Красный импортный огненный муравей
| Красный импортный огненный муравей | |
|---|---|

| |
| Группа огненных муравьев | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Членистоногие |
| Сорт: | Насекомое |
| Заказ: | Перепончатокрылые |
| Семья: | Формициды |
| Подсемейство: | Мирмицины |
| Род: | Соленопсис |
| Разновидность: | Святой непобедимый
|
| Биномиальное имя | |
| Соленопсис инвикта Бурен, 1972 г.
| |
| Синонимы [1] | |
| |
Solenopsis invicta , огненный муравей , или красный импортированный огненный муравей ( RIFA ), — вид муравьев , обитающий в Южной Америке . Член рода Solenopsis подсемейства Myrmicinae , он был описан швейцарским энтомологом Феликсом Санчи как вариант S. saevissima в 1916 году. Его нынешнее видовое название invicta было дано муравью в 1972 году как отдельный вид. Однако вариант и вид были одним и тем же муравьем, а название сохранилось благодаря его широкому использованию. Несмотря на южноамериканское происхождение, красный импортированный огненный муравей был случайно завезён в Австралию , Новую Зеландию , несколько Азии и Карибского бассейна стран , Европу и США . Красный импортированный огненный муравей полиморфен , так как рабочие появляются в разных формах и размерах. Цвет муравья красный и несколько желтоватый с коричневым или черным брюшком , но самцы полностью черные. Красные завезенные огненные муравьи доминируют на измененных территориях и обитают в самых разных средах обитания. Их можно найти в тропических лесах , нарушенных территориях, пустынях , лугах. , вдоль дорог и зданий , а также в электрооборудовании. Колонии образуют большие насыпи, построенные из почвы, без видимых входов, поскольку строятся туннели для корма, а рабочие выходят далеко от гнезда.
Эти муравьи демонстрируют самые разнообразные модели поведения, например, строят плоты, когда чувствуют, что уровень воды повышается. Они также демонстрируют некрофорное поведение , когда товарищи по гнезду выбрасывают отходы или мертвых муравьев на кучи мусора за пределами гнезда. Собирание пищи происходит в теплые или жаркие дни, хотя ночью они могут оставаться на улице. Рабочие общаются с помощью ряда семиохимических веществ и феромонов , которые используются для вербовки, добывания пищи и защиты. Они всеядны и едят мертвых млекопитающих , членистоногих , насекомых , семена и сладкие вещества, такие как медвяная роса полужесткокрылых насекомых , с которыми у них сложились отношения . Хищники включают паукообразных , птиц и многих насекомых, включая других муравьев , стрекоз , уховерток и жуков . Муравей является хозяином паразитов и ряда патогенов , нематод и вирусов , которые рассматриваются как потенциальные агенты биологической борьбы. Брачный полет происходит в теплое время года, и крылатые особи могут спариваться на срок до 30 минут. Основание колонии может быть осуществлено одной королевой или группой королев, которые позже будут бороться за доминирование, как только появятся первые рабочие. Рабочие особи могут жить несколько месяцев, а королевы – годы; Численность колоний может варьироваться от 100 000 до 250 000 особей. У красного импортированного огненного муравья существуют две формы общества: полигинные колонии (гнезда с несколькими матками) и моногинные колонии (гнезда с одной маткой).
Яд играет важную роль в жизни муравьев, поскольку он используется для захвата добычи или защиты. [2] Около 95% яда состоит из водонерастворимых пиперидиновых алкалоидов, известных как соленопсины , а остальная часть состоит из смеси токсичных белков, которые могут быть особенно эффективны для чувствительных людей; Название «огненный муравей» происходит от ощущения жжения, вызываемого их укусом. [3] ими жалят более 14 миллионов человек Ежегодно в Соединенных Штатах , где у многих, как ожидается, разовьется аллергия на яд. У большинства пострадавших наблюдается сильное жжение и отек с последующим образованием стерильных пустул , которые могут сохраняться в течение нескольких дней. Однако от 0,6% до 6,0% людей могут страдать от анафилаксии , которая может привести к летальному исходу, если ее не лечить. Общие симптомы включают головокружение , боль в груди , тошноту , сильное потоотделение , низкое кровяное давление, одышку и невнятную речь . В результате нападений завезенных красных огненных муравьев зарегистрировано более 80 смертей. Лечение зависит от симптомов; тем, кто испытывает только боль и образование пустул, медицинская помощь не требуется, но тем, кто страдает анафилаксией, дают адреналин . Иммунотерапия экстрактами всего тела используется для лечения жертв и считается высокоэффективной. [4]
Муравей считается печально известным вредителем, ежегодно наносящим миллиарды долларов ущерб на и наносящим ущерб дикой природе. Муравьи процветают в городских районах, поэтому их присутствие может препятствовать выходу на улицу. Гнезда можно строить под такими конструкциями, как тротуары и фундаменты, что может вызвать структурные проблемы или привести к их обрушению. Они не только могут повредить или разрушить постройки, но и завезенные красные огненные муравьи также могут повредить оборудование и инфраструктуру и повлиять на стоимость бизнеса, земли и собственности. В сельском хозяйстве они могут повредить посевы и технику, а также угрожать пастбищам. Известно, что они поражают самые разнообразные сельскохозяйственные культуры, а насыпи, построенные на сельскохозяйственных угодьях, могут помешать сбору урожая. Они также представляют угрозу для животных и домашнего скота, способные нанести им серьезные травмы или убить, особенно молодых, слабых или больных животных. Несмотря на это, они могут быть полезны, поскольку поедают распространенных насекомых-вредителей сельскохозяйственных культур. Общие методы борьбы с этими муравьями включают травлю и окуривание; другие методы могут быть неэффективными или опасными. Благодаря своей известности и важности муравей стал одним из наиболее изученных насекомых на планете, соперничая даже с западная медоносная пчела ( Apis mellifera ). [5] [6]
Этимология и общие имена
[ редактировать ]Видовой эпитет красного огненного муравья, invicta , происходит от латинского языка и означает «непобедимый» или «непобедимый». [7] [8] [9] Эпитет происходит от фразы Roma invicta («непокоренный Рим»), которая использовалась в качестве вдохновляющей цитаты до падения Западной Римской империи в 476 году нашей эры . Родовое название Solenopsis переводится как « вид трубки». Это соединение двух древнегреческих слов: solen , означающего «труба» или «канал», и opsis , означающего «видимость» или «зрение». [10] [11] Муравей широко известен как «красный импортированный огненный муравей» (сокращенно RIFA). Часть « огненного муравья » связана с ощущением жжения, вызванным его укусом. [12] [13] Альтернативные названия включают: «огненный муравей», «красный муравей» или «муравей-бродяга». [14] [15] В Бразилии местные жители называют муравья toicinhera , что происходит от португальского слова toicinho (свиной жир). [16]
Таксономия
[ редактировать ]
Красный импортированный огненный муравей был впервые описан швейцарским энтомологом Феликсом Санчи в журнальной статье 1916 года, опубликованной Physis . [17] Первоначально названный Solenopsis saevissima wagneri от рабочего -синтиписта , собранного в Сантьяго-дель-Эстеро , Аргентина, Санчи полагал, что муравей был вариантом S. saevissima ; видовой эпитет wagneri происходит от фамилии Э. Р. Вагнера, собравшего первые экземпляры. [18] Шрифтовой , Швейцария, но дополнительные работники по материал в настоящее время хранится в Музее естествознания в Базеле шрифтам, возможно, размещены в Национальном музее естественной истории в Париже. [19] В 1930 году американский мирмеколог Уильям Крейтон проанализировал род Solenopsis и реклассифицировал таксон как Solenopsis saevissimalectra wagneri во внутриподвидовом ранге, отметив, что он не смог собрать ни одного рабочего вида, который ссылался на первоначальное описание Санчи. [20] В 1952 году комплекс видов S. saevissima был исследован и вместе с девятью другими названиями видовых групп S. saevissimalectra wagneri был синонимом S. saevissima saevissima . [21] Эта реклассификация была принята австралийским энтомологом Джорджем Эттершанком в его пересмотре рода и в каталоге неотропических муравьев Уолтера Кемпфа 1972 года. [22] [23]
В 1972 году американский энтомолог Уильям Бюрен описал, по его мнению, новый вид, назвав его Solenopsis invicta . [24] Бюрен собрал рабочий- голотип из Куябы в Мату-Гросу , Бразилия, и предоставил первое официальное описание муравья в журнальной статье, опубликованной Энтомологическим обществом Джорджии. Он случайно написал invicta с ошибкой как invica [ sic ] над страницами с описанием вида, хотя было ясно, что invicta было предполагаемым написанием из-за постоянного использования этого названия в статье. [25] Типовой материал в настоящее время хранится в Национальном музее естественной истории в Вашингтоне, округ Колумбия. [24]

В обзоре комплекса видов в 1991 году американский энтомолог Джеймс Трэгер как синонимы объединил S. saevissimalectra wagneri и S. wagneri . [25] Трагер неправильно цитирует Solenopsis saevissimalectra wagneri в качестве оригинального названия, ошибочно полагая, что имя S. wagneri недоступно, и использовал имя Бюрена S. invicta . Ранее Трагер считал, что S. invicta конспецифичен S. saevissima, пока не сравнил материал с S. wagneri . Трагер отмечает, что хотя S. wagneri имеет приоритет над S. invicta , это название никогда не использовалось выше инфраподвидового ранга. Использование названия со времен Санчи не было связано с собранными экземплярами, и в результате это nomen nudum . [25] В 1995 году английский мирмеколог Барри Болтон исправил ошибку Трагера, признав S. wagneri действительным названием и превратив S. invicta в синоним . [26] Он заявляет, что Трагер ошибочно классифицировал S. wagneri как недоступное название, и ссылается на S. saevissimalectra wagneri как на исходный таксон. Он заключает, что S. wagneri на самом деле является оригинальным названием и имеет приоритет над S. invicta . [26] [27]
В 1999 году Стив Шаттак и его коллеги предложили сохранить название S. invicta . [19] С момента первого описания S. invicta было опубликовано более 1800 научных статей с использованием этого названия, в которых обсуждался широкий круг тем, касающихся его экологического поведения, генетики, химической связи, экономических последствий, методов контроля, популяций и физиологии. Они заявляют, что использование S. wagneri представляет собой «угрозу» номенклатурной стабильности по отношению к ученым и неученым; систематики, возможно, смогли адаптироваться к такому изменению названия, но если такой случай произошел, может возникнуть путаница в названиях. В связи с этим Шаттук и его коллеги предложили продолжать использовать S. invicta , а не S. wagneri , поскольку это название использовалось редко; между 1995 и 1998 годами было опубликовано более 100 статей с использованием S. invicta и только три с использованием S. wagneri . Они потребовали, чтобы Международная комиссия по зоологической номенклатуре (МКЗН) использовала свои полномочия для подавления S. wagneri в целях соблюдения принципа приоритета, а не принципа омонимии . Кроме того, они потребовали, чтобы название S. invicta быть добавлен в Официальный список видовых названий в зоологии, а S. wagneri быть добавлен в Официальный указатель отклоненных недействительных видовых названий в зоологии. [19] После рассмотрения предложение было проголосовано энтомологическим сообществом и было поддержано всеми, кроме двух избирателей. нет никаких оснований Они отмечают, что подавлению S. wagneri ; вместо этого было бы лучше отдать предпочтение S. invicta перед S. wagneri всякий раз, когда автор рассматривал их как виды одного вида. МКЗН сохранит S. invicta и подавит S. wagneri в обзоре 2001 года. [28] По современной классификации красный импортный огненный муравей является представителем рода Solenopsis трибы Solenopsidini , подсемейства Myrmicinae . Это член семейства Formicidae , принадлежащий к отряду Hymenoptera , отряду насекомых, в состав которого входят муравьи, пчелы и осы . [29]


Филогения
[ редактировать ]Красный завезенный огненный муравей является членом S. saevissima видовой группы . Членов можно отличить по двухсуставчатым булавам на конце канатика у рабочих и королев, а у более крупных рабочих второй и третий сегменты канатика вдвое длиннее и шире. Полиморфизм встречается у всех видов, а нижние челюсти несут четыре зуба. [24] На следующей кладограмме показано положение завезенного красного огненного муравья среди других представителей S. saevissima : видовой группы [а] [30]
Фенотипические и генетические данные позволяют предположить, что красный завезенный огненный муравей и черный завезенный огненный муравей ( Solenopsis richteri ) отличаются друг от друга, но имеют близкое генетическое родство. [31] [32] [33] [34] Гибридизация двух муравьев происходит в тех местах, где они вступают в контакт, причем гибридная зона расположена в Миссисипи. Такая гибридизация возникла в результате вторичного контакта между этими двумя муравьями несколько десятилетий назад. [ когда? ] , когда они впервые встретились друг с другом на юге Алабамы . [31] [35] На основе митохондриальной ДНК исследованные гаплотипы не образуют монофилетическую кладу . Некоторые из исследованных гаплотипов образуют более близкое родство с S. megergates , S. quinquecuspis и S. richteri , чем с другими гаплотипами S. invicta . Возникновение возможной парафилетической группировки позволяет предположить, что красный импортированный огненный муравей и S. quinquecuspis являются возможными загадочными группами видов, состоящими из нескольких видов, которые невозможно различить морфологически. [34] [36]
Генетика
[ редактировать ]Исследования показывают, что митохондриальной ДНК вариации в значительной степени происходят в полигинных обществах (гнездах с несколькими матками). [37] вариаций не обнаружено . но в моногинных обществах (гнезда с одной маткой) [38] Триплоидия (хромосомная аномалия) встречается у красных завезенных огненных муравьев с высокой частотой (до 12% у нерепродуктивных самок), что связано с высокой частотой диплоидных самцов. [39] Красный импортированный огненный муравей — первый вид, обладающий геном зеленой бороды , с помощью которого естественный отбор может способствовать альтруистическому поведению . Рабочие, содержащие этот ген, способны различать маток, содержащих его, от маток, у которых его нет, по-видимому, по запаху. Рабочие убивают маток, не содержащих этот ген. [40] [41] В 2011 году ученые объявили, что полностью секвенировали красного импортированного огненного муравья геном самца. [42]
Описание
[ редактировать ]
Завезенные красные огненные муравьи имеют размер от маленького до среднего, что делает их полиморфными . У рабочих размеры составляют от 2,4 до 6,0 мм (от 0,094 до 0,236 дюйма). [43] Головка имеет размеры от 0,66 до 1,41 мм (от 0,026 до 0,056 дюйма) и ширину от 0,65 до 1,43 мм (от 0,026 до 0,056 дюйма). У более крупных рабочих (как и у основных рабочих) ширина головы составляет от 1,35 до 1,40 мм (от 0,053 до 0,055 дюйма) и от 1,39 до 1,42 мм (от 0,055 до 0,056 дюйма). Размер антенных лопаток составляет от 0,96 до 1,02 мм (от 0,038 до 0,040 дюйма), а длина грудной клетки - от 1,70 до 1,73 мм (от 0,067 до 0,068 дюйма). [24] Голова становится шире за глазами с наличием округлых затылочных долей , и в отличие от похожего на вид S. richteri , доли достигают вершины дальше средней линии, но затылочное иссечение не такое складчатое. Скапус у основных рабочих не выходит за пределы затылочной вершины на один-два диаметра скапуса; эта особенность более заметна у S. richteri . У рабочих средних размеров скапусы достигают затылочных вершин, а у самых мелких рабочих превышают заднюю границу. У мелких и средних работников голова имеет более эллиптические стороны. Голова мелких рабочих спереди шире, чем сзади. [24] У основных рабочих переднеспинка не имеет ни угловатых плеч, ни впалой заднемедиальной области. Промезонотум выпуклый, основание проподеума округлое и также выпуклое. Основание и скат имеют одинаковую длину. У более крупных рабочих шов . промезонотума бывает прочным или слабым Черешок ; имеет толстую и тупую чешуйку если смотреть сзади, то сверху он не такой закругленный, как у S. richteri , а иногда может быть усеченным. Раструб большой и широкий, а у более крупных рабочих он шире своей длины. Раструб, как правило, менее широкий спереди и более широкий сзади. На задней стороне спинной поверхности имеется поперечный вдавление. У S. richteri эта особенность также присутствует, но гораздо слабее. [24]
Скульптура очень похожа на С. Рихтери . [24] Точки возникают там, где возникает волосатость, и они часто имеют удлиненную форму на дорсальной и вентральной частях головы. На груди имеются борозды, но они менее выражены и имеют меньше точек, чем у S. richteri . На черешке точки расположены по бокам. Раструб, если смотреть сверху, имеет сильную шагрень с отчетливыми поперечными точками. Бока покрыты глубокими точками, где они кажутся меньше, но глубже. У S. richteri точки крупнее и неглубоки. Это придает поверхности более непрозрачный вид. В некоторых случаях вокруг задней части могут присутствовать пунктострии. [24] Волосистость похожа на волосистость S. richteri . Эти волосы стоят прямостоячие и различаются по длине, кажутся длинными с каждой стороны переднеспинки и мезонотума ; на голове длинные волосы расположены продольными рядами. На черешковой чешуе многочисленные прижатые опушенные волоски; все наоборот у S. richteri , так как эти волоски редкие. [24] Рабочие кажутся красными и несколько желтоватыми с коричневым или полностью черным брюшком . [17] Желудочные пятна иногда наблюдаются у более крупных рабочих, у которых они не так ярко окрашены, как у S. richteri . Желудочное пятно обычно покрывает небольшую часть первого тергита желудка . Грудь одноцветная, от светло-красновато-коричневого до темно-коричневого. Ноги и тазики обычно слегка заштрихованы. У крупных рабочих голова имеет однородный цвет, при этом затылок и макушка кажутся коричневыми. Остальные части головы, включая переднюю часть, щеки и центральную часть наличника , желтоватые или желтовато-коричневые. Передние края щек и мандибул темно-коричневые; Кроме того, оба они, кажется, имеют один и тот же оттенок с затылком. Скапусы и канатики варьируются от того же цвета, что и голова, до того же оттенка, что и затылок. Светлые участки головы у рабочих малого и среднего размера ограничены только лобной областью, при этом присутствует темная отметина, напоминающая стрелу или ракету. Иногда гнезда могут иметь разные цвета. Например, рабочие могут быть значительно темнее, а желудочное пятно может совсем отсутствовать или выглядеть темно-коричневым. [24]
Королевы имеют длину головы от 1,27 до 1,29 мм (от 0,050 до 0,051 дюйма) и ширину от 1,32 до 1,33 мм (от 0,052 до 0,052 дюйма). [24] Размер скапусов составляет от 0,95 до 0,98 мм (от 0,037 до 0,039 дюйма), а грудная клетка - от 2,60 до 2,63 мм (от 0,102 до 0,104 дюйма). Голова почти неотличима от S. richteri , но затылочный вырез менее складчатый, а черешки значительно короче. Его черешковая чешуя выпуклая и напоминает таковую у S. richteri . Раструб имеет прямые стороны, которые никогда не вогнуты, в отличие от S. richteri, у которого они вогнуты. Грудная клетка почти идентична, но свободное пространство между метаплевральной полосатой областью и проподеальными дыхальцами либо представляет собой узкую складку, либо отсутствует. Боковые части черешка пунктированные. Бока раструба непрозрачные , имеются точки, но неправильной шероховатости не наблюдается. Передняя часть спины шагреневая , средний и задний отделы несут поперечные точечные штрихи. Во всех этих регионах волосы стоят стоячие. Передние части как черешка, так и раструба имеют прижатое опушение, которое также заметно на проподеуме. Окраска королевы аналогична рабочей: брюшко темно-коричневое, ноги, скапусы и грудь светло-коричневые с темными пестринами на мезоскутум . Голова в центральной части желтоватая или желтовато-коричневая, затылок и жвалы такого же цвета, как грудная клетка, а жилки крыльев варьируются от бесцветных до бледно-коричневых. [24] Самцы внешне похожи на S. richteri , но верхние края черешковых чешуек более вогнутые. У обоих видов дыхальца раструба и черешка сильно выступают. Все тело самца однотонно-черное, но усики беловатые. Как и у королевы, жилки крыльев бесцветные или бледно-коричневые. [24]

Красный импортированный муравей может быть ошибочно принят за похожего на него S. richteri . [24] Эти два вида можно отличить друг от друга с помощью морфологического исследования головы, грудной клетки и раструба. У S. richteri бока головы широко эллиптические, а сердцевидная форма, наблюдаемая у красного завезенного огненного муравья, отсутствует. Область затылочных долей, расположенная вблизи средней линии и затылочной вырезки, у S. richteri выглядит более складчатой , чем у красного завезенного огненного муравья. Скребки S. richteri длиннее, чем у красного завезенного огненного муравья, а переднеспинка имеет сильные угловатые плечи. Такой признак практически отсутствует у красного привозного огненного муравья. Неглубокий, но затонувший участок известен только у более крупных рабочих S. richteri и расположен в задней части спинки переднеспинки. Эта особенность полностью отсутствует у более крупных импортированных рабочих огненных муравьев красного цвета. Промезонотум красного импортного огненного муравья сильно выпуклый, тогда как у S. richteri этот признак слабо выпуклый . При осмотре основание проподеума удлиненное и прямое. S. richteri , хотя и выпуклый и более короткий, красного цвета, импортированный огненный муравей. У него также есть широкий раструб с прямыми или расходящимися сторонами. Раструб у S. richteri уже, со сходящимися сторонами. У S. richteri поперечное отпечаток на заднедорсальной части раструба сильный, но слабый или отсутствует у красного завезенного огненного муравья. [24] Кроме того, рабочие S. richteri на 15% крупнее импортных красных огненных муравьев, имеют черновато-коричневый цвет и имеют желтую полосу на спинной стороне брюшка. [44] [45]
Выводок
[ редактировать ]
Яйца крошечные, овальной формы, сохраняют одинаковый размер около недели. Через неделю яйцо принимает форму эмбриона и превращается в личинку при удалении яичной скорлупы. [46] Размер личинок составляет 3 мм (0,12 дюйма). [47] По внешнему виду они похожи на личинок S. geminata , но их можно отличить по покровам с шипиками на вершине дорсальной части задних сомитов. Волосы на теле имеют длину от 0,063 до 0,113 мм (от 0,0025 до 0,0044 дюйма) с зубчатым кончиком. Оба усика имеют по две или три сенсиллы . меньше Верхняя губа по размеру, с двумя волосками на передней поверхности размером 0,013 мм (0,00051 дюйма). На верхней челюсти имеется склеротизированная полоса между кардо и ножкой. также На губе имеется небольшая склеротизированная полоса. [47] трубочки губных желез Известно, что производят или секретируют белковое вещество, имеющее богатый уровень пищеварительных ферментов, в том числе протеазы и амилазы, которые выполняют функцию внекишечного переваривания твердой пищи. Средняя кишка также содержит амилазы , ротазы и упазы. Узкие клетки в его резервуаре практически не участвуют в секреции. [48] Куколки напоминают взрослых особей любой касты, за исключением того, что их ноги и усики плотно прижаты к телу. Они кажутся белыми, но со временем куколка темнеет, когда они почти готовы созреть. [3]
четыре личиночных возраста . На основе отличительных морфологических признаков описаны [46] [49] [50] Личинок второстепенных и основных рабочих невозможно отличить до финального возраста, когда становятся очевидными различия в размерах. [50] После окукливания разница в ширине головы между кастами становится более очевидной. Репродуктивные личинки крупнее рабочих личинок и имеют отдельные морфологические различия в ротовом аппарате. [50] Личинок четвертого возраста самцов и королев можно отличить по относительной форме и окраске тела. [50] а также могут различаться внутренние гоноподальные имагинальные диски.
