Сперматозоид
| Сперматозоид | |
|---|---|
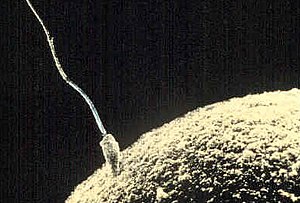 Сперматозоид пытается проникнуть в оболочку яйцеклетки , чтобы оплодотворить ее. | |
 Схема человеческого сперматозоида | |
| Подробности | |
| Идентификаторы | |
| латинский | сперматозоид |
| Греческий | сперма |
| МеШ | D013094 |
| Анатомические термины микроанатомии | |
Сперматозоид æ ( / s p ər ˌ m t ə ˈ z s oʊ . ən , ˌ p ɜːr m ə t ə - / ; [ 1 ] также пишется как сперматозоид ; пл. : сперматозоиды ; от древнегреческого σπέρμα ( spérma ) «семя» и ζῷον ( zôion ) «животное») — сперматозоид , подвижный или подвижная форма гаплоидной клетки , представляющая собой мужскую гамету . Сперматозоид присоединяется к , яйцеклетке образуя зиготу . (Зигота — это отдельная клетка с полным набором хромосом , которая обычно развивается в эмбрион .)
Сперматозоиды передают примерно половину ядерной генетической информации диплоидному митохондриальной потомству (за исключением, в большинстве случаев, ДНК ). У млекопитающих пол потомства определяется сперматозоидом: сперматозоид, несущий Х-хромосому , приведет к потомству женского пола (XX), а сперматозоид, несущий Y-хромосому, приведет к потомству мужского пола (XY). Сперматозоиды были впервые обнаружены в Антони ван Левенгука в 1677 году. лаборатории [ 2 ]
Структура, функции и размер сперматозоидов млекопитающих
[ редактировать ]Люди
[ редактировать ]Человеческий сперматозоид является репродуктивной клеткой у мужчин и выживает только в теплой среде; как только он покидает мужской организм, вероятность выживания спермы снижается, и она может погибнуть, тем самым снижая общее качество спермы . Сперматозоиды бывают двух типов: «женские» и «мужские». Сперматозоиды, дающие после оплодотворения потомство женского пола (XX), отличаются тем, что несут Х-хромосому, тогда как сперматозоиды, дающие потомство мужского пола (XY), несут Y-хромосому. [ 3 ]
Сперматозоид человека состоит из плоской дискообразной головки размером 5,1 на 3,1 мкм и хвоста, известного как жгутик, длиной 50 мкм. [ 4 ] Жгутик приводит в движение сперматозоид (у человека со скоростью около 1–3 мм/мин), образуя эллиптический конус. [ 5 ] Сперматозоиды имеют обонятельный механизм направления и после достижения маточных труб должны пройти период капацитации перед проникновением в яйцеклетку. [ 6 ]
Голова: имеет компактное ядро, состоящее только из хроматического вещества, и окружено лишь тонким краем цитоплазмы. Над ядром лежит колпачковая структура, называемая акросомой , образованная в результате модификации тельца Гольджи, который секретирует фермент спермлизин ( гиалуронидазу, корона-проникающий фермент, зона лизин или акрозин ), необходимый для оплодотворения. Когда сперматозоид приближается к яйцеклетке, он подвергается акросомной реакции , при которой мембрана, окружающая акросому, сливается с плазматической мембраной головки спермия, обнажая содержимое акросомы. [ 7 ]
Шея: это самая маленькая часть (0,03 мкм), имеющая проксимальную центриоль , параллельную основанию ядра, и дистальную центриоль, перпендикулярную предыдущей. Проксимальная центриоль присутствует и в зрелом сперматозоиде; дистальная центриоль исчезает после сборки аксонемы. Проксимальная центриоль входит в яйцеклетку во время оплодотворения и начинает первое деление яйцеклетки, не имеющей центриоли. Дистальная центриоль дает начало осевой нити, которая образует хвост и имеет расположение (9+2). переходная мембрана, называемая манжетой В средней части находится .
