Видимый спектр

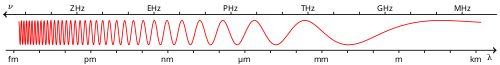
— Видимый спектр это диапазон электромагнитного спектра , видимый человеческим глазом . Электромагнитное излучение в этом диапазоне длин волн называется видимым светом (или просто светом). Оптический спектр иногда считают таким же, как видимый спектр, но некоторые авторы определяют этот термин более широко, включая ультрафиолетовую и инфракрасную также части электромагнитного спектра, известные под общим названием оптическое излучение . [ 1 ]
Типичный человеческий глаз реагирует на длины волн от 380 до 750 нанометров . [ 2 ] По частоте это соответствует полосе в районе 400–790 ТГц . Эти границы не четко определены и могут варьироваться в зависимости от человека. [ 3 ] В оптимальных условиях эти пределы человеческого восприятия могут простираться до 310 нм (ультрафиолетовый свет) и 1100 нм (ближний инфракрасный диапазон). [ 4 ] [ 5 ] [ 6 ]
Спектр не содержит всех цветов человека , которые может различать зрительная система . Ненасыщенные цвета, такие как розовый , или фиолетового вариации , такие как пурпурный , например, отсутствуют, поскольку они могут быть получены только из смеси нескольких длин волн. Цвета, содержащие только одну длину волны, также называются чистыми цветами или спектральными цветами . [ 7 ]
Видимые длины волн проходят практически без ослабления через атмосферу Земли через область « оптического окна » электромагнитного спектра. Примером этого явления является то, что чистый воздух рассеивает синий свет больше, чем красный, поэтому полуденное небо кажется синим (за исключением области вокруг Солнца, которая кажется белой, потому что свет рассеивается не так сильно). Оптическое окно также называют «видимым окном», поскольку оно перекрывает спектр видимой реакции человека. Окно ближнего инфракрасного диапазона (NIR) находится вне поля зрения человека, а также окно средневолнового инфракрасного диапазона (MWIR) и окно длинноволнового или дальнего инфракрасного диапазона (LWIR или FIR), хотя другие животные могут их воспринимать.
Спектральные цвета
[ редактировать ] | |||
|---|---|---|---|
| Цвет | Длина волны ( нм ) |
Частота ( ТГц ) |
Фотонная энергия ( эВ ) |
| 380–450 | 670–790 | 2.75–3.26 | |
| 450–485 | 620–670 | 2.56–2.75 | |
| 485–500 | 600–620 | 2.48–2.56 | |
| 500–565 | 530–600 | 2.19–2.48 | |
| 565–590 | 510–530 | 2.10–2.19 | |
| 590–625 | 480–510 | 1.98–2.10 | |
| 625–750 | 400–480 | 1.65–1.98 | |
Цвета, которые можно получить с помощью видимого света узкого диапазона длин волн ( монохроматический свет ), называются чистыми спектральными цветами . Различные цветовые диапазоны, указанные на иллюстрации, являются приблизительными: спектр непрерывен, без четких границ между одним цветом и другим. [ 8 ]
История
[ редактировать ]
В 13 веке Роджер Бэкон предположил, что радуга возникает в результате процесса, аналогичного прохождению света через стекло или кристалл. [ 9 ]
В 17 веке Исаак Ньютон обнаружил, что призмы могут разбирать и собирать белый свет, и описал это явление в своей книге «Оптика» . Он был первым, кто использовал слово спектр ( лат. «видимость» или «видение») в этом смысле в печати в 1671 году при описании своих экспериментов по оптике . Ньютон заметил, что, когда узкий луч солнечного света падает на поверхность стеклянной призмы под углом, часть луча отражается , а часть проходит в стекло и сквозь него, образуя полосы разного цвета. Ньютон предположил, что свет состоит из «корпускул» (частиц) разных цветов, причем свет разных цветов движется с разной скоростью в прозрачном веществе, причем красный свет движется быстрее, чем фиолетовый в стекле. В результате красный свет преломляется ( преломляется ) менее резко, чем фиолетовый, когда он проходит через призму, создавая спектр цветов.