Полиморфизм
[ редактировать ]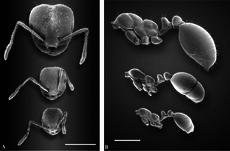
Красный импортированный огненный муравей полиморфен с двумя разными кастами рабочих: мелкими рабочими и крупными рабочими (солдатами). Как и многие муравьи, демонстрирующие полиморфизм, молодые, более мелкие муравьи не добывают пищу, а вместо этого склоняются к выводку, в то время как более крупные рабочие выходят и добывают пищу. [51] [52] [53] [54] В зарождающихся колониях полиморфизма не существует, а вместо них их заселяют мономорфные рабочие, называемые «минимами» или «нанитиками». Средняя ширина головы в тестируемых семьях увеличивается в течение первых шести месяцев развития. [55] [56] В пятилетних колониях ширина головы у мелких рабочих уменьшается, а у крупных рабочих ширина головы остается прежней. Общий вес крупных рабочих в два раза превышает вес мелких рабочих на момент их первого прибытия, а к шести месяцам основные рабочие становятся в четыре раза тяжелее мелких рабочих. Как только основные рабочие вырастут, они смогут составить значительную часть рабочей силы, при этом до 35% будут основными рабочими в одной колонии. [55] Это не влияет на продуктивность семей, поскольку полиморфные семьи и гнезда с небольшими рабочими производят выводок примерно с одинаковой скоростью, а полиморфизм не является преимуществом или недостатком, если источники пищи не ограничены. Однако полиморфные семьи более энергетически эффективны, и в условиях ограниченности пищи полиморфизм может дать небольшое преимущество в производстве потомства, но это зависит от уровня пищевого стресса. [57]
По мере роста рабочих муравьев форма головы меняется, поскольку длина головы увеличивается одновременно с общей длиной тела, а ширина головы может увеличиваться на 20%. Длина усиков растет медленно; усики могут вырасти только на 60% длиннее к тому времени, когда тело удваивает свою длину, таким образом, относительная длина усиков уменьшается на 20% по мере увеличения длины тела вдвое. [56] Все отдельные ноги тела изометричны, а длина тела означает, что даже если длина тела удваивается, ноги также удваиваются. Однако не все ноги одинаковой длины; переднегрудная часть составляет 29% длины ноги, среднегрудная 31% и заднегрудная 41%. Первые две пары ног имеют одинаковую длину, тогда как последняя пара длиннее. [56] В целом морфологический облик рабочего резко меняется по мере его роста. Голова демонстрирует наибольшее изменение формы, а высота алинотума растет быстрее, чем его длина, при этом соотношение высоты к длине составляет 0,27 у второстепенных рабочих и 0,32 у основных рабочих. [56] Из-за этого более крупные рабочие, в отличие от более мелких рабочих, обычно имеют горбатую форму и крепкий алинотум. Ни один сегмент черешка не меняет форму по мере изменения размера тела. Ширина брюшка растет быстрее, чем его длина, при этом ширина может составлять 96% его длины, но увеличивается до 106%. [56]
Физиология
[ редактировать ]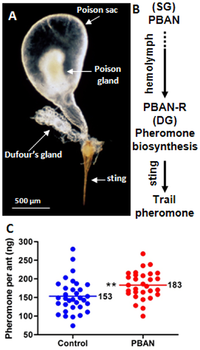
Как и другие насекомые, красный завезенный огненный муравей дышит через систему заполненных газом трубок, называемых трахеями, соединенных с внешней средой посредством дыхалец. Конечные ветви трахеи ( трахеолы ) непосредственно контактируют с внутренними органами и тканями. Транспорт кислорода к клеткам (и углекислого газа из клеток) происходит посредством диффузии газов между трахеолами и окружающими тканями и сопровождается прерывистым газообменом . [58] Как и у других насекомых, прямая связь между трахейной системой и тканями устраняет необходимость в сети циркулирующей жидкости для транспортировки O 2 . [59] Таким образом, завезенные красные огненные муравьи и другие членистоногие могут иметь скромную систему кровообращения, хотя их метаболические потребности очень высоки. [60]
Выделительная система состоит из трех отделов. Базальная область имеет три клетки, расположенные в задней части средней кишки. Передняя и верхняя полости образованы основаниями четырех мальпигиевых канальцев . [61] Верхняя полость открывается в просвет тонкой кишки. Прямая кишка представляет собой большой, но тонкостенный мешок, занимающий заднюю пятую часть личинки. Выброс отходов контролируется ректальными клапанами , ведущими в задний проход. [61] Иногда личинки выделяют жидкость, состоящую из мочевой кислоты , воды и солей. [62] Это содержимое часто выносится рабочими на улицу и выбрасывается, но колонии, находящиеся в условиях дефицита воды, могут потреблять это содержимое. [61] В репродуктивной системе королевы выделяют феромон , который предотвращает размножение и оогенез у девственных самок; те, которые были протестированы в семьях без матки, начинают развитие ооцитов после сдачи и берут на себя роль яйцекладки. [63] Дегенерация летательных мышц инициируется брачными и ювенильными гормонами и предотвращается аллатэктомией тела . [64] [65] Гистолиз начинается с растворения миофибрилл и медленного разрушения миофиламентов . Такое растворение продолжается до тех пор, пока не достигнет единственного свободного материала Z-линии, который также исчезнет; остаются только ядра и пластинчатые тельца . [61] В одном исследовании количество аминокислот в гемолимфе увеличивается после осеменения. [66]
Железистая система содержит четыре железы: нижнечелюстные, верхнечелюстные, губные и заглоточные железы. [61] У королевы хорошо развита заглоточная железа, у рабочих другие железы крупнее. Заглоточная железа функционирует как вакуум для поглощения жирных кислот и триглицеридов , а также как слепая кишка желудка . [67] Функции других желез остаются малоизученными. В одном исследовании, посвященном ферментам пищеварительной системы взрослых муравьев, активность липазы была обнаружена в нижнечелюстных и губных железах, а также активность инвертазы . обнаруженная Железа Дюфура, у муравья, действует как источник следовых феромонов , хотя ученые считали, что источником феромона королевы является ядовитая железа. [61] [68] [69] У муравья обнаружен нейропептид , активирующий биосинтез феромонов нейрогормонов из железы Дюфура. [70] . У маток имеется сперматека, которая отвечает за поддержание спермы У самцов эти железы, по-видимому, отсутствуют, но те, что связаны с его головой, морфологически аналогичны тем, что есть у рабочих, но эти железы могут действовать по-другому. [67]

Муравей сталкивается со многими респираторными проблемами из-за очень изменчивой окружающей среды, которая может вызвать повышенное высыхание , гипоксию и гиперкапнию . Жаркий и влажный климат вызывает увеличение частоты сердечных сокращений и дыхания, что увеличивает потерю энергии и воды. [59] [71] Гипоксия и гиперкапния могут возникнуть в результате завоза красных колоний огненных муравьев, живущих в плохо вентилируемых терморегуляторных курганах и подземных гнездах. Прерывистый газообмен (DGE) может позволить муравьям выжить в гиперкапнических и гипоксических условиях, часто встречающихся в их норах; [58] он идеален для адаптации к этим условиям, поскольку позволяет муравьям самостоятельно увеличивать период потребления O 2 и выделения CO 2 посредством манипуляций с дыхалцами. Успех инвазии красного завезенного огненного муравья, возможно, может быть связан с его физиологической толерантностью к абиотическому стрессу , поскольку он более устойчив к жаре и более адаптируется к стрессу, связанному с высыханием, чем S. richteri . Это означает, что муравей менее уязвим к жаре и стрессу от высыхания. Хотя S. richteri имеет более высокое содержание воды в организме, чем красный завезенный огненный муравей, S. richteri был более уязвим к стрессу от высыхания. Меньшая чувствительность к высыханию обусловлена меньшей скоростью потери воды. [72] Колонии, живущие в незатененных и более теплых местах, как правило, имеют более высокую жароустойчивость, чем те, которые живут в затененных и более прохладных местах. [73]
На скорость обмена веществ, которая косвенно влияет на дыхание, также влияет температура окружающей среды. Пик метаболизма происходит при температуре около 32 °C. [74] Метаболизм и, следовательно, частота дыхания постоянно увеличиваются с повышением температуры. ДГЭ прекращается при температуре выше 25 °C, хотя причина этого на данный момент неизвестна. [75]
Судя по всему, на частоту дыхания также существенно влияет каста. У самцов частота дыхания значительно выше, чем у самок и рабочих, отчасти из-за их способности к полету и более высокой мышечной массы. В целом у мужчин больше мышц и меньше жира, что приводит к более высокой метаболической потребности в О 2 . [75] Хотя скорость метаболизма самая высокая при температуре 32 °C, колонии часто процветают при немного более низких температурах (около 25 °C). Высокая скорость метаболической активности, связанная с более высокими температурами, является ограничивающим фактором роста колоний, поскольку потребность в потреблении пищи также увеличивается. В результате более крупные колонии, как правило, встречаются в более прохладных условиях, поскольку метаболические потребности, необходимые для поддержания колонии, снижаются. [74]
Распространение и среда обитания
[ редактировать ]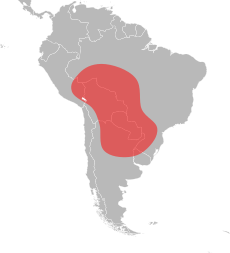
Красные завезенные огненные муравьи родом из тропических районов Центральной Южной Америки, где они имеют обширный географический ареал, простирающийся от юго-востока Перу до центральной Аргентины и юга Бразилии. [76] [77] [78] [79] В отличие от его географического ареала в Северной Америке, его ареал в Южной Америке существенно отличается. Он имеет чрезвычайно длинный ареал с севера на юг, но очень узкое распространение с востока на запад. Самая северная встреча красного завезенного огненного муравья - это Порту-Велью в Бразилии, а самая южная - Ресистенсия в Аргентине; это расстояние около 3000 км (1900 миль). Для сравнения, ширина его узкого ареала составляет около 350 км (220 миль), и, скорее всего, он уже, в южной Аргентине и Парагвае, а также в северных районах бассейна реки Амазонки. [80] Наиболее известные случаи завоза красного огненного муравья происходят в регионе Пантанал в Бразилии. Однако внутренняя часть этой территории досконально не исследована, но можно с уверенностью сказать, что этот вид встречается в благоприятных местах вокруг нее. Считается, что регион Пантанал является родиной завезенного красного огненного муравья; Распространение гидрохора через плавучие муравьиные плоты может легко объяснить популяцию крайнего юга вокруг рек Парагвай и Гуапоре . Западная протяженность его ареала точно неизвестна, но его численность там может быть ограничена. Он может быть обширным в самой восточной части Боливии из-за присутствия региона Пантанал. [80]
Эти муравьи родом из Аргентины, а красные огненные муравьи, скорее всего, пришли отсюда, когда они впервые вторглись в Соединенные Штаты; в частности, популяции этих муравьев были обнаружены в провинциях Чако , Корриентес , Формоза , Сантьяго-дель-Эстеро , Санта-Фе и Тукуман . [25] [80] [81] Северо-восточные регионы Аргентины являются наиболее вероятным местом происхождения вторгшихся муравьев. [78] В Бразилии они встречаются на севере Мату-Гросу, в Рондонии и в штате Сан-Паулу . Красный завезенный огненный муравей и S. saevissima являются парапатрическими в Бразилии, зоны контакта известны в Мату-Гросу-ду-Сул , штате Парана и Сан-Паулу. [77] [82] Они находятся в департаментах Бокерон , Каагуасу , Каниндейу , Центральный , Гуайра , Ньембуку , Парагуари и Президенти-Хейс ; Трагер утверждает, что муравей распространен во всех регионах страны. [83] [84] [85] Они также встречаются на большей части северо-востока Боливии и, в меньшей степени, на северо-западе Уругвая. [80] [86]

Красный завезенный огненный муравей способен доминировать на измененных территориях и жить в самых разных средах обитания. Он может пережить экстремальные погодные условия тропических лесов Южной Америки, а в нарушенных районах его гнезда часто можно увидеть вдоль дорог и зданий. [87] [88] Муравей часто наблюдался в поймах реки Парагвай. [89] В районах, где присутствует вода, они обычно встречаются вокруг: оросительных каналов, озер, прудов, водохранилищ, рек, ручьев, берегов рек и мангровых болот. [83] [87] Гнезда встречаются в сельскохозяйственных районах, на побережьях, водно-болотных угодьях, остатках прибрежных дюн, пустынях, лесах, лугах, естественных лесах, дубовых лесах, мезических лесах , в листовой подстилке, на окраинах пляжей , в кустарниках, вдоль железных дорог и дорог, а также в городских районах. [90] В частности, они встречаются на обрабатываемых землях, в управляемых лесах и плантациях, на нарушенных территориях, в системах интенсивного животноводства и в теплицах. [87] [91] Было обнаружено, что красные завезенные огненные муравьи проникают в здания, в том числе в медицинские учреждения. [92] В городской местности колонии обитают на открытой местности, особенно если местность солнечная. [92] Сюда входят: городские сады, места для пикников, лужайки, игровые площадки, школьные дворы, парки и поля для гольфа. [83] [92] В некоторых районах в среднем насчитывается 200 курганов на акр. [93] Зимой колонии перемещаются под тротуары или в здания, а недавно спарившиеся королевы перемещаются на пастбища. [87] [92] Красные завезенные огненные муравьи в основном встречаются на высоте от 5 до 145 м (от 16 до 476 футов) над уровнем моря. [83]
Курганы варьируются от маленьких до больших, их размеры составляют от 10 до 60 см (от 3,9 до 23,6 дюйма) в высоту и 46 см (18 дюймов) в диаметре без видимых входов. [87] [94] Рабочие могут получить доступ к своим гнездам только через серию туннелей, выступающих из центральной части. Такие выступы могут простираться на расстояние до 25 футов от центральной насыпи либо прямо вниз к земле, либо, что чаще, в сторону от первоначальной насыпи. [95] Построенные из почвы курганы ориентированы так, что длинные части кургана обращены к солнцу ранним утром и перед закатом. [87] [94] Курганы обычно имеют овальную форму, длинная ось гнезда ориентирована в направлении с севера на юг. [96] Эти муравьи также тратят большое количество энергии на строительство гнезда и транспортировку расплода, что связано с терморегуляцией . Выводок транспортируют в места с высокими температурами; рабочие отслеживают температурный режим кургана и не полагаются на поведенческие привычки. [97] Внутри гнезд насыпи содержат ряд узких горизонтальных туннелей с подземными шахтами и узлами, достигающими корней травы на глубине от 10 до 20 см (от 3,9 до 7,9 дюйма) под поверхностью; эти валы и узлы соединяют туннели кургана с подземными камерами. Эти камеры имеют размер около 5 см. 2 (0,77 дюйма 2 ) и достигают глубины от 10 до 80 см (от 3,9 до 31,5 дюйма). Среднее количество муравьев в одной подземной камере составляет около 200. [98] [99] [100]
Введение
[ редактировать ]Красные завезенные огненные муравьи являются одними из самых инвазивных видов в мире. [101] [102] Некоторые учёные считают завезённого красного огненного муравья «специалистом по беспокойству »; Нарушение человеком окружающей среды может быть основным фактором воздействия муравьев (огненные муравьи, как правило, предпочитают нарушенные территории). Об этом свидетельствует один эксперимент, показывающий, что покос и вспашка на изученных территориях уменьшили разнообразие и численность местных видов муравьев, тогда как красные завезенные огненные муравьи, обнаруженные на ненарушенных лесных участках, сократили лишь пару видов. [103] [104]
В США красный огненный муравей впервые прибыл в морской порт Мобил, штат Алабама , на грузовом корабле в период с 1933 по 1945 год. [б] [с] [80] [107] [108] [109] Прибыв примерно с 9–20 неродственными матками, [110] [111] красный завезенный огненный муравей в то время был редкостью, поскольку энтомологи не смогли собрать ни одного экземпляра (самые ранние наблюдения были впервые сделаны в 1942 году, чему предшествовало увеличение популяции в 1937 году); популяция этих муравьев резко возросла к 1950-м годам. [112] [113] [114] С момента своего появления в Соединенных Штатах завезенный красный огненный муравей распространился по южным штатам и северо-востоку Мексики, отрицательно влияя на дикую природу и нанося экономический ущерб. [76] [115] [116] [117] Распространение завезенных красных огненных муравьев может быть ограничено, поскольку зимой в Теннесси они практически истребляются и, таким образом, могут достичь [ когда? ] их самый северный ареал. [118] [119] [120] Однако глобальное потепление может позволить завезённому красному огненному муравью расширить свой географический ареал. [121] По состоянию на 2004 год муравей обитает в 13 штатах и занимает более 128 миллионов гектаров земли, а на одном акре земли можно найти до 400 курганов. [122] [123] , По оценкам Министерства сельского хозяйства США они расширяются на 120 миль (193 км) на запад в год. [92] Вероятно, из-за отсутствия южноамериканских конкурентов - и меньшего количества местных конкурентов - S. invicta доминирует над большим количеством внецветочных нектарников и полужесткокрылых источников пади на юге США, чем в своем родном ареале. [124]
Красные завезенные огненные муравьи были впервые обнаружены в Квинсленде , Австралия, в 2001 году. [125] [126] Считалось, что муравьи присутствовали в транспортных контейнерах, прибывших в порт Брисбена , скорее всего, из Северной Америки. [127] По неофициальным данным, огненные муравьи могли обитать в Австралии в течение шести-восьми лет до того, как их официально идентифицировали. Потенциальный ущерб от завезенного красного огненного муравья побудил правительство Австралии быстро отреагировать. совместное финансирование на уровне штата и федерального правительства в размере 175 миллионов австралийских долларов . На шестилетнюю программу искоренения было выделено [128] [129] [130] Сообщалось, что после нескольких лет ликвидации уровень ликвидации на ранее зараженных объектах недвижимости составил более 99%. Программа получила расширенное финансирование Содружества в размере около 10 миллионов австралийских долларов как минимум еще на два года для лечения остаточных заражений, обнаруженных совсем недавно. [131] В декабре 2014 года гнездо было обнаружено в Порт-Ботанике , Сидней , в Новом Южном Уэльсе . Порт был помещен на карантин, и была проведена операция по его удалению. [132] В сентябре 2015 года в аэропорту Брисбена были обнаружены популяции выходцев из США. [133] С тех пор на их искоренение были выделены сотни миллионов долларов. В августе 2023 года Совет по инвазивным видам заявил, что без дополнительного финансирования огненные муравьи, вероятно, распространится на север Нового Южного Уэльса и на запад, потенциально в бассейн Мюррей-Дарлинг. [134]

Красные завезенные огненные муравьи распространились за пределы Северной Америки. Группа специалистов по инвазивным видам (ISSG) сообщает, что муравей обитает на трех Каймановых островах . Однако источники, на которые ссылается ISSG, не сообщают о них на острове, но недавние коллекции указывают на их присутствие. [76] В 2001 году в Новой Зеландии были обнаружены завезенные красные огненные муравьи, но через несколько лет их успешно искоренили. [76] [135] В Индии зарегистрированы завезенные красные огненные муравьи. [136] Малайзия, [137] Филиппины [138] и Сингапур. [76] Однако эти отчеты оказались неверными, поскольку собранные там муравьи были ошибочно идентифицированы как импортированные красные огненные муравьи. В Сингапуре муравьев, скорее всего, тоже опознали неправильно. В Индии обследованные муравьи в Саттур-Талуке, Индия, включили туда красные завезенные огненные муравьи в высокие популяции; при этом за пределами обследуемой территории сообщений о появлении муравья не поступало. В 2016 году ученые заявили, что, несмотря на отсутствие муравья в Индии, красный завезенный огненный муравей, скорее всего, найдет подходящие места обитания в экосистеме Индии, если ему представится такая возможность. [139] В сообщениях на Филиппинах, скорее всего, собранный материал был ошибочно идентифицирован как красный завезенный огненный муравей, поскольку там не было обнаружено никаких популяций. [76] Он был обнаружен в Гонконге и материковом Китае в 2004 году, откуда распространился в несколько провинций, а также в Макао и Тайвань. [140] [141] [142] [143] Никакие географические или климатические барьеры не препятствуют дальнейшему распространению этих муравьев, поэтому они могут распространиться по тропическим и субтропическим регионам Азии. [122] [144] В Европе единственное гнездо было обнаружено в Нидерландах в 2002 году. [79] Впервые в 2023 году колонии муравьев были обнаружены в Европе. [145]
Примерно в 1980 году завезенные красные огненные муравьи начали распространяться по всей Вест-Индии , где о них впервые сообщили в Пуэрто-Рико и на Виргинских островах США . [146] [147] В период с 1991 по 2001 год муравей был зарегистрирован на Тринидаде и Тобаго, в нескольких районах Багамских островов, Британских Виргинских островов, Антигуа и островов Тёркс и Кайкос. [76] [148] [149] С тех пор красные завезенные огненные муравьи были зарегистрированы на большем количестве островов и регионов, а новые популяции обнаружены в Ангилье , Сен-Мартене , Барбуде , Монтсеррате , Сент-Китсе , Невисе , Арубе и Ямайке. [76] Муравьев, зарегистрированных на Арубе и Ямайке, можно встретить только на полях для гольфа; эти курсы импортируют дерн из Флориды , поэтому такой импорт может быть важным способом распространения муравьев по всей Вест-Индии. [76]
Популяции, обнаруженные за пределами Северной Америки, происходят из Соединенных Штатов. В 2011 году была проанализирована ДНК образцов из Австралии, Китая и Тайваня, и результаты показали, что они родственны образцам из Соединенных Штатов. [150] Несмотря на распространение красного завозного огненного муравья ( S. invicta ), S. geminata имеет больший географический ареал, но его легко вытесняет S. invicta . Из-за этого почти весь экзотический ареал S. geminata в Северной Америке утрачен и он там почти исчез. На обочинах дорог во Флориде на 83% этих участков присутствовал S. Geminata , когда красный завезенный огненный муравей отсутствовал, но только на 7%, когда он присутствовал. [76] Это означает, что муравей, вероятно, может проникнуть во многие тропические и субтропические регионы, где S. Geminata . присутствуют популяции [76]
Поведение и экология
[ редактировать ]
Красные завезенные огненные муравьи чрезвычайно устойчивы и приспособились противостоять как наводнениям, так и засухе. Если муравьи чувствуют повышение уровня воды в своих гнездах, они объединяются и образуют плавающий шар или плот с рабочими снаружи и королевой внутри. [151] [152] [153] Выводок транспортируется на самую высокую поверхность. [154] Они также используются в качестве основы плота, за исключением яиц и более мелких личинок. Прежде чем погрузиться в воду, муравьи ныряют в воду и разрывают связь с сушей. В некоторых случаях рабочие могут намеренно убрать всех самцов с плота, в результате чего самцы утонут. Срок службы плота может достигать 12 дней. Муравьи, попавшие в ловушку под водой, спасаются, поднимаясь на поверхность с помощью пузырьков, собранных из подводного субстрата. [154] Из-за большей уязвимости для хищников красные завезенные огненные муравьи значительно более агрессивны при сплаве. Работники, как правило, вводят более высокие дозы яда, что снижает угрозу нападения других животных. Из-за этого, а также из-за наличия более высокой численности муравьев, плоты потенциально опасны для тех, кто с ними сталкивается. [155]
Некрофорное поведение наблюдается у красного завезенного огненного муравья. Рабочие выбрасывают несъеденную пищу и другие подобные отходы подальше от гнезда. Активный компонент не был идентифицирован, но в нем участвовали жирные кислоты, накапливающиеся в результате разложения, а кусочки бумаги, покрытые синтетической олеиновой кислотой, обычно вызывали некрофорную реакцию. Процесс, лежащий в основе такого поведения импортированных красных огненных муравьев, был подтвержден Блюмом (1970): ненасыщенные жиры, такие как олеиновая кислота, вызывают поведение по удалению трупов. [156] Рабочие также по-разному реагируют на мертвых рабочих и куколок. Мертвых рабочих обычно забирают из гнезда, тогда как у куколок может пройти день, прежде чем произойдет некрофорная реакция. Куколки, зараженные Metarhizium anisopliae , обычно чаще выбрасываются рабочими; В течение суток выбрасывают 47,5% непораженных трупов, а для пораженных трупов этот показатель составляет 73,8%. [157]
Красные завезенные огненные муравьи отрицательно влияют на прорастание семян . Однако степень ущерба зависит от того, как долго семена уязвимы (высохнут и прорастают), а также от численности муравьев. [158] Одно исследование показало, что, хотя этих муравьев привлекают семена, которые адаптировались к распространению муравьев, и удаляют их, красные завезенные огненные муравьи повреждают эти семена или перемещают их в неблагоприятные места для прорастания. В семенах, переданных колониям, 80% семян Sanguinaria canadensis были скарифицированы и 86% семян Viola rotundifolia были уничтожены. [159] Небольшой процент семян сосны длиннолистной ( Pinus palustris ), оставленные рабочими, успешно прорастают, тем самым доказывая, что красные завезенные огненные муравьи помогают движению семян в экосистеме длиннолистной сосны. Семена, несущие элайосомы, собираются быстрее, чем семена, не несущие элайосомы, и складируют их не в гнездах, а в поверхностных кучах мусора в окрестностях кургана. [160]
Собирательство и общение
[ редактировать ]
Колонии завезенных красных огненных муравьев имеют поверхность туннелей, выступающую из поверхностей, на которых рабочие добывают корм. [161] [162] Эти области выступа, как правило, находятся на своей территории, но на это может повлиять большая колонизация муравьев. [163] Туннели спроектированы таким образом, чтобы обеспечить эффективное взаимодействие тела, конечностей и усиков со стенами, а внутри них рабочий может перемещаться исключительно быстро (более девяти длин тела в секунду). [164] Норы выходят из любой точки территории колонии, и работникам-собирателям, возможно, придется пройти полметра, чтобы добраться до поверхности. Если предположить, что среднестатистический собиратель преодолевает расстояние 5 м, то более 90% времени поиска пищи приходится на туннели днем и редко ночью. Рабочие добывают корм при температуре почвы, достигающей 27 ° C (81 ° F), и температуре поверхности 12–51 ° C (54–124 ° F). [162] Рабочие, подвергающиеся воздействию температуры 42 °C (108 °F), рискуют умереть от жары. [162] К осени количество рабочих, добывающих пищу, быстро падает, и зимой они редко выходят на поверхность. Это может быть связано с воздействием температуры почвы и снижением предпочтения источников пищи. Эти предпочтения уменьшаются только тогда, когда производство расплода низкое. В северных регионах США слишком холодно, чтобы муравьи могли добывать пищу, но в других регионах, таких как Флорида и Техас , добыча пищи может происходить круглый год. Во время дождя рабочие не добывают корм на улице, так как выходные отверстия временно блокируются, следы феромонов смываются, а собиратели могут физически пострадать от дождя. Влажность почвы также может влиять на пищевое поведение рабочих. [162]
Когда рабочие добывают пищу, он состоит из трех этапов: поиск, вербовка и транспортировка. [165] Рабочие, как правило, ищут мед чаще, чем другие источники пищи, и вес еды не влияет на время поиска. Рабочие могут нанять других товарищей по гнезду, если найденная ими еда слишком тяжелая, и для прибытия максимального количества набранных рабочих требуется до 30 минут. Более легкие источники пищи занимают меньше времени и обычно транспортируются быстро. [165] Рабочие-собиратели становятся разведчиками и ищут пищу исключительно за пределами поверхности, а через две недели могут умереть от старости. [166]