Средняя часть: имеет 10–14 спиралей митохондрий, окружающих осевую нить в цитоплазме. Он обеспечивает подвижность и поэтому называется электростанцией спермы. Он также имеет кольцевую центриоль (кольцо), которая образует диффузионный барьер между средней частью и основной частью и служит стабилизирующей структурой для жесткости хвоста. [ 8 ]
Хвост: Жгутик - самая длинная часть (50 мкм), имеющая осевую нить, окруженную цитоплазмой и плазматической мембраной, но на заднем конце осевая нить обнажена. Жгутик обеспечивает движение клетке.
Сперма имеет щелочную природу, и сперматозоиды не достигают полной подвижности (гиперподвижности), пока не достигнут влагалища , где щелочной pH нейтрализуется кислыми вагинальными жидкостями. Этот постепенный процесс занимает 20–30 минут. В этот период фибриноген из семенных пузырьков образует сгусток, закрепляющий и защищающий сперму. Как только они становятся гиперподвижными, фибринолизин из предстательной железы растворяет сгусток, позволяя сперматозоидам оптимально развиваться.
Сперматозоид характеризуется минимумом цитоплазмы и наиболее плотно упакованной ДНК, известной у эукариот . По сравнению с митотическими хромосомами в соматических клетках ДНК сперматозоидов как минимум в шесть раз более конденсирована. [ 9 ]
Образец вносит свой вклад в ДНК / хроматин , центриоль и, возможно, также фактор активации ооцитов (OAF). [ 10 ] Он также может вносить вклад в отцовскую информационную РНК (мРНК), что также способствует эмбриональному развитию. [ 10 ]
-
 Электронная микрофотография сперматозоидов человека, увеличенная в 3140 раз.
Электронная микрофотография сперматозоидов человека, увеличенная в 3140 раз. -
 Сперматозоиды в образце мочи мужчины 45 лет, находящегося под наблюдением с диагнозом доброкачественная гиперплазия предстательной железы .
Сперматозоиды в образце мочи мужчины 45 лет, находящегося под наблюдением с диагнозом доброкачественная гиперплазия предстательной железы . -
 Размеры головки сперматозоида человека, измеренные у 39-летнего здорового субъекта.
Размеры головки сперматозоида человека, измеренные у 39-летнего здорового субъекта.



Сперматозоид человека содержит не менее 7500 различных белков . [ 11 ]
генетика спермы человека связана с эволюцией человека . Согласно исследованию 2020 года, [ 12 ] [ 13 ] [ 14 ]
Повреждение и восстановление ДНК
[ редактировать ]Повреждения ДНК, присутствующие в сперматозоидах в период после мейоза , но до оплодотворения , могут быть устранены в оплодотворенной яйцеклетке, но если их не устранить, они могут иметь серьезные пагубные последствия для фертильности и развивающегося эмбриона. Человеческие сперматозоиды особенно уязвимы к атаке свободных радикалов и окислительному повреждению ДНК. [ 15 ] [ 16 ] (см., например, 8-оксо-2'-дезоксигуанозин )
Воздействие на мужчин определенного образа жизни, окружающей среды или профессиональных опасностей может увеличить риск образования анеуплоидных сперматозоидов. [ 17 ] В частности, риск анеуплоидии увеличивается при курении табака. [ 18 ] [ 19 ] и профессиональное воздействие бензола, [ 20 ] инсектициды, [ 21 ] [ 22 ] и перфторированные соединения. [ 23 ] Повышенная анеуплоидия сперматозоидов часто возникает в связи с повышенным повреждением ДНК. Фрагментация ДНК и повышенная восприимчивость ДНК к денатурации in situ, признаки, аналогичные тем, которые наблюдаются при апоптозе соматических клеток, характеризуют аномальные сперматозоиды в случаях мужского бесплодия . [ 24 ] [ 25 ]
Уклонение от реакции иммунной системы
[ редактировать ]Молекулы гликопротеина на поверхности эякулированных сперматозоидов распознаются всеми женскими иммунными системами человека и интерпретируются как сигнал о том, что клетка не должна быть отторгнута. В противном случае женская иммунная система могла бы атаковать сперму в репродуктивном тракте . Специфические гликопротеины, покрывающие сперматозоиды, также используются некоторыми раковыми и бактериальными клетками, некоторыми паразитическими червями и ВИЧ-инфицированными лейкоцитами, тем самым избегая иммунного ответа со стороны организма-хозяина . [ 26 ]
Гемато -тестикулярный барьер , поддерживаемый плотными соединениями между клетками Сертоли семенных канальцев, предотвращает сообщение между формирующимися сперматозоидами в семенниках и кровеносными сосудами (и циркулирующими в них иммунными клетками) внутри интерстициального пространства . Это не позволяет им вызвать иммунный ответ. Гематотестикулярный барьер также важен для предотвращения нарушения сперматогенеза токсичными веществами. [ нужна ссылка ]
Сперматозоиды в других организмах
[ редактировать ]
Животные
[ редактировать ]У большинства животных, имеющих половое размножение, оплодотворение происходит за счет сперматозоидов.