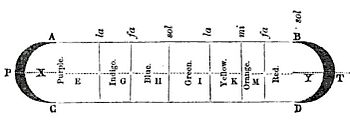
Ньютон первоначально разделил спектр на шесть названных цветов: красный , оранжевый , желтый , зеленый , синий и фиолетовый . Позже он добавил индиго в качестве седьмого цвета, так как считал, что семь — это совершенное число, заимствованное у древнегреческих софистов , о существовании связи между цветами, музыкальными нотами, известными объектами Солнечной системы и днями неделя. [ 10 ] Человеческий глаз относительно нечувствителен к частотам индиго, и некоторые люди с хорошим зрением не могут отличить индиго от синего и фиолетового. По этой причине некоторые более поздние комментаторы, в том числе Айзек Азимов , [ 11 ] предположили, что индиго следует рассматривать не как самостоятельный цвет, а просто как оттенок синего или фиолетового. Факты показывают, что то, что Ньютон имел в виду под «индиго» и «синим», не соответствует современным значениям этих цветных слов. Сравнение наблюдений Ньютона за призматическими цветами с цветным изображением спектра видимого света показывает, что «индиго» соответствует тому, что сегодня называется синим, тогда как его «синий» соответствует голубому цвету . [ 12 ] [ 13 ] [ 14 ]
В 18 веке Иоганн Вольфганг фон Гете написал об оптических спектрах в своей «Теории цвета» . Гете использовал слово спектр ( Spektrum ) для обозначения призрачного оптического остаточного изображения , как это делал Шопенгауэр в «О зрении и цветах» . Гете утверждал, что непрерывный спектр представляет собой сложное явление. В то время как Ньютон сузил луч света, чтобы изолировать явление, Гете заметил, что более широкая апертура дает не спектр, а скорее красновато-желтые и сине-голубые края с белым между ними. Спектр появляется только тогда, когда эти края находятся достаточно близко, чтобы перекрываться.
В начале 19 века понятие видимого спектра стало более определенным, поскольку свет за пределами видимого диапазона был открыт и охарактеризован Уильямом Гершелем ( инфракрасный ) и Иоганном Вильгельмом Риттером ( ультрафиолетовый ), Томасом Янгом , Томасом Иоганном Зеебеком и другими. [ 15 ] Янг был первым, кто измерил длины волн света разных цветов в 1802 году. [ 16 ]
Связь между видимым спектром и цветовым зрением исследовали Томас Янг и Герман фон Гельмгольц в начале 19 века. Их теория цветового зрения правильно предположила, что глаз использует три различных рецептора для восприятия цвета.
Пределы видимого диапазона
[ редактировать ]
Видимый спектр ограничен длинами волн, которые могут достигать сетчатки и вызывать зрительную фототрансдукцию (возбуждать зрительный опсин ). Нечувствительность к УФ-излучению обычно ограничивается его пропусканием через линзу . Нечувствительность к ИК-излучению ограничена функциями спектральной чувствительности зрительных опсинов. Диапазон определяется психометрически функцией светоотдачи , которая учитывает все эти факторы. У человека существует отдельная функция для каждой из двух зрительных систем: одна для фотопического зрения , используемого при дневном свете, опосредованного колбочками , и одна для скотопического зрения , используемого при тусклом свете, опосредованного палочковидными клетками . Каждая из этих функций имеет разные видимые диапазоны. Однако обсуждение видимого диапазона обычно предполагает фотопическое зрение.
Атмосферная передача
[ редактировать ]Видимый диапазон большинства животных эволюционировал, чтобы соответствовать оптическому окну , то есть диапазону света, который может проходить через атмосферу. Озоновый слой поглощает почти весь ультрафиолетовый свет (ниже 315 нм). [ 17 ] Однако это влияет только на космический свет (например, солнечный свет ), а не на земной свет (например, биолюминесценцию ).
