Открытие и разработка ингибиторов интегразы
Эту статью необходимо обновить . ( ноябрь 2013 г. ) |
Первый случай заражения вирусом иммунодефицита человека ( ВИЧ ) был зарегистрирован в США в начале 1980-х годов. Было обнаружено множество лекарств для лечения этого заболевания, но мутации вируса и устойчивость к лекарствам затрудняют разработку. Интеграза — вирусный фермент , который интегрирует ретровирусную ДНК в геном клетки-хозяина . Ингибиторы интегразы — новый класс препаратов, используемых для лечения ВИЧ. Первый ингибитор интегразы, ралтегравир , был одобрен в 2007 году, а в 2011 году другие препараты проходили клинические испытания.
История
[ редактировать ]Организм использует свою иммунную систему для защиты от бактерий, вирусов и других болезнетворных организмов, а когда он не может этого сделать, возникают заболевания иммунодефицита. Одним из таких заболеваний является синдром приобретенного иммунодефицита ( СПИД ), который чаще всего является результатом заражения вирусом иммунодефицита человека (ВИЧ). [ 1 ] Были идентифицированы два близкородственных типа ВИЧ: ВИЧ-1 и ВИЧ-2 . В то время как ВИЧ-2 распространяется в Индии и Западной Африке, ВИЧ-1 более вирулентен и является причиной номер один СПИДа во всем мире. Хотя у некоторых пациентов результаты различаются, в большинстве случаев у людей, инфицированных ВИЧ, развивается СПИД и в конечном итоге они умирают от оппортунистических инфекций или рака . Интеграция в ретровирусный геном имеет решающее значение для экспрессии генов и репликации вируса . Вирусный геном обратно транскрибируется в ДНК инфицированной клетки с помощью вирусной обратной транскриптазы , затем ДНК интегрируется в хромосомы клетки-хозяина с помощью вирусной интегразы. Транскрипты РНК производятся из интегрированной вирусной ДНК и служат как мРНК, направляющими синтез вирусных белков, так и позже как геномы РНК новых вирусных частиц. Вирусные частицы покидают клетку путем отпочкования от плазматической мембраны, каждая из которых заключена в мембранную оболочку . [ 2 ]
В этом процессе интеграза ВИЧ-1 играет важную роль и, следовательно, является очень многообещающей мишенью для разработки лекарств против СПИДа. Селективная разработка лекарств возможна, поскольку интеграза ВИЧ-1 не имеет известного клеточного эквивалента. [ 3 ] Многие ингибиторы интегразы были открыты и разработаны, но лишь немногие из молекул получили дальнейшее развитие и дошли до фазы II или фазы III клинических испытаний . Ралтегравир (торговая марка Isentress) получил ускоренное одобрение США Управления по контролю за продуктами и лекарствами (FDA) в октябре 2007 года и региона EMEA (ныне EMA ) в декабре 2007 года. [ 4 ] [ 5 ] Он продавался как антиретровирусный препарат (АРВ) для взрослых, инфицированных ВИЧ-1, которые уже подвергались воздействию как минимум трех классов АРВ-препаратов и демонстрировали множественную лекарственную устойчивость. В целом существует две основные группы ингибиторов интегразы; Ингибиторы переноса цепи интегразы (INSTI) и ингибиторы связывания интегразы (INBI). INSTI сдерживают связывание преинтеграционного комплекса ( PIC ) и ДНК хозяина, а INBI сдерживают связывание интегразы и вирусной ДНК. Ралтегравир является ингибитором интегразы INSTI, который ингибирует репликацию ВИЧ-1 и ВИЧ-2. Он более эффективен, чем другие ранее известные ингибиторы интегразы, а также вызывает меньше побочных эффектов. Ралтегравир, Элвитегравир, Долутегравир и Биктегравир — единственные ингибиторы интегразы ВИЧ-1, используемые для лечения ВИЧ-инфекции S/GSK1349572 . [ 4 ] [ 6 ] [ 7 ] [ 8 ]
Фермент интеграза ВИЧ-1
[ редактировать ]Интеграза ВИЧ-1 (ИН) является ключевым ферментом механизма репликации ретровирусов. [ 9 ] Он отвечает за перенос кодируемой вирусом ДНК в хромосому хозяина, что является необходимым событием при репликации ретровируса. [ 10 ] Поскольку ИН не имеет эквивалента в клетке-хозяине, ингибиторы интегразы обладают высоким терапевтическим индексом , поскольку не влияют на нормальные клеточные процессы. [ 11 ]