Некоторые виды плодовых мух производят самые крупные из известных в природе сперматозоидов. [ 27 ] [ 28 ] Drosophila melanogaster производит сперматозоиды размером до 1,8 мм. [ 29 ] в то время как его родственник Drosophila bifurca производит самый крупный из известных сперматозоидов длиной более 58 мм. [ 27 ] У Drosophila melanogaster весь сперматозоид, включая хвост, включается в ооцита цитоплазму , однако у Drosophila bifurca в ооцит проникает лишь небольшая часть хвоста. [ 30 ]
Древесная мышь Apodemus sylvaticus обладает сперматозоидами серповидной морфологии. Еще одной характеристикой, которая делает эти гаметоциты уникальными, является наличие апикального крючка на головке спермия. Этот крючок используется для прикрепления к крючкам или жгутикам других сперматозоидов. Эти навесные устройства вызывают агрегацию, в результате чего образуются мобильные поезда. Эти поезда обеспечивают улучшение моторики женских репродуктивных путей и являются средством, способствующим оплодотворению. [ 31 ]
Постмейотическая фаза сперматогенеза мышей очень чувствительна к генотоксичным агентам окружающей среды, поскольку по мере того, как мужские половые клетки образуют зрелые сперматозоиды, они постепенно теряют способность восстанавливать повреждения ДНК. [ 32 ] Облучение мышей-самцов во время позднего сперматогенеза может вызвать повреждение, которое сохраняется в течение как минимум 7 дней в оплодотворяющих сперматозоидах, а нарушение путей восстановления двухцепочечных разрывов материнской ДНК увеличивает хромосомные аберрации, происходящие из сперматозоидов. [ 33 ] Лечение самцов мышей мелфаланом , бифункциональным алкилирующим агентом , часто используемым в химиотерапии, вызывает повреждения ДНК во время мейоза, которые могут сохраняться в невосстановленном состоянии по мере того, как зародышевые клетки проходят через компетентные к репарации ДНК фазы сперматогенного развития. [ 34 ] Такие невосстановленные повреждения ДНК в сперматозоидах после оплодотворения могут привести к появлению потомства с различными аномалиями.
Морские ежи, такие как Arbacia punctulata, являются идеальными организмами для исследования спермы. Они выбрасывают в море большое количество сперматозоидов, что делает их хорошо подходящими в качестве модельных организмов для экспериментов. [ 35 ]
Сперматозоиды сумчатых обычно длиннее, чем у плацентарных млекопитающих . [ 36 ]
Растения, водоросли и грибы
[ редактировать ]Гаметофиты покрытосеменных мохообразных голосеменных , папоротников и некоторых растений производят подвижные сперматозоиды , в отличие от пыльцевых зерен, используемых у большинства голосеменных и всех растений . Это делает невозможным половое размножение при отсутствии воды , поскольку вода является необходимой средой для встречи сперматозоида и яйцеклетки. Сперматозоиды водорослей и низших растений часто имеют множество жгутиков (см. изображение) и поэтому морфологически отличаются от сперматозоидов животных. [ 37 ]
Некоторые водоросли и грибы производят неподвижные сперматозоиды, называемые сперматиями. У высших растений, а также у некоторых водорослей и грибов оплодотворение включает миграцию ядра спермия через оплодотворительную трубку (например, пыльцевую трубку у высших растений) для достижения яйцеклетки. [ нужна ссылка ]
Производство сперматозоидов у млекопитающих
[ редактировать ]Сперматозоиды производятся в семенных канальцах яичек называемом в процессе, сперматогенезом . Круглые клетки, называемые сперматогониями, делятся и дифференцируются, в конечном итоге превращаясь в сперматозоиды. Во время совокупления клоака внутри или влагалище оплодотворяются , а затем сперматозоиды перемещаются посредством хемотаксиса к яйцеклетке яйцевода .
