембридж, Великобритания: RSC Pub. ISBN 0-85404-654-2 .
- ^ Jump up to: а б Йегер Л., Мишель Ф., Вестхоф Э. (март 1994 г.). «Участие тетрапетли GNRA в третичных взаимодействиях РНК дальнего действия». Дж. Мол. Биол . 236 (5): 1271–6. дои : 10.1016/0022-2836(94)90055-8 . ПМИД 7510342 .
- ^ Мишель Ф., Вестхоф Э (декабрь 1990 г.). «Моделирование трехмерной архитектуры каталитических интронов группы I на основе сравнительного анализа последовательностей». Дж. Мол. Биол . 216 (3): 585–610. дои : 10.1016/0022-2836(90)90386-Z . ПМИД 2258934 .
- ^ Jump up to: а б с ПДБ : 1ФФК ; Бан Н., Ниссен П., Хансен Дж., Мур П.Б., Стейц Т.А. (август 2000 г.). «Полная атомная структура большой субъединицы рибосомы с разрешением 2,4 А». Наука . 289 (5481): 905–20. Бибкод : 2000Sci...289..905B . дои : 10.1126/science.289.5481.905 . ПМИД 10937989 . ; визуализируется с помощью PyMOL
- ^ Ниссен П., Ипполито Дж.А., Бан Н., Мур П.Б., Стейц Т.А. (апрель 2001 г.). «Третичные взаимодействия РНК в большой субъединице рибосомы: А-минорный мотив» . Учеб. Натл. акад. наук. США . 98 (9): 4899–903. Бибкод : 2001PNAS...98.4899N . дои : 10.1073/pnas.081082398 . ПМК 33135 . ПМИД 11296253 .
- ^ Ёсидзава С., Фурми Д., Пуглиси Дж.Д. (сентябрь 1999 г.). «Распознавание спирали кодон-антикодон рибосомальной РНК». Наука . 285 (5434): 1722–5. дои : 10.1126/science.285.5434.1722 . ПМИД 10481006 .
- ^ Боков К., Штейнберг С.В. (февраль 2009 г.). «Иерархическая модель эволюции 23S рибосомальной РНК». Природа . 457 (7232): 977–80. Бибкод : 2009Natur.457..977B . дои : 10.1038/nature07749 . ПМИД 19225518 . S2CID 4400869 .
- ^ Бэти РТ, Рэмбо Р.П., Дудна Дж.А. (август 1999 г.). «Третичные мотивы в структуре и сворачивании РНК». Энджью. хим. Межд. Эд. англ . 38 (16): 2326–2343. doi : 10.1002/(SICI)1521-3773(19990816)38:16<2326::AID-ANIE2326>3.0.CO;2-3 . ПМИД 10458781 .
- ^ Тамура М., Холбрук С.Р. (июль 2002 г.). «Последовательность и структурная консервативность рибозных застежек РНК» . Дж. Мол. Биол . 320 (3): 455–74. дои : 10.1016/S0022-2836(02)00515-6 . ПМИД 12096903 .
- ^ ВВП : 3IGI ; Тур Н., Китинг К.С., Федорова О., Раджашанкар К., Ван Дж., Пайл А.М. (январь 2010 г.). «Третичная архитектура интрона Oceanobacillus iheyensis группы II» . РНК . 16 (1): 57–69. дои : 10.1261/rna.1844010 . ПМК 2802037 . ПМИД 19952115 . ; визуализируется с использованием PyMOL .
- ^ ПДБ : 1ZZN ; Стейли М.Р., Strobel SA (сентябрь 2005 г.). «Структурные доказательства двухметаллического механизма сплайсинга интронов группы I». Наука . 309 (5740): 1587–90. Бибкод : 2005Sci...309.1587S . дои : 10.1126/science.1114994 . ПМИД 16141079 . S2CID 40099718 . ; визуализируется с помощью PyMOL
- ^ Селандер Д.В., Чехия ТР (январь 1991 г.). «Визуализация сворачивания каталитической молекулы РНК более высокого порядка». Наука . 251 (4992): 401–7. Бибкод : 1991Sci...251..401C . дои : 10.1126/science.1989074 . ПМИД 1989074 .
- ^ Пайл AM (сентябрь 2002 г.). «Ионы металлов в структуре и функциях РНК». Ж. Биол. Неорг. Хим . 7 (7–8): 679–90. дои : 10.1007/s00775-002-0387-6 . ПМИД 12203005 . S2CID 42008484 .
- ^ Морроу-младший, Андолина СМ (2012). «Глава 6. Спектроскопические исследования связывания ионов лантаноидов с нуклеиновыми кислотами». В Sigel A, Sigel H, Sigel RK (ред.). Взаимодействие между ионами металлов и нуклеиновыми кислотами . Ионы металлов в науках о жизни. Том. 10. Спрингер. стр. 171–197. дои : 10.1007/978-94-007-2172-2_6 . ISBN 978-94-007-2171-5 . ПМИД 22210339 .