Глазная передача
[ редактировать ]Прежде чем достичь сетчатки , свет должен сначала пройти через роговицу и хрусталик . Свет UVB (<315 нм) фильтруется в основном роговицей, а свет UVA (315–400 нм) фильтруется преимущественно хрусталиком. [ 18 ] Хрусталик также желтеет с возрастом, ослабляя пропускание наиболее сильно в синей части спектра. [ 18 ] Это может вызвать ксантопсию , а также небольшое усечение коротковолновой (синей) границы видимого спектра. У субъектов с афакией отсутствует хрусталик, поэтому свет UVA может достигать сетчатки и возбуждать зрительные опсины; это расширяет видимый диапазон и может также привести к цианопсии .
Поглощение опсина
[ редактировать ]Каждый опсин имеет функцию спектральной чувствительности , которая определяет, насколько вероятно, что он поглотит фотон каждой длины волны. Функция светоотдачи представляет собой приблизительно суперпозицию вносящих вклад зрительных опсинов . Таким образом, изменение положения отдельных функций спектральной чувствительности опсина влияет на функцию светоотдачи и видимый диапазон. Например, длинноволновой (красный) предел меняется пропорционально положению L-опсина. Положения определяются пиковой длиной волны (длиной волны наибольшей чувствительности), поэтому, когда пиковая длина волны L-опсина смещается на 10 нм, длинноволновая граница видимого спектра также смещается на 10 нм. Большие отклонения пиковой длины волны L-опсина приводят к форме цветовой слепоты , называемой протаномалией , а отсутствие L-опсина ( протанопия ) укорачивает видимый спектр примерно на 30 нм в длинноволновом пределе. Формы дальтонизма, затрагивающие М-опсин и S-опсин, существенно не влияют на функцию светоотдачи и пределы видимого спектра.
Различные определения
[ редактировать ]Независимо от реальных физических и биологических различий, определение пределов не является стандартным и будет меняться в зависимости от отрасли. Например, некоторые отрасли могут быть обеспокоены практическими ограничениями, поэтому консервативно сообщают о длине волны 420–680 нм. [ 19 ] [ 20 ] в то время как другие могут быть заинтересованы в психометрии и для достижения самого широкого спектра обычно сообщают о 380–750 или даже 380–800 нм. [ 21 ] [ 22 ] Функция светоотдачи в ближнем ИК-диапазоне не имеет жесткого ограничения, а имеет экспоненциальное затухание, так что значение функции (или чувствительность зрения) на длине волны 1050 нм составляет около 10 9 раз слабее, чем при 700 нм; Поэтому для восприятия света с длиной волны 1050 нм требуется гораздо более высокая интенсивность, чем для света с длиной волны 700 нм. [ 23 ]
Видение за пределами видимого спектра
[ редактировать ]В идеальных лабораторных условиях испытуемые могут воспринимать инфракрасный свет длиной не менее 1064 нм. [ 23 ] Хотя БИК-свет с длиной волны 1050 нм может вызывать красный цвет, что предполагает прямое поглощение L-опсином, есть также сообщения о том, что импульсные БИК-лазеры могут вызывать зеленый цвет, что предполагает, что двухфотонное поглощение может обеспечить расширенную чувствительность БИК. [ 23 ]
Точно так же молодые люди могут воспринимать ультрафиолетовые волны длиной примерно до 310–313 нм. [ 24 ] [ 25 ] [ 26 ] но обнаружение света с длиной волны ниже 380 нм может быть связано с флуоресценцией среды глаза, а не с прямым поглощением УФ-света опсинами. Поскольку свет UVA поглощается средами глаза (хрусталиком и роговицей), он может флуоресцировать и выделяться с более низкой энергией (более длинной волной), которая затем может быть поглощена опсинами. Например, когда линза поглощает свет с длиной волны 350 нм, спектр излучения флуоресценции сосредоточен на длине волны 440 нм. [ 27 ]
Невизуальное обнаружение света
[ редактировать ]Помимо фотопической и скотопической систем, у людей есть и другие системы обнаружения света, которые не участвуют в работе первичной зрительной системы . Например, меланопсин имеет диапазон поглощения 420–540 нм и регулирует циркадный ритм и другие рефлекторные процессы. [ 28 ] Поскольку меланопсиновая система не формирует изображения, она строго не считается зрением и не вносит вклад в видимый диапазон.