В вспомогательных репродуктивных технологиях нормозооспермией называют общее количество эякулированных >39 млн , >32% с прогрессивной моторикой и >4% нормальной морфологии. Также нормальная эякуляция у человека должна иметь объем более 1,5 мл, а избыточный объем – 6 мл на одно семяизвержение ( гиперспермия ). Недостаточный объем называется гипоспермией . Эти проблемы связаны с рядом осложнений при производстве сперматозоидов, например:
- Гиперспермия: обычно провоцируется воспалением простаты .
- Гипоспермия: неполная эякуляция, обычно связанная с дефицитом андрогенов ( гипоандрогенией ) или обструкцией какой-либо части семявыбрасывающего протока . В лабораторных условиях это происходит также из-за частичной потери образца.
- Аспермия : нет эякуляции. Это могло произойти из-за ретроградной эякуляции , анатомических или неврологических заболеваний или приема антигипертензивных препаратов.
Активация сперматозоидов
[ редактировать ]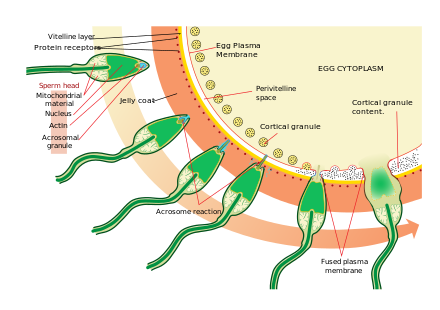
Приближение к яйцеклетке — это довольно сложный, многоэтапный процесс хемотаксиса, управляемый различными химическими веществами/стимулами на отдельных уровнях филогении. Одним из наиболее значимых, общих сигнальных признаков этого события является то, что в поверхностная мембрана даже в случае человеческих сперматозоидов. [ 38 ] Сперматозоиды млекопитающих становятся еще более активными, когда они приближаются к яйцеклетке в процессе, называемом активацией спермы . Было показано, что активация спермы вызвана кальция ионофорами in vitro , прогестероном, высвобождаемым близлежащими кумулюсными клетками , и связыванием с ZP3 зоны пеллюцида . Клетки кумулюса погружены в гелеобразное вещество, состоящее в основном из гиалуроновой кислоты , развиваются в яичнике вместе с яйцеклеткой и поддерживают ее по мере ее роста.
Первоначальное изменение называется «гиперактивацией», которое вызывает изменение подвижности сперматозоидов. Они плавают быстрее, а движения их хвоста становятся более резкими и беспорядочными.
Недавнее открытие связывает гиперактивацию с внезапным притоком ионов кальция в хвосты. Кнутообразный хвост (жгутик) сперматозоида усеян ионными каналами, образованными белками CatSper . Эти каналы являются селективными, пропуская только ионы кальция. Открытие каналов CatSper отвечает за приток кальция. Внезапное повышение уровня кальция приводит к тому, что жгутик изгибается глубже, что приводит к более сильному продвижению сперматозоидов через вязкую среду. Гиперактивность спермы необходима для преодоления двух физических барьеров, защищающих яйцеклетку от оплодотворения.
Вторым процессом активации сперматозоидов является акросомная реакция . Это включает в себя высвобождение содержимого акросомы, которое рассеивается, и воздействие ферментов, прикрепленных к внутренней акросомальной мембране сперматозоида. Это происходит после того, как сперматозоид впервые встречается с яйцеклеткой. Этот механизм типа «замок и ключ» является видоспецифичным и предотвращает слияние сперматозоидов и яйцеклеток разных видов. Есть некоторые свидетельства того, что именно это связывание заставляет акросому высвобождать ферменты, которые позволяют сперматозоиду сливаться с яйцеклеткой.
ZP3, один из белков, составляющих пеллюцидную зону, затем связывается с молекулой-партнером сперматозоида. Ферменты внутренней акросомальной мембраны переваривают пеллюцидную зону. После того, как сперматозоид проникает в зону пеллюцида, часть клеточной мембраны спермия сливается с мембраной яйцеклетки, и содержимое головки диффундирует в яйцеклетку.