Структура
[ редактировать ]ИН принадлежит как механистически, так и структурно к суперсемейству полинуклеотидилтрансфераз 10 и состоит из 288 аминокислот, которые образуют белок массой 32 кДа. [ 9 ] Ретровирусы кодируют свои ферменты (протеазу, обратную транскриптазу и интегразу) геном POL , 3'-конец которого кодирует IN. [ 11 ]

IN состоит из 3 структурно независимых функциональных доменов (см. рисунок 1): [ 9 ]
1. N-концевой домен (NTD) включает аминокислоты 1–50 и содержит два остатка гистидина (His12 и His16) и два остатка цистеина (Cys40 и Cys43), все из которых абсолютно консервативны и образуют мотив цинковых пальцев HHCC. . [ 9 ] [ 12 ] Одиночные мутации любого из этих четырех остатков снижают ферментативную активность ИН. [ 11 ] Мотив «цинковых пальцев» HHCC хелатирует один атом цинка на мономер IN . NTD необходим для образования мультимеров более высокого порядка , что, по-видимому, является его основной ролью. [ 12 ] [ 13 ] Для мультимеризации требуется атом цинка, который стабилизирует складку. [ 12 ]
2. Каталитический коровый домен (CCD), который включает аминокислоты 51–212, содержит активный сайт IN, но не может катализировать интеграцию в отсутствие NTD и CTD (С-концевой домен). [ 11 ] CCD содержит три абсолютно консервативные отрицательно заряженные аминокислоты; Д64, Д116 и Е152. [ 9 ] Эти аминокислоты образуют мотив DDE, который координирует ионы двухвалентных металлов (Mg 2+ или Мн 2+ ). Эти ионы металлов необходимы для катализа интеграции. [ 12 ] [ 13 ] CCD имеет смешанную β- и α-структуру с пятью β-листами и шестью α-спиралями , соединенными гибкими петлями. [ 12 ] Гибкие петли допускают конформационные изменения, необходимые для 3- процессинга вирусной ДНК и реакций переноса цепи (STF), которые являются двумя ключевыми этапами реакции интеграции. [ 9 ] CCD необходим для этих стадий, и замена любого из остатков в мотиве DDE резко ингибирует активность IN. [ 12 ]
3. С-концевой домен (CTD), который включает аминокислоты 213–288, неспецифически связывает ДНК, и его взаимодействие с NTD и CCD необходимо для активности IN 3'-процессинга и переноса цепи. [ 11 ] [ 12 ] CTD является наименее консервативным из трех доменов. [ 12 ] IN действует как мультимер, и димеризация для стадии 3´-процессинга необходима , при этом тетрамерный IN катализирует реакцию переноса цепи. [ нужна ссылка ]
Функция
[ редактировать ]Интеграция ВИЧ-1 происходит посредством многоэтапного процесса, который включает две каталитические реакции: 3’эндонуклеолитический процессинг концов провирусной ДНК (называемый 3’процессингом) и интеграцию 3’-процессированной вирусной ДНК в клеточную ДНК (называемую переносом цепи). [ 6 ] При 3'-процессинге ИН связывается с короткой последовательностью, расположенной на обоих концах длинного концевого повтора (LTR) вирусной ДНК, и катализирует расщепление эндонуклеотида. Это приводит к удалению динуклеотида с каждого из 3'-концов LTR. Расщепленная ДНК затем используется в качестве субстрата для интеграции или переноса цепи. [ 9 ] Перенос цепи представляет собой реакцию трансэтерификации, включающую прямую нуклеофильную атаку 3'гидроксигруппы двух вновь процессированных концов 3'-ДНК вируса на фосфодиэфирный остов ДНК-мишени хозяина. [ 14 ] Это приводит к ковалентному внедрению вирусной ДНК в геном инфицированной клетки. Перенос цепи происходит одновременно на обоих концах молекулы вирусной ДНК со смещением ровно в пять пар оснований между двумя противоположными точками вставки. [ 9 ] Реакция интеграции завершается удалением неспаренных динуклеотидов с 5'-концов вирусной ДНК, восстановлением одноцепочечных разрывов, образовавшихся между молекулами вирусной и целевой ДНК, и лигированием 3'-концов с 5'-концами вирусной ДНК. ДНК хозяина. [ 9 ] [ 14 ] Двухвалентные металлы, Мг 2+ или Мн 2+ , необходимы для стадий 3'-процессинга и переноса цепи, а также для сборки IN на специфической вирусной донорской ДНК с образованием комплекса, способного выполнять любую функцию. Поскольку обилие магния (Mg 2+ ) по сравнению с марганцем (Mn 2+ ) в клетках человека увеличивается в 1 000 000 раз, магний кажется более доступным двухвалентным кофактором для интеграции. [ 6 ]