- ^ Кейт Дж. Х., Дудна Дж. А. (октябрь 1996 г.). «Металлосвязывающие сайты в большой бороздке большого рибозимного домена» . Структура . 4 (10): 1221–9. дои : 10.1016/S0969-2126(96)00129-3 . ПМИД 8939748 .
- ^ Кифт Дж. С., Тиноко I (май 1997 г.). «Структура раствора места связывания металла в большой бороздке РНК в комплексе с гексаммином кобальта (III)» . Структура . 5 (5): 713–21. дои : 10.1016/S0969-2126(97)00225-6 . ПМИД 9195889 .
- ^ Рюдиссер С., Тиноко I (февраль 2000 г.). «Структура раствора гексаммина кобальта (III), образующего комплекс с тетрапетлей GAAA, и связывание ионов металла с несовпадениями G·A». Дж. Мол. Биол . 295 (5): 1211–23. дои : 10.1006/jmbi.1999.3421 . ПМИД 10653698 .
- ^ Буркхардт С., Захариас М. (октябрь 2001 г.). «Моделирование связывания ионов с мотивами платформы AA в РНК: исследование непрерывных растворителей, включая конформационную адаптацию» . Нуклеиновые кислоты Рез . 29 (19): 3910–8. дои : 10.1093/нар/29.19.3910 . ПМК 60250 . ПМИД 11574672 .
- ^ Панютин И.Г., Бисвас И., Се П. (апрель 1995 г.). «Ключевая роль структуры соединения Холлидея в миграции ветвей ДНК» . Журнал ЭМБО . 14 (8): 1819–26. дои : 10.1002/j.1460-2075.1995.tb07170.x . ПМЦ 398275 . ПМИД 7737132 .
- ^ Фу Т.Дж., Симан, Северная Каролина (апрель 1993 г.). «Молекулы двойного кроссовера ДНК». Биохимия . 32 (13): 3211–20. дои : 10.1021/bi00064a003 . ПМИД 8461289 .
- ^ Уотсон Дж. Д., Крик Ф. Х. (апрель 1953 г.). «Молекулярная структура нуклеиновых кислот; структура нуклеиновой кислоты дезоксирибозы» (PDF) . Природа . 171 (4356): 737–738. Бибкод : 1953Natur.171..737W . дои : 10.1038/171737a0 . ПМИД 13054692 . S2CID 4253007 .
- ^ Холли, Р.В., Апгар, Дж., Эверетт, Дж.А., Мэдисон, Дж.Т., Маргисс, М., Меррилл, С.Х., Пенвик, Дж.Р., Замир (март 1965 г.). «Структура рибонуклеиновой кислоты». Наука . 147 (3664): 1462–5. Бибкод : 1965Sci...147.1462H . дои : 10.1126/science.147.3664.1462 . ПМИД 14263761 . S2CID 40989800 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Ким Ш., Куигли Дж., Суддат Флорида, Рич А. (апрель 1971 г.). «Рентгеновские дифрактограммы высокого разрешения кристаллической транспортной РНК, на которых видны спиральные области» . Учеб. Натл. акад. наук. США . 68 (4): 841–5. дои : 10.1073/pnas.68.4.841 . ПМК 389056 . ПМИД 5279525 .
- ^ Шен LX, Цай Z, Тиноко I (август 1995 г.). «Структура РНК в высоком разрешении» . ФАСЕБ Дж . 9 (11): 1023–33. дои : 10.1096/fasebj.9.11.7544309 . ПМИД 7544309 . S2CID 40621440 .
- ^ Чех Т.Р., Зауг А.Я., Грабовский П.Ю. (декабрь 1981 г.). «Сплайсинг in vitro предшественника рибосомальной РНК Tetrahymena: участие гуанозинового нуклеотида в вырезании промежуточной последовательности». Клетка . 27 (3, часть 2): 487–96. дои : 10.1016/0092-8674(81)90390-1 . ПМИД 6101203 . S2CID 17674600 .
- ^ Старк Б.К., Коле Р., Боуман Э.Дж., Альтман С. (август 1978 г.). «Рибонуклеаза P: фермент с важным компонентом РНК» . Учеб. Натл. акад. наук. США . 75 (8): 3717–21. Бибкод : 1978PNAS...75.3717S . дои : 10.1073/pnas.75.8.3717 . ПМЦ 392857 . ПМИД 358197 .
- ^ Проди Г.А., Бакос Дж.Т., Бузаян Дж.М., Шнайдер И.Р., Брюнинг Г. (март 1986 г.). «Автолитический процессинг димерной сателлитной РНК вируса растений». Наука . 231 (4745): 1577–1580. Бибкод : 1986Sci...231.1577P . дои : 10.1126/science.231.4745.1577 . ПМИД 17833317 . S2CID 21563490 .
- ^ Плей Х.В., Флаэрти К.М., Маккей Д.Б. (ноябрь 1994 г.). «Трехмерная структура рибозима «головка молотка». Природа . 372 (6501): 68–74. Бибкод : 1994Natur.372...68P . дои : 10.1038/372068a0 . ПМИД 7969422 . S2CID 4333072 .