Говорят, что после проникновения яйцеклетка активируется . Он подвергается вторичному мейотическому делению, и два гаплоидных ядра (отцовское и материнское) сливаются, образуя зиготу . Чтобы предотвратить полиспермию и свести к минимуму возможность образования триплоидной зиготы, некоторые изменения в зоне пеллюцида яйцеклетки делают ее непроницаемой вскоре после того, как первый сперматозоид попадает в яйцеклетку.
Искусственное хранилище
[ редактировать ]Сперматозоиды можно хранить в разбавителях, таких как разбавитель переменной температуры Иллини (IVT), который, как сообщается, способен сохранять высокую фертильность сперматозоидов в течение более семи дней. [ 39 ] Разбавитель IVT состоит из нескольких солей, сахаров и антибактериальных веществ и насыщен CO 2 . [ 39 ]
Криоконсервацию спермы можно использовать для гораздо более длительного хранения. Самый длительный успешный срок хранения человеческих сперматозоидов при использовании этого метода составляет 21 год. [ 40 ]
ММП и емкость
[ редактировать ]Капацитация — заключительная фаза развития сперматозоидов, когда они приобретают способность оплодотворять яйцеклетку. In vivo это происходит во время эякуляции, когда сперматозоиды покидают влагалище и попадают в верхние репродуктивные пути женщины. In vitro это происходит, когда сперматозоиды промываются и очищаются. Почти 30-40% бесплодия обусловлено мужским фактором, поэтому было разработано несколько стратегий для восстановления функциональных сперматозоидов. Показатель MMP (миллион подвижных прогрессивных клеток на миллилитр) является синонимом капацитации и является очень полезным параметром для принятия решения, наряду со спермиограммой , о типе необходимого лечения. он представляет собой соотношение между процентом прогрессивно подвижных сперматозоидов, полученных в капацитированном состоянии, и процентом прогрессивно подвижных сперматозоидов, полученных в эякулированном состоянии. Он основан на проценте восстановления. В зависимости от процентного содержания мы решаем качество восстановления подвижных сперматозоидов: от 15 до 25 миллионов сперматозоидов на мл считается оптимальным, от 5 до 15 миллионов считается достаточным, а менее 5 миллионов считается неоптимальным или недостаточным. Что касается полученных нами значений, а также результатов спермиограммы, будут отображаться различные методы.
Например, если более 1,0×10 6 будут обнаружены прогрессивные подвижные сперматозоиды на миллилитр, будет рекомендовано вступить в половой акт, а если это не удастся, следующим шагом будет внутриматочная инсеминация , а затем обычное экстракорпоральное оплодотворение .
Менее 1,0×10 6 прогрессивные подвижные сперматозоиды на миллилитр, мы выполним интрацитоплазматическую инъекцию сперматозоидов . В случае азооспермии (отсутствие сперматозоидов в эякуляте) мы проведем биопсию яичек, чтобы проверить, есть ли в яичках сперматозоиды или сперматозоиды не производятся.
История
[ редактировать ]- В 1677 году микробиолог Антони ван Левенгук открыл сперматозоиды.
- В 1841 году швейцарский анатом Альберт фон Кёлликер написал о сперматозоидах в своей работе «Исследования важности сперматозоидов».
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ «сперматозоид» . Dictionary.com Полный (онлайн). nd
- ^ «Хронология: вспомогательная репродукция и контроль над рождаемостью» . Новости ЦБК . Архивировано из оригинала 28 апреля 2016 г. Проверено 6 апреля 2006 г.
- ^ Шейнфельд, Амрам (1939). Ты и наследственность . Нью-Йорк: Компания Фредерика А. Стокса. п. 39.
- ^ Смит, диджей; Гаффни, Э.А.; Блейк-младший; Киркман-Браун, Дж. К. (25 февраля 2009 г.). «Накопление человеческой спермы возле поверхностей: моделирование» (PDF) . Журнал механики жидкости . 621 : 289–320. Бибкод : 2009JFM...621..289S . дои : 10.1017/S0022112008004953 . S2CID 3942426 . Архивировано (PDF) из оригинала 27 января 2022 года . Проверено 10 сентября 2021 г.
- ^ Исиджима, Сумио; Ошио, Сигэру; Мори, Хидео (1986). « Жгутиковое движение сперматозоидов человека ». Исследование гамет . 13 (3): 185–197. дои : 10.1002/mrd.1120130302 .