Механизм действия
[ редактировать ]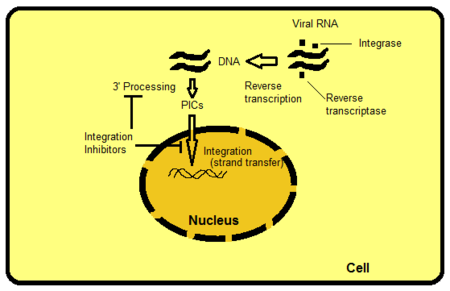
Существует несколько способов воздействия на интегразу, но ингибирование переноса цепи является наиболее интуитивно очевидным и легко реализуемым на сегодняшний день. Другие мишени включают, например, белковые домены за пределами активного сайта ИН. Домены взаимодействуют с ДНК вируса или хозяина и важны для связывания с ферментом. Можно препятствовать функциям фермента, нарушая или удаляя эти связывания. PIC представляет собой мультимерную белковую структуру внутри клетки-хозяина, состоящую как из вирусных белков, так и из белков-хозяев. Интеграза является частью вирусного компонента PIC. Считается, что вирусные белки PIC и белки хозяина модулируют внутреннюю активность фермента, доставляют PIC в ядро и осуществляют прямую интеграцию вирусной ДНК в транскрипционно активную область генома хозяина. Если бы можно было исключить определенные белки из PIC, это заблокировало бы способность вируса интегрироваться в геном хозяина. Процесс, при котором ретровирусная РНК транскрибируется в ДНК и затем интегрируется в геном клетки-хозяина, показан на рисунке 2. [ 8 ]
Ингибиторы переноса цепи IN (INSTI)
[ редактировать ]мг 2+ и Мн 2+ являются важнейшими кофакторами на этапе интеграции. Инактивация этих кофакторов (например, посредством хелатирования) вызывает функциональные нарушения ИН. Эта концепция дает исследователям возможность проектировать и разрабатывать высокоэффективные ингибиторы ИН (ИНИ). Фактически, все низкомолекулярные INI ВИЧ-1, которые сейчас исследуются, содержат структурный мотив, который координирует два двухвалентных иона магния в активном центре фермента. [ 6 ]
Ралтегравир и элвитегравир имеют одинаковый механизм действия против интегразы: связываются с активным центром Mg. 2+ ионы. [ 8 ] Конкурентные ингибиторы напрямую конкурируют с вирусной ДНК за связывание с интегразой, чтобы ингибировать процессинг 3'-конца. [ 15 ] При этом ингибиторы полностью блокируют связывание активного сайта с целевой ДНК. Это ингибирование называется ингибированием переноса цепи . [ 8 ]
Ингибирование взаимодействия LEDGF/p75-интеграза
[ редактировать ]Фактор роста, полученный из эпителия хрусталика ( LEDGF/p75 ), представляет собой белок-хозяин, который связывается с интегразой и имеет решающее значение для репликации вируса. Механизм действия точно не известен, но данные свидетельствуют о том, что LEDGF/p75 направляет интегразу для вставки вирусной ДНК в транскрипционно активные сайты генома хозяина. Ингибиторы этого белка уже разрабатываются и запатентованы. Они, вероятно, будут в высшей степени целенаправленными и менее склонны к развитию резистентности. [ 8 ]
Ингибиторы связывания ИН
[ редактировать ]Другим классом ИНИ могут быть ингибиторы связывания ИН (ИНБИ), такие как V-165. V-165 представляет собой соединение, которое, как было показано, ингибирует интеграцию, но не оказывает очевидного влияния на синтез вирусной ДНК. При изучении механизма действия выяснилось, что V-165 препятствует образованию комплекса вирусной ДНК-ИН. Из-за своего мешающего действия его классифицируют как ингибитор связывания ИН. Другие соединения, такие как стирилхинолины, имеют аналогичный механизм, конкурируя с субстратом LTR за связывание IN. [ 16 ]
Дизайн лекарств
[ редактировать ]Связывание