Рабочие общаются с помощью ряда семиохимических веществ и феромонов . Эти методы общения используются в различных видах деятельности, таких как вербовка товарищей по гнезду, добыча пищи, привлечение и защита; например, рабочий может выделять следовые феромоны , если обнаруженный им источник пищи слишком велик, чтобы его можно было нести. [167] Эти феромоны синтезируются железой Дюфура и могут перемещаться от обнаруженного источника пищи обратно в гнездо. [168] [169] Компоненты этих следовых феромонов также видоспецифичны только для этого муравья, в отличие от других муравьев с общими хвостовыми феромонами. [170] Ядовитый мешок этого вида был идентифицирован как новое место хранения феромона королевы; Известно, что этот феромон вызывает ориентацию у рабочих особей, что приводит к отложению расплода. [69] [171] Это также аттрактант: рабочие собираются в районах, где был выпущен феромон. [167] Возможно, присутствует выводковый феромон, поскольку рабочие могут разделять выводок по возрасту и касте, после чего следует облизывание, уход и наращивание усиков . [167] Если колония подверглась нападению, рабочие выпустят тревожные феромоны. [172] Однако у рабочих эти феромоны развиты слабо. Рабочие могут обнаружить пиразины , вырабатываемые крыльями; эти пиразины могут участвовать в брачном полете , а также в реакции тревоги. [173]
Красные завезенные огненные муравьи могут отличать соседей по гнезду от несоседей по химическому общению и специфическому запаху колонии. [167] [174] Рабочие предпочитают копать материал для гнезд из своей собственной колонии, а не из почвы на негнездовых участках или из других завезенных красных колоний огненных муравьев. Одно исследование предполагает, что, поскольку диета колоний одинакова, единственная разница между гнездовой и негнездовой почвой заключалась в гнездовании самих муравьев. Таким образом, рабочие могут переносить запах колоний в почву. [174] На запах колонии может влиять окружающая среда, поскольку рабочие в колониях, выращенных в лаборатории, менее агрессивны, чем в дикой природе. [175] Сигналы, полученные от королевы, способны регулировать распознавание товарищей по гнезду у рабочих и уровень аминов . Однако эти сигналы не играют большой роли в распознавании на уровне колонии, но могут служить формой распознавания касты внутри гнезд. [176] [177] Рабочие, живущие в моногинных обществах, склонны быть чрезвычайно агрессивными и нападать на злоумышленников из соседних гнезд. В колониях без маток добавление чужеродных маток или рабочих не увеличивает агрессию среди населения. [178]
Диета
[ редактировать ]
Красные импортированные огненные муравьи всеядны , а собиратели считаются падальщиками, а не хищниками. [179] Рацион муравьев состоит из мертвых млекопитающих, членистоногих , [180] insects, earthworms, vertebrates, and solid food matter such as seeds. However, this species prefers liquid over solid food. The liquid food the ants collect is sweet substances from plants or honeydew-producing hemipterans.[94][179][181][182] Arthropod prey may include dipteran adults, larvae and pupae, and termites. The consumption of sugar amino acid is known to affect recruitment of workers to plant nectars. Mimic plants with sugar rarely have workers to feed on them, whereas those with sugar and amino acids have considerable numbers.[183][184] The habitats where they live may determine the food they collect the most; for example, forage success rates for solid foods are highest in lakeshore sites, while high levels of liquid sources were collected from pasture sites.[185] Specific diets can also alter the growth of a colony, with laboratory colonies showing high growth if fed honey-water. Colonies that feed on insects and sugar-water can grow exceptionally large in a short period of time, whereas those that do not feed on sugar-water grow substantially slower. Colonies that do not feed on insects cease brood production entirely.[182] Altogether, the volume of food digested by nestmates is regulated within colonies.[186] Larvae are able to display independent appetites for sources such as solid proteins, amino acid solutions, and sucrose solutions, and they also prefer these sources over dilute solutions. Such behaviour is due to their capability to communicate hunger to workers. The rate of consumption depends on the type, concentration, and state of the food on which they feed. Workers tend to recruit more nestmates to food sources filled with high levels of sucrose than to protein.[186]
Food distribution plays an important role in a colony. This behaviour varies in colonies, with small workers receiving more food than larger workers if a small colony is seriously deprived of food. In larger colonies, however, the larger workers receive more food. Workers can donate sugar water efficiently to other nestmates, with some acting as donors. These "donors" distribute their food sources to recipients, which may also act as donors. Workers may also share a greater portion of their food with other nestmates.[187] In colonies that are not going through starvation, food is still distributed among the workers and larvae. One study shows that honey and soybean oil were fed to the larvae after 12 to 24 hours of being retained by the workers. The ratio distribution of these food sources was 40% towards the larvae and 60% towards the worker for honey, and for soybean oil this figure was around 30 and 70%, respectively.[188] Red imported fire ants also stockpile specific food sources such as insect pieces rather than consuming them immediately. These pieces are usually transported below the mound surface and in the driest and warmest locations.[189]
This species engages in trophallaxis with the larvae.[190] Regardless of the attributes and conditions of each larva, they are fed roughly the same amount of liquid food. The rate of trophallaxis may increase with larval food deprivation, but such increase depends on the size of each larva. Larvae that are fed regularly tend to be given small amounts. To reach satiation, all larvae regardless of their size generally require the equivalent of eight hours of feeding.[191]
Predators
[edit]
A number of insects, arachnids, and birds prey on these ants, especially when queens are trying to establish a new colony.[192] While in the absence of defending workers, the fire ant queens must rely on their venom to keep off competitor species [2] Many species of dragonfly, including Anax junius, Pachydiplax longipennis, Somatochlora provocans, and Tramea carolina, capture the queens while they are in flight; 16 species of spiders, including the wolf spider Lycosa timuga and the southern black widow spider (Latrodectus mactans), actively kill red imported fire ants. L. mactans captures all castes of the species (the workers, queens, and males) within its web. These ants constitute 75% of prey captured by the spider. Juvenile L. mactans spiders have also been seen capturing the ants.[192][193] Other invertebrates that prey on red imported fire ants are earwigs (Labidura riparia) and tiger beetles (Cicindela punctulata).[192] Birds that eat these ants include the chimney swift (Chaetura pelagica), the eastern kingbird (Tyrannus tyrannus), and the eastern bobwhite (Colinus virginianus virginianus). The eastern bobwhite attacks these ants by digging out the mounds looking for young queens.[192] Red imported fire ants have been found in stomach contents inside of armadillos.[194]
Many species of ants have been observed attacking queens and killing them. Apparently, the venom of fire ant queens is chemically adapted to rapidly subdue offending competitor ants.[2] Predatory ants include: Ectatomma edentatum, Ephebomyrmex spp., Lasius neoniger, Pheidole spp., Pogonomyrmex badius, and Conomyrma insana, which is among the most significant.[192][195][196] C. insana ants are known to be effective predators against founding queens in studied areas of Northern Florida. The pressure of attacks initiated by C. insana increase over time, causing queens to exhibit different reactions, including escaping, concealment, or defence. Most queens that are attacked by these ants are ultimately killed.[195] Queens that are in groups have higher chances of survival than solitary queens if they are attacked by S. geminata.[197] Ants can attack queens on the ground and invade nests by stinging and dismembering them. Other ants such as P. porcula try to take the head and gaster, and C. clara invade in groups. Also, certain ants try to drag queens out of their nests by pulling on the antennae or legs.[196] Small, monomorphic ants rely on recruitment to kill queens and do not attack them until reinforcements arrive. Aside from killing the queen, some ants may steal the eggs for consumption or emit a repellent that is effective against red imported fire ants.[196] Certain ant species may raid colonies and destroy them.[198]
Parasites, pathogens and viruses
[edit]
Flies in the genus Pseudacteon (phorid flies) are known to be parasitic to ants. Some species within this genus, such as Pseudacteon tricuspis, have been introduced into the environment for the purpose of controlling the imported fire ant. These flies are parasitoids of the red imported fire ant in its native range in South America, and can be attracted through the ants' venom alkaloids.[199] One species, Pseudacteon obtusus, attacks the ant by landing on the posterior portion of the head and laying an egg. The location of the egg makes it impossible for the ant to successfully remove it.[200] The larvae migrate to the head, then develop by feeding on the hemolymph, muscle tissue, and nervous tissue. After about two weeks, they cause the ant's head to fall off by releasing an enzyme that dissolves the membrane attaching the head to its body. The fly pupates in the detached head capsule, emerging two weeks later.[200][201] P. tricuspis is another phorid fly that is a parasitoid to this species. Although parasitism pressures by these flies do not affect the ants' population density and activity, it has a small effect on a colony population.[202] The strepsipteran insect Caenocholax fenyesi is known to infect male ants of this species and attack the eggs,[203][204] and the mite Pyemotes tritici has been considered a potential biological agent against red imported fire ants, capable of parasitising every caste within the colony.[205] Bacteria, such as Wolbachia, has been found in the red imported fire ant; three different variants of the bacteria are known to infect the red imported fire ant. However, its effect on the ant is unknown.[206][207] Solenopsis daguerrei is a reproductive parasite to red imported fire ant colonies.[208]
A large variety of pathogens and nematodes also infect red imported fire ants. Pathogens include Myrmecomyces annellisae, Mattesia spp., Steinernema spp.,[209] a mermithid nematode,[210] Vairimorpha invictae, which can be transmitted via live larvae and pupae and dead adults[211][212] and Tetradonema solenopsis, which can be fatal to a large portion of a colony.[213] Individuals infected by Metarhizium anisopliae tend to perform trophallaxis more frequently and have an enhanced preference to quinine, an alkaloid substance.[214] Phorid flies with Kneallhazia solenopsae can serve as vectors in transmitting the disease to the ants.[215] Weakening the colony, infections from this disease are localised within the body fat, with spores only occurring in adult individuals.[216][217] The mortality of an infected colony tends to be greater in contrast to those that are healthy.[210] These ants are a host to Conidiobolus,[218] Myrmicinosporidium durum, and Beauveria bassiana, each of which are parasitic fungi. Infected individuals have spores all over their bodies and appear darker than usual.[219][220][221] The toxicity from antimicrobial property of volatiles produced by the ants can significantly reduce the germination rate of B. bassiana within the colony.[222]
A virus, S. invicta 1 (SINV-1), has been found in about 20% of fire ant fields, where it appears to cause the slow death of infected colonies. It has proven to be self-sustaining and transmissible. Once introduced, it can eliminate a colony within three months. Researchers believe the virus has potential as a viable biopesticide to control fire ants.[223][224][225] Two more viruses have also been discovered: S. invicta 2 (SINV-2) and S. invicta 3 (SINV-3). Polygynous colonies tend to face greater infections in contrast to monogynous colonies. Multiple virus infections can also occur.[226][227]
Lifecycle and reproduction
[edit]
Nuptial flight in red imported fire ants begins during the warmer seasons of the year (spring and summer), usually two days after rain. The time alates emerge and mate is between noon and 3:00 pm.[228][229] Nuptial flights recorded in North Florida have, on average, 690 female and male alates participating in a single flight.[230] Males are the first to leave the nest, and both sexes readily undertake flight with little to no preflight activity. However, workers swarm the mound excitedly stimulated by mandibular glands within the head of the alates.[231][232] As mounds do not have holes, workers form holes during nuptial flight as a way for the alates to emerge. This behaviour in workers, elicited by the pheromones, includes rapid running and back-and-forth movements, and increased aggression. Workers also cluster themselves around the alates as they climb up on vegetation, and in some cases, attempt to pull them back down before they take flight. Chemical cues from males and females during nuptial flight attract workers, but chemical cues released by workers do not attract other nestmates. It also induces alarm-recruitment behaviour in workers which results in a higher rate of alate retrieval.[231][233]
Males fly at higher elevations than females: captured males are usually 100 to 300 m (330 to 980 ft) above the surface, whereas the females are only 60 to 120 m (200 to 390 ft) above the surface. A nuptial flight takes place for roughly half an hour and females generally fly for less than 1.6 km (0.99 mi) before landing. About 95% of queens successfully mate and only mate once; some males may be infertile due to the testicular lobes failing to develop.[228][234][235][236][237][238] In polygyne colonies, males do not play a significant role and most are, therefore, sterile; one of the reasons for this is to avoid mating with other ant species. This also makes male mortality selective, which may affect the breeding system, mating success and, gene flow.[239][240] Ideal conditions for a nuptial flight to begin is when humidity levels are above 80% and when the soil temperature is above 18 °C (64 °F). Nuptial flights only occur when the ambient temperature is 24–32 °C (75–90 °F).[228]
Queens are often found 1–2.3 miles from the nest they flew from. Colony founding can be done by an individual or in groups, known as pleometrosis.[241] This joint effort of the co-foundresses contributes to the growth and survival of the incipient colony; nests founded by multiple queens begin the growth period with three times as many workers when compared to colonies founded by a single queen. Despite this, such associations are not always stable.[242][243][244][245] The emergence of the first workers instigates queen-queen and queen-worker fighting. In pleometrotic conditions, only one queen emerges victorious, whereas the queens that lost are subsequently killed by the workers.[246] The two factors that could affect the survival of individual queens are their relative fighting capabilities and their relative contribution to worker production. Size, an indicator of fighting capacity, positively correlates with survival rates. However, manipulation of the queen's relative contribution to worker production had no correlation with survival rate.[247]
A single queen lays around 10 to 15 eggs 24 hours after mating.[94] In established nests, a queen applies venom onto each egg that perhaps contains a signal calling for workers to move it.[248] These eggs remain unchanged in size for one week until they hatch into larvae. By this time, the queen will have laid about 75 to 125 more eggs. The larvae that hatch from their eggs are usually covered in their shell membranes for several days. The larvae can free their mouthparts from their shells using body movements, but still need assistance from workers with hatching. The larval stage is divided into four instars, as observed through the moulting stages. At the end of each moult, a piece of unknown material is seen connected to the exuviae if they are isolated from the workers. The larval stage lasts between six and 12 days before their bodies expand significantly and become pupae; the pupal stage lasts between nine and 16 days.[46][94]
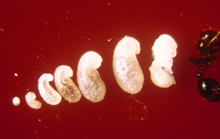
As soon as the first individuals reach the pupal stage, the queen ceases egg production until the first workers mature. This process takes two weeks to one month. The young larvae are fed oils which are regurgitated from her crop, as well as trophic eggs or secretions. She also feeds the young her wing muscles, providing the young with needed nutrients. The first generation of workers are always small because of the limit of nutrients needed for development. These workers are known as minims or nanitics, which burrow out of the queen's chamber and commence foraging for food needed for the colony. Mound construction also occurs at this time. Within a month after the first generation is born, larger workers (major workers) start to develop, and within six months, the mound will be noticeable, if viewed, and houses several thousand residents. A mature queen is capable of laying 1,500 eggs per day; all workers are sterile, so cannot reproduce.[94][249][250][251]
A colony can grow exceptionally fast. Colonies that housed 15–20 workers in May grew to over 7,000 by September. These colonies started to produce reproductive ants when they were a year old, and by the time they were two years old, they had over 25,000 workers. The population doubled to 50,000 when these colonies were three years old.[252] At maturity, a colony can house 100,000 to 250,000 individuals, but other reports suggest that colonies can hold more than 400,000.[d][253][254][255][256] Polygyne colonies have the potential to grow much larger than monogyne colonies.[254]
Several factors contribute to colony growth. Temperature plays a major role in colony growth and development; colony growth ceases below 24 °C and developmental time decreases from 55 days at temperatures of 24 °C to 23 days at 35 °C. Growth in established colonies only occurs at temperatures between 24 and 36 °C. Nanitic brood also develops far quicker than minor worker brood (around 35% faster), which is beneficial for founding colonies.[257] Colonies that have access to an unlimited amount of insect prey are known to grow substantially, but this growth is further accelerated if they are able to access plant resources colonised by hemipteran insects.[258] In incipient monogyne colonies where diploid males are produced, colony mortality rates are significantly high and colony growth is slow. In some cases, monogyne colonies experience 100% mortality rates in the early stages of development.[259]

The life expectancy of a worker ant depends on its size, although the overall average is around 62 days.[260] Minor workers are expected to live for about 30 to 60 days, whereas the larger workers live much longer. Larger workers, which have a life expectancy of 60 to 180 days, live 50–140% longer than their smaller counterparts, but this depends on the temperature.[94][261] However, workers kept in laboratory conditions have been known to live for 10 to 70 weeks (70 days to 490 days); the maximum recorded longevity of a worker is 97 weeks (or 679 days).[262] The queens live much longer than the workers, with a lifespan ranging from two years to nearly seven years.[94][262]
In colonies, queens are the only ants able to alter sex ratios which can be predicted. For example, queens originating from male-producing colonies tend to produce predominantly males, while queens that came from female-favoured sex ratio colonies tend to produce females.[263][264] Queens also exert control over the production of sexuals through pheromones that influence the behaviours of workers toward both male and female larvae.[265]
Monogyny and polygyny
[edit]There are two forms of society in the red imported fire ant: polygynous colonies and monogynous colonies.[266][267][266] Polygynous colonies differ substantially from monogynous colonies in social insects. The former experience reductions in queen fecundity, dispersal, longevity, and nestmate relatedness.[268][269][270][271] Polygynous queens are also less physogastric than monogynous queens and workers are smaller.[272][273] Understanding the mechanisms behind queen recruitment is integral to understanding how these differences in fitness are formed. It is unusual that the number of older queens in a colony does not influence new queen recruitment. Levels of queen pheromone, which appears to be related to queen number, play important roles in the regulation of reproduction. It would follow that workers would reject new queens when exposed to large quantities of this queen pheromone. Moreover, evidence supports the claim that queens in both populations enter nests at random, without any regard for the number of older queens present.[274] There is no correlation between the number of older queens and the number of newly recruited queens. Three hypotheses have been posited to explain the acceptance of multiple queens into established colonies: mutualism, kin selection, and parasitism.[275] The mutualism hypothesis states that cooperation leads to an increase in the personal fitness of older queens. However, this hypothesis is not consistent with the fact that increasing queen number decreases both queen production and queen longevity.[276] Kin selection also seems unlikely given that queens have been observed to cooperate under circumstances where they are statistically unrelated.[277] Therefore, queens experience no gain in personal fitness by allowing new queens into the colony. Parasitism of preexisting nests appears to be the best explanation of polygyny. One theory is that so many queens attempt to enter the colony that the workers get confused and inadvertently allow several queens to join it.[277]
Monogyne workers kill foreign queens and aggressively defend their territory. However, not all behaviours are universal, primarily because worker behaviours depend on the ecological context in which they develop, and the manipulation of worker genotypes can elicit change in behaviours. Therefore, behaviours of native populations can differ from those of introduced populations.[278] In a study to assess the aggressive behaviour of monogyne and polygyne red fire ant workers by studying interaction in neutral arenas, and to develop a reliable ethogram for readily distinguishing between monogyne and polygyne colonies of red imported fire ants in the field,[279] monogyne and polygyne workers discriminated between nestmates and foreigners as indicated by different behaviours ranging from tolerance to aggression. Monogyne ants always attacked foreign ants independently if they were from monogyne or polygyne colonies, whereas polygyne ants recognised, but did not attack, foreign polygyne ants, mainly by exhibiting postures similar to behaviours assumed after attacks by Pseudacteon phorids. Hostile versus warning behaviours were strongly dependent on the social structure of workers. Therefore, the behaviour toward foreign workers was a method of characterising monogyne and polygyne colonies.[280] Most colonies in the southeastern and south-central US tend to be monogynous.[245]
The monogynous red imported fire ant colony territorial area and the mound size are positively correlated, which, in turn, is regulated by the colony size (number and biomass of workers), distance from neighbouring colonies, prey density, and by the colony's collective competitive ability. In contrast, nestmate discrimination among polygynous colonies is more relaxed as workers tolerate conspecific ants alien to the colony, accept other heterozygote queens, and do not aggressively protect their territory from polygyne conspecifics.[281] These colonies might increase their reproductive output as a result of having many queens and the possibility of exploiting greater territories by means of cooperative recruitment and interconnected mounds.[282]
A social chromosome is present in the red imported fire ant. This chromosome can differentiate the social organisation of a colony carrying one of two variants of a supergene (B and b) which contains more than 600 genes. The social chromosome has often been compared to sexual chromosomes because they share similar genetic features[283] and they define colony phenotype in a similar way. For example, colonies exclusively carrying the B variant of this chromosome accept single BB queens, but colonies with both B and b variants will accept multiple Bb queens only.[284] Differences in another single gene can also determine whether the colony will have single or multiple queens.[285]
Relationship with other animals
[edit]Competition
[edit]
When polygyne forms invade areas where colonies have not yet been established, the diversity of native arthropods and vertebrates declines greatly.[286] This is evident as populations of isopods, mites and tumblebug scarabs decline significantly. They can also significantly alter the populations of many fly and beetle families, including: Calliphoridae, Histeridae, Muscidae, Sarcophagidae, Silphidae, and Staphylinidae. Despite this, one review found that certain insects may be unaffected by red imported fire ants; for example, the density of isopods decreases in red imported fire ant infested areas, but crickets of the genus Gryllus are unaffected. There are some cases where the diversity of certain insect and arthropod species increase in areas where red imported fire ants are present.[287][288][289] Red imported fire ants are important predators on cave invertebrates, some of which are endangered species. This includes harvestmen, pseudoscorpions, spiders, ground beetles, and pselaphid beetles. The biggest concern is not the ant itself, but the bait used to treat them because this can prove fatal. Stock Island tree snails (Orthalicus reses) are extinct in the wild; predation by red imported fire ants is believed to be the major factor in the snail's extinction.[287] Overall, red imported fire ants prefer specific arthropods to others, although they attack and kill any invertebrate that cannot defend itself or escape.[290] Arthropod biodiversity increases once red imported fire ant populations are either reduced or eradicated.[115][291]
Interactions between red imported fire ants and mammals have been rarely documented. However, deaths of live-trapped animals by red imported fire ants have been observed.[287][292][293] Mortality rates in eastern cottontail (Sylvilagus floridanus) young range from 33 to 75% because of red imported fire ants.[294] It is believed that red imported fire ants have a strong impact on many herpetofauna species; scientists have noted population declines in the Florida kingsnake (Lampropeltis getula floridana), and eggs and adults of the eastern fence lizard (Sceloporus undulatus) and six-lined racerunner (Aspidoscelis sexlineata) are a source of food.[295][296][297] Because of this, eastern fence lizards have adapted to have longer legs and new behaviours to escape the red imported fire ant.[298][299] Additionally, another lizard species, Sphaerodactylus macrolepis are also a target of the fire ants' and have developed tactics to fend them off, such as tail flicks.[300] Adult three-toed box turtles (Terrapene carolina triunguis), Houston toad (Anaxyrus houstonensis) juveniles, and American alligator (Alligator mississippiensis) hatchlings are also attacked and killed by these ants.[287][301][302][303] Despite this mostly-negative association, one study shows that red imported fire ants may be capable of impacting vector-borne disease transmissions by regulating tick populations and altering vector and host dynamics, thereby reducing transmission rates not only to animals, but to humans as well.[304]