- ^ Айзенбах, Майкл; Джохалас, Лаура К. (апрель 2006 г.). «Наведение спермы у млекопитающих — грунтовая дорога к яйцеклетке». Nature Reviews Молекулярно-клеточная биология . 7 (4): 276–285. дои : 10.1038/nrm1893 . hdl : 11336/57585 . ПМИД 16607290 . S2CID 32567894 .
- ^ дель Рио, Мария Хосе; Годой, Ана; Торо, Алехандра; Орельяна, Ренан; Кортес, Мануэль Э.; Морено, Рикардо Д.; Виджил, Пилар (октябрь 2007 г.). «Акросомальная реакция спермы: последние достижения». Международный журнал андрологии . 5 (4): 368–373. дои : 10.1016/S1698-031X(07)74086-4 .
- ^ «кольцо спермы | SGD» . www.yeastgenome.org . Архивировано из оригинала 22 февраля 2019 г. Проверено 22 февраля 2019 г.
- ^ Уорд В.С., Коффи Д.С. (1991). «Упаковка и организация ДНК в сперматозоидах млекопитающих: сравнение с соматическими клетками» . Биология размножения . 44 (4): 569–74. дои : 10.1095/biolreprod44.4.569 . ПМИД 2043729 .
- ^ Jump up to: а б Баррозу, Херардо; Вальдеспин, Карлос; Вега, Ева; Кершенович, Рубенс; Авила, Розаура; Авенданьо, Конрадо; Энингер, Серджио (сентябрь 2009 г.). «Вклад спермы в развитие: оплодотворение и не только» . Фертильность и бесплодие . 92 (3): 835–848. doi : 10.1016/j.fertnstert.2009.06.030 . ПМИД 19631936 .
- ^ Амарал, Александра; Кастильо, Юдит; Рамалью-Сантос, Жуан; Олива, Рафаэль (1 января 2014 г.). «Комбинированный протеом спермы человека: клеточные пути и значение для фундаментальной и клинической науки» . Обновление репродукции человека . 20 (1): 40–62. дои : 10.1093/humupd/dmt046 . ПМИД 24082039 .
- ^ Ся, Бо; Ян, Юн; Барон, Смерть; Вагнер, Флориан; Баркли, Далия; Хиодин, Марта; Ким, Санг Ю.; Киф, Дэвид Л.; Алукал, Джозеф П.; Буке, Джефф Д.; Янаи, Итай (январь 2020 г.). «Широко распространенное транскрипционное сканирование в семенниках модулирует скорость эволюции генов» . Ячейка 180 (2): 248–262.e21. дои : 10.1016/j.cell.2019.12.015 . ПМЦ 7891839 . ПМИД 31978344 .
- ^ «Система сканирования спермы может контролировать скорость эволюции человека» . Архивировано из оригинала 28 января 2020 г. Проверено 24 января 2020 г.
- ^ «Система генетического сканирования спермы может контролировать скорость эволюции человека» . Архивировано из оригинала 11 августа 2020 г. Проверено 24 января 2020 г.
- ^ Гаврилюк, Дэн; Эйткен, Роберт Джон (2015). «Повреждение ДНК спермы, опосредованное активными формами кислорода: его влияние на репродукцию человека и траекторию здоровья потомства». Мужская роль в потере беременности и неудачной имплантации эмбриона . Достижения экспериментальной медицины и биологии. Том. 868. стр. 23–47. дои : 10.1007/978-3-319-18881-2_2 . ISBN 978-3-319-18880-5 . ПМИД 26178844 .
- ^ Лозано, генеральный менеджер; Бехарано, И.; Эспино, Дж.; Гонсалес, Д.; Ортис, А.; Гарсия, Дж. Ф.; Родригес, AB; Париенте, JA (2009). «Капаситация в градиенте плотности является наиболее подходящим методом для улучшения оплодотворения и уменьшения фрагментации ДНК сперматозоидов у бесплодных мужчин» . Анатолийский журнал акушерства и гинекологии . 3 (1): 1–7. Архивировано из оригинала 30 апреля 2022 г. Проверено 8 марта 2016 г.
- ^ Темпландо С, Уроз Л, Эстоп А (2013). «Новые сведения о происхождении и значении анеуплоидии в сперматозоидах человека». Молекулярная репродукция человека . 19 (10): 634–43. дои : 10.1093/моль/gat039 . ПМИД 23720770 .