[ редактировать ]INSTI прочно и специфично связываются с IN, который связан с концами ДНК, путем хелатирования ионов двухвалентных металлов (Mg 2+ ), которая координируется каталитической триадой, т.е. мотивом DDE. [ 9 ] Мотив DDE расположен в CCD IN и является активным центром фермента, и, следовательно, INSTI являются так называемыми ингибиторами активного центра. INSTI связываются со специфическим сайтом, близким к мотиву DDE IN, сайтом, который присутствует только в той конформации, которая возникает после процессинга 3'-концов вирусной ДНК. Вирусная ДНК вполне может составлять часть сайта связывания ингибитора. Связывание является формой аллостерического ингибирования , поскольку оно подразумевает блокировку специфического комплекса интеграза-вирусная ДНК. [ 12 ] Это приводит к избирательному ингибированию реакции переноса цепи без существенного влияния на реакцию 3'-процессинга. [ 9 ] Таким образом, INSTI могут быть более специфичными и избирательно связываться с целевым сайтом связывания ДНК и, следовательно, быть менее токсичными, чем бифункциональные ингибиторы, которые способны связываться как с донорным, так и с целевым сайтом связывания. [ 12 ]
INBI также связываются с IN, но механизм действия неизвестен, поэтому подробное описание связывания невозможно. [ 16 ]
Отношения структура-активность (SAR)
[ редактировать ]
Для связывания интегразы необходимы два структурных компонента: гидрофобный бензильный фрагмент, который скрыт в высокогидрофобном кармане рядом с активным центром; и хелатирующая триада, которая связывается с двумя Mg 2+ ионы в достаточно гидрофильной области, закрепляя ингибитор на поверхности белка (см. рисунок 3). [ 17 ] Фактически, все мощные ингибиторы интегразы содержат замещенный бензильный компонент, который имеет решающее значение для поддержания эффективности соединения 3'-концов. Удаление бензильной группы предотвращает ингибирующую функцию. [ 15 ] Таким образом, липофильные заместители полезны для ингибирования переноса цепи, в частности тиофениловые , фуранильные и (тиофен-2-ил)фенильные замены. Гетероароматический амин и амид также вызывают усиление ингибирующего действия 3'-процессинга. [ 6 ]
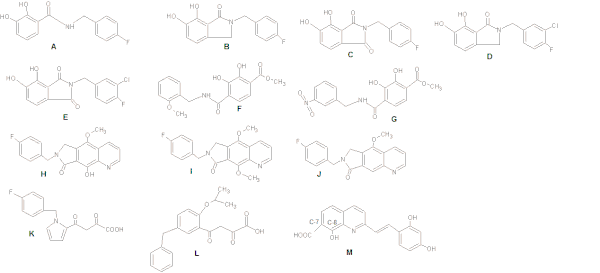
Когда были исследованы ингибиторы ИН на основе катехолов, было замечено, что поддержание плоского взаимодействия с бис-гидроксилированным арильным кольцом увеличивает эффективность. Ингибирующую активность можно дополнительно оптимизировать за счет включения заместителя метахлора, усиливающего взаимодействие бензильной группы с соседним гидрофобным карманом (см. рисунок 4: Структуры AG). [ 8 ]
Бензилзамещенная гидроксильная группа (рис. 4 H) улучшает способность к хелатированию металлов (по сравнению со структурой J на рис. 4), тогда как метоксигруппа (I) гораздо менее эффективна из-за стерического столкновения дополнительной метильной группы с каталитическими металлами. . [ 15 ]
При исследовании дикетопроизводных центральное пиррольное кольцо структуры К на рис. 4 был заменен рядом ароматических систем, имеющих различный характер замещения. Это обеспечивало оптимальную относительную ориентацию цепей бензильных и дикетокислотных (ДКА) участков. Структура L на рис. 4 привело к 100-кратному увеличению эффективности. [ 18 ]
Бенард и др . (2004) синтезировали INI с хинолиновой субъединицей и вспомогательным ароматическим кольцом, связанным функционализированными спейсерами, такими как амид, гидразид , мочевина и гидроксипроп-1-ен-3-оновая группа. Они обнаружили, что наиболее перспективными являются производные, содержащие амидную группу. [ 18 ] [ 19 ] Синтезируя ряд стирилхинонов, исследователи обнаружили, что для ингибирования необходимы карбоксильная группа при С-7, гидроксильная группа при С-8 в хинолиновой субъединице и вспомогательное фенильное кольцо (рис. 4: Структура M), хотя изменения кольца возможны. терпимо. Две гидроксильные группы на вспомогательном фенильном кольце также необходимы для ингибирующей активности. [ 18 ]