В нечеловеческом
[ редактировать ]Видимый спектр определяется как видимый для человека, но различия между видами велики. не только могут Опсины колбочек быть спектрально смещены для изменения видимого диапазона, но и позвоночные с 4 колбочками (тетрахроматические) или 2 колбочками (дихроматические) по сравнению с тремя колбочками у человека (трихроматические), также будут иметь тенденцию иметь более широкий или более узкий видимый спектр, чем люди. соответственно.
Позвоночные животные, как правило, имеют 1-4 различных класса опсинов: [ 17 ]
- длинноволновая чувствительность (LWS) с пиковой чувствительностью 500–570 нм,
- средневолновая чувствительность (MWS) с пиковой чувствительностью 480–520 нм,
- Коротковолновая чувствительность (SWS) с пиковой чувствительностью 415–470 нм и
- чувствительный к фиолетовому/ультрафиолетовому излучению (VS/UVS) с максимальной чувствительностью в диапазоне 355–435 нм.
Поведенческое тестирование зрительных систем животных сложно, поэтому видимый диапазон животных обычно оценивается путем сравнения пиковых длин волн опсинов с пиковыми длинами волн опсинов у типичных людей (S-опсин при 420 нм и L-опсин при 560 нм).
Млекопитающие
[ редактировать ]У большинства млекопитающих сохранились только два класса опсинов (LWS и VS), что, вероятно, связано с ночным узким местом . Однако с тех пор приматы Старого Света (включая людей) развили две версии класса LWS, чтобы восстановить трихроматию. [ 17 ] В отличие от большинства млекопитающих, опсины UVS грызунов имеют более короткие волны. Помимо отсутствия УФ-фильтров в хрусталике мышей есть опсин UVS, способный обнаруживать свет на длине волны до 340 нм. Хотя попадание ультрафиолетового света на сетчатку может привести к повреждению сетчатки, короткая продолжительность жизни мышей по сравнению с другими млекопитающими может свести к минимуму этот недостаток по сравнению с преимуществами ультрафиолетового зрения. [ 29 ] У собак есть два конусных опсина с длиной волны 429 и 555 нм, поэтому они видят почти весь видимый спектр человека, несмотря на то, что они дихроматические. [ 30 ] У лошадей есть два конусных опсина с длиной волны 428 и 539 нм, что обеспечивает немного более усеченное красное зрение. [ 31 ]
Птицы
[ редактировать ]Большинство других позвоночных (птицы, ящерицы, рыбы и т. д.) сохранили свою тетрахроматию , включая опсины UVS, которые простираются дальше в ультрафиолет, чем опсин VS человека. [ 17 ] Чувствительность опсинов UVS птиц сильно варьируется от 355–425 нм, а опсинов LWS – от 560–570 нм. [ 32 ] Это означает, что некоторые птицы имеют видимый спектр наравне с людьми, а другие птицы имеют значительно повышенную чувствительность к ультрафиолетовому излучению. Иногда сообщается, что опсин LWS птиц имеет пиковую длину волны выше 600 нм, но это эффективная пиковая длина волны, которая включает в себя фильтр капель птичьего жира . [ 32 ] Пиковая длина волны опсина LWS сама по себе является лучшим предиктором длинноволнового предела. Возможная польза от ультрафиолетового зрения птиц заключается в том, что на их оперении появляются зависящие от пола отметины , которые видны только в ультрафиолетовом диапазоне. [ 33 ] [ 34 ]
Рыба
[ редактировать ]Костистые рыбы (костистые рыбы) обычно четырехцветные. Чувствительность опсинов UVS рыб варьируется в пределах 347–383 нм, а опсинов LWS – в диапазоне 500–570 нм. [ 35 ] Однако некоторые рыбы, использующие альтернативные хромофоры, могут увеличить чувствительность опсина LWS до 625 нм. [ 35 ] Распространено мнение, что обыкновенная золотая рыбка — единственное животное, способное видеть как инфракрасный, так и ультрафиолетовый свет. [ 36 ] неверно, поскольку золотая рыбка не видит инфракрасного света. [ 37 ]