Mortality rates have been well observed in birds; there have been instances where no young have survived to adulthood in areas with high fire ant density. Many birds including cliff nesting swallows, ducks, egrets, quail, and terns have been affected by red imported fire ants.[305] Ground nesting birds, particularly the least tern (Sterna antillarum), are vulnerable to fire ant attacks.[306][303] The impact of red imported fire ants on colonial breeding birds is especially severe; waterbirds can experience a mortality rate of 100%, although this factor was lower for early-nesting birds. Brood survival decreases in American cliff swallows (Petrochelidon pyrrhonota) if they are exposed to foraging workers.[307] Songbird nest survival decreases in areas with red imported fire ants present, but survival rates in white-eyed vireo (Vireo griseus) and black-capped vireo (Vireo atricapilla) nests increase from 10% to 31% and 7% to 13% whenever fire ants are not present or when they are unable to attack them.[308] Red imported fire ants may indirectly contribute to low brood survival in the Attwater's prairie chicken.[309] It was first thought that the ants were linked to the decline of overwintering birds such as the loggerhead shrike (Lanius ludovicianus), but a later study showed that ant eradication efforts using the pesticide Mirex, which was known to have toxic side effects, was largely to blame.[310][311]
Red imported fire ants are strong competitors with many ant species. They have managed to displace many native ants which has led to a number of ecological consequences.[312] However, studies show that these ants are not always superior competitors that suppress native ants. Habitat disturbance prior to their arrival, and recruitment limitations, are more plausible reasons why native ants are suppressed.[313] Between Tapinoma melanocephalum and Pheidole fervida, the red imported fire ant is stronger than both species but shows different levels of aggression. For example, they are less hostile towards T. melanocephalum in contrast to P. fervida. Mortality rates in T. melanocephalum and P. fervida when fighting with red imported fire ants are high, being 31.8% and 49.9% respectively. The mortality rate for red imported fire ant workers, however, is only 0.2% to 12%.[314] The imported crazy ant (Nylanderia fulva) exhibits greater dominance than the red imported fire ant and has been known to displace them in habitats where they encounter each other in.[315] Larger colonies of pavement ants (Tetramorium caespitum) can destroy red imported fire ant colonies, leading entomologists to conclude that this conflict between the two species may help impede the spread of the red imported fire ant.[316][317] Individuals infected by SINV-1 can be killed faster than healthy individuals by Monomorium chinense. This means that ants infected with SINV-1 are weaker than their healthy counterparts and more than likely will be eliminated by M. chinense. However, major workers, whether they are infected or not, are rarely killed.[318]
In areas it is native to, the red imported fire ant is still a dominant species and coexists with 28 ant species in gallery forest gaps and ten species in xerophytic forest grassland, winning most aggressive interactions with other ants. However, some ants can be co-dominant in areas where they coexist such as the Argentine ant, where they compete symmetrically.[319][320] Workers regularly engage in food competition with other ants, and can suppress the exploitation of food resources from honeydew-producing hemipterans (specifically from Phenacoccus solenopsis) from native ants; however, red imported fire ants are unable to eliminate T. melanocephalum completely although they consume a higher proportion of food. Instead, the two ants may peacefully coexist and share the honeydew.[321][322] When encountering neighboring fire ants, workers may take on death-feigning behaviours to avoid them with success. However, such behaviour is only seen in young workers, as older workers either flee or fight back when threatened.[323]
As mentioned, red imported fire ants and Argentine ants compete with each other. Mortality rates vary in different scenarios (i.e. mortality rates in colony confrontation is lower than those confronting each other in the field). Major workers can also withstand more injuries to their bodies, thus increasing the mortality rate of Argentine ants. For an Argentine ant colony to successfully wipe out a monogyne colony of 160,000 workers, the colony would need 396,800 workers. A colony that has reduced in size due to successful bait treatment are prone to predation by Argentine ants. The ants may play a vital role in removing weakened fire ant colonies, and they may also be important in slowing the spread of these fire ants, especially in heavily infested Argentine ant areas.[324] Despite this, Argentine ant populations in the southeastern United States have declined following the introduction of the red imported fire ant.[325]
Red imported fire ants may reduce butterfly numbers due to eggs and caterpillar predation[326][327][328] and have been linked to butterfly species declines including Schaus swallowtails (Papilio aristodemus).[329]
Mutualism
[edit]
Red imported fire ants have formed a relationship with an invasive mealybug, Phenacoccus solenopsis. Colony growth is known to increase if ants have access to resources produced by P. solenopsis, and the population density of these mealybugs is significantly higher in areas where red imported fire ants are present (the spread of P. solenopsis in areas it is not native is attributed to the presence of the ants). As well as that, the life expectancy and reproductive rate of P. solenopsis both increase. These mealybugs may even be transported by workers back to their nest.[330] Predation of P. solenopsis decreases due to tending workers interfering with the predation and parasitism by natural enemies. Mealybugs are preyed on by the lady beetle (Menochilus sexmaculatus) and are hosts of two parasites (Aenasius bambawalei and Acerophagus coccois) if there are no ants present, but this is different if ants are present. Lady beetles are less frequently seen on plants with red imported fire ants, and the rate of "mummy" nymphs is significantly lower.[330] This is clearly evident as scientists observed that cotton aphid (Aphis gossypii) populations, and the predation of sentinel bollworm eggs, increased in areas with red imported fire ant presence.[331][332][333] Red imported fire ants have developed a mutual relationship with another mealybug (Dysmicoccus morrisoni). The ants promote the colony growth of D. morrisoni through protection, covering the colonies up with debris and collecting the honeydew they secrete.[334]
Toxicology
[edit]The toxicology of red imported fire ant venom has been relatively well studied. The venom is important to the ant, enabling it to capture its prey and to defend itself. Some 14 million people are stung annually in the United States,[335] who may suffer reactions ranging from mild eczema to anaphylactic shock.[336] Most common reactions to fire ant stings are a burning sensation at the sting site, followed by urticaria and pustule formation.[337] Some victims may develop a more serious allergic reaction which can be life-threatening.[338][339] The venom is mainly composed of insoluble alkaloids with a minor watery phase[340] containing some 46 proteins[341] of which four are reported as allergens.[342][343] Effective immunotherapy is available.[344]
Relationship with humans
[edit]As pests
[edit]
Because of their notoriety and invasive behavior, red imported fire ants are considered pests. In the United States, the Food and Drug Administration estimates more than $5 billion is spent annually on medical treatment, damage, and control in infested areas.[345][346] In Texas alone, red imported fire ants caused $300 million in damages for livestock, wildlife and public health.[347] Approximately $36 per American household, and more than $250 million has been spent to control and eradicate red imported fire ants. Private agencies spend $25 to $40 million on pesticides annually.[94] Unsuccessful eradication in Australia may cost the economy billions in damages annually and, based on a Queensland government study, the estimated cost could reach $43 billion over 30 years.[348]
Red imported fire ants thrive in urban areas, especially in backyards, golf courses, parks, recreational areas, school grounds, and street verges, in which their presence may deter outdoor activities. If they enter homes or properties, they can harm pets if they are caged, penned, tied or cannot escape.[346][349] Nests can be built under pavements or even roads, as well as under driveways, foundations, lawns, edges of sidewalks, under patio slabs, in electrical boxes or near power lines. A colony can excavate huge quantities of soil, resulting in structural problems in driveways, pavings and walls, and can also cause the formation of potholes in roads.[94][346][350][351][352] Additional damage by mounds can be inflicted on trees, yard plants, and pipes; some structures may also collapse. Colonies may migrate into human homes after heavy rain to take refuge from saturated soil.[3][94]
Not only do they thrive in urban areas, red imported fire ants can damage equipment and infrastructure and impact business, land, and property values.[349] They are also attracted to electricity; electrically stimulated workers release venom alkaloids, alarm pheromones, and recruitment pheromones, which in return attracts more workers to the site.[353] As a result, red imported fire ants can destroy electrical equipment.[354] This is known as magnetism, where scientists have identified internal magnetic materials which may play a role in orientation behaviors.[355][356] They are known to chew through electrical insulation which causes damage to electric motors, irrigation lines, pumps, signal boxes, transformers, telephone exchanges, and other equipment.[346][357] Colonies aggregate near electrical fields and are capable of causing short circuits or interfering with switches and equipment such as air conditioners, computers, and water pumps. They are also known to infest airport landing areas and traffic lights.[357] In 2002, the farm gate value of decorative producing nurseries in Orange, Los Angeles, and San Diego counties was $1.3 billion. According to author Les Greenberg of the Center for Invasive Species Research, roughly 40% of crops are produced in these three counties, making it one of the major crops in these areas. “The value of California's entire nursery industry in 2002 was estimated at $2.6 billion” (Greenberg, 1). This business is already feeling the effects of quarantine laws that need pesticide soaking before imports can begin. Immediate costs include the spraying of insecticides to all plants in restricted zones.[358]
In agriculture
[edit]
The red imported fire ant is a significant agricultural pest in areas where it is not native. They are capable of damaging crops, and they threaten pastures and orchards.[349] The mounds themselves can destroy agricultural equipment such as irrigation systems and damage machinery during harvest time.[359] The ants are known to invade soybean crops causing lower yields, and could cause $156 million in losses for soybean crops in the southeastern United States.[94][360][361] Reports from Georgia and North Carolina claim that 16.8 to 49.1 kg/ha of soybeans could not be harvested due to interference from fire ant mounds; combine harvesters skipped over the mounds preventing the crops from being harvested, and farmers raised the cutter bars on their combine harvesters to avoid impacting the mounds.[362][363][364] The feeding behaviour of red imported fire ants can result in significant damage to many other crops, including: beans, cabbage, citrus, corn, cucumber, eggplant, okra, peanut, potato, sorghum, sunflower, and sweet potato. The ants also interfere with plants' root systems and feed on young growth.[94][365] Sometimes, colonies build mounds around or near the base of citrus trees chew off new growth and feed on the flowers or developing fruit. Citrus trees are often girdled or killed.[366]
Despite its pest status and notoriety, the red imported fire ant can be beneficial. The ant is an effective insect predator, so it may serve as a biological agent against other pest species, especially in sugarcane fields.[367] Pest insects the ant kills include: boll weevils (Anthonomus grandis) in cotton crops,[368][369][370] sugarcane borers (Diatraea saccharalis) in sugarcane fields,[371][372][373] horn flies (Haematobia irritans) in manure,[374] velvetbean caterpillars (Anticarsia gemmatalis) in soybeans, and whiteflies that are found in greenhouses.[375][376] A number of studies indicate that red imported fire ants do not interfere or attack insects of economic importance in cotton fields, which has resulted in many farmers in the southeastern regions of the United States viewing red imported fire ants as beneficial.[377] However, some scientists have suggested that the beneficial status of the red imported fire ant is hard to predict when geography, plant size, season, soil moisture and the use of insecticides are not considered. These factors may lower the efficacy of red imported fire ants as pest control agents. Another factor is that workers are indiscriminate and kill beneficial insects such as predators that eat horn flies and other pests in pastures and predators of aphids and scale insects.[377][378][379] They also reduce the efficacy of parasitic wasps against pest species by eating the larvae and pupae.[377][380]
Control
[edit]In comparison to other ant species like Anoplolepis gracilipes, which rapidly take over areas they have been contained in, red imported fire ants are fairly easy to control.[381] The first proposals to controlling the ant occurred in 1957 when the United States Congress authorised an eradication program using federal and state funding. Research on the ant and its biology has been continuous after the establishment of an eradication program, and many chemicals were used to eliminate them. However, scientists discovered that these insecticides were killing native fauna, and the Environmental Protection Agency subsequently outlawed them. Some scientists even questioned whether the ants were pests or not.[382][383] Today, it is unlikely that the red imported fire ant will be eradicated in areas such as the United States. Populations can be managed properly if an integrated approach is used.[384] Some scientists have considered using the ants' natural enemies against it; this includes Kneallhazia solenopsae and B. bassiana. Phorid flies have also been viewed as potential biological agents, as they can reduce foraging activity in red imported fire ants and affect population levels. However, they are unable to affect colony growth rate.[385] In addition, parasitic ants, parasitic wasps, mites, other pathogens, nematodes, and fungi have been considered to be potential biological agents.[386] Others suggest that populations can be maintained or reduced by manipulating several ecological factors.[387]

Several baits have been used to control populations. Mounds are destroyed in a matter of weeks if baits are used on them. Baits are considered to be effective and simple to use against red imported fire ants, in comparison to drenching, dusting, or fumigating. They are sprinkled onto the mound then the ants take and consume them.[388][389] Certain baits such as growth regulator baits, and boric acid-sucrose water baits, benefit native fauna and low concentrations are usually required to kill a colony.[390][391] Others baits used against red imported fire ants include Amdro, Ascend, hydramethylnon, indoxacarb, and Maxforce.[392]
Solid and liquid bait insecticides, if improperly applied in a location, may be moved through waterflow in to waterways. As snow melts and rainfall moves over and through the ground, the water picks up improperly applied insecticides and deposits them in to larger bodies of water such as rivers and wetlands, or percolates in to watersheds.
Researchers have also been experimenting with other methods, such as fumigation, injecting the mound, mound drenching, and surface dusting. Mound drenching entails pouring large volumes of toxic liquid into the mound. While this may affect a large portion of the mound, it is possible it may not reach the queen, thus preventing the destruction of the colony. Surface dusting is similar to mound drenching, except insecticides are applied on top of the mound and soak into the soil when wet. Mound injections use pressurising and injecting insecticides into the mound, but the queen may not be affected by this. Mounds can be eliminated effectively through fumigation. Broadcast treatment is sometimes used by spraying infected areas with agricultural equipment. Finally, spot treatment with insecticides can be used by drilling and injecting the mound with residual insecticide.[94][388]
Aside from chemical control, other methods that can be employed against these ants include mechanical and electrical devices. However, it is unknown whether or not these devices are effective. Ant-proofing can be effective against colonies nesting inside buildings by caulking and sealing cracks, which successfully suppresses the population outside the walls. Homeowners have used their own methods to remove mounds by pouring boiling water on them or igniting them with flammable liquids. Although these methods may be effective, they are not recommended because they can be harmful to humans and the environment.[94][388]
Notes
[edit]- ^ Although S. geminata appears in the cladogram, it is not a member of the species-group. It is only included to serve as an "attachment point"; to be precise, it is showing where the cladogram under investigation connects with the tree of life.[30]
- ^ Some sources suggest they first arrived in 1929.[105]
- ^ Red imported fire ants most likely reached New Orleans in Louisiana, as well, but the presence of Argentine ants there prevented the ants from settling and spreading.[106]
- ^ The population of a single colony can vary seasonally.[253]
References
[edit]Footnotes
[edit]- ^ Johnson, N.F. (19 December 2007). "Solenopsis invicta Buren". Hymenoptera Name Server version 1.5. Columbus, Ohio, USA: Ohio State University. Archived from the original on 20 September 2016. Retrieved 1 April 2015.
- ^ Jump up to: a b c Fox, Eduardo G.P.; Wu, Xiaoqing; Wang, Lei; Chen, Li; Lu, Yong-Yue; Xu, Yijuan (February 2019). "Queen venom isosolenopsin A delivers rapid incapacitation of fire ant competitors". Toxicon. 158: 77–83. Bibcode:2019Txcn..158...77F. doi:10.1016/j.toxicon.2018.11.428. PMID 30529381. S2CID 54481057.
- ^ Jump up to: a b c Greenberg, L.; Kabashima, J.N. (2014). "Pest Notes: Red Imported Fire Ant". Statewide Integrated Pest Management Program. Agriculture and Natural Resources, University of California. Retrieved 4 April 2016.
- ^ "Red Imported Fire Ant | National Invasive Species Information Center". www.invasivespeciesinfo.gov. Retrieved 23 August 2021.
- ^ Tschinkel 2006, p. vii.
- ^ Taber 2000, p. 12.
- ^ Bowersock, G.W.; Brown, P. (1999). Late Antiquity: a Guide to the Postclassical World (2nd ed.). Cambridge: Belknap Press of Harvard University Press. p. 284. ISBN 978-0-674-51173-6.
- ^ Starr, C.; Evers, C.; Starr, L. (2010). Biology: Concepts and Applications without Physiology (8th ed.). Belmont, California: Cengage Learning. p. 431. ISBN 978-0-538-73925-2.
- ^ Lewis, P.H. (24 July 1990). "Mighty Fire Ants March Out of the South". The New York Times. Retrieved 2 November 2016.
- ^ Vanderwoude, C.; Elson-Harris, M.; McNaught, M. K. "Solenopsis invicta (Family Formicidae)". Species Bank. Department of the Environment and Energy (Australia). Archived from the original on 21 September 2016.
- ^ Tschinkel 2006, pp. 13–14.
- ^ Drees, B.M. (2002). "Medical problems and treatment considerations for the red imported fire ant" (PDF). Texas Imported Fire Ant Research and Management Project. Texas A&M University. Retrieved 23 August 2016.
- ^ Booth, K.; Dhami, M. (2008). Red Imported Fire Ant (Solenopsis invicta): A review of the literature regarding the determination of colony age (PDF). Centre for Biodiversity and Biosecurity (Report). Ministry for Primary Industries: Biosecurity New Zealand. p. 2. Archived from the original (PDF) on 9 October 2016. Retrieved 29 November 2018.
- ^ Masterson, J. (2007). "Species Name: Solenopsis invicta". Smithsonian Marine Station. Archived from the original on 20 September 2016. Retrieved 2 April 2016.
- ^ Carmichael, A. (2005). "Red imported fire ant Solenopsis invicta Buren (Hymenoptera: Formicidae: Myrmicinae)". PaDIL (Australian Biosecurity). Archived from the original on 29 November 2020. Retrieved 2 April 2016.
- ^ Buhs 2005, p. 13.
- ^ Jump up to: a b Santschi, F. (1916). "Formicides sudaméricains nouveaux ou peu connus". Physis. 2: 365–399. doi:10.5281/ZENODO.14374.
- ^ Taber 2000, p. 25.
- ^ Jump up to: a b c Shattuck, S.O.; Porter, S.D.; W., D.P. (1999). "Case 3069. Solenopsis invicta Buren, 1972 (Insecta, Hymenoptera): proposed conservation of the specific name". Bulletin of Zoological Nomenclature. 56 (1): 27–30. doi:10.5962/bhl.part.23022. ISSN 0007-5167.
- ^ Creighton, W.S. (1930). "The new world species of the genus Solenopsis (Hymenop. Formicidae)" (PDF). Proceedings of the American Academy of Arts and Sciences. 66 (2): 39–152. doi:10.2307/20026320. JSTOR 20026320. Archived from the original (PDF) on 4 March 2016.
- ^ Wilson, E.O. (1952). "The Solenopsis saevissima complex in South America (Hymenoptera: Formicidae)". Memórias do Instituto Oswaldo Cruz. 50: 60–68. doi:10.1590/S0074-02761952000100003. ISSN 0074-0276. PMID 13012835.
- ^ Ettershank, G. (1966). "A generic revision of the world Myrmicinae related to Solenopsis and Pheidologeton (Hymenoptera : Formicidae)". Australian Journal of Zoology. 14 (1): 73–171. doi:10.1071/ZO9660073.
- ^ Kempf, W.W. (1972). "Catalogo abreviado das formigas da Regiao Neotropical". Studia Entomologica. New Series. 15: 3–344.
- ^ Jump up to: a b c d e f g h i j k l m n o Buren, W.F. (1972). "Revisionary studies on the taxonomy of the imported fire ants". Journal of the Georgia Entomological Society. 7: 1–26. doi:10.5281/zenodo.27055.
- ^ Jump up to: a b c d Trager, J.C. (1991). "A revision of the fire ants, Solenopsis geminata group (Hymenoptera: Formicidae: Myrmicinae)". Journal of the New York Entomological Society. 99 (2): 141–198. doi:10.5281/zenodo.24912. JSTOR 25009890.
- ^ Jump up to: a b Bolton, B. (1995). A New General Catalogue of the Ants of the World. Cambridge: Harvard University Press. pp. 388–391. ISBN 978-0-674-61514-4.
- ^ Taber 2000, p. 26.
- ^ International Commission on Zoological Nomenclature (2001). "Opinion 1976 Solenopsis invicta Buren, 1972 (Insecta, Hymenoptera): specific name conserved". Bulletin of Zoological Nomenclature. 58 (2): 156–157. ISSN 0007-5167.
- ^ Bolton, B. (2016). "Solenopsis invicta". AntCat. Retrieved 19 August 2016.
- ^ Jump up to: a b Tschinkel 2006, p. 14.
- ^ Jump up to: a b Ross, K.G.; Meer, R.K.V.; Fletcher, D.J.C.; Vargo, E.L. (1987). "Biochemical phenotypic and genetic studies of two introduced fire ants and their hybrid (Hymenoptera: Formicidae)". Evolution. 41 (2): 280–293. doi:10.2307/2409138. JSTOR 2409138. PMID 28568766.
- ^ Ross, K.G.; Vargo, E.L.; Fletcher, David J.C. (1987). "Comparative biochemical genetics of three fire ant species in North America, with special reference to the two social forms of Solenopsis invicta (Hymenoptera: Formicidae)". Evolution. 41 (5): 979–990. doi:10.2307/2409186. JSTOR 2409186. PMID 28563420.
- ^ Tschinkel 2006, p. 15–16.
- ^ Jump up to: a b Pitts, J.P.; McHugh, J.V.; Ross, K.G. (2005). "Cladistic analysis of the fire ants of the Solenopsis saevissima species-group (Hymenoptera: Formicidae)". Zoologica Scripta. 34 (5): 493–505. doi:10.1111/j.1463-6409.2005.00203.x. S2CID 85792334.
- ^ Tschinkel 2006, p. 503.
- ^ Pitts, J.P. (2002). A cladistic analysis of the Solenopsis saevissima species-group (Hymenoptera: Formicidae) (PDF) (Ph.D.). University of Georgia, Athens. pp. 42–43.
- ^ Taber 2000, p. 43.
- ^ Ross, K.G.; Shoemaker, D.D. (1997). "Nuclear and mitochondrial genetic structure in two social forms of the fire ant Solenopsis invicta: insights into transitions to an alternate social organization". Heredity. 78 (6): 590–602. doi:10.1038/hdy.1997.98. S2CID 19225329.
- ^ Krieger, M.J.B.; Ross, K.G.; Chang, C..W.Y.; Keller, L. (1999). "Frequency and origin of triploidy in the fire ant Solenopsis invicta". Heredity. 82 (2): 142–150. doi:10.1038/sj.hdy.6884600. S2CID 13144037.
- ^ Keller, L.; Ross, K.G. (1998). "Selfish genes: a green beard in the red fire ant" (PDF). Nature. 394 (6693): 573–575. Bibcode:1998Natur.394..573K. doi:10.1038/29064. S2CID 4310467. Archived from the original (PDF) on 20 July 2011. Retrieved 26 February 2011.
- ^ Grafen, A. (1998). "Evolutionary biology: Green beard as death warrant" (PDF). Nature. 394 (6693): 521–522. Bibcode:1998Natur.394..521G. doi:10.1038/28948. S2CID 28124873.
- ^ Wurm, Y.; Wang, J.; Riba-Grognuz, O.; Corona, M.; Nygaard, S.; Hunt, B.G.; Ingram, K.K.; Falquet, L.; Nipitwattanaphon, M.; Gotzek, D.; Dijkstra, M.B.; Oettler, J.; Comtesse, F.; Shih, C.-J.; Wu, W.-J.; Yang, C.-C.; Thomas, J.; Beaudoing, E.; Pradervand, S.; Flegel, V.; Cook, E.D.; Fabbretti, R.; Stockinger, H.; Long, L.; Farmerie, W.G.; Oakey, J.; Boomsma, J.J.; Pamilo, P.; Yi, S.V.; Heinze, J.; Goodisman, M. A.D.; Farinelli, L.; Harshman, K.; Hulo, N.; Cerutti, L.; Xenarios, I.; Shoemaker, D.; Keller, L. (2011). "The genome of the fire ant Solenopsis invicta". Proceedings of the National Academy of Sciences. 108 (14): 5679–5684. Bibcode:2011PNAS..108.5679W. doi:10.1073/pnas.1009690108. PMC 3078418. PMID 21282665.
- ^ Hedges, S.A. (1997). Moreland, D. (ed.). Handbook of Pest Control (8th ed.). Mallis Handbook and Technical Training Company. pp. 531–535.
- ^ Tschinkel 2006, p. 24.
- ^ Buhs 2005, p. 11.
- ^ Jump up to: a b c Petralia, R. S.; Vinson, S. B. (1979). "Developmental morphology of larvae and eggs of the Imported fire ant, Solenopsis invicta". Annals of the Entomological Society of America. 72 (4): 472–484. doi:10.1093/aesa/72.4.472.
- ^ Jump up to: a b Wheeler, G.C.; Wheeler, J. (1977). "Supplementary studies on ant larvae: Myrmicinae". Transactions of the American Entomological Society. 103: 581–602. doi:10.5281/zenodo.25100.
- ^ Petralia, R.S.; Sorensen, A.A.; Vinson, S.B. (1980). "The labial gland system of larvae of the imported fire ant, Solenopsis invicta Buren". Cell and Tissue Research. 206 (1): 145–156. doi:10.1007/BF00233615. PMID 6153574. S2CID 28786982.
- ^ O'Neal, J.; Markin, G.P. (1975). "The larval instars of the imported fire ant, Solenopsis invicta Buren (Hymenoptera: Formicidae)". Journal of the Kansas Entomological Society. 48 (2): 141–151. JSTOR 25082731.
- ^ Jump up to: a b c d Fox, Eduardo G P; Solis, Daniel Russ; Rossi, Mônica Lanzoni; Delabie, Jacques Hubert Charles; de Souza, Rodrigo Fernando; Bueno, Odair Correa (2012). "Comparative Immature Morphology of Brazilian Fire Ants (Hymenoptera: Formicidae: Solenopsis)". Psyche: A Journal of Entomology. 2012: 1–10. doi:10.1155/2012/183284. hdl:11449/73193.