- ^ Ши К., Ко Э., Барклай Л., Хоанг Т., Радемейкер А., Мартин Р. (2001). «Курение сигарет и анеуплоидия человеческой спермы». Молекулярное воспроизводство и развитие . 59 (4): 417–21. дои : 10.1002/mrd.1048 . ПМИД 11468778 . S2CID 35230655 .
- ^ Рубс Дж., Лоу Х., Мур Д., Перро С., Слотт В., Эвенсон Д., Селеван С.Г., Выробек А.Дж. (1998). «Курение сигарет связано с увеличением дисомии сперматозоидов у мужчин-подростков» . Фертильность и бесплодие . 70 (4): 715–23. дои : 10.1016/S0015-0282(98)00261-1 . ПМИД 9797104 .
- ^ Син С., Маркетти Ф., Ли Г., Уэлдон Р.Х., Куртович Э., Янг С., Шмид Т.Е., Чжан Л., Раппапорт С., Вайдьянатха С., Выробек А.Дж., Эскенази Б (2010). «Воздействие бензола, близкое к допустимому пределу в США, связано с анеуплоидией сперматозоидов» . Перспективы гигиены окружающей среды . 118 (6): 833–9. дои : 10.1289/ehp.0901531 . ПМЦ 2898861 . ПМИД 20418200 .
- ^ Ся Ю, Бянь Ц, Сюй Л, Ченг С, Сун Л, Лю Дж, Ву В, Ван С, Ван Икс (2004). «Генотоксическое воздействие на сперматозоиды человека среди рабочих пестицидных заводов, подвергшихся воздействию фенвалерата». Токсикология . 203 (1–3): 49–60. Бибкод : 2004Toxgy.203...49X . дои : 10.1016/j.tox.2004.05.018 . ПМИД 15363581 . S2CID 36073841 .
- ^ Ся Ю, Ченг С, Бянь Ц, Сюй Л, Коллинз МД, Чанг ХК, Сонг Л, Лю Дж, Ван С, Ван Х (2005). «Генотоксическое воздействие на сперматозоиды рабочих, подвергшихся воздействию карбарила» . Токсикологические науки . 85 (1): 615–23. дои : 10.1093/toxsci/kfi066 . ПМИД 15615886 .
- ^ Говернини Л., Герранти С., Де Лео В., Боски Л., Лудди А., Гори М., Орвието Р., Пьомбони П. (2014). «Хромосомные анеуплоидии и фрагментация ДНК сперматозоидов человека от пациентов, подвергшихся воздействию перфторированных соединений» . Андрология . 47 (9): 1012–9. дои : 10.1111/and.12371 . hdl : 11365/982323 . ПМИД 25382683 . S2CID 13484513 .
- ^ Горчица, Ж; Траганос, Ф; Есионовска, Х; Дажинкевич, З. (1993). «Наличие разрывов нитей ДНК и повышенная чувствительность ДНК in situ к денатурации в аномальных сперматозоидах человека. Аналогия с апоптозом соматических клеток». Exp Cell Res . 207 (1): 202–205. дои : 10.1006/excr.1993.1182 . ПМИД 8391465 .
- ^ Эвенсон, ДП; Дажинкевич, З; Меламед, MR (1980). «Связь гетерогенности хроматина сперматозоидов млекопитающих с фертильностью». Наука . 210 (4474): 1131–1133. Бибкод : 1980Sci...210.1131E . дои : 10.1126/science.7444440 . ПМИД 7444440 .
- ^ «Сперматозоиды являются ключом к «иммунитету к болезням» » . Новости Би-би-си . 17 декабря 2007 г. Архивировано из оригинала 4 ноября 2013 г. Проверено 3 ноября 2013 г.
- ^ Jump up to: а б Питник, С; Спайсер, Дж.С.; Марков, Т.А. (11 мая 1995 г.). «Какова длина гигантской спермы?» . Природа . 375 (6527): 109. Бибкод : 1995Natur.375Q.109P . дои : 10.1038/375109a0 . ПМИД 7753164 . S2CID 4368953 .
- ^ Питник, С; Марков, Т.А. (27 сентября 1994 г.). «Преимущества крупных самцов связаны с затратами на производство спермы у Drosophila hydei, вида с гигантскими сперматозоидами» . Труды Национальной академии наук Соединенных Штатов Америки . 91 (20): 9277–81. Бибкод : 1994PNAS...91.9277P . дои : 10.1073/pnas.91.20.9277 . ПМК 44795 . ПМИД 7937755 .