Беспозвоночные
[ редактировать ]Зрительные системы беспозвоночных сильно отличаются от позвоночных, поэтому прямые сравнения затруднительны. Однако чувствительность к УФ-излучению зарегистрирована у большинства видов насекомых. [ 38 ] Пчелы и многие другие насекомые способны улавливать ультрафиолетовый свет, который помогает им находить нектар в цветах. Виды растений, которые зависят от опыления насекомыми, могут быть обязаны репродуктивным успехом своему внешнему виду в ультрафиолетовом свете, а не тому, насколько красочными они кажутся людям. Длинноволновой предел пчел составляет около 590 нм. [ 39 ] Креветки-богомолы содержат до 14 опсинов, что обеспечивает видимый диапазон от менее 300 нм до более 700 нм. [ 17 ]
Тепловое зрение
[ редактировать ]Некоторые змеи могут «видеть» [ 40 ] лучистое тепло на длинах волн от 5 до 30 мкм с такой степенью точности, что слепая гремучая змея может нацеливаться на уязвимые части тела жертвы, на которую она нападает, [ 41 ] а другие змеи с этим органом могут обнаруживать теплые тела на расстоянии метра. [ 42 ] Его также можно использовать для терморегуляции и обнаружения хищников . [ 43 ] [ 44 ]
Спектроскопия
[ редактировать ]
Спектроскопия — это исследование объектов на основе спектра цветов, которые они излучают, поглощают или отражают. Спектроскопия видимого света — важный инструмент в астрономии (как и спектроскопия на других длинах волн), где ученые используют его для анализа свойств удаленных объектов. Химические элементы и небольшие молекулы можно обнаружить в астрономических объектах, наблюдая линии излучения и линии поглощения . Например, гелий был впервые обнаружен путем анализа спектра Солнца . Сдвиг частоты спектральных линий используется для измерения доплеровского смещения ( красного или синего смещения ) удаленных объектов, чтобы определить их скорости по направлению к наблюдателю или от него. Астрономическая спектроскопия высокой дисперсией использует дифракционные решетки с для наблюдения спектров с очень высоким спектральным разрешением.
См. также
[ редактировать ]- Видимый свет высокой энергии
- Визуальные явления космических лучей
- Электромагнитное поглощение водой
- Двухфотонное поглощение - метод видения за пределами видимого спектра.
Ссылки
[ редактировать ]- ^ Педротти, Фрэнк Л.; Педротти, Лено М.; Педротти, Лено С. (21 декабря 2017 г.). Введение в оптику . Издательство Кембриджского университета. стр. 100-1 7–8. ISBN 9781108428262 .
- ^ Старр, Сеси (2005). Биология: концепции и приложения . Томсон Брукс/Коул. п. 94 . ISBN 978-0-534-46226-0 .
- ^ «Видимый спектр» . Британика. 27 мая 2024 г.
- ^ Д. Х. Слайни (февраль 2016 г.). «Что такое свет? Видимый спектр и за его пределами» . Глаз . 30 (2): 222–229. дои : 10.1038/eye.2015.252 . ISSN 1476-5454 . ПМЦ 4763133 . ПМИД 26768917 .
- ^ У. К. Ливингстон (2001). Цвет и свет в природе (2-е изд.). Кембридж, Великобритания: Издательство Кембриджского университета. ISBN 0-521-77284-2 .
- ^ Гражина Пальчевска; и др. (декабрь 2014 г.). «Инфракрасное зрение человека обусловлено двухфотонной изомеризацией хромофора» . Труды Национальной академии наук . 111 (50): Е5445–Е5454. Бибкод : 2014PNAS..111E5445P . дои : 10.1073/pnas.1410162111 . ПМЦ 4273384 . ПМИД 25453064 .