- ^ Mirenda, J.T.; Vinson, S.B. (1981). "Division of labour and specification of castes in the red imported fire ant Solenopsis invicta Buren". Animal Behaviour. 29 (2): 410–420. doi:10.1016/S0003-3472(81)80100-5. S2CID 53199246.
- ^ Schmid-Hempel 1998, p. 144.
- ^ Hölldobler & Wilson 1990, p. 311.
- ^ Hölldobler & Wilson 1990, p. 318.
- ^ Jump up to: a b Tschinkel, W.R. (1988). "Colony growth and the ontogeny of worker polymorphism in the fire ant, Solenopsis invicta". Behavioral Ecology and Sociobiology. 22 (2): 103–115. doi:10.1007/BF00303545. S2CID 8455126.
- ^ Jump up to: a b c d e Tschinkel, W.R.; Mikheyev, A.S.; Storz, S.R. (2003). "Allometry of workers of the fire ant, Solenopsis invicta". Journal of Insect Science. 3 (2): 2. doi:10.1673/031.003.0201. PMC 524642. PMID 15841219.
- ^ Porter, S.D.; Tschinkel, W.R. (1985). "Fire ant polymorphism: the ergonomics of brood production". Behavioral Ecology and Sociobiology. 16 (4): 323–336. doi:10.1007/BF00295545. JSTOR 4599785. S2CID 5967997.
- ^ Jump up to: a b Vogt, J.T.; Appel, A.G. (2000). "Discontinuous gas exchange in the fire ant, Solenopsis invicta Buren: Caste differences and temperature effects". Journal of Insect Physiology. 46 (4): 403–416. Bibcode:2000JInsP..46..403V. doi:10.1016/S0022-1910(99)00123-7. PMID 12770204.
- ^ Jump up to: a b Klowden, M.J. (2007). Physiological Systems in Insects. Amsterdam; Boston: Elsevier/Academic Press. pp. 357–383, 433–449. ISBN 978-0-12-369493-5.
- ^ Hill, R.W.; Wyse, G.A.; Anderson, M. (2012). Animal Physiology (3rd ed.). Sunderland, Massachusetts: Sinauer Associates. pp. 612–614. ISBN 978-0-87893-662-5.
- ^ Jump up to: a b c d e f Vinson, S.B. (1983). "The physiology of the imported fire ant revisited". The Florida Entomologist. 66 (1): 126–139. doi:10.2307/3494559. JSTOR 3494559.
- ^ Petralia, R.S.; Williams, H.J.; Vinson, S.B. (1982). "The hindgut ultrastructure, and excretory products of larvae of the imported fire ant, Solenopsis invicta Buren". Insectes Sociaux. 29 (2): 332–345. doi:10.1007/BF02228760. S2CID 7324976.
- ^ Fletcher, D.J.C.; Blum, M.S. (1981). "Pheromonal control of dealation and oogenesis in virgin queen fire ants". Science. 212 (4490): 73–75. Bibcode:1981Sci...212...73F. doi:10.1126/science.212.4490.73. PMID 17747633.
- ^ Barker, J.F. (1978). "Neuroendocrine regulation of oocyte maturation in the imported fire ant Solenopsis invicta". General and Comparative Endocrinology. 35 (3): 234–237. doi:10.1016/0016-6480(78)90067-9. PMID 689357.
- ^ Jones, R.G.; Davis, W.L.; Hung, A.C.F.; Vinson, S.B. (1978). "Insemination-induced histolysis of the flight musculature in fire ants (Solenopsis, spp.): An ultrastructural study". American Journal of Anatomy. 151 (4): 603–610. doi:10.1002/aja.1001510411. PMID 645619.
- ^ Toom, P.M.; Johnson, C.P.; Cupp, E.W. (1976). "Utilization of body reserves during preoviposition activity by Solenopsis invicta". Annals of the Entomological Society of America. 69 (1): 145–148. doi:10.1093/aesa/69.1.145.
- ^ Jump up to: a b Vinson, S.B.; Phillips, S.A.; Williams, H.J. (1980). "The function of the post-pharyngeal glands of the red imported fire ant, Solenopsis invicta buren". Journal of Insect Physiology. 26 (9): 645–650. Bibcode:1980JInsP..26..645V. doi:10.1016/0022-1910(80)90035-9.
- ^ Vander Meer, R.K.; Lofgren, C.S. (1989). "Biochemical and behavioral evidence foe hybridization between fire ants, Solenopsis invicta and Solenopsis richteri (Hymenoptera: Formicidae)". Journal of Chemical Ecology. 15 (6): 1757–1765. Bibcode:1989JCEco..15.1757V. doi:10.1007/BF01012263. PMID 24272179. S2CID 23144401.
- ^ Jump up to: a b Vander Meer, R.K.; Glancey, B.M.; Lofgren, C.S.; Glover, A.; Tumlinson, J.H.; Rocca, J. (1980). "The poison sac of red imported fire ant queens: source of a pheromone attractant". Annals of the Entomological Society of America. 73 (5): 609–612. doi:10.1093/aesa/73.5.609.
- ^ Choi, M.Y.; Vander Meer, R.K.; Renou, M. (2012). "Ant trail pheromone biosynthesis is triggered by a neuropeptide hormone". PLOS ONE. 7 (11): e50400. Bibcode:2012PLoSO...750400C. doi:10.1371/journal.pone.0050400. PMC 3511524. PMID 23226278.
- ^ Elzen, G.W (1986). "Oxygen consumption and water loss in the imported fire ant Solenopsis invicta Buren". Comparative Biochemistry and Physiology A. 84 (1): 13–17. doi:10.1016/0300-9629(86)90035-6.
- ^ Chen, J.; Rashid, T.; Feng, G.; Hughes, W. (2014). "A comparative study between Solenopsis invicta and Solenopsis richteri on tolerance to heat and desiccation stresses". PLOS ONE. 9 (6): e96842. Bibcode:2014PLoSO...996842C. doi:10.1371/journal.pone.0096842. PMC 4051589. PMID 24915009.
- ^ Boyles, J.G.; Aubrey, D.P.; Hickman, C.R.; Murray, K.L.; Timpone, J.C.; Ops, C.H. (2009). "Variation in physiological response of red imported fire ants (Solenopsis invicta) to small-scale thermal heterogeneity". Journal of Thermal Biology. 34 (2): 81–84. Bibcode:2009JTBio..34...81B. doi:10.1016/j.jtherbio.2008.10.005.
- ^ Jump up to: a b Porter, S.D.; Tschinkel, W.R. (1993). "Fire ant thermal preferences: behavioral control of growth and metabolism". Behavioral Ecology and Sociobiology. 32 (5). doi:10.1007/BF00183787. S2CID 9840395.
- ^ Jump up to: a b Vogt, J.T.; Appel, A.G. (1999). "Standard metabolic rate of the fire ant, Solenopsis invicta Buren: effects of temperature, mass, and caste". Journal of Insect Physiology. 45 (7): 655–666. Bibcode:1999JInsP..45..655V. doi:10.1016/S0022-1910(99)00036-0. PMID 12770351.
- ^ Jump up to: a b c d e f g h i j k Wetterer, J.K. (2013). "Exotic spread of Solenopsis invicta (Hymenoptera: Formicidae) beyond North America". Sociobiology. 60: 50–55. doi:10.13102/sociobiology.v60i1.50-55.
- ^ Jump up to: a b Allen, G. E.; Buren, W. F.; Williams, R. N.; Menezes, M. D.; Whitcomb, W. H. (1974). "The red imported fire ant, Solenopsis invicta; distribution and habitat in Mato Grosso, Brazil". Annals of the Entomological Society of America. 67 (1): 43–46. doi:10.1093/aesa/67.1.43.
- ^ Jump up to: a b Mescher, M.C.; Ross, K.G.; Shoemaker, D.D.; Keller, L.; Krieger, M.J. B. (2003). "Distribution of the two social forms of the fire ant Solenopsis invicta (Hymenoptera: Formicidae) in the native South American range" (PDF). Annals of the Entomological Society of America. 96 (6): 810–817. doi:10.1603/0013-8746(2003)096[0810:DOTTSF]2.0.CO;2. S2CID 41810551.[permanent dead link]
- ^ Jump up to: a b Noordijk, J. (2010). "A risk analysis for fire ants in the Netherlands". Leiden: Stichting European Invertebrate Survey. p. 15. Archived from the original on 20 October 2018. Retrieved 5 April 2016.
- ^ Jump up to: a b c d e Buren, W.F.; Allen, G.E.; Whitcomb, W.H.; Lennartz, F.E.; Williams, R.N. (1974). "Zoogeography of the imported fire ants". Journal of the New York Entomological Society. 82 (2): 113–124. JSTOR 25008914.
- ^ Caldera, E.J.; Ross, K.G.; DeHeer, Ch.J.; Shoemaker, D. D.. (2008). "Putative native source of the invasive fire ant Solenopsis invicta in the USA". Biological Invasions. 10 (8): 1457–1479. Bibcode:2008BiInv..10.1457C. doi:10.1007/s10530-008-9219-0. S2CID 4471306.
- ^ Pesquero, M.A.; Dias, A.M.P.M (2011). "Geographical transition zone of Solenopsis fire ants (Hymenoptera: Formicidae) and Pseudacteon fly parasitoids (Diptera: Phoridae) in the state of São Paulo, Brazil". Neotropical Entomology. 40 (6): 647–652. doi:10.1590/S1519-566X2011000600003. PMID 23939270.
- ^ Jump up to: a b c d "Species: Solenopsis invicta Buren, 1972". AntWeb. The California Academy of Sciences. Retrieved 5 April 2016.
- ^ Wild, A.L. (2007). "A catalogue of the ants of Paraguay (Hymenoptera: Formicidae)" (PDF). Zootaxa. 1622: 1–55. doi:10.11646/zootaxa.1622.1.1. ISSN 1175-5334.
- ^ Santschi, F. (1923). "Solenopsis et autres fourmis néotropicales". Revue Suisse de Zoologie. 30 (8): 245–273. doi:10.5281/ZENODO.14217.
- ^ Ahrens, M.E.; Ross, K.G.; Shoemaker, D.D. (2005). "Phylogeographic structure of the fire ant Solenopsis invicta in its native South American range: roles of natural barriers and habitat connectivity" (PDF). Evolution. 59 (8): 1733–1743. doi:10.1111/j.0014-3820.2005.tb01822.x. PMID 16329243. S2CID 34788394.[permanent dead link]
- ^ Jump up to: a b c d e f "Solenopsis invicta (red imported fire ant)". Invasive Species Compendium. CABI. 2014. Retrieved 12 April 2016.
- ^ Taber 2000, p. 28.
- ^ Cokendolpher, J.C.; Phillips, S.A. Jr. (1989). "Rate of spread of the red imported fire ant, Solenopsis invicta (Hymenoptera: Formicidae), in Texas". The Southwestern Naturalist. 34 (3): 443–449. doi:10.2307/3672182. JSTOR 3672182.
- ^ "Solenopsis invicta (insect)". Global Invasive Species Database. Invasive Species Specialist Group. 2010. Archived from the original on 3 March 2016. Retrieved 2 April 2016.
- ^ Buhs 2005, p. 12.
- ^ Jump up to: a b c d e Kemp, S.F.; deShazo, R.D.; Moffitt, J.E.; Williams, D.F.; Buhner, W.A. (2000). "Expanding habitat of the imported fire ant (Solenopsis invicta): a public health concern". The Journal of Allergy and Clinical Immunology. 105 (4): 683–691. doi:10.1067/mai.2000.105707. PMID 10756216. S2CID 8280947.
- ^ Desforges, J.F.; deShazo, R.D.; Butcher, B.T.; Banks, W.A. (1990). "Reactions to the stings of the imported fire ant". New England Journal of Medicine. 323 (7): 462–466. doi:10.1056/NEJM199008163230707. PMID 2197555.
- ^ Jump up to: a b c d e f g h i j k l m n o Collins, L.; Scheffrahn, R.H. (2001). "Red imported fire ant, Solenopsis invicta Buren". UF/IFAS Featured Creatures. University of Florida. Archived from the original on 21 September 2016. Retrieved 13 April 2016.
- ^ "Ant Colonies and Social Structure | Terminix". www.terminix.com. Retrieved 19 November 2019.
- ^ Hubbard, M.D.; Cunningham, W.G. (1977). "Orientation of mounds in the ant Solenopsis invicta (Hymenoptera, Formicidae, Myrmicinae)". Insectes Sociaux. 24 (1): 3–7. doi:10.1007/BF02223276. S2CID 5851856.
- ^ Penick, C.A.; Tschinkel, W. R. (2008). "Thermoregulatory brood transport in the fire ant, Solenopsis invicta". Insectes Sociaux. 55 (2): 176–182. doi:10.1007/s00040-008-0987-4. S2CID 15284230.
- ^ Cassill, D.; Tschinkel, W.R.; Vinson, S.B. (2002). "Nest complexity, group size and brood rearing in the fire ant, Solenopsis invicta". Insectes Sociaux. 49 (2): 158–163. doi:10.1007/s00040-002-8296-9. hdl:10806/3290. S2CID 13938249.
- ^ Taber 2000, p. 29.
- ^ Taber 2000, p. 31.
- ^ "100 of the World's Worst Invasive Alien Species". Global Invasive Species Database. Invasive Species Specialist Group. Retrieved 26 January 2014.
- ^ Mooallem, J. (5 December 2013). "There's a Reason They Call Them 'Crazy Ants'". The New York Times. Retrieved 23 August 2016.
- ^ Fountain, H. (8 December 2008). "Fire Ants Win Out Through Land Changes, Not a Better Build". The New York Times. Retrieved 31 October 2016.
- ^ King, J.R.; Tschinkel, W.R. (2008). "Experimental evidence that human impacts drive fire ant invasions and ecological change". Proceedings of the National Academy of Sciences. 105 (51): 20339–20343. Bibcode:2008PNAS..10520339K. doi:10.1073/pnas.0809423105. PMC 2629336. PMID 19064909.
- ^ Capinera 2008, p. 3116.
- ^ Buhs 2005, p. 20.
- ^ Wilcove, D.S.; Rothstein, D.; Dubow, J.; Phillips, A.; Losos, E. (1998). "Quantifying threats to imperiled species in the United States" (PDF). BioScience. 48 (8): 607–615. doi:10.2307/1313420. JSTOR 1313420. S2CID 7168138. Archived from the original (PDF) on 12 August 2011.
- ^ Tschinkel 2006, p. 28.
- ^ Buhs 2005, p. 9.
- ^ Fountain, H. (8 July 2008). "Tracing an Ant Invasion to a Handful of Queens". The New York Times. Retrieved 2 November 2016.
- ^ Ross, K.G; Shoemaker, D.D. (2008). "Estimation of the number of founders of an invasive pest insect population: the fire ant Solenopsis invicta in the USA". Proceedings of the Royal Society B: Biological Sciences. 275 (1648): 2231–2240. doi:10.1098/rspb.2008.0412. PMC 2603238. PMID 18577505.
- ^ Tschinkel 2006, p. 25.
- ^ Buhs 2005, p. 23.
- ^ Adkins, H.G. (1970). "The imported fire ant in the Southern United States". Annals of the Association of American Geographers. 60 (3): 578–592. doi:10.1111/j.1467-8306.1970.tb00742.x.
- ^ Jump up to: a b Epperson, D.M.; Allen, C.R. (2010). "Red imported fire ant impacts on upland arthropods in southern Mississippi". The American Midland Naturalist. 163 (1): 54–63. doi:10.1674/0003-0031-163.1.54. S2CID 12992133.
- ^ Callcott, A.-M.; Collins, H.L. (1996). "Invasion and range expansion of imported fire ants (Hymenoptera: Formicidae) in North America from 1918–1995". The Florida Entomologist. 79 (2): 240–251. doi:10.2307/3495821. JSTOR 3495821.
- ^ Korzukhin, M.D.; Porter, S.D.; Thompson, L.C.; Wiley, S. (2001). "Modeling temperature-dependent range limits for the fire ant Solenopsis invicta (Hymenoptera: Formicidae) in the United States" (PDF). Environmental Entomology. 30 (4): 645–655. doi:10.1603/0046-225X-30.4.645. S2CID 53065271.
- ^ Callcott, A.-M.; Oi, D.H.; Collins, H.L.; Williams, D.F.; Lockley, T.C. (2000). "Seasonal studies of an isolated red imported fire ant (Hymenoptera: Formicidae) population in eastern Tennessee" (PDF). Environmental Entomology. 29 (4): 788–794. doi:10.1603/0046-225X-29.4.788. S2CID 53073021.
- ^ Pimm, S.L.; Bartell, D.P. (1980). "Statistical model for predicting range expansion of the red imported fire ant, Solenopsis invicta, in Texas". Environmental Entomology. 9 (5): 653–658. doi:10.1093/ee/9.5.653.
- ^ Capinera 2008, pp. 2034–2035.
- ^ Morrison, L.W.; Korzukhin, M.D.; Porter, S.D. (2005). "Predicted range expansion of the invasive fire ant, Solenopsis invicta, in the eastern United States based on the VEMAP global warming scenario". Diversity and Distributions. 11 (3): 199–204. Bibcode:2005DivDi..11..199M. doi:10.1111/j.1366-9516.2005.00142.x. S2CID 46089176.
- ^ Jump up to: а б Моррисон, LW; Портер, SD; Дэниэлс, Э.; Корзухин, доктор медицинских наук (2004). «Потенциальное глобальное расширение ареала инвазивного огненного муравья Solenopsis invicta » (PDF) . Биологические инвазии . 6 (2): 183–191. Бибкод : 2004BiInv...6..183M . дои : 10.1023/B:BINV.0000022135.96042.90 . S2CID 9973667 . [ постоянная мертвая ссылка ]
- ^ Бухс 2005 , с. 5.
- ^ Уайлдер, Шон М.; Холуэй, Дэвид А.; Суарес, Эндрю В.; ЛеБрун, Эдвард Г.; Юбэнкс, Микки Д. (20 декабря 2011 г.). «Межконтинентальные различия в использовании ресурсов показывают важность мутуализма при вторжениях огненных муравьев» . Труды Национальной академии наук . 108 (51). Национальная академия наук : 20639–20644. Бибкод : 2011PNAS..10820639W . дои : 10.1073/pnas.1115263108 . ISSN 0027-8424 . ПМЦ 3251143 . ПМИД 22143788 .
- ^ «Красный импортный огненный муравей Solenopsis invicta » . Департамент окружающей среды . Правительство Австралии. Архивировано из оригинала 21 сентября 2016 года . Проверено 21 марта 2015 г.
- ^ МакКаббин, Кентукки; Вайнер, Дж. М. (2002). «Огненные муравьи в Австралии: новая медицинская и экологическая опасность». Медицинский журнал Австралии . 176 (11): 518–519. дои : 10.5694/j.1326-5377.2002.tb04547.x . ПМИД 12064981 . S2CID 43444266 .
- ^ Хеншоу, Майкл Т; Кунцманн, Н.; Вандервуд, К.; Санетра, М.; Крозье, Р.Х. (2005). «Популяционная генетика и история завезенного огненного муравья Solenopsis invicta Buren (Hymenoptera: Formicidae) в Австралию». Австралийский журнал энтомологии . 44 (1): 37–44. дои : 10.1111/j.1440-6055.2005.00421.x .
- ^ Кондон, М. (27 июля 2013 г.). «Квинсленд начал войну против вторжения огненных муравьев, но 12 лет спустя они все еще на марше» . Курьерская почта . Проверено 30 декабря 2014 г.
- ^ Уиллис, П. (22 февраля 2007 г.). «Обновление огненного муравья» . Новости АВС . Архивировано из оригинала 21 сентября 2016 года . Проверено 4 мая 2015 г.
- ^ «Национальная программа по уничтожению огненных муравьев» . Департамент первичной промышленности . Правительство Квинсленда. 2007. Архивировано из оригинала 21 октября 2009 года.
- ^ «Импорт красных импортированных огненных муравьев Solenopsis invicta Buren 1972 - профиль» . Управление окружающей среды и наследия . Правительство Нового Южного Уэльса. 2014. Архивировано из оригинала 21 сентября 2016 года . Проверено 5 мая 2015 г.
- ^ Кридон, К. (4 декабря 2014 г.). «Гонка на время: карантин в Сиднее, вспышка красных огненных муравьев» . 9 Новости . Сидней, Австралия. Архивировано из оригинала 21 сентября 2016 года . Проверено 6 декабря 2014 г.
- ^ Австралийское агентство Associated Press (25 февраля 2016 г.). «Американские огненные муравьи обнаружены в аэропорту Брисбена» . Новости СБС . Архивировано из оригинала 21 сентября 2016 года . Проверено 23 августа 2016 г. .
- ^ Кансдейл, Доминик (7 августа 2023 г.). «Огненные муравьи выигрывают битву против искоренения из-за «бюрократических проволочек», - говорит Совет по инвазивным видам» . Австралийская радиовещательная корпорация .
- ^ Молони, С.; Вандервуд, К. (2002). «Красные импортированные огненные муравьи: угроза дикой природе восточной Австралии?». Экологический менеджмент и восстановление . 3 (3): 167–175. Бибкод : 2002ЭкоМР...3..167М . дои : 10.1046/j.1442-8903.2002.t01-1-00109.x . S2CID 54926916 .
- ^ Раджагопал, Т.; Севаркодийоне, ИП; Секар, М. (2005). «Индекс видового богатства, разнообразия и сходства муравьев в пяти выбранных местонахождениях Саттур Талук» . Индийский журнал экологического образования . 5 :7–12.
- ^ На, JPS; Ли, Калифорния (2001). «Ключ для идентификации распространенных городских муравьев-вредителей в Малайзии» (PDF) . Тропическая биомедицина . 18 (1): 1–17.
- ^ Куо, К.К. (2008). Борьба с красными инвазивными огненными муравьями и плодовыми мухами: опыт Тайваня (PDF) . Тайбэй, Тайвань: Центр технологий пищевых продуктов и удобрений (FFTC). п. 1.
- ^ Бхарти, Х.; Генар, Б.; Бхарти, М.; Экономо, EP (2016). «Обновленный контрольный список муравьев Индии с указанием их конкретного распространения в индийских штатах (Hymenoptera, Formicidae)» . ZooKeys (551): 1–83. Бибкод : 2016ZooK..551....1B . дои : 10.3897/zookeys.551.6767 . ПМЦ 4741291 . ПМИД 26877665 .
- ^ Чинкель 2006 , с. 72.
- ^ Ван, Л.; Юнъюэ, Л.; Ицзюань, X.; Линг, З. (2013). «Текущий статус исследований Solenopsis invicta Buren (Hymenoptera: Formicidae) в материковом Китае» (PDF) . Азиатская мирмекология . 5 : 125–137. ISSN 1985-1944 гг .
- ^ Цзэн, Л.; Лу, ГГ; Он, XF; Чжан, WQ; Лян, GW (2005). «Идентификация завезенного красного огненного муравья Solenopsis invicta, вторгшегося в материковый Китай и заразившего Учуань, Гуандун». Китайский бюллетень энтомологии . 42 (2): 144–148. ISSN 0452-8255 .
- ^ Чжан, Р.; Ли, Ю.; Лю, Н.; Портер, SD (2007). «Обзор красного завезенного огненного муравья (Hymenoptera: Formicidae) в материковый Китай» . Энтомолог Флориды . 90 (4): 723–731. doi : 10.1653/0015-4040(2007)90[723:aootri]2.0.co;2 . S2CID 73532042 . Проверено 12 апреля 2016 г.
- ^ Сазерст, RW; Мэйвальд, Г. (2005). «Климатическая модель красного завезенного огненного муравья Solenopsis invicta Buren (Hymenoptera: Formicidae): последствия для вторжения в новые регионы, особенно в Океанию» . Экологическая энтомология . 34 (2): 317–335. дои : 10.1603/0046-225X-34.2.317 . S2CID 85951260 .
- ^ Менчетти, Маттиа; Скифани, Энрико; Аликата, Антонио; Кардадор, Лаура; Сбрега, Элизабетта; Торо-Дельгадо, Эрик; Вила, Роджер (сентябрь 2023 г.). «Инвазивный муравей Solenopsis invicta обосновался в Европе» . Современная биология . 33 (17): Р896–Р897. Бибкод : 2023CBio...33R.896M . дои : 10.1016/j.cub.2023.07.036 . ПМИД 37699343 . S2CID 261688271 .
- ^ Бюрен, ВФ (1982). «Красный импортированный огненный муравей теперь в Пуэрто-Рико». Энтомолог Флориды . 65 (1): 188–189. дои : 10.2307/3494163 . JSTOR 3494163 .
- ^ Веттерер, Дж. К.; Снеллинг, Р.Р. (2006). «Красный завезенный огненный муравей Solenopsis invicta на Виргинских островах (Hymenoptera: Formicidae)» (PDF) . Энтомолог Флориды . 89 (4): 431–434. doi : 10.1653/0015-4040(2006)89[431:TRIFAS]2.0.CO;2 . S2CID 85996855 .