Фармакофор
[ редактировать ]
Поскольку информации о критической структуре катализа ВИЧ-интегразы недостаточно, трудно найти точный фармакофор для ее ингибирования. Wang et al (2010) надеялись, что, изучая SAR и фармакофор каркаса с двойным ингибитором, уделяя особое внимание как интегразе, так и обратной транскриптазе (RT), можно будет наблюдать антиинтегразную активность. Изучая SAR ингибиторов интегразы ВИЧ, удалось обнаружить, что для оптимального ингибирования интегразы фармакофору необходим региоспецифический (N-1) ДКА определенной длины. Функциональность DKA или ее гетероциклический биоизостер , которые избирательно ингибируют перенос цепи, по-видимому, присутствуют во всех основных хемотипах ингибиторов интегразы. [ 17 ] Как подробно описано выше в обсуждении SAR, двумя необходимыми структурными компонентами INI являются бензильная гидрофобная группа и хелатирующая триада, связывающая Mg. 2+ ионы. Для триады, связывающей Mg 2+ ионы должны быть ионизированы (см. рис. 5), и, следовательно, биоизостер фармакофора также должен быть ионизирован, а биоизостер бензил-фармакофора должен быть очень гидрофобным. [ 11 ] [ 17 ]
Однако, несмотря на предыдущие успехи в клинических разработках (ралтегравир), детальная модель связывания отсутствует, поэтому оказалось сложно структурировать основу для разработки ингибиторов интегразы. Когда фармакофор салициловой кислоты и катехина были объединены, были созданы новые химические каркасы. Соседние гидроксильные и карбоксильные группы салициловой кислоты могут связываться с ионами металлов и служить их фармакофором. Полигидроксилированные ароматические ингибиторы в основном активны в отношении реакций переноса цепи и 3'-процессинга, что предполагает механизм, нацеленный на обе стадии. Это очень важная часть соединения, поскольку ее можно использовать для связывания двухвалентного металла в активном сайте ИН и, как таковую, она эффективна против вирусных штаммов, устойчивых к специфическим ингибиторам переноса цепи. [ 6 ] [ 17 ]
Сопротивление
[ редактировать ]Было обнаружено, что более 60 вариантов мутаций INSTI вызывают устойчивость in vivo и in vitro . Из-за этих мутаций и развития резистентности ингибиторы менее эффективны против вируса. [ 9 ] Резистентность ИНИ соответствует таковой у других АРВ-препаратов. Первая устойчивость к IN вызвана первичными мутациями, которые снижают чувствительность к INI, в сочетании со вторичными мутациями, которые еще больше снижают чувствительность вируса и/или восстанавливают пониженную приспособленность вируса. Во-вторых, существует генетический барьер устойчивости к ИНИ, определяемый количеством мутаций, необходимых для потери клинической активности ИНИ. существует обширная, но неполная перекрестная резистентность . В-третьих, среди ИНИ [ 13 ] Петля, содержащая аминокислотные остатки 140–149, расположена в домене каталитического ядра и важна для функции IN, как упоминалось ранее. Эта петля является гибкой, и хотя ее роль не совсем известна, считается, что она важна, а ее функции критически важны для связывания ДНК. Эта устойчивость проявляется в результате мутаций в этой IN-кодирующей области. [ 9 ] Устойчивость к ралтегравиру и элвитегравиру в первую очередь обусловлена одними и теми же двумя путями мутаций, но для каждого из препаратов также задействованы и другие первичные мутации. [ 10 ] Некоторые мутации повышают устойчивость к лекарствам в большей степени, чем другие. Например, один из наиболее распространенных путей мутации увеличивает устойчивость к ралтегравиру до 100 раз больше, чем второй по распространенности. [ 9 ] Устойчивость к ингибитору интегразы S/GSK1349572 все еще разрабатывается, и устойчивость еще не полностью охарактеризована. Когда его оценивали вместе с первичными мутациями ралтегравира и элвитегравира, он не выявил перекрестной устойчивости, что означает, что он может быть полезен против вирусов, устойчивых к лекарственным средствам. [ 7 ] Ралтегравир имеет ограниченную кишечную абсорбцию, поэтому резистентность невозможно преодолеть назначением более высоких доз. Новые лекарства должны преодолеть этот фармакологический недостаток и достичь достаточно высоких концентраций в плазме, чтобы воздействовать на устойчивые к ралтегравиру вирусы. [ 7 ]