- ^ Нейв, Р. «Спектральные цвета» . Гиперфизика . Проверено 11 мая 2022 г.
- ^ Бруно, Томас Дж. и Своронос, Париж DN (2005). Справочник CRC по диаграммам фундаментальной спектроскопической корреляции. ЦРК Пресс. ISBN 9781420037685
- ^ Коффи, Питер (1912). Наука логики: исследование принципов точного мышления . Лонгманс. п. 185 .
Призма Роджера Бэкона.
- ^ Исаков, Стюарт (16 января 2009 г.). Темперамент: как музыка стала полем битвы великих умов западной цивилизации . Издательская группа Кнопфа Doubleday. стр. 12–13. ISBN 978-0-307-56051-3 . Проверено 18 марта 2014 г.
- ^ Азимов, Исаак (1975). Глаза на Вселенную: история телескопа . Бостон: Хоутон Миффлин. п. 59 . ISBN 978-0-395-20716-1 .
- ^ Эванс, Ральф М. (1974). Восприятие цвета (нулевое изд.). Нью-Йорк: Wiley-Interscience. ISBN 978-0-471-24785-2 .
- ^ Макларен, К. (март 2007 г.). «Индиго Ньютона». Исследование и применение цвета . 10 (4): 225–229. дои : 10.1002/col.5080100411 .
- ^ Уолдман, Гэри (2002). Введение в свет: физика света, зрения и цвета (изд. Дувра). Минеола: Dover Publications. п. 193. ИСБН 978-0-486-42118-6 .
- ^ Мэри Джо Най, изд. (2003). Кембриджская история науки: современные физико-математические науки . Том. 5. Издательство Кембриджского университета. п. 278. ИСБН 978-0-521-57199-9 .
- ^ Джон С.Д. Брэнд (1995). Линии света: источники дисперсионной спектроскопии, 1800–1930 гг . ЦРК Пресс. стр. 30–32. ISBN 978-2-88449-163-1 .
- ^ Jump up to: а б с д и Хант, DM; Уилки, ЮВ; Боумейкер, Дж. К.; Пупаласундарам, С. (октябрь 2001 г.). «Зрение в ультрафиолете». Клеточные и молекулярные науки о жизни . 58 (11): 1583–1598. дои : 10.1007/PL00000798 . ПМИД 11706986 . S2CID 22938704 .
Излучение ниже 320 нм [ультрафиолетовое (УФ)А] в значительной степени экранируется озоновым слоем в верхних слоях атмосферы Земли и поэтому недоступно для зрительной системы.
- ^ Jump up to: а б Беттнер, Эдвард А.; Уолтер, Дж. Реймер (декабрь 1962 г.). «Передача глазных медиа». Исследовательская офтальмология и визуальные науки . 1 : 776-783.
- ^ Лауфер, Габриэль (1996). «Геометрическая оптика» . Введение в оптику и лазеры в технике . п. 11. Бибкод : 1996iole.book.....L . дои : 10.1017/CBO9781139174190.004 . ISBN 978-0-521-45233-5 . Проверено 20 октября 2013 г.
- ^ Брэдт, Хейл (2004). Астрономические методы: физический подход к астрономическим наблюдениям . Издательство Кембриджского университета. п. 26. ISBN 978-0-521-53551-9 . Проверено 20 октября 2013 г.
- ^ Оганесян, Лена; Стритер, Энтони (2001). Справочник по фармацевтическому анализу . ЦРК Пресс. п. 187. ИСБН 978-0-8247-4194-5 . Проверено 20 октября 2013 г.
- ^ Ахлувалия, В.К.; Гоял, Мадхури (2000). Учебник органической химии . Розовый. п. 110. ИСБН 978-81-7319-159-6 . Проверено 20 октября 2013 г.