- ^ Дейруп, М.; Дэвис, Л.; Бакнер, С. (1998). Состав муравьиной фауны трех Багамских островов (PDF) . Сан-Сальвадор: материалы 7-го симпозиума по естественной истории Багамских островов. п. 27. ISBN 978-0-935909-66-1 .
- ^ Дэвис, ЛР; Вандер Меер, РК; Портер, SD (2001). «Красные завезенные огненные муравьи расширяют свой ареал по всей Вест-Индии» (PDF) . Энтомолог Флориды . 84 (4): 735. дои : 10.2307/3496416 . JSTOR 3496416 . Архивировано из оригинала (PDF) 25 января 2012 года . Проверено 12 апреля 2016 г.
- ^ Аскунс, MS; Ян, К.-К.; Оки, Дж.; Калькатерра, Л.; Ву, В.-Дж.; Ши, К.-Дж.; Гуде, Ж.; Росс, КГ; Шумейкер, Д. (2011). «Глобальная история вторжения огненного муравья Solenopsis invicta ». Наука . 331 (6020): 1066–1068. Бибкод : 2011Sci...331.1066A . дои : 10.1126/science.1198734 . ПМИД 21350177 . S2CID 28149214 .
- ^ Млот, Нью-Джерси; Крейг, AT; Ху, Д.Л. (2011). «Огненные муравьи сами собираются в водонепроницаемые плоты, чтобы пережить наводнения» . Труды Национальной академии наук . 108 (19): 7669–7673. Бибкод : 2011PNAS..108.7669M . дои : 10.1073/pnas.1016658108 . ПМК 3093451 . ПМИД 21518911 .
- ^ Моррилл, WL (1974). «Распространение красных завезенных огненных муравьев водой». Энтомолог Флориды . 57 (1): 39–42. дои : 10.2307/3493830 . JSTOR 3493830 .
- ^ Хёллдоблер и Уилсон 1990 , с. 171.
- ^ Jump up to: а б Адамс, Би Джей; Хупер-Буи, LM; Стрекер, Р.М.; О'Брайен, DM (2011). «Формирование плота красным огненным муравьем Solenopsis invicta » . Журнал науки о насекомых . 11 (171): 171. дои : 10.1673/031.011.17101 . ПМЦ 3462402 . ПМИД 22950473 .
- ^ Хейт, КЛ (2006). «Защитная способность огненного муравья Solenopsis invicta повышается во время сплава колонии». Общество насекомых . 53 (1): 32–36. дои : 10.1007/s00040-005-0832-y . S2CID 24420242 .
- ^ Ховард, DF; Чинкель, WR (1976). «Аспекты некрофорного поведения красного импортированного огненного муравья Solenopsis invicta » (PDF) . Поведение . 56 (1): 157–178. дои : 10.1163/156853976x00334 . ISSN 0005-7959 .
- ^ Цю, Х.-Л.; Лу, Л.-Х.; Ши, К.-Х.; Ту, К.-К.; Лин, Т.; Он, Ю.-Р. (2015). «Дифференциальное некрофорное поведение муравья Solenopsis invicta по отношению к зараженным грибком трупам рабочих и куколок». Бюллетень энтомологических исследований . 105 (5): 607–614. дои : 10.1017/S0007485315000528 . ПМИД 26082426 . S2CID 25475122 .
- ^ Смит, CW; Фредериксен, Р.А. (2000). Сорго: происхождение, история, технология и производство . Нью-Йорк, Нью-Йорк: Джон Уайли и сыновья. п. 456. ИСБН 978-0-471-24237-6 .
- ^ Зеттлер, Дж. А.; Спира, ТП; Аллен, CR (2001). «Муравьи и семена муравьев: могут ли завезенные красные огненные муравьи испортить отношения?» . Биологическая консервация . 101 (2): 249–253. Бибкод : 2001BCons.101..249Z . дои : 10.1016/S0006-3207(01)00074-X .
- ^ Камберленд, штат Массачусетс; Киркман, Л.К. (2013). «Влияние завезенного красного огненного муравья на судьбу семян в экосистеме длиннолистной сосны». Экология растений . 214 (5): 717–724. Бибкод : 2013PlEco.214..717C . дои : 10.1007/s11258-013-0201-2 . S2CID 2648579 .
- ^ Маркин, врач общей практики; О'Нил, Дж.; Диллер, Дж. (1975). «Кормовые туннели красного завезенного огненного муравья Solenopsis invicta (Hymenoptera: Formicidae)». Журнал Канзасского энтомологического общества . 48 (1): 83–89. JSTOR 25082717 .
- ^ Jump up to: а б с д Портер, SD; Чинкель, WR (1987). «Собирательство Solenopsis invicta (Hymenoptera: Formicidae): влияние погоды и сезона» . Экологическая энтомология . 16 (3): 802–808. дои : 10.1093/ee/16.3.802 .
- ^ Шоулер, AT; Кнаус, РМ; Рейган, Т.Э. (1989). «Территориальность поиска завезенного огненного муравья Solenopsis invicta Buren в сургартростнике, определенная нейтронно-активационным анализом». Общество насекомых . 36 (3): 235–239. дои : 10.1007/BF02226306 . S2CID 36832531 .
- ^ Гравиш, Н.; Монаенкова Д.; Гудисман, MAD; Гольдман, Д.И. (2013). «Подъем, падение и застревание во время передвижения муравьев в замкнутом пространстве» . Труды Национальной академии наук . 110 (24): 9746–9751. arXiv : 1305.5860 . Бибкод : 2013PNAS..110.9746G . дои : 10.1073/pnas.1302428110 . ПМЦ 3683784 . ПМИД 23690589 .
- ^ Jump up to: а б Ицзюань, X.; Юнъюэ, Л.; Линг, З.; Гуангвэнь, Л. (2007). «Собирательное поведение и пополнение красного импортированного огненного муравья Solenopsis invicta Buren в типичных местах обитания Южного Китая». Акта Экологика Синика . 27 (3): 855–860. Бибкод : 2007AcEcS..27..855Y . дои : 10.1016/S1872-2032(07)60022-5 .
- ^ Чинкель, WR (2011). «Организация питания огненного муравья Solenopsis invicta » . Журнал науки о насекомых . 11 (26): 26. дои : 10.1673/031.011.0126 . ПМК 3391925 . ПМИД 21529150 .
- ^ Jump up to: а б с д Вандер Меер, РК (1983). «Полухимикаты и красный импортированный огненный муравей ( Solenopsis invicta Buren) (Hymenoptera: Formicidae)». Энтомолог Флориды . 66 (1): 139–161. дои : 10.2307/3494560 . JSTOR 3494560 .
- ^ Уильямс, HJ; Стрэнд, MR; Винсон, С.Б. (1981). «Следовый феромон красного завезенного огненного муравья Solenopsis invicta (Buren)». Эксперименты . 37 (11): 1159–1160. дои : 10.1007/BF01989893 . S2CID 46572483 .
- ^ Вандер Меер, РК; Альварес, Ф.; Лофгрен, CS (1988). «Выделение феромона рекрутирования следа Solenopsis invicta ». Журнал химической экологии . 14 (3): 825–838. Бибкод : 1988JCEco..14..825V . дои : 10.1007/BF01018776 . ПМИД 24276134 . S2CID 32388563 .
- ^ Барлин, MR; Блюм, М.С.; Бранд, ММ (1976). «Феромоны огненных муравьев: анализ видовой специфичности после газохроматографического фракционирования». Журнал физиологии насекомых . 22 (6): 839–844. Бибкод : 1976JInsP..22..839B . дои : 10.1016/0022-1910(76)90253-5 .
- ^ Капинера 2008 , с. 3445.
- ^ Табер 2000 , стр. 41.
- ^ Вандер Меер, РК; Престон, Калифорния; Цой, М.-Ю. (2010). «Выделение компонента феромона тревоги пиразин из огненного муравья Solenopsis invicta ». Журнал химической экологии . 36 (2): 163–170. Бибкод : 2010JCEco..36..163В . дои : 10.1007/s10886-010-9743-0 . ПМИД 20145982 . S2CID 10622781 .
- ^ Jump up to: а б Хаббард, доктор медицины (1974). «Влияние материала гнезда и запаха колонии на копание муравья Solenopsis invicta (Hymenoptera: Formicidae)». Энтомологическое общество Джорджии . 9 : 127–132.
- ^ Обин, Мартин С. (1986). «Признаки распознавания гнездовья в лабораторных и полевых колониях Solenopsis invicta Buren (Hymenoptera: Formicidae)». Журнал химической экологии . 12 (9): 1965–1975. Бибкод : 1986JCEco..12.1965O . дои : 10.1007/BF01041858 . ПМИД 24305971 . S2CID 19517580 .
- ^ Обин, М.С.; Вандер Меер, РК (1989). «Распознавание товарищей по гнезду у огненных муравьев ( Solenopsis invicta Buren). Маркируют ли королевы рабочих?». Этология . 80 (1–4): 255–264. Бибкод : 1989Этол..80..255О . дои : 10.1111/j.1439-0310.1989.tb00744.x .
- ^ Вандер Меер, РК; Престон, Калифорния; Хефец, А. (2008). «Королева регулирует уровень биогенных аминов и распознавание партнеров по гнезду у рабочих огненного муравья Solenopsis invicta ». Naturwissenschaften . 95 (12): 1155–1158. Бибкод : 2008NW.....95.1155V . дои : 10.1007/s00114-008-0432-6 . ПМИД 18704354 . S2CID 7640153 .
- ^ Обин, М.С.; Вандер Меер, РК (1988). «Источники сигналов распознавания товарищей по гнезду у завезенного огненного муравья Solenopsis invicta Buren (Hymenoptera: Formicidae)». Поведение животных . 36 (5): 1361–1370. дои : 10.1016/S0003-3472(88)80205-7 . S2CID 53205306 .
- ^ Jump up to: а б Табер 2000 , стр. 37.
- ^ Табер 2000 , стр. 38.
- ^ Теннант, Луизиана; Портер, SD (1991). «Сравнение рациона двух видов огненных муравьев (Hymenoptera: Formicidae): твердые и жидкие компоненты». Журнал энтомологической науки . 26 (4): 450–465. дои : 10.18474/0749-8004-26.4.450 . ISSN 0749-8004 .
- ^ Jump up to: а б Портер, SD (1989). «Влияние диеты на рост колоний лабораторных огненных муравьев (Hymenoptera: Formicidae)». Журнал Канзасского энтомологического общества . 62 (2): 288–291. JSTOR 25085088 .
- ^ Спир, Дж.; Варго, THE; Пулим, С.; Чанг, YZ (1993). «Предпочтения огненных муравьев Solenopsis invicta и S. geminata (Hymenoptera: Formicidae) к аминокислотным и сахарным компонентам внефлоральных нектаров». Экологическая энтомология . 22 (2): 411–417. дои : 10.1093/да/22.2.411 . S2CID 17682096 .
- ^ Ланца, Дж. (1991). «Реакция огненных муравьев (Formicidae: Solenopsis invicta и S.gerninata ) на искусственные нектары с аминокислотами» . Экологическая энтомология . 16 (2): 203–210. Бибкод : 1991EcoEn..16..203L . дои : 10.1111/j.1365-2311.1991.tb00210.x . S2CID 84907757 .
- ^ Фогт, Дж.Т.; Грэнтэм, РА; Корбетт, Э.; Райс, ЮАР; Райт, RE (2002). «Пищевые привычки Solenopsis invicta (Hymenoptera: Formicidae) в четырех средах обитания в Оклахоме» . Экологическая энтомология . 31 (1): 47–53. дои : 10.1603/0046-225X-31.1.47 . S2CID 85586177 .
- ^ Jump up to: а б Кассилл, Д.Л.; Чинкель, WR (1999). «Регулирование диеты огненного муравья Solenopsis invicta » (PDF) . Журнал поведения насекомых . 12 (3): 307–328. дои : 10.1023/А:1020835304713 . S2CID 18581840 .
- ^ Ховард, DF; Чинкель, WR (1980). «Влияние размера колонии и голодания на поток пищи у огненного муравья Solenopsis invicta (Hymenoptera: Formicidae)». Поведенческая экология и социобиология . 7 (4): 293–300. дои : 10.1007/BF00300670 . JSTOR 4599344 . S2CID 18507664 .
- ^ Соренсен, А.А.; Винсон, С.Б. (1981). «Количественные исследования распределения пищи в лабораторных колониях завезенного огненного муравья Solenopsis invicta Buren». Общество насекомых . 28 (2): 129–160. дои : 10.1007/BF02223701 . S2CID 28896246 .
- ^ Гаяхан, Г.Г.; Чинкель, WR (2008). «Огненные муравьи, Solenopsis invicta , высушите и сохраните кусочки насекомых для дальнейшего использования» . Журнал науки о насекомых . 8 (39): 39. дои : 10.1673/031.008.3901 . ПМК 3127378 .
- ^ Табер 2000 , стр. 39.
- ^ Кассилл, Д.Л.; Чинкель, WR (1995). «Распределение жидкой пищи личинкам посредством трофаллаксиса в колониях огненного муравья Solenopsis invicta ». Поведение животных . 50 (3): 801–813. дои : 10.1016/0003-3472(95)80140-5 . S2CID 18395869 .
- ^ Jump up to: а б с д и Уиткомб, Вашингтон; Бхаткар, А.; Никерсон, Дж. К. (1973). «Хищники королев Solenopsis invicta до успешного создания колонии». Экологическая энтомология . 2 (6): 1101–1103. дои : 10.1093/ee/2.6.1101 .
- ^ Ниффелер, М.; Дин, округ Колумбия; Стерлинг, WL (1988). «Южный паук-черная вдова, Latrodectus mactans (Araneae, Theridiidae), как хищник красного завезенного огненного муравья Solenopsis invicta (Hymenoptera, Formicidae) на хлопковых полях Техаса». Журнал прикладной энтомологии . 106 (1–5): 52–57. дои : 10.1111/j.1439-0418.1988.tb00563.x . S2CID 83876695 .
- ^ Дейруп, М.; Трагер, Дж. (1986). «Муравьи биологической станции Арчболд, округ Хайлендс, Флорида (Hymenoptera: Formicidae)» . Энтомолог Флориды . 69 (1): 206–228. дои : 10.2307/3494763 . JSTOR 3494763 .
- ^ Jump up to: а б Никерсон, Дж. К.; Уиткомб, Вашингтон; Бхаткар, АП; Нейвс, Массачусетс (1975). «Хищничество на маток-основательниц Solenopsis invicta рабочих Conomyrma insana ». Энтомолог Флориды . 58 (2): 75–82. дои : 10.2307/3493384 . JSTOR 3493384 .
- ^ Jump up to: а б с Николс, Би Джей; Сайты, RW (1991). «Муравьи-хищники королев-основательниц Solenopsis invicta (Hymenoptera: Formicidae) в центральном Техасе» . Экологическая энтомология . 20 (4): 1024–1029. дои : 10.1093/ee/20.4.1024 .
- ^ Джером, Калифорния; Макиннес, Д.А.; Адамс, ЕС (1998). «Групповая защита маток-основателей колоний огненного муравья Solenopsis invicta » . Поведенческая экология . 9 (3): 301–308. дои : 10.1093/beheco/9.3.301 .
- ^ Рао, Аша; Винсон, С.Б. (2004). «Способность местных муравьев уничтожать небольшие колонии Solenopsis invicta (Hymenoptera: Formicidae)» . Экологическая энтомология . 33 (3): 587–598. дои : 10.1603/0046-225X-33.3.587 . S2CID 85657533 .
- ^ Чен, Ли; Шарма, КР; Фадамиро, Хай (2009). «Алкалоиды яда огненных муравьев действуют как ключевые аттрактанты для паразитической форидной мухи Pseudacteon tricuspis (Diptera: Phoridae)». Naturwissenschaften . 96 (12): 1421–1429. Бибкод : 2009NW.....96.1421C . дои : 10.1007/s00114-009-0598-6 . ПМИД 19714317 . S2CID 24205012 .
- ^ Jump up to: а б Уильямс, DF; Бэнкс, Вашингтон (1987). « Pseudacteon obtusus (Diptera: Phoridae) атакует Solenopsis invicta (Hymenoptera: Formicidae) в Бразилии» . Психика: журнал энтомологии . 94 (1–2): 9–13. дои : 10.1155/1987/85157 .
- ^ Эренберг, Р. (18 сентября 2009 г.). «Яд привлекает обезглавливающих мух» . Новости науки . Архивировано из оригинала 22 сентября 2016 года . Проверено 30 декабря 2014 г. (требуется подписка)
- ^ Моррисон, LW; Портер, SD (2005). «Тестирование воздействия на популяционном уровне интродуцированных мух Pseudacteon tricuspis , форидных паразитоидов огненных муравьев Solenopsis invicta ». Биологический контроль . 33 (1): 9–19. Бибкод : 2005БиолС..33....9М . CiteSeerX 10.1.1.542.4377 . doi : 10.1016/j.biocontrol.2005.01.004 .
- ^ Шмид-Хемпель 1998 , с. 94.
- ^ Катиритамби, Дж.; Джонстон, Дж. С. (1992). «Стилопизация Solenopsis invicta (Hymenoptera: Formicidae) Caenocholax fenyesi (Strepsiptera: Myrmecolacidae) в Техасе». Анналы Энтомологического общества Америки . 85 (3): 293–297. дои : 10.1093/aesa/85.3.293 .
- ^ Брюс, Вашингтон; ЛеКато, GL (1980). « Pyemotes tritici : потенциальный новый агент биологической борьбы с завезенным красным огненным муравьем Solenopsis invicta (Acari: Pyemotidae)». Международный журнал акарологии . 6 (4): 271–274. Бибкод : 1980IJAca...6..271B . дои : 10.1080/01647958008683230 .
- ^ Аренс, Мэн; Шумейкер, Д. (2005). «Эволюционная история инфекций Wolbachia у огненного муравья Solenopsis invicta » . Эволюционная биология BMC . 5 (1): 35. Бибкод : 2005BMCEE...5...35A . дои : 10.1186/1471-2148-5-35 . ПМК 1175846 . ПМИД 15927071 .
- ^ Шмид-Хемпель 1998 , с. 34.
- ^ Капинера 2008 , с. 188.
- ^ Шмид-Хемпель 1998 , с. 59.
- ^ Jump up to: а б Бриано, Дж.; Калькатерра, Л.; Вароне, Л. (2012). «Огненные муравьи ( Solenopsis spp.) и их естественные враги на юге Южной Америки» . Психика: журнал энтомологии . 2012 : 1–19. дои : 10.1155/2012/198084 .
- ^ Жувеназ, ДП; Эллис, Э.А. (1986). « Vairimorpha invictae N. Sp. (Microspora: Microsporida), паразит красного завезенного огненного муравья Solenopsis invicta Buren (Hymenoptera: Formicidae)». Журнал протозоологии . 33 (4): 457–461. дои : 10.1111/j.1550-7408.1986.tb05641.x .
- ^ Ой, ДХ; Бриано, JA; Валлес, С.М.; Уильямс, Д.Ф. (2005). «Передача инфекций Vairimorpha invictae (Microsporidia: Burenellidae) между колониями красных завезенных огненных муравьев (Hymenoptera: Formicidae)». Журнал патологии беспозвоночных . 88 (2): 108–115. Бибкод : 2005JInvP..88..108O . дои : 10.1016/j.jip.2004.11.006 . ПМИД 15766927 .
- ^ Никль, WR; Жувеназ, ДП (1987). « Tetradonema solenopsis n. sp. (Nematoda: Tetradonematidae), паразитирующая на красном огненном муравье Solenopsis invicta Buren из Бразилии» . Журнал нематологии . 19 (3): 311–313. ПМК 2618658 . ПМИД 19290149 .
- ^ Цю, Х.-Л.; Лу, Л.-Х.; Залуцкий, депутат парламента; Он, Ю.-Р. (2016). « Инфекция Metarhizium anisopliae изменяет пищевое и трофаллактическое поведение муравья Solenopsis invicta » (PDF) . Журнал патологии беспозвоночных . 138 : 24–29. Бибкод : 2016JInvP.138...24Q . дои : 10.1016/j.jip.2016.05.005 . ПМИД 27234423 .
- ^ Дарем, С. (2010). «Коллекции паразитов ARS помогают в исследованиях и диагностике» . Служба сельскохозяйственных исследований . Министерство сельского хозяйства США . Проверено 25 августа 2014 г.
- ^ Нелл, JD; Аллен, GE; Опасность, Э.И. (1977). «Исследование Thelohania solenopsae n. sp. (Microsporida: Protozoa) с помощью светового и электронного микроскопа у красного завезенного огненного муравья Solenopsis invicta ». Журнал патологии беспозвоночных . 29 (2): 192–200. Бибкод : 1977JInvP..29..192K . дои : 10.1016/0022-2011(77)90193-8 . ПМИД 850074 .
- ^ «Юг борется с огненными муравьями с помощью возбудителя» . CNN . 1 июня 1998 года . Проверено 1 ноября 2016 г.
- ^ Шмид-Хемпель 1998 , с. 295.
- ^ Перейра, РМ (2004). «Встреча Myrmicinosporidium durum у красного завезенного огненного муравья, Solenopsis invicta и других новых муравьев-хозяев на востоке США». Журнал патологии беспозвоночных . 86 (1–2): 38–44. Бибкод : 2004JInvP..86...38P . дои : 10.1016/j.jip.2004.03.005 . ПМИД 15145249 .
- ^ Шмид-Хемпель 1998 , с. 44.
- ^ Чинкель 2006 , с. 632.
- ^ Ван, Л.; Эллиотт, Б.; Джин, X.; Цзэн, Л.; Чен, Дж. (2015). «Противомикробные свойства летучих веществ гнезд красных завезенных огненных муравьев Solenopsis invicta (перепончатокрылые: formicidae)». Naturwissenschaften . 102 (11–12): 66. Бибкод : 2015SciNa.102...66W . дои : 10.1007/s00114-015-1316-1 . ПМИД 26467352 . S2CID 1650319 .
- ^ Валлес, С.М.; Стронг, Калифорния; Данг, премьер-министр; Хантер, ВБ; Перейра, РМ; Ой, ДХ; Шапиро, AM; Уильямс, Д.Ф. (2004). «Пикорноподобный вирус красного импортированного огненного муравья Solenopsis invicta : первоначальное открытие, последовательность генома и характеристика» (PDF) . Вирусология . 328 (1): 151–157. дои : 10.1016/j.virol.2004.07.016 . ПМИД 15380366 .
- ^ Валлес, С.М.; Стронг, Калифорния; Ой, ДХ; Портер, SD; Перейра, РМ; Вандер Меер, РК; Хасимото, Ю.; Хупер-Буи, LM; Санчес-Арройо, Х.; Дэвис, Т.; Карпакакунджарам, В.; Вейл, К.М.; Фадд Грэм, LC; Бриано, JA; Калькатерра, Луизиана; Гилберт, Ле; Уорд, Р.; Уорд, К.; Оливер, Дж.Б.; Танигучи, Г.; Томпсон, округ Колумбия (2007). «Фенология, распространение и специфичность хозяина вируса Solenopsis invicta -1». Журнал патологии беспозвоночных . 96 (1): 18–27. Бибкод : 2007JInvP..96...18V . дои : 10.1016/j.jip.2007.02.006 . ПМИД 17412359 .
- ^ «Огненные муравьи, возможно, встретили свою пару» . CNN . Ассошиэйтед Пресс. 7 мая 2007 г. Архивировано из оригинала 13 мая 2007 г.
- ^ Валлес, С.М.; Хасимото, Ю. (2009). «Выделение и характеристика вируса Solenopsis invicta 3, нового РНК-вируса с положительной цепью, заражающего красного завезенного огненного муравья Solenopsis invicta ». Вирусология . 388 (2): 354–61. дои : 10.1016/j.virol.2009.03.028 . ПМИД 19403154 .
- ^ Аллен, К.; Валлес, С.М.; Стронг, Калифорния (2011). «Множественные вирусные инфекции встречаются у отдельных полигинных и моногинных Solenopsis invicta муравьев » . Журнал патологии беспозвоночных . 107 (2): 107–111. Бибкод : 2011JInvP.107..107A . дои : 10.1016/j.jip.2011.03.005 . ПМИД 21439294 .
- ^ Jump up to: а б с Трагер 1988 , с. 419.
- ^ Бухс 2005 , с. 21.
- ^ Моррилл, WL (1974). «Производство и полет крылатых красных импортных огненных муравьев». Экологическая энтомология . 3 (2): 265–271. дои : 10.1093/ee/3.2.265 .
- ^ Jump up to: а б Алонсо, Ле; Вандер Меер, РК (1997). «Источник крылатых возбуждающих феромонов у красного завезенного огненного муравья Solenopsis invicta (Hymenoptera: Formicidae)». Журнал поведения насекомых . 10 (4): 541–555. Бибкод : 1997JIBeh..10..541A . дои : 10.1007/BF02765376 . S2CID 12295884 .