- ^ Купер, К.В. (1950). Демерец, М. (ред.). Биология дрозофилы . Нью-Йорк: Уайли. стр. 1–61.
- ^ Питник, С.; Спайсер, Дж.С.; Марков, Т.А. (1995). «Какова длина гигантской спермы» . Природа . 375 (6527): 109. Бибкод : 1995Natur.375Q.109P . дои : 10.1038/375109a0 . ПМИД 7753164 . S2CID 4368953 .
- ^ Мур, Х; Дворакова, К; Дженкинс, Н; Брид, В. (2002). «Исключительное сотрудничество сперматозоидов у Wood Mouse» (PDF) . Природа . 418 (6894): 174–177. Бибкод : 2002Natur.418..174M . дои : 10.1038/nature00832 . ПМИД 12110888 . S2CID 4413444 .
- ^ Маркетти Ф., Выробек А.Дж. (2008). «Снижение восстановления ДНК во время спермиогенеза у мышей приводит к накоплению наследственных повреждений ДНК» (PDF) . Восстановление ДНК . 7 (4): 572–81. дои : 10.1016/j.dnarep.2007.12.011 . ПМИД 18282746 . S2CID 1316244 . Архивировано (PDF) из оригинала 25 июля 2018 г. Проверено 12 мая 2024 г.
- ^ Маркетти Ф., Эссерс Дж., Канаар Р., Выробек А.Дж. (2007). «Нарушение восстановления материнской ДНК увеличивает хромосомные аберрации, происходящие из сперматозоидов» . Труды Национальной академии наук Соединенных Штатов Америки . 104 (45): 17725–9. Бибкод : 2007PNAS..10417725M . дои : 10.1073/pnas.0705257104 . ПМК 2077046 . ПМИД 17978187 .
- ^ Маркетти Ф., Бишоп Дж., Джинджерич Дж., Выробек А.Дж. (2015). «Повреждения мейотической межцепочечной ДНК избегают отцовской репарации и вызывают хромосомные аберрации в зиготе из-за материнской неправильной репарации» . Научные отчеты . 5 : 7689. Бибкод : 2015NatSR...5E7689M . дои : 10.1038/srep07689 . ПМЦ 4286742 . ПМИД 25567288 .
- ^ Вакье, Виктор Д. (август 2011 г.). «Лаборатория по оплодотворению морских ежей». Молекулярное воспроизводство и развитие . 78 (8): 553–564. дои : 10.1002/mrd.21360 . ПМИД 21805525 . S2CID 13452188 .
- ^ Ларри Фогельнест; Тимоти Портас (1 мая 2019 г.). Современная терапия в медицине австралийских млекопитающих . Издательство Csiro. ISBN 978-1-4863-0752-4 .
- ^ Уайт-Купер, Хелен; Баусек, Нина (27 мая 2010 г.). «Эволюция и сперматогенез» . Философские труды Королевского общества B: Биологические науки . 365 (1546): 1465–1480. дои : 10.1098/rstb.2009.0323 . ISSN 0962-8436 . ПМК 2871925 . ПМИД 20403864 .
- ^ Гнесси Л., Фаббри А., Сильвестрони Л., Моретти С., Фрайоли Ф., Перт С.Б., Исидори А. (1986). «Доказательства наличия специфических рецепторов N-формиловых хемотаксических пептидов на сперматозоидах человека». Журнал клинической эндокринологии и метаболизма . 63 (4): 841–6. doi : 10.1210/jcem-63-4-841 . ПМИД 3018025 .
- ^ Jump up to: а б Уотсон, П.Ф. (1993). «Потенциальное влияние технологии инкапсуляции спермы на важность выбора времени искусственного оплодотворения: перспектива в свете опубликованных работ». Воспроизводство, рождаемость и развитие . 5 (6): 691–9. дои : 10.1071/RD9930691 . ПМИД 9627729 .
- ^ НОВОСТИ и пресс-релизы Planer > Ребенок, рожденный после 21 года хранения спермы в морозильной камере с регулируемой скоростью Planer. Архивировано 3 марта 2018 г. в Wayback Machine 14.10.2004.