- ^ Jump up to: а б с Слайни, Дэвид Х.; Вангеманн, Роберт Т.; Фрэнкс, Джеймс К.; Вольбаршт, Майрон Л. (1976). «Зрительная чувствительность глаза к инфракрасному лазерному излучению». Журнал Оптического общества Америки . 66 (4): 339–341. Бибкод : 1976JOSA...66..339S . дои : 10.1364/JOSA.66.000339 . ПМИД 1262982 .
Была измерена фовеальная чувствительность к нескольким длинам волн лазера ближнего инфракрасного диапазона. Было обнаружено, что глаз может реагировать на излучение с длиной волны по крайней мере до 1064 нм. Непрерывный лазерный источник с длиной волны 1064 нм выглядел красным, а импульсный лазерный источник с длиной волны 1060 нм — зеленым, что предполагает наличие генерации второй гармоники в сетчатке.
- ^ Линч, Дэвид К.; Ливингстон, Уильям Чарльз (2001). Цвет и свет в природе (2-е изд.). Кембридж: Издательство Кембриджского университета. п. 231. ИСБН 978-0-521-77504-5 . Архивировано из оригинала 8 октября 2022 года . Проверено 12 октября 2013 г.
Пределы общего диапазона чувствительности глаза простираются от 310 до 1050 нанометров.
- ^ Даш, Мадхаб Чандра; Даш, Сатья Пракаш (2009). Основы экологии 3Е . Тата МакГроу-Хилл Образование. п. 213. ИСБН 978-1-259-08109-5 . Архивировано из оригинала 8 октября 2022 года . Проверено 18 октября 2013 г.
В норме человеческий глаз реагирует на световые лучи длиной от 390 до 760 нм. В искусственных условиях этот диапазон можно расширить до диапазона от 310 до 1050 нм.
- ^ Саидман, Жан (15 мая 1933 г.). «О видимости ультрафиолета до длины волны 3130» . Доклады Академии наук (на французском языке). 196 : 1537–9. Архивировано из оригинала 24 октября 2013 года . Проверено 21 октября 2013 г.
- ^ Курзель, Ричард Б.; Вольбаршт, Майрон Л.; Яманаси, Билл С. (1977). «Влияние ультрафиолетового излучения на человеческий глаз». Фотохимические и фотобиологические обзоры . стр. 133–167. дои : 10.1007/978-1-4684-2577-2_3 . ISBN 978-1-4684-2579-6 .
- ^ Энези Дж., Ревелл В., Браун Т., Винн Дж., Шланген Л., Лукас Р. (август 2011 г.). «Функция «меланопической» спектральной эффективности предсказывает чувствительность фоторецепторов меланопсина к полихроматическому свету» . Журнал биологических ритмов . 26 (4): 314–323. дои : 10.1177/0748730411409719 . ПМИД 21775290 . S2CID 22369861 .
- ^ Гурас, Питер; Экестен, Бьорн (декабрь 2004 г.). «Почему у мышей ультрафиолетовое зрение?». Экспериментальное исследование глаз . 79 (6): 887–892. дои : 10.1016/j.exer.2004.06.031 . ПМИД 15642326 .
- ^ Нейтц, Джей; Гейст, Тимоти; Джейкобс, Джеральд Х. (август 1989 г.). «Цветовое зрение у собаки». Визуальная нейронаука . 3 (2): 119–125. дои : 10.1017/S0952523800004430 . ПМИД 2487095 . S2CID 23509491 .
- ^ Кэрролл, Джозеф; Мерфи, Кристофер Дж.; Нейтц, Морин; Вер Хув, Джеймс Н.; Нейтц, Джей (3 октября 2001 г.). «Фотопигментная основа дихроматического цветового зрения у лошадей» . Журнал видения . 1 (2): 80–87. дои : 10.1167/1.2.2 . ПМИД 12678603 .
- ^ Jump up to: а б Харт, Натан С.; Хант, Дэвид М. (январь 2007 г.). «Птичьи визуальные пигменты: характеристики, спектральная настройка и эволюция». Американский натуралист . 169 (С1): С7–С26. дои : 10.1086/510141 . ПМИД 19426092 . S2CID 25779190 .