- ^ Маркин, врач общей практики; Дилье, Дж. Х.; Хилл, СО; Блюм, М.С.; Герман, HR (1971). «Брачный полет и дальность полета завезенного огненного муравья Solenopsis saevissima richteri (Hymenoptera: Formicidae)». Журнал Энтомологического общества Джорджии . 6 (3): 145–156.
- ^ Обин, М.С.; Вандер Меер, РК (1994). «Алатные семиохимические вещества влияют на поведение рабочих во время брачных полетов огненных муравьев» (PDF) . Журнал энтомологической науки . 29 (1): 143–151. дои : 10.18474/0749-8004-29.1.143 .
- ^ Хунг, АКФ; Винсон, SB; Саммерлин, JW (1974). «Мужская стерильность у красного завезенного огненного муравья Solenopsis invicta ». Анналы Энтомологического общества Америки . 67 (6): 909–912. дои : 10.1093/aesa/67.6.909 .
- ^ Трагер 1988 , с. 422.
- ^ Капинера 2008 , с. 3444.
- ^ Капинера 2008 , с. 3112.
- ^ Хёллдоблер и Уилсон 1990 , с. 156.
- ^ Фриц, Дж.Н.; Вандер Меер, РК; Престон, Калифорния (2006). «Выборочная смертность самцов красного завезенного огненного муравья Solenopsis invicta » . Генетика . 173 (1): 207–213. doi : 10.1534/genetics.106.056358 . ПМЦ 1461442 . ПМИД 16489215 .
- ^ Хёллдоблер и Уилсон 1990 , с. 145.
- ^ Чинкель, В. (1992). «Набеги на выводок и динамика численности основанных и зарождающихся колоний огненного муравья Solenopsis invicta » . Экологическая энтомология . 17 (2): 179–188. Бибкод : 1992EcoEn..17..179T . дои : 10.1111/j.1365-2311.1992.tb01176.x . S2CID 85060765 .
- ^ Гудисман, М.; Росс, К. (1999). «Набор королевы в популяции огненного муравья Solenopsis invicta, состоящей из нескольких маток ». Поведенческая экология . 10 (4): 428–435. дои : 10.1093/beheco/10.4.428 .
- ^ Чинкель, WR; Ховард, Д.Ф. (1983). «Колония, основанная в результате плеометроза у огненного муравья Solenopsis invicta » (PDF) . Поведенческая экология и социобиология . 12 (2): 103–113. дои : 10.1007/BF00343200 . S2CID 3017347 .
- ^ Трагер 1988 , с. 217.
- ^ Jump up to: а б Трагер 1988 , с. 218.
- ^ Манфредини, Ф.; Риба-Грогнуз, О.; Вурм, Ю.; Келлер, Л.; Шумейкер, Д.; Грозингер, CM ; Цуцуи, Северная Дакота (2013). «Социогеномика сотрудничества и конфликта при создании колонии огненного муравья Solenopsis invicta » . ПЛОС Генетика . 9 (8): e1003633. дои : 10.1371/journal.pgen.1003633 . ПМЦ 3738511 . ПМИД 23950725 .
- ^ Бернаскони, Дж.; Келлер, Л. (1996). «Репродуктивные конфликты в кооперативных объединениях королев огненных муравьев ( Solenopsis invicta )» (PDF) . Труды Королевского общества B: Биологические науки . 263 (1369): 509–513. Бибкод : 1996РСПСБ.263..509Б . дои : 10.1098/rspb.1996.0077 . S2CID 84276777 .
- ^ Табер 2000 , стр. 42.
- ^ Дебели, Мичиган; Абухейф, Э. (2016). «Моделирование эволюционных переходов у социальных насекомых» . электронная жизнь . 5 : е12721. дои : 10.7554/eLife.12721 . ПМЦ 4744194 . ПМИД 26780668 .
- ^ Хёллдоблер и Уилсон 1990 , с. 157.
- ^ Хёллдоблер и Уилсон 1990 , с. 168.
- ^ Маркин, врач общей практики; Дилье, Дж. Х.; Коллинз, Х.Л. (1973). «Рост и развитие колоний красного завозного огненного муравья Solenopsis invicta ». Анналы Энтомологического общества Америки . 66 (4): 803–808. дои : 10.1093/aesa/66.4.803 .
- ^ Jump up to: а б Хёльдоблер и Уилсон 1990 , с. 163.
- ^ Jump up to: а б Табер 2000 , стр. 32.
- ^ Шмид-Хемпель 1998 , с. 10.
- ^ Шмид-Хемпель 1998 , с. 16.
- ^ Портер, SD (1988). «Влияние температуры на рост колонии и скорость развития муравья Solenopsis invicta » . Журнал физиологии насекомых . 34 (12): 1127–1133. Бибкод : 1988JInsP..34.1127P . дои : 10.1016/0022-1910(88)90215-6 .
- ^ Хелмс, КР; Винсон, С.Б. (2008). «Растительные ресурсы и рост колоний инвазивных муравьев: важность полужесткокрылых, производящих медвяную росу, в переносе углеводов через трофические уровни» . Экологическая энтомология . 37 (2): 487–493. дои : 10.1093/ee/37.2.487 . ПМИД 18419921 .
- ^ Росс, КГ; Флетчер, DJC (1986). «Производство диплоидных самцов — важный фактор смертности колоний огненных муравьев Solenopsis invicta (Hymenoptera: Formicidae)» . Поведенческая экология и социобиология . 19 (4): 283–291. дои : 10.1007/BF00300643 . S2CID 25932018 .
- ^ Ван, Л.; Сюй, Ю.; Ди, З.; Ренер, Б.М. (2013). «Как групповое взаимодействие и его разрыв влияют на продолжительность жизни?» (PDF) . Библиотека Корнельского университета : 1–28. arXiv : 1304.2935 . Бибкод : 2013arXiv1304.2935W . S2CID 14848627 . Архивировано из оригинала (PDF) 3 ноября 2016 года.
- ^ Калаби, П.; Портер, SD (1989). «Продолжительность жизни рабочих у огненного муравья Solenopsis invicta : эргономические соображения о корреляции между температурой, размером и скоростью метаболизма» . Журнал физиологии насекомых . 35 (8): 643–649. Бибкод : 1989JInsP..35..643C . дои : 10.1016/0022-1910(89)90127-3 .
- ^ Jump up to: а б Хёльдоблер и Уилсон 1990 , с. 170.
- ^ Дэвис, Северная Каролина; Кребс, младший; Вест, ЮАР (2012). Введение в поведенческую онкологию (4-е изд.). Чичестер: Уайли-Блэквелл. п. 385. ИСБН 978-1-4051-1416-5 .
- ^ Пассера, Л.; Арон, С.; Варго, Эль; Келлер, Л. (2001). «Королевский контроль над соотношением полов у огненных муравьев» (PDF) . Наука . 293 (5533): 1308–1310. Бибкод : 2001Sci...293.1308P . дои : 10.1126/science.1062076 . ПМИД 11509728 . S2CID 7483158 .
- ^ Варго, Эль; Флетчер, DJC (1986). «Свидетельства контроля феромонной королевы над производством мужских и женских половых органов у огненного муравья Solenopsis invicta ». Журнал сравнительной физиологии А. 159 (6): 741–749. дои : 10.1007/BF00603727 . ISSN 1432-1351 . S2CID 1165247 .
- ^ Jump up to: а б Чинкель 2006 , с. 95.
- ^ Хёллдоблер и Уилсон 1990 , с. 187.
- ^ Гудисман, Майкл А.Д.; Росс, КГ (1998). «Тестирование моделей набора королевы с использованием ядерных и митохондриальных маркеров у огненного муравья Solenopsis invicta » (PDF) . Эволюция . 52 (5): 1416–1422. дои : 10.2307/2411311 . JSTOR 2411311 . ПМИД 28565383 . Архивировано из оригинала (PDF) 5 мая 2015 года . Проверено 4 мая 2015 г.
- ^ Морель, Л.; Меер, РКВ; Лофгрен, CS (1990). «Сравнение распознавания гнездящихся особей моногинных и полигинных популяций Solenopsis invicta (Hymenoptera: Formicidae)». Анналы Энтомологического общества Америки . 83 (3): 642–647. дои : 10.1093/aesa/83.3.642 .
- ^ Шмид-Хемпель 1998 , с. 191.
- ^ Трагер 1988 , с. 204.
- ^ Флетчер, DJC; Блюм, М.С.; Уитт, ТВ; Темпл, Н. (1980). «Моногиния и полигиния огненного муравья Solenopsis invicta ». Анналы Энтомологического общества Америки . 73 (6): 658–661. дои : 10.1093/aesa/73.6.658 .
- ^ Гринберг, Л.; Флетчер, DJC; Винсон, С.Б. (1985). «Различия в размерах рабочих и распределении холмиков в моногинных и полигинных колониях огненного муравья Solenopsis invicta Buren». Журнал Канзасского энтомологического общества . 58 (1): 9–18. JSTOR 25084596 .
- ^ Харрисон, Р.Г. (1980). «Полиморфизмы распространения у насекомых». Ежегодный обзор экологии и систематики . 11 (1): 95–118. doi : 10.1146/annurev.es.11.110180.000523 . JSTOR 2096904 .
- ^ ДеХир, CJ; Гудисман, MAD; Росс, КГ (1999). «Стратегии расселения королевы в форме огненного муравья Solenopsis invicta с несколькими королевами » (PDF) . Американский натуралист . 153 (6): 660–675. дои : 10.1086/303205 . hdl : 1853/31356 . ISSN 0003-0147 . JSTOR 303205 . S2CID 12981148 . Архивировано из оригинала (PDF) 24 сентября 2015 года . Проверено 4 мая 2015 г.
- ^ Келлер, Л. (1995). «Социальная жизнь: парадокс колоний с несколькими королевами» . Тенденции в экологии и эволюции . 10 (9): 355–360. Бибкод : 1995TEcoE..10..355K . дои : 10.1016/S0169-5347(00)89133-8 . ПМИД 21237068 .
- ^ Jump up to: а б Росс, Кеннет Г. (1993). «Система размножения огненного муравья Solenopsis invicta : влияние на генетическую структуру колонии» . Американский натуралист . 141 (4): 554–76. дои : 10.1086/285491 . JSTOR 2462750 . ПМИД 19425999 . S2CID 6556261 .
- ^ Гудисман, MAD; Санкович, К.А.; Ковач, Дж. Л. (2006). «Генетические и морфологические изменения в пространстве и времени у инвазивного огненного муравья Solenopsis invicta » (PDF) . Биологические инвазии . 9 (5): 571–584. дои : 10.1007/s10530-006-9059-8 . S2CID 25341015 . Архивировано из оригинала (PDF) 24 сентября 2015 года . Проверено 4 мая 2015 г.
- ^ Адамс, ЕС; Балас, МТ (1999). «Рабочая дискриминация маток во вновь основанных колониях огненного муравья Solenopsis invicta » . Поведенческая экология и социобиология . 45 (5): 330–338. дои : 10.1007/s002650050568 . S2CID 23876496 .
- ^ Чирино, МГ; Гилберт, Ле; Фольгарайт, П.Дж. (2012). «Поведенческая дискриминация между моногинными и полигинными красными огненными муравьями (Hymenoptera: Formicidae) в их естественном ареале» . Анналы Энтомологического общества Америки . 105 (5): 740–745. дои : 10.1603/AN11073 . S2CID 86326110 .
- ^ Гудисман, М.; Росс, КГ (1997). «Взаимосвязь числа маток и родства маток в многоматочных колониях огненного муравья Solenopsis invicta » . Экологическая энтомология . 22 (2): 150–157. Бибкод : 1997EcoEn..22..150G . дои : 10.1046/j.1365-2311.1997.t01-1-00052.x . S2CID 86706296 .
- ^ Адамс, ЕС; Чинкель, WR (2001). «Механизмы регуляции численности огненного муравья Solenopsis invicta : экспериментальное исследование» . Журнал экологии животных . 70 (3): 355–369. Бибкод : 2001JAnEc..70..355A . дои : 10.1046/j.1365-2656.2001.00501.x . S2CID 18657087 .
- ^ Прачана, Р.; Приям, А.; Левантис, И.; Николс, РА; Вурм, Ю. (2017). «Супергенный вариант Sb социальной хромосомы огненного муравья демонстрирует низкое разнообразие, но сильное отличие от SB» . Молекулярная экология . 26 (11): 2864–2879. Бибкод : 2017MolEc..26.2864P . дои : 10.1111/mec.14054 . ПМК 5485014 . ПМИД 28220980 .
- ^ Ван, Дж.; Вурм, Ю.; Нипитваттанафон, М.; Риба-Грогнуз, О.; Хуанг, Ю.-К.; Шумейкер, Д.; Келлер, Л. (2013). «Y-подобная социальная хромосома вызывает альтернативную организацию колоний у огненных муравьев» (PDF) . Природа . 493 (7434): 664–668. Бибкод : 2013Natur.493..664W . дои : 10.1038/nature11832 . ПМИД 23334415 . S2CID 4359286 .
- ^ Росс, КГ; Кригер, MJ; Шумейкер, Д.Д. (2003). «Альтернативные генетические основы ключевого социального полиморфизма огненных муравьев» . Генетика . 165 (4): 1853–1867. дои : 10.1093/генетика/165.4.1853 . ПМЦ 1462884 . ПМИД 14704171 .
- ^ «Сокращение биоразнообразия местной фауны и флоры Австралии из-за завезенного красного огненного муравья Solenopsis invicta (огненный муравей)» . Департамент окружающей среды и энергетики . Правительство Австралии. Архивировано из оригинала 22 сентября 2016 года . Проверено 10 августа 2016 г.
- ^ Jump up to: а б с д Войчик, Д.; Аллен, ЧР; Бреннер, Р.Дж.; Форис, Э.А.; Жувеназ, ДП; Лутц, Р.С. (2001). «Красные завезенные огненные муравьи: влияние на биоразнообразие» (PDF) . Американский энтомолог . 47 (1): 16–23. дои : 10.1093/ae/47.1.16 . Архивировано из оригинала (PDF) 16 декабря 2010 года.
- ^ Портер, SD; Савиньяно, Д.А. (1990). «Нашествие полигинных огненных муравьев уничтожает местных муравьев и разрушает сообщество членистоногих». Экология . 71 (6): 2095–2106. Бибкод : 1990Экол...71.2095П . дои : 10.2307/1938623 . JSTOR 1938623 . S2CID 56244004 .
- ^ Стокер, РЛ; Грант, МЫ; Брэдли Винсон, С. (1995). « Влияние Solenopsis invicta (Hymenoptera: Formicidae) на беспозвоночных, разлагающих падаль, в центральном Техасе» . Экологическая энтомология . 24 (4): 817–822. дои : 10.1093/ee/24.4.817 .
- ^ Рикс, БЛ; Винсон, бакалавр наук (1970). «Приемлемость кормления некоторыми насекомыми и различными водорастворимыми соединениями двух разновидностей импортированного огненного муравья» . Журнал экономической энтомологии . 63 (1): 145–148. дои : 10.1093/джи/63.1.145 .
- ^ Кук, Дж.Л. (2003). «Сохранение биоразнообразия на территории, пострадавшей от завезенного красного огненного муравья Solenopsis invicta (Hymenoptera: Formicidae)». Биоразнообразие и сохранение . 12 (2): 187–195. дои : 10.1023/А:1021986927560 . S2CID 28425331 .
- ^ Массер, член парламента; Грант, МЫ (1986). «Смертность мелких млекопитающих, вызванная огненными муравьями, в восточно-центральном Техасе». Юго-западный натуралист . 31 (4): 540–542. дои : 10.2307/3671712 . JSTOR 3671712 .
- ^ Фликинджер, Э.Л. (1989). «Наблюдения за хищничеством красных завезенных огненных муравьев на диких хлопковых крыс, отловленных живыми ловушками». Техасский научный журнал . 41 (2): 223–224.
- ^ Хилл, EP (1970). «Наблюдения за завозным хищничеством огненных муравьев на птенцов хлопчатобумажных муравьев». Материалы ежегодной конференции Юго-Восточной ассоциации уполномоченных по охоте и рыбной ловле . 23 : 171–181.
- ^ Тоули, CJ; Лангкилде, Т. (2016). «Инвазивный огненный муравей ( Solenopsis invicta ) похищает яйца ящерицы восточного забора ( Sceloporus undulatus )». Журнал герпетологии . 50 (2): 284–288. дои : 10.1670/15-017 . S2CID 85855854 .
- ^ Крепление, правая сторона; Траут, ЮВ; Мейсон, WH (1981). «Хищничество красного завезенного огненного муравья Solenopsis invicta (Hymenoptera: Formicidae) на яйца ящерицы Cnemidophorus sexlineatus (Squamata: Teiidae)» . Журнал Академии наук Алабамы . 52 : 66–70.
- ^ Бартлетт, Д. (1997). «40 лет мыслей о прерии Пейнс». Рептилии . 5 (7): 68, 70–73.
- ^ Лангкилде, Т. (2009). «Агрессивные огненные муравьи изменяют поведение и морфологию местных ящериц». Экология . 90 (1): 208–217. Бибкод : 2009Ecol...90..208L . дои : 10.1890/08-0355.1 . ПМИД 19294926 . S2CID 670780 .
- ^ Брайнер, Дж. (26 января 2009 г.). «Танец ящериц спасает от смертоносных муравьев» . ЖиваяНаука . Проверено 20 августа 2016 г.
- ^ Келехеар, Кристалл; Грэм, Шон П.; Лангкилде, Трейси (март 2017 г.). «Защитные стратегии пуэрториканских карликовых гекконов (Sphaerodactylus macrolepis) против инвазивных огненных муравьев» . Герпетологика . 73 (1): 48–54. doi : 10.1655/HERPETOLOGICA-D-16-00042 . ISSN 0018-0831 . S2CID 90559576 .
- ^ Фрид, PS; Нейтман, К. (1988). «Заметки о хищничестве находящейся под угрозой исчезновения хьюстонской жабы Bufo houstonensis ». Техасский научный журнал . 40 : 454–456.
- ^ Аллен, ЧР; Райс, КГ; Войчик, Д.П.; Персиваль, HF (1997). «Влияние завезенных красных огненных муравьев на новорожденных американских аллигаторов» . Журнал герпетологии . 31 (2): 318–321. дои : 10.2307/1565408 . JSTOR 1565408 .
- ^ Jump up to: а б Капинера 2008 , с. 1080.
- ^ Кастелланос, Адриан А.; Медейрос, Мэтью К.И.; Хамер, Габриэль Л.; Морроу, Майкл Э.; Юбэнкс, Микки Д.; Тил, Пит Д.; Хамер, Сара А.; Лайт, Джессика Э. (2016). «Снижение численности мелких млекопитающих и клещей на хозяине в связи с инвазивными красными огненными муравьями ( Solenopsis invicta )» . Письма по биологии . 12 (9): 20160463. doi : 10.1098/rsbl.2016.0463 . ПМК 5046925 . ПМИД 27651533 .
- ^ Молони, SD; Вандервуд, К. (2003). «Потенциальное экологическое воздействие завезенных красных огненных муравьев в восточную Австралию» (PDF) . Журнал сельскохозяйственной и городской энтомологии . 20 (3): 131–142. Архивировано из оригинала (PDF) 9 октября 2016 года . Проверено 20 августа 2016 г.
- ^ Локли, TC (1995). «Влияние завозного хищничества огненных муравьев на популяцию наименьшей крачки: исчезающий вид». Юго-западный энтомолог . 20 : 517–519.
- ^ Сайкс, П.Дж.; Арнольд, Калифорния (1986). «Красный завезенный огненный муравей ( Solenopsis invicta ) хищничал на птенцов скальной ласточки ( Hirundopyrhorota ) в восточно-центральном Техасе». Юго-западный натуралист . 31 (1): 105–106. дои : 10.2307/3670967 . JSTOR 3670967 .
- ^ Кампомицци, Эй Джей; Моррисон, ML; Фаррелл, СЛ; Уилкинс, Р.Н.; Дрис, Б.М.; Паккард, Дж. М. (2009). «Красные завезенные огненные муравьи могут снизить выживаемость гнезда певчих птиц» . Кондор . 111 (3): 534–537. дои : 10.1525/cond.2009.090066 . S2CID 84552544 .
- ^ Морроу, МЭ; Честер, RE; Ленен, SE; Дрис, Б.М.; Топфер, Дж. Э. (2015). «Косвенное влияние завезенных красных огненных муравьев на выживаемость выводка прерийных кур Аттуотера» . Журнал управления дикой природой . 79 (6): 898–906. Бибкод : 2015JWMan..79..898M . дои : 10.1002/jwmg.915 . ПМК 4745021 . ПМИД 26900176 .
- ^ Йосеф, Р.; Лорер, Ф.Е. (1995). «Логгерхедские сорокопуты, красные огненные муравьи и отвлекающие маневры?» (PDF) . Кондор . 97 (4): 1053–1056. дои : 10.2307/1369544 . JSTOR 1369544 .
- ^ «Окружающая среда: борьба с огненным муравьем» . Время . 2 ноября 1970 года. Архивировано из оригинала 22 сентября 2016 года . Проверено 28 августа 2016 г.
- ^ Капинера 2008 , с. 1771.
- ^ Кинг, младший; Чинкель, WR (2006). «Экспериментальные доказательства того, что завезенный огненный муравей Solenopsis invicta не подавляет конкурентно встречающихся муравьев в нарушенной среде обитания». Журнал экологии животных . 75 (6): 1370–1378. Бибкод : 2006JAnEc..75.1370K . дои : 10.1111/j.1365-2656.2006.01161.x . JSTOR 4125078 . ПМИД 17032369 . S2CID 14719552 .
- ^ Гао, Ю.; Лу, ЛХ.; Он, Ю.Р.; Ци, ГДж.; Чжан, JQ. (2011). «Конкуренция между красным завезенным огненным муравьем ( Solenopsis invicta Buren ) и двумя местными видами муравьев (Hymenoptera: Formicidae)». Acta Entomologica Sinica . 54 (5): 602–608.
- ^ ЛеБрун, Е.Г.; Эбботт, Дж.; Гилберт, Л.Е. (2013). «Импортированный сумасшедший муравей вытесняет импортированного огненного муравья, уменьшает и гомогенизирует сообщества луговых муравьев и членистоногих». Биологические инвазии . 15 (11): 2429–2442. Бибкод : 2013BiInv..15.2429L . дои : 10.1007/s10530-013-0463-6 . S2CID 15260861 .
- ^ Витоне, Т.; Лаки, А. (2014). «Муравей тротуарный — Tetramorium caespitum » . Избранные существа UF/IFAS . Университет Флориды. Архивировано из оригинала 19 апреля 2016 года . Проверено 4 апреля 2016 г.
- ^ Кинг, Т.Г.; Филлипс, Ю.А. (1992). «Уничтожение молодых колоний красного завезенного огненного муравья тротуарным муравьем». Энтомологические новости . 103 (3): 72–77. ISSN 0013-872X .
- ^ Чен, ЮК; Кафле, Л.; Ши, CJ (2011). «Межвидовая конкуренция между Solenopsis invicta и двумя местными видами муравьев, Pheidolefervens и Monomorium chinense » . Журнал экономической энтомологии . 104 (2): 614–621. дои : 10.1603/ec10240 . ПМИД 21510213 . S2CID 44631810 .
- ^ Калькатерра, Луизиана; Ливор, Япония; Дельгадо, А.; Бриано, JA (2008). «Экологическое доминирование красного завезенного огненного муравья Solenopsis invicta в его естественном ареале» . Экология . 156 (2): 411–421. Бибкод : 2008Oecol.156..411C . дои : 10.1007/s00442-008-0997-y . ПМИД 18305962 . S2CID 21842183 . [ постоянная мертвая ссылка ]
- ^ ЛеБрун, Е.Г.; Тиллберг, CV; Суарес, А.В.; Фолгарайт, П.Дж.; Смит, ЧР; Холуэй, окружной прокурор (2007). «Экспериментальное исследование конкуренции между огненными муравьями и аргентинскими муравьями в их естественном ареале» (PDF) . Экология . 88 (1): 63–75. doi : 10.1890/0012-9658(2007)88[63:AESOCB]2.0.CO;2 . ПМИД 17489455 .
- ^ Чжоу, А.; Лян, Г.; Лу, Ю.; Цзэн, Л.; Сюй, Ю. (2014). «Межвидовая конкуренция между красным завезенным огненным муравьем Solenopsis invicta Buren и муравьем-призраком Tapinoma melanocephalum Fabricius за ресурсы падевой росы, производимые инвазивным мучнистым червецом Phenacoccus solenopsis Tinsiley». Взаимодействие членистоногих и растений . 8 (5): 469–474. Бибкод : 2014APInt...8..469Z . дои : 10.1007/s11829-014-9324-1 . S2CID 18102670 .
- ^ Ву, Б.; Ван, Л.; Лян, Г.; Лу, Ю.; Цзэн, Л. (2014). «Механизм пищевой конкуренции между Solenopsis invicta Buren и Tapinoma melanocephalum Fabricius» . Социобиология . 61 (3): 265–273. doi : 10.13102/sociobiology.v61i3.265-273 (неактивен 21 мая 2024 г.). Архивировано из оригинала 22 сентября 2016 года.