Текущий статус
[ редактировать ]
Поиск новых путей улучшения лечения пациентов, инфицированных ВИЧ, ведется постоянно. Учитывая опыт, накопленный с 1980-х годов по разработке АРВ-препаратов, появление INSTI в качестве нового мощного класса АРВ-препаратов сигнализирует о новой эре в лечении ВИЧ. Разработка успешного метода лечения INSTI была завершена, когда компания Merck Sharp & Dohme Limited обнаружила ралтегравир. [ 12 ] Условное регистрационное удостоверение было лицензировано Европейской Комиссией в декабре 2007 года и действовало на всей территории Европейского Союза . [ 20 ] В 2009 году это разрешение было преобразовано в полное регистрационное удостоверение , и в том же году FDA изменило разрешение с ускоренного на традиционное и включило препарат в список средств АРВ-терапии первого ряда. [ 12 ] [ 21 ] Второй препарат INSTI, элвитегравир, был разработан компанией Japan Tobacco, и его клинические испытания начались в 2005 году. В 2011 году препарат все еще находился на третьей фазе клинических испытаний, где его сравнивали с ралтегравиром, на пациентах, уже опытных в лечении, а также находится на второй фазе разработки. у наивных субъектов в рамках комбинированного лечения. [ 12 ] S/GSK1349572 — это ингибитор интегразы, открытый компанией ViiV/Shinongi, который вступал в третью фазу клинических испытаний в 2011 году. Этот новый препарат многообещающий, хорошо переносится и на данный момент показывает лучшие результаты, чем ралтегравир и элвитегравир. [ 22 ]
Поскольку возникли проблемы с устойчивостью к ралтегравиру и элвитегравиру, ученые начали работать над новыми ингибиторами интегразы второго поколения, такими как MK-2048 , который в 2009 году разработала компания Merck. Это прототип INSTI второго поколения, который остается эффективным против вирусов, содержащих мутации против ралтегравира и элвитегравира. Механизм действия и SAR МК-2048 такие же, как и у других INSTI, структура МК-2048 показана на рисунке 6 с выделенным основным фармакофором. [ 23 ] [ 24 ]
Несмотря на то, что обсуждаемые выше лекарства являются многообещающими, их разработка еще долгая, и многое еще неизвестно об эффективности, безопасности и механизме действия этих лекарств. [ 7 ]
См. также
[ редактировать ]- Ралтегравир
- Элвитегравир
- Интеграза
- Ингибитор интегразы
- ВИЧ
- Обратная транскриптаза
- МК-2048
- Долутегравир
- Биктегравир
Ссылки
[ редактировать ]- ^ Джонсон, Ди Унглауб Сильверторн; с Уильямом К. Обером, координатором иллюстраций; Клэр В. Гаррисон, иллюстратор; Эндрю С. Силверторн, клинический консультант; при участии Брюса Р. (2007). Физиология человека: комплексный подход (4-е изд.). Сан-Франциско: Пирсон/Бенджамин Каммингс. ISBN 978-0-8053-6849-9 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Мерфи, Кеннет; Трэверс, Пол; Уолпорт, Марк (2008). Иммунобиология Джейнвей (7-е изд.). Нью-Йорк: Garland Science. ISBN 978-0-8153-4123-9 .
- ^ Помье, Ю; Маршан, К; Неамати, Н. (сентябрь 2000 г.). «Ингибиторы ретровирусной интегразы, 2000 год: обновленная информация и перспективы». Противовирусные исследования . 47 (3): 139–48. дои : 10.1016/S0166-3542(00)00112-1 . ПМИД 10974366 .
- ^ Jump up to: а б Домбровская, Магдалена Моника; Верциньска-Драпало, Алисия (1 января 2007 г.). «Ингибиторы интегразы как новый класс АРВ-терапии» . Обзор ВИЧ и СПИДа . 6 (4): 10–14. дои : 10.1016/S1730-1270(10)60053-7 .