- ^ Катхилл, Иннес С. (1997). «Ультрафиолетовое зрение у птиц». В Питере Дж. Б. Слейтере (ред.). Достижения в изучении поведения . Том. 29. Оксфорд, Англия: Академик Пресс. п. 161. ИСБН 978-0-12-004529-7 .
- ^ Джеймисон, Барри GM (2007). Репродуктивная биология и филогения птиц . Шарлоттсвилл, Вирджиния: Университет Вирджинии. п. 128. ИСБН 978-1-57808-386-2 .
- ^ Jump up to: а б Карлтон, Карен Л.; Эскобар-Камачо, Даниэль; Стиб, Сара М.; Кортези, Фабио; Маршалл, Н. Джастин (15 апреля 2020 г.). «Видеть радугу: механизмы, лежащие в основе спектральной чувствительности костистых рыб» . Журнал экспериментальной биологии . 223 (8). дои : 10.1242/jeb.193334 . ПМЦ 7188444 . ПМИД 32327561 .
- ^ «Правда или ложь? «Обыкновенная золотая рыбка — единственное животное, которое может видеть как инфракрасный, так и ультрафиолетовый свет » . Скептический . 2013. Архивировано из оригинала 24 декабря 2013 года . Проверено 28 сентября 2013 г.
- ^ Ноймайер, Криста (2012). «Глава 2: Цветовое зрение золотых рыбок и других позвоночных». У Лазаревой Ольги; Симидзу, Тору; Вассерман, Эдвард (ред.). Как животные видят мир: сравнительное поведение, биология и эволюция зрения . Оксфордская стипендия онлайн. ISBN 978-0-19-533465-4 .
- ^ Бриско, Адриана Д.; Читтка, Ларс (январь 2001 г.). "T<SCP>HE</SCP>E<SCP>ВОЛЮЦИЯ</SCP>C<SCP>ЦВЕТА</SCP>V<SCP>ISION В</SCP>I<SCP>NSECTS</SCP>". Ежегодный обзор энтомологии . 46 (1): 471–510. дои : 10.1146/annurev.ento.46.1.471 . ПМИД 11112177 .
- ^ Скорупский, Питер; Читтка, Ларс (10 августа 2010 г.). «Спектральная чувствительность фоторецепторов у шмелей Bombus impatiens (Hymenoptera: Apidae)» . ПЛОС ОДИН . 5 (8): e12049. Бибкод : 2010PLoSO...512049S . дои : 10.1371/journal.pone.0012049 . ПМК 2919406 . ПМИД 20711523 .
- ^ Ньюман, Э.А.; Хартлайн, штат Пенсильвания (1981). «Интеграция зрительной и инфракрасной информации в бимодальных нейронах зрительного покрова гремучей змеи» . Наука . 213 (4509): 789–91. Бибкод : 1981Sci...213..789N . дои : 10.1126/science.7256281 . ПМК 2693128 . ПМИД 7256281 .
- ^ Кардонг, КВ; Макесси, СП (1991). «Ударное поведение врожденно слепой гремучей змеи». Журнал герпетологии . 25 (2): 208–211. дои : 10.2307/1564650 . JSTOR 1564650 .
- ^ Фанг, Джанет (14 марта 2010 г.). «Раскрыто инфракрасное обнаружение змей» . Новости природы . дои : 10.1038/news.2010.122 .
- ^ Крохмаль, Аарон Р.; Джордж С. Баккен; Трэвис Дж. ЛаДюк (15 ноября 2004 г.). «Жара на кухне эволюции: эволюционные взгляды на функции и происхождение лицевой ямки гадюк (Viperidae: Crotalinae)» . Журнал экспериментальной биологии . 207 (Часть 24): 4231–4238. дои : 10.1242/jeb.01278 . ПМИД 15531644 .
- ^ Грин HW. (1992). «Экологический и поведенческий контекст эволюции гадюк», в книге Кэмпбелл Дж.А., Броди Э.Д. младший. Биология гадюк . Техас: Сельва. ISBN 0-9630537-0-1 .
Цветовые темы |
|---|