{{cite journal}}: CS1 maint: DOI неактивен по состоянию на май 2024 г. ( ссылка ) - ^ Кассилл, Д.Л.; Во, К.; Беккер, Б. (2008). «Молодые огненные муравьи симулируют смерть и выживают среди агрессивных соседей». Naturwissenschaften . 95 (7): 617–624. Бибкод : 2008NW.....95..617C . дои : 10.1007/s00114-008-0362-3 . ПМИД 18392601 . S2CID 2942824 .
- ^ Кабашима, JN; Гринберг, Л.; Руст, МК; Пейн, Т.Д. (2007). «Агрессивное взаимодействие между Solenopsis invicta и Linepithemahumile (Hymenoptera: Formicidae) в лабораторных условиях» (PDF) . Журнал экономической энтомологии . 100 (1): 148–154. doi : 10.1603/0022-0493(2007)100[148:AIBSIA]2.0.CO;2 . ПМИД 17370822 . S2CID 5764682 . Архивировано из оригинала (PDF) 9 октября 2016 года . Проверено 11 августа 2016 г.
- ^ Капинера 2008 , с. 290.
- ^ Эпперсон, Дебора М.; Аллен, Крейг Р.; Хоган, Кэтрин Ф.Е. (январь 2021 г.). «Красные завезенные огненные муравьи уменьшают численность, богатство и разнообразие беспозвоночных в норах черепах-сусликов» . Разнообразие . 13 (1): 7. дои : 10.3390/d13010007 . ISSN 1424-2818 .
- ^ Германн, Сара Л.; Блэкледж, Карисса; Хаан, Натан Л.; Майерс, Эндрю Т.; Лэндис, Дуглас А. (4 октября 2019 г.). «Хищники яиц бабочки-монарха и личинок новорожденных более разнообразны, чем считалось ранее» . Научные отчеты . 9 (1): 14304. Бибкод : 2019NatSR...914304H . дои : 10.1038/s41598-019-50737-5 . ISSN 2045-2322 . ПМЦ 6778129 . ПМИД 31586127 .
- ^ Гист, Эмили А.; Берман, Дэвид Д.; Баум, Кристен А. (ноябрь 2023 г.). «Численность бабочек выше на территориях, обработанных от огненных муравьев» . Журнал управления дикой природой . 87 (8). Бибкод : 2023JWMan..87E2483G . дои : 10.1002/jwmg.22483 . ISSN 0022-541X . S2CID 261627655 .
- ^ Форис, Элизабет А.; Квисторф, Анна; Аллен, Крейг Р. (2001). «Потенциальное воздействие огненных муравьев (Hymenoptera: Formicidae) на находящихся под угрозой исчезновения махаонов (Lepidoptera: Papilionidae)» . Энтомолог Флориды . 84 (2): 254–258. дои : 10.2307/3496176 . ISSN 0015-4040 . JSTOR 3496176 .
- ^ Jump up to: а б Чжоу, А.; Лу, Ю.; Цзэн, Л.; Сюй, Ю.; Лян, Г. (2012). «Способствует ли мутуализм вторжению двух чужеродных видов? Случай Solenopsis invicta и Phenacoccus solenopsis » . ПЛОС ОДИН . 7 (7): е41856. Бибкод : 2012PLoSO...741856Z . дои : 10.1371/journal.pone.0041856 . ПМК 3402455 . ПМИД 22911859 .
- ^ Каплан, И.; Юбэнкс, доктор медицины (2002). «Нарушение численности хлопковой тли (Homoptera: Aphididae) — динамика естественного врага красными завезенными огненными муравьями (Hymenoptera: Formicidae)» . Экологическая энтомология . 31 (6): 1175–1183. дои : 10.1603/0046-225X-31.6.1175 . S2CID 34957731 .
- ^ Копплер, Л.Б.; Мерфи, Дж. Ф.; Юбэнкс, доктор медицины (2007). «Красные завезенные огненные муравьи (Hymenoptera: Formicidae) увеличивают численность тли в томатах» . Энтомолог Флориды . 90 (3): 419–425. doi : 10.1653/0015-4040(2007)90[419:RIFAHF]2.0.CO;2 . hdl : 10919/95478 . S2CID 41237855 .
- ^ Диас, Р.; Кнутсон, А.; Бернал, Дж.С. (2004). «Влияние красного завезенного огненного муравья на плотность популяции хлопковой тли и хищничество яиц коробочной совки и свекловичной совки» . Журнал экономической энтомологии . 97 (2): 222–229. дои : 10.1603/0022-0493-97.2.222 . ПМИД 15154439 . S2CID 198130004 .
- ^ Теддерс, WL; Рейли, CC; Вуд, BW; Моррисон, РК; Лофгрен, CS (1990). «Поведение Solenopsis invicta (Hymenoptera: Formicidae) в садах пекана» . Экологическая энтомология . 19 (1): 44–53. дои : 10.1093/ee/19.1.44 .
- ^ Табер, Стивен (август 2000 г.). Огненные муравьи (1-е изд.). Издательство Техасского университета A&M. п. 308. ISBN 0890969450
- ^ Подробнее, Дэниел Р.; Кольмайер, Р.Э.; Хоффман, Дональд Р. (март 2008 г.). «Смертельная анафилаксия на укусы местных огненных муравьев у младенца». Американский журнал судебной медицины и патологии . 29 (1): 62–63. дои : 10.1097/PAF.0b013e3181651b53 . ПМИД 19749619 .
- ^ Фицджеральд, Кевин Т.; Флуд, Арын А. (ноябрь 2006 г.). «Укусы перепончатокрылых». Клинические методы в практике мелких животных . 21 (4): 194–204. дои : 10.1053/j.ctsap.2006.10.002 . ПМИД 17265905 .
- ^ Роудс, Роберт Б.; Стаффорд, Честер Т.; Джеймс, Фрэнк К. (август 1989 г.). «Обзор фатальных анафилактических реакций на укусы завезенных огненных муравьев». Журнал аллергии и клинической иммунологии . 84 (2): 159–162. дои : 10.1016/0091-6749(89)90319-9 . ПМИД 2760357 .
- ^ Замит-Миранда, Дэниел; Фокс, Эдвард GP; МОНТЕЙРО, Ана Паула; ГАМА, Диого; ПУБЛАН, Луис Э.; де Араужо, Альмаир Феррейра; АРАУЖО, Мария ФК; Ателла, Джорджия К.; МАЧАДО, Эднильдо А.; Диас, Бруно Л. (26 сентября 2018 г.). «Аллергическая реакция, опосредованная белками яда огненных муравьев» . Научные отчеты . 8 (1): 14427. Бибкод : 2018NatSR...814427Z . дои : 10.1038/s41598-018-32327-z . ПМК 6158280 . ПМИД 30258210 .
- ^ Fox EGP (2016) Ядовитые токсины огненных муравьев. В: Гопалакришнаконе П., Кальвете Дж. (ред.) Геномика и протеомика яда. Токсинология. Спрингер, Дордрехт
- ^ дос Сантос Пинту, Хосе Р.А.; Фокс, Эдвард GP; Сайдемберг, Дэниел М.; САНТОС, Люсилин Д.; да Силва Менегассо, Аналли Р.; Коста-Мансо, Элиуде; МАЧАДО, Эднильдо А.; Буэно, Одаир К.; Пальма, Марио С. (22 августа 2012 г.). «Протеомный взгляд на яд огненного муравья Бурена». Журнал исследований протеома . 11 (9): 4643–4653. дои : 10.1021/pr300451g . ПМИД 22881118 .
- ^ Баер, Х.; Лю, Т.-Ю.; Андерсон, MC; Блюм, М.; Шмид, WH; Джеймс, Ф.Дж. (1979). «Белковые компоненты яда огненного муравья ( Solenopsis invicta )». Токсикон . 17 (4): 397–405. Бибкод : 1979Txcn...17..397B . дои : 10.1016/0041-0101(79)90267-8 . ПМИД 494321 .
- ^ Хоффман, доктор медицинских наук; Дав, Делавэр; Джейкобсон, Р.С. (ноябрь 1988 г.). «Аллергены в яде перепончатокрылых. XX. Выделение четырех аллергенов из завезенного яда огненных муравьев (Solenopsis invicta)». Журнал аллергии и клинической иммунологии . 82 (5, часть 1): 818–27. дои : 10.1016/0091-6749(88)90084-X . ПМИД 3192865 .
- ^ ФРИМАН, Т; ХИЛАНДЕР, Р; ОРТИЗ, А; МАРТИН, М (1992). «Импортная иммунотерапия огненными муравьями: эффективность экстрактов всего тела» . Журнал аллергии и клинической иммунологии . 90 (2). Эльзевир Б.В.: 210–215. дои : 10.1016/0091-6749(92)90073-б . ISSN 0091-6749 . ПМИД 1500625 .
- ^ Макдональд, М. (2006). «Красные под ногами» . Новый учёный . 189 (2538): 50. Архивировано из оригинала 25 апреля 2016 года.
- ^ Jump up to: а б с д «Красный завезенный огненный муравей: объявлен вредителем в Западной Австралии» . Министерство сельского хозяйства и продовольствия . Правительство Западной Австралии. 2014. Архивировано из оригинала 22 сентября 2016 года . Проверено 21 августа 2016 г.
- ^ Пиментел, Д.; Лах, Л.; Зунига, Р.; Моррисон, Д. (2000). «Экологические и экономические издержки неместных видов в Соединенных Штатах» . Бионаука . 50 (1): 53–65. doi : 10.1641/0006-3568(2000)050[0053:EAECON]2.3.CO;2 . S2CID 44703574 .
- ^ Дай, Дж. (8 декабря 2014 г.). «Вспышка красных огненных муравьев в Сиднее может стоить экономике миллиарды» . Сидней Морнинг Геральд . Проверено 8 декабря 2014 г.
- ^ Jump up to: а б с «Огненные муравьи» . Сельское хозяйство Виктория . Правительство Виктории. 2004. Архивировано из оригинала 22 сентября 2016 года . Проверено 20 августа 2016 г.
- ^ Смит, MR (1965). Муравьи, наводняющие дома на востоке США: их распознавание, биология и экономическое значение (PDF) . Вашингтон, округ Колумбия: Министерство сельского хозяйства США. п. 43. ОСЛК 6078460 .
- ^ Стайлз, Дж. Х.; Джонс, Р.Х. (1998). «Распространение красного завезенного огненного муравья формы Solenopsis invicta в местах обитания на дорогах и линиях электропередач». Ландшафтная экология . 13 (6): 335–346. дои : 10.1023/А:1008073813734 . S2CID 21131952 .
- ^ «США импортируют врагов огненных муравьев» . CNN . 30 ноября 1999 года . Проверено 1 ноября 2016 г.
- ^ Вандер Меер, Р.К. Вандер; Словик, Ти Джей; Торвилсон, Х.Г. (2002). «Полухимикаты, выделяемые электрически стимулированными красными огненными муравьями Solenopsis invicta ». Журнал химической экологии . 28 (12): 2585–2600. дои : 10.1023/А:1021448522147 . ПМИД 12564802 . S2CID 4705569 .
- ^ Маккей, В.П.; Винсон, SB; Ирвинг, Дж.; Мажди, С.; Мессер, К. (1992). «Влияние электрических полей на красного завезенного огненного муравья (Hymenoptera: Formicidae)» . Экологическая энтомология . 21 (4): 866–870. дои : 10.1093/ee/21.4.866 .
- ^ Словик, Ти Джей; Грин, БЛ; Торвилсон, Х.Г. (1997). «Обнаружение магнетизма у красного импортированного огненного муравья ( Solenopsis invicta ) с помощью магнитно-резонансной томографии». Биоэлектромагнетизм . 18 (5): 396–399. doi : 10.1002/(SICI)1521-186X(1997)18:5<396::AID-BEM7>3.0.CO;2-Y . ПМИД 9209721 .
- ^ Андерсон, Дж. Б.; Вандер Меер, РК (1993). «Магнитная ориентация огненного муравья Solenopsis invicta » . естественные науки . 80 (12): 568–570. Бибкод : 1993NW.....80..568A . дои : 10.1007/BF01149274 . S2CID 41502040 .
- ^ Jump up to: а б Джеттер, К.М.; Гамильтон, Дж.; Клотц, Дж. Х. (2002). «Рассчитаны затраты на искоренение: завезенные красные огненные муравьи угрожают сельскому хозяйству, дикой природе и домам» (PDF) . Калифорнийское сельское хозяйство . 56 (1): 26–34. дои : 10.3733/ca.v056n01p26 . S2CID 84157527 .
- ^ Гринберг, Лес (2018). «Красный импортный огненный муравей Solenopsis invicta» . Калифорнийский университет Риверсайд : 1–2.
- ^ «Воздействие огненных муравьев – почему они представляют собой проблему» . Департамент сельского хозяйства и рыболовства . Правительство Квинсленда. 2016. Архивировано из оригинала 22 сентября 2016 года . Проверено 20 августа 2016 г.
- ^ Лофгрен, CS; Адамс, Коннектикут (1981). «Снижение урожайности соевых бобов на полях, зараженных красным огненным муравьем Solenopsis invicta ». Энтомолог Флориды . 64 (1): 199–202. дои : 10.2307/3494619 . JSTOR 3494619 .
- ^ Бэнкс, Вашингтон; Адамс, Коннектикут; Лофгрен, CS; Войчик, Д.П. (1990). «Завозное заражение огненными муравьями соевых полей на юге США» . Энтомолог Флориды . 73 (3): 503–504. дои : 10.2307/3495467 . JSTOR 3495467 .
- ^ Адамс, Коннектикут; Пламли, Дж. К.; Лофгрен, CS; Бэнкс, Вашингтон (1976). «Экономическое значение завезенного красного огненного муравья Solenopsis invicta Buren. I. Предварительные исследования воздействия на урожай сои». Журнал Энтомологического общества Джорджии . 11 : 165–169.
- ^ Адамс, Коннектикут; Пламли, Дж. К.; Бэнкс, Вашингтон; Лофгрен, CS (1977). «Влияние красного завезенного огненного муравья Solenopsis invicta Buren (Hymenoptera: Formicidae) на урожай сои в Северной Каролине». Журнал Общества Элиши Митчелла . 93 : 150–153.
- ^ Апперсон, CS; Адамс, Коннектикут (1983). «Медицинское и сельскохозяйственное значение красного завезенного огненного муравья». Энтомолог Флориды . 66 (1): 121–126. дои : 10.2307/3494558 . JSTOR 3494558 .
- ^ Капинера 2008 , с. 903.
- ^ Бэнкс, Вашингтон; Адамс, Коннектикут; Лофгрен, CS (1991). «Повреждение молодых цитрусовых деревьев красным огненным муравьем (Hymenoptera: Formicidae)» . Журнал экономической энтомологии . 84 (1): 241–246. дои : 10.1093/джи/84.1.241 .
- ^ Капинера 2008 , с. 1084.
- ^ Стерлинг, WL (1978). «Случайное биологическое подавление коробочного долгоносика завезенным красным огненным муравьем». Экологическая энтомология . 7 (4): 564–568. дои : 10.1093/ee/7.4.564 .
- ^ Филман, Д.А.; Стерлинг, WL (1983). «Убойная сила красного импортированного огненного муравья [Hym.: Formicidae]: основного хищника коробочного долгоносика [Col.: Curculionidae]». Энтомофага . 28 (4): 339–344. дои : 10.1007/BF02372186 . S2CID 38550501 .
- ^ Стерлинг, WL; Дин, округ Колумбия; Филман, Д.А.; Джонс, Д. (1984). «Естественный биологический контроль над долгоносиком [колонка: Curculionidae]». Энтомофага . 29 (1): 1–9. дои : 10.1007/BF02372203 . S2CID 33045477 .
- ^ Негм, А.А.; Хенсли, SD (1967). «Взаимосвязь членистоногих хищников с ущербом урожаю, нанесенным мотыльком сахарного тростника» . Журнал экономической энтомологии . 60 (6): 1503–1506. дои : 10.1093/джи/60.6.1503 .
- ^ Рейган, Т.Э.; Коберн, Г.; Хенсли, SD (1972). «Влияние мирекса на фауну членистоногих поля сахарного тростника в Луизиане» . Экологическая энтомология . 1 (5): 588–591. дои : 10.1093/ee/1.5.588 .
- ^ Капинера 2008 , с. 4173.
- ^ Лемке, Луизиана; Киссам, Дж. Б. (1988). «Воздействие красного завезенного огненного муравья (Hymenoptera: Formicidae) на мух-рогосов (Diptera: Muscidae) на пастбище для крупного рогатого скота, обработанном продроном» . Журнал экономической энтомологии . 81 (3): 855–858. дои : 10.1093/джи/81.3.855 . ПМИД 3403766 .
- ^ Ли, Дж.-Х.; Джонсон, С.Дж.; Райт, В.Л. (1990). «Количественный анализ выживаемости куколок бархатной фасоли (Lepidoptera: Noctuidae) на соевых полях в Луизиане» . Экологическая энтомология . 19 (4): 978–986. дои : 10.1093/ee/19.4.978 .
- ^ Моррилл, WL (1977). «Красный завезенный огненный муравей кормится в теплице». Экологическая энтомология . 6 (3): 416–418. дои : 10.1093/ee/6.3.416 .
- ^ Jump up to: а б с Юбэнкс, доктор медицины (2001). «Оценки прямого и косвенного воздействия красных завезенных огненных муравьев на биологический контроль полевых культур». Биологический контроль . 21 (1): 35–43. Бибкод : 2001BiolC..21...35E . дои : 10.1006/bcon.2001.0923 . S2CID 86682078 .
- ^ Уилсон, Нидерланды; Олив, AD (1969). «Пищевые привычки завезенных огненных муравьев на пастбищах и в сосновых лесах на юго-востоке Луизианы» . Журнал экономической энтомологии . 62 (6): 1268–1271. дои : 10.1093/джи/62.6.1268 .
- ^ Ху, ГЯ; Фрэнк, Дж. Х. (1996). «Влияние красного завезенного огненного муравья (Hymenoptera: Formicidae) на членистоногих, обитающих в навозе во Флориде» . Экологическая энтомология . 25 (6): 1290–1296. дои : 10.1093/ee/25.6.1290 .
- ^ Лопес, Джей Ди (1982). «Схема появления зимующей популяции Cardiochiles nigriceps в центральном Техасе». Экологическая энтомология . 11 (4): 838–842. дои : 10.1093/ee/11.4.838 .
- ^ Эллиот, Д. (1 июля 2013 г.). « Безумные муравьи кишат в юго-восточных штатах» . Новости CBS . Проверено 22 ноября 2016 г.
- ^ Лофгрен, CS; Бэнкс, Вашингтон; Гланси, Б.М. (1975). «Биология и борьба с завозными огненными муравьями». Ежегодный обзор энтомологии . 20 (1): 1–30. doi : 10.1146/annurev.en.20.010175.000245 . ПМИД 1090234 .
- ^ Бухс 2005 , стр. 1–2.
- ^ Уильямс, DF; деШазо, Р.Д. (2004). «Биологический контроль над огненными муравьями: обновленная информация о новых методах». Анналы аллергии, астмы и иммунологии . 93 (1): 15–22. дои : 10.1016/S1081-1206(10)61442-1 . ПМИД 15281467 .
- ^ Моттерн, Дж.Л.; Хайнц, К.М.; Ода, Пи Джей (2004). «Оценка биологического контроля огненных муравьев с помощью форидных мух: влияние на конкурентные взаимодействия». Биологический контроль . 30 (3): 566–583. Бибкод : 2004BiolC..30..566M . doi : 10.1016/j.biocontrol.2004.02.006 .
- ^ Дрис, Б.М.; Пакетт, Р. «Потенциальные агенты биологической борьбы с завезенными красными огненными муравьями» (PDF) . Техасский импортный проект по исследованию и управлению огненными муравьями . Техасский университет A&M . Проверено 21 августа 2016 г.
- ^ Бурен, ВФ; Аллен, GE; Уильямс, Р.Н. (1978). «Подходы к возможной борьбе с вредителями завезенных огненных муравьев» . Бюллетень Энтомологического общества Америки . 24 (4): 418–421. дои : 10.1093/беса/24.4.418 .
- ^ Jump up to: а б с «Руководство по комплексной борьбе с вредителями: пожарные вещества» . Служба национальных парков . Министерство внутренних дел США. 2010. Архивировано из оригинала 22 сентября 2016 года . Проверено 21 августа 2016 г.
- ^ Боннефой, X.; Кампен, Х.; Суини, К. (2008). Значение городских вредителей для общественного здравоохранения . Копенгаген, Дания: Всемирная организация здравоохранения, Европейское региональное бюро. п. 193 . ISBN 978-92-890-7188-8 .
- ^ Каликсто, А.А.; Харрис, МК; Кнутсон, А.; Барр, CL (2007). «Реакция местных муравьев на сокращение количества Solenopsis invicta Buren с использованием приманок для трансляции» . Экологическая энтомология . 36 (5): 1112–1123. doi : 10.1603/0046-225X(2007)36[1112:НАРТСИ]2.0.CO;2 . ПМИД 18284735 . S2CID 42073024 .
- ^ Клотц, Дж. Х.; Вейл, К.М.; Уильямс, Д.Ф. (1997). «Токсичность водной приманки борная кислота-сахароза для Solenopsis invicta (Hymenoptera: Formicidae)» . Журнал экономической энтомологии . 90 (2): 488–491. дои : 10.1093/джи/90.2.488 .
- ^ Бенсон, EP; Зунголи, Пенсильвания; Райли, МБ (2003). «Влияние загрязняющих веществ на прием приманки Solenopsis invicta (Hymenoptera: Formicidae)» . Журнал экономической энтомологии . 96 (1): 94–97. дои : 10.1093/джи/96.1.94 . ПМИД 12650350 .
Библиография
[ редактировать ]- Бухс, Дж. Б. (2005). Войны огненных муравьев: природа, наука и государственная политика в Америке двадцатого века . Чикаго, Иллинойс: Издательство Чикагского университета. ISBN 978-0-226-07981-3 .
- Капинера, Дж.Л. (2008). Энциклопедия энтомологии (2-е изд.). Дордрехт, Нидерланды: Springer. ISBN 978-1-4020-6242-1 .
- Хёльдоблер, Б.; Уилсон, Э.О. (1990). Муравьи . Кембридж, Массачусетс: Belknap Press издательства Гарвардского университета. ISBN 978-0-674-04075-5 .
- Шмид-Хемпель, П. (1998). Паразиты у социальных насекомых . Принстон, Нью-Джерси: Издательство Принстонского университета. ISBN 978-0-691-05924-2 .
- Табер, SW (2000). Огненные муравьи . Колледж-Стейшн, Техас: Издательство Техасского университета A&M. ISBN 978-1-60344-711-9 .
- Трагер, Дж. К. (1988). Достижения мирмекологии (1-е изд.). Лейден, Нидерланды: Э.Дж. Брилл. ISBN 978-0-916846-38-1 .
- Чинкель, В. (2006). Огненные муравьи . Кембридж, Массачусетс: Belknap Press издательства Гарвардского университета. ISBN 978-0-674-02207-2 .
Внешние ссылки
[ редактировать ]Веб-сайт
[ редактировать ]- Книги и журналы о красном завезенном огненном муравье в Библиотеке наследия биоразнообразия.
- Книги и журналы о красном импортированном огненном муравье в Интернет-архиве.
- Национальная программа по искоренению завезенных красных огненных муравьев. Архивировано 19 февраля 2017 г. в Wayback Machine - Министерство сельского хозяйства и рыболовства Квинсленда.
- Импортированные огненные муравьи – Министерство сельского хозяйства США
- Solenopsis invicta в AntWiki – появление муравьев в мире
- Проект S. invicta. генома Архивировано 11 сентября 2019 г. в Wayback Machine - проект базы данных генома перепончатокрылых.
- Профиль видов — красный импортированный огненный муравей ( Solenopsis invicta ) , Национальный информационный центр по инвазивным видам, Национальная сельскохозяйственная библиотека США .
СМИ
[ редактировать ]- Кадры форидных мух, паразитирующих на огненных муравьях. Архивировано 7 ноября 2022 года на Wayback Machine . Снято BBC, опубликовано Техасским университетом в Остинском колледже естественных наук.
- Красные огненные муравьи - опубликовано National Geographic на YouTube.
- Заливка колонии огненных муравьев расплавленным алюминием на YouTube
Освещение новостей
[ редактировать ]- Невероятный парящий огненный муравей – The Washington Post , 2011 г.
| Базы данных органов управления : Национальные |
|---|
- Соленопсис (муравей)
- Перепончатокрылые Азии
- Перепончатокрылые Австралии
- Перепончатокрылые Северной Америки
- Перепончатокрылые Южной Америки
- Насекомые описаны в 1916 году.
- Насекомые описаны в 1972 году.
- Мусорщики
- Сельскохозяйственные вредители
- Биологические средства борьбы с вредителями
- Насекомые, используемые в качестве средств борьбы с вредителями
- Насекомые-вредители
- Бытовые насекомые-вредители