- ^ «Одобрение FDA препарата Изентресс (ралтегравир)» . Управление по санитарному надзору за качеством пищевых продуктов и медикаментов США (FDA) . Проверено 25 сентября 2011 г.
- ^ Jump up to: а б с д и ж Веер, Х; Чжан, Ф.Х.; Аль-Сафи, Род-Айленд; Цзэн, LF; Шабайк Ю.; Дебнат, Б; Санчес, ТВ; Одде, С; Неамати, Н; Лонг, YQ (15 августа 2011 г.). «Разработка ингибиторов интегразы ВИЧ-1, нацеленных на каталитический домен, а также его взаимодействие с LEDGF/p75: подход с использованием каркасных прыжков с использованием салицилатных и катехоловых групп» . Биоорганическая и медицинская химия . 19 (16): 4935–52. дои : 10.1016/j.bmc.2011.06.058 . ПМК 3163123 . ПМИД 21778063 .
- ^ Jump up to: а б с д Ленц, Дж. К.; Рокстро, Дж. К. (апрель 2011 г.). «S/GSK1349572, новый ингибитор интегразы для лечения ВИЧ: перспективы и проблемы». Экспертное заключение об исследуемых препаратах . 20 (4): 537–48. дои : 10.1517/13543784.2011.562189 . ПМИД 21381981 . S2CID 5442539 .
- ^ Jump up to: а б с д и ж Пендри, А; Минвелл, Северная Каролина; Пиз, К.М.; Уокер, Массачусетс (август 2011 г.). «Новые ингибиторы интегразы вируса иммунодефицита человека-1 первого и второго поколения». Экспертное заключение о терапевтических патентах . 21 (8): 1173–89. дои : 10.1517/13543776.2011.586631 . ПМИД 21599420 . S2CID 23324357 .
- ^ Jump up to: а б с д и ж г час я дж к л м н Мусаде, Ж. Ф.; Делелис, О; Марселин, АГ; Чертанов, Л. (август – октябрь 2010 г.). «Устойчивость к ингибиторам интегразы ВИЧ-1: структурная перспектива». Обновления в области лекарственной устойчивости: обзоры и комментарии по противомикробной и противораковой химиотерапии . 13 (4–5): 139–50. дои : 10.1016/j.drup.2010.05.001 . ПМИД 20570551 .
- ^ Jump up to: а б Кокохоба, Дж; Донг, Би Джей (октябрь 2008 г.). «Ралтегравир: первый ингибитор интегразы ВИЧ». Клиническая терапия . 30 (10): 1747–65. дои : 10.1016/j.clinthera.2008.10.012 . ПМИД 19014832 .
- ^ Jump up to: а б с д и ж Помье, Ив; Джонсон, Эллисон А.; Маршан, Кристоф (24 февраля 2005 г.). «Ингибиторы интегразы для лечения ВИЧ/СПИДа». Nature Reviews Открытие лекарств . 4 (3): 236–248. дои : 10.1038/nrd1660 . ПМИД 15729361 . S2CID 11505680 .
- ^ Jump up to: а б с д и ж г час я дж к л м Макколл, диджей; Чен, X (январь 2010 г.). «Ингибиторы переноса цепи интегразы ВИЧ-1: новая эра антиретровирусной терапии». Противовирусные исследования . 85 (1): 101–18. doi : 10.1016/j.antiviral.2009.11.004 . ПМИД 19925830 .
- ^ Jump up to: а б с Бланко, JL; Варгезе, В; Ри, С.Ю.; Гателл, Дж. М.; Шафер, RW (01 мая 2011 г.). «Резистентность к ингибитору интегразы ВИЧ-1 и ее клинические последствия» . Журнал инфекционных болезней . 203 (9): 1204–14. дои : 10.1093/infdis/jir025 . ПМК 3069732 . ПМИД 21459813 .
- ^ Jump up to: а б Де Лука, Лаура; Де Грация, Сара; Ферро, Стефания; Гитто, Розария; Господи, Фрауке; Дебизер, Зегер; Чимирри, Альба (февраль 2011 г.). «Ингибиторы переноса цепи интегразы ВИЧ-1: разработка, синтез и исследование молекулярного моделирования». Европейский журнал медицинской химии . 46 (2): 756–764. дои : 10.1016/j.ejmech.2010.12.012 . ПМИД 21227550 .
- ^ Jump up to: а б с Чен, X; Цзян, М; Ю, Ф; Хунг, М; Джонс, Г.С.; Зейналзадеган, А; Ци, Х; Джин, Х; Ким, CU; Сваминатан, С; Чен, Дж. М. (11 июля 2008 г.). «Моделирование, анализ и проверка новой структуры интегразы ВИЧ дают представление о способах связывания мощных ингибиторов интегразы». Журнал молекулярной биологии . 380 (3): 504–19. дои : 10.1016/j.jmb.2008.04.054 . ПМИД 18565342 .
- ^ Jump up to: а б Хомбрук, А.; Хантсон, А.; ван Ремортель, Б.; Михилс, М.; Веркаммен, Дж.; Родс, Д.; Тец, В.; Энгельборгс, Ю.; Христос, Ф.; Дебизер, З.; Витвроу, М. (июнь 2007 г.). «Выбор устойчивости вируса иммунодефицита человека типа 1 к пиранодипиримидину V-165 указывает на мультимодальный механизм действия» . Журнал антимикробной химиотерапии . 59 (6): 1084–95. дои : 10.1093/jac/dkm101 . ПМИД 17470918 .
- ^ Jump up to: а б с д Ван, З; Тан, Дж; Саломон, CE; Дрейс, CD; Винс, Р. (15 июня 2010 г.). «Фармакофор и взаимосвязь структура-активность ингибирования интегразы в каркасе с двойным ингибитором обратной транскриптазы и интегразы ВИЧ». Биоорганическая и медицинская химия . 18 (12): 4202–11. дои : 10.1016/j.bmc.2010.05.004 . ПМИД 20576573 .
- ^ Jump up to: а б с Дубей, С; Сатьянараяна, Ю.Д.; Лавания, Х (сентябрь 2007 г.). «Разработка ингибиторов интегразы для лечения СПИДа: обзор». Европейский журнал медицинской химии . 42 (9): 1159–68. дои : 10.1016/j.ejmech.2007.01.024 . ПМИД 17367896 .
- ^ Бенар, К; Зухири, Ф; Норманд-Бейл, М; Данет, М; Десмаэль, Д; Лех, Х; Мусаде, Ж. Ф.; Мбемба, Г; Томас, CM; Бонненфан, С; Ле Бре, М; д'Анджело, Дж (17 мая 2004 г.). «Линкер-модифицированные производные хинолина, нацеленные на интегразу ВИЧ-1: синтез и биологическая активность». Письма по биоорганической и медицинской химии . 14 (10): 2473–6. дои : 10.1016/j.bmcl.2004.03.005 . ПМИД 15109635 .
- ^ «Изентресс» (PDF) . Европейское агентство по лекарственным средствам . Проверено 17 сентября 2011 г.
- ^ «Традиционное одобрение Изентресса (ралтегравира)» . Управление по санитарному надзору за качеством пищевых продуктов и медикаментов США (FDA) . Проверено 25 сентября 2011 г.
- ^ Барнхарт, Мэтью; Джеймс Шелтон (апрель 2011 г.). «Улучшение состояния АРТ, улучшение схем антиретровирусной терапии для расширения глобального доступа к лечению ВИЧ». Журнал исследований СПИДа и ВИЧ . 3 (4): 71–78.
- ^ Бар-Маген, Т; Слоан, РД; Донахью, округ Колумбия; Куль, Б.Д.; Забейда, А; Сюй, Х; Оливейра, М; Хазуда, диджей; Вайнберг, Массачусетс (сентябрь 2010 г.). «Идентификация новых мутаций, ответственных за устойчивость к МК-2048, ингибитору интегразы ВИЧ-1 второго поколения» . Журнал вирусологии . 84 (18): 9210–6. дои : 10.1128/JVI.01164-10 . ПМЦ 2937597 . ПМИД 20610719 .
- ^ Гетальс, О; Вос, А; Ван Гиндерен, М; Гелюйкенс, П; Смитс, В; Шолс, Д; Хертогс, К; Клейтон, Р. (5 июля 2010 г.). «Первичные мутации, выбранные in vitro с помощью ралтегравира, приводят к значительным кратным изменениям чувствительности к ингибиторам интегразы первого поколения, но незначительным кратным изменениям к ингибиторам с профилем устойчивости второго поколения». Вирусология . 402 (2): 338–46. дои : 10.1016/j.virol.2010.03.034 . ПМИД 20